Thomas W. Myers
This chapter focuses largely on the global postural assessment of compensation patterns in the neuromyofascial web, using the map of ‘myofascial meridians’ known as the Anatomy Trains
®
(Myers 2014).
Chapter 4
has additional global, as well as a number of local, palpation and assessment methods that complement the content of this chapter.
Fascial versus neurological patterns
Since this book is about fascial dysfunction, we should first address the question of the degree to which postural patterns are neuromotor patterns versus actual patterning within the fascial tissues. The answer depends primarily on how long they have been established and whether they are sequelae from trauma or simply ingrained habits.
Every movement begins as an experiment, a novel expression of neuromotor firing. Any parent who has been with their 6-month-old when he or she first turns over knows that the child is just as surprised at this accomplishment as the parent.
To our knowledge there is no specific program for ‘