The last decade has seen a veritable explosion in the manner in which fluorescence microscopy has been applied to studies of the immune system. Since students will be exposed to many of these techniques in the current primary literature, we present here a summary of the common variations of immunofluorescence. See also Advances Box 20-1, which introduces students to dynamic imaging techniques that visualize cells in real time during an ongoing immune response and offers hints on how to view the resulting movies with an educated and critical eye.
Fluorescence Can Be Used to Visualize Cells and Molecules
The phenomenon of fluorescence results from the property of some molecules (fluorescent dyes) to absorb light at one wavelength and emit it at a longer wavelength. If the emitted light has a wavelength in the visible region of the spectrum, the fluorescent dye can be used to detect any molecules bound by that dye.
Some fluorescence imaging experiments take advantage of the fact that particular dyes specifically bind to particular macromolecules. For example, the blue dye 4′,6-diamidino-2-phenylindole (DAPI) specifically stains DNA. Other protocols utilize the affinities of easily obtained proteins (which can be readily conjugated with fluorescent dyes) to bind to biologically important molecules. For example, the protein phalloidin, which specifically binds filamentous actin, can easily be conjugated to fluorescent probes. Similarly, the soluble protein annexin A5 binds to phosphatidylserine, which is exposed on the outer surface of cells undergoing apoptosis; fluorescently labeled annexin A5 is easy to obtain and use as a measure of apoptosis. In immunofluorescence measurements, antibodies or streptavidin can be artificially conjugated to a host of dyes. These various approaches to fluorescence labeling can be combined with one another to provide spectacular images of cellular and subcellular structures (Figure 20-14a).
FIGURE 20-14 Fluorescently labeled cells and the passage of light through a fluorescence microscope. (a) Fluorescence microscopic image of a human dendritic cell infected with engineered Listeria monocytogenes. The green fluorescence is generated by Alexa Fluor 488–conjugated phalloidin (which binds actin filaments). Red fluorescence derives from red fluorescent protein expressed by the bacterium. DNA is stained with DAPI (blue). (b) Light from a source passes through a barrier filter that only allows passage of blue light of particular wavelengths. The light is then directed onto the sample by a dichroic mirror that reflects light of short wavelengths (below approximately 510 nm) but allows passage of higher wavelengths. When the blue light interacts with the sample, any fluorescent molecules excited by it emit fluorescence that then passes through the dichroic mirror, through a second barrier filter, and then is transmitted to the eyepiece.
Such images can be visualized by fluorescence microscopy, which uses short-wavelength light to excite the fluorescent dyes. A series of filters and mirrors that can be adjusted by the investigator can then be employed to select which wavelengths of light will reach the eyepiece and therefore which molecules will be visualized (see Figure 20-14b). Modern instruments use combinations of several filters and mirrors that allow the investigator to detect light emitted at multiple different fluorescence wavelengths. The different colored images generated from the various dyes can then be overlaid by the instrument’s software to provide a single representation in which the locations of the antibody-bound molecules can be compared.
In addition to artificially synthesized dyes, some naturally occurring proteins such as GFP and RFP (green and red fluorescent protein, respectively) contain fluorescent chromophores. Figure 20-15 shows a striking image of three neonatal mice engineered to express GFP under the control of an actin promoter, along with their non-GFP littermates. In some experiments, investigators will place the fluorescent protein genes under the control of particular promoters or make fusion proteins that contain both the native protein and GFP sequences. These adaptations allow researchers to use fluorescence imaging to determine where and when proteins under the control of those same promoters are expressed.
FIGURE 20-15 Fluorescence labeling of whole animals with green fluorescent protein. Three neonatal transgenic mice express GFP under the control of an actin promoter.
Confocal Fluorescence Microscopy Provides Three-Dimensional Images of Extraordinary Clarity
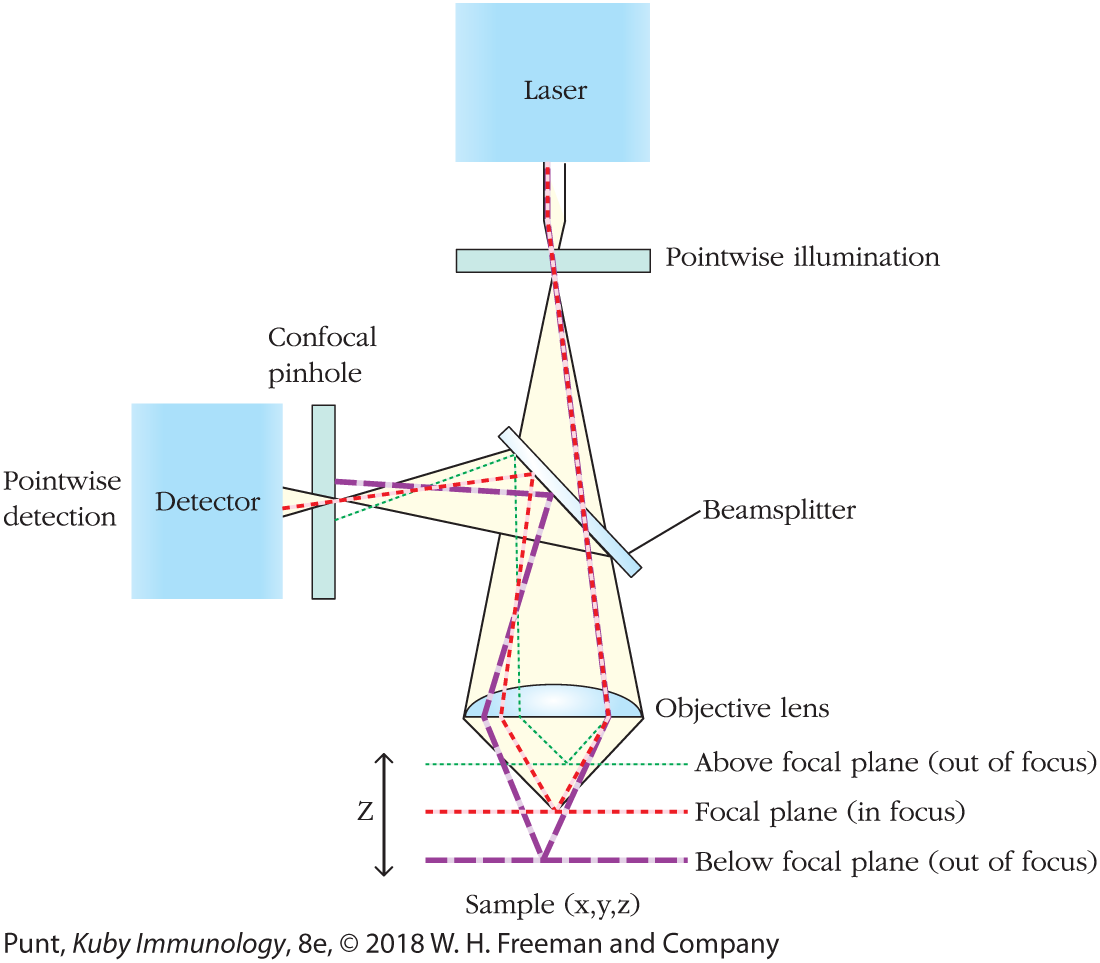
One of the limiting factors in obtaining clear images by fluorescence microscopy is that fluorescent molecules lying above and below the focal plane can contribute to the light that reaches the objective, leading to a blurred image. In confocal microscopy, that artifact is eliminated by using an objective lens that focuses the light from the desired focal plane directly onto a pinhole aperture in front of the detector (Figure 20-16). Light emitted from molecules located at other levels within the sample is stopped at the perimeter of the pinhole, and remarkably clear images of a single plane within the sample can thus be generated. In laser scanning confocal microscopy, investigators use lasers to provide the exciting light and computing power to move the focal plane in all three dimensions, thus enabling them to scan an x, y plane at different depths of focus, and reconstitute powerful three-dimensional images.
FIGURE 20-16 The principle of confocal microscopy. The sample is illuminated by a laser beam that excites fluorescence from dyes in several different focal planes, represented here by the green, red, and purple lines. However, passage of light through the confocal pinhole (shown on the left-hand side of the image) filters out light emitted from all but a single focal plane, resulting in an extraordinarily clear image. Computer control of the exact plane from which light can be received through the pinhole allows the development of images from a number of focal planes and the creation of a composite three-dimensional representation.
Multiphoton Fluorescence Microscopy Is a Variation of Confocal Microscopy
Two-photon and multiphoton microscopy are variations on confocal microscopy that offer even greater resolution in the development of three-dimensional images. In standard confocal microscopy, excitation of the fluorescent probes (dyes) occurs along the whole path of the laser beam through the tissue (Figure 20-17a). This means that, although the emission beams are derived only from a single level within the sample, fluorescent probes are being excited throughout many levels of the tissue. Since fluorescent probes will eventually photobleach (i.e., cease to emit light) after extensive excitation, this limits the useful life of the sample. It also means that some additional light is emitted from the sample that must be filtered out of the final image.
FIGURE 20-17 Fluorescence excitation by one-photon versus two-photon laser excitation. (a) On the left-hand side, we visualize excitation by a short-wavelength, high-energy laser that excites fluorescence from all molecules in its path. (b) On the right-hand side, we see the fluorescence that results when molecules are excited with near simultaneity by two beams of longer wavelength, lower energy laser light. Since excitation can occur only in the region where the two beams intersect, it is limited to a single focal plane.
In multiphoton fluorescence microscopy, long-wavelength lasers are used that emit in the infrared region of the spectrum. The laser beams are relatively low energy, and so more than one photon must impinge on a fluorescent molecule in order to provide sufficient energy to excite the electrons. The low energy of these infrared lasers minimizes the extent of photobleaching and enhances the useful lifetime of the sample, since those parts of the sample that interact with only a single laser beam are not typically damaged. Furthermore, the fact that at least two beams of light are required to bring about fluorescence excitation ensures that excitation occurs only within the plane of intersection of the laser beams (see Figure 20-17b). By moving the focal point of excitation within the x and y planes, information about a full optical section can be generated, and that whole process can then be repeated on additional z levels, thus giving rise to a three-dimensional image. In Chapter 14, you have seen some of the powerful images developed with this technique.
Intravital Imaging Allows Observation of Immune Responses in Vivo
Intravital imaging takes advantage of the ability to maintain both the blood and the lymphatic circulations within lymph nodes, sections of intestine, or other organs, after they have been gently lifted out of an anesthetized donor and onto a warmed perfused microscope stage. Using a multiphoton microscope, three-dimensional images of fluorescently labeled cells and structures can be generated and information gleaned about the behavior of immune cells and molecules essentially in vivo. Advances Box 20-1 describes this approach in more detail and Chapter 14 describes significant advances that have been made using this, and related technologies.
Visualization and Analysis of DNA Sequences in Intact Chromatin
Three-dimensional fluorescence in situ hybridization (3-D FISH) is a relatively new technique in which fluorescent RNA probes labeled with several different colors are generated that hybridize with known DNA sequences. For example, probes have been developed that hybridize with VH sequences known to be situated farthest from the DH regions (distal VH sequences), or with VH sequences closest to the DH region (proximal VH sequences). Investigators have used combinations of such probes to study the relative arrangements of BCR and TCR genes in cell nuclei at defined stages of lymphoid development. Sophisticated analytical tools then use the distances measured between the individual probes to develop topological maps of the chromosomes. Figure 20-18 is a striking image that allows us to see with our own eyes how the distance between D-JH distal and D-JH proximal VH genes is altered in pro-B cells as the Ig-encoding chromosome undergoes contraction during V-D recombination.
FIGURE 20-18 Three-dimensional fluorescence in situ hybridization (3-D FISH). 3-D FISH is used to show the distances between proximal (VH7183—red) and distal (VHJ558—green) genes in EL4 T cells (left) relative to those in pro-B cells (right). On the left-hand side, we see that, in T cells, the proximal and distal gene families (red and green, respectively) are at a distance where they can be readily discriminated from one another. In contrast, in pro-B cells, where the Ig chromosome is undergoing contraction, the two gene families are closer together and the two colors can barely be resolved, if at all.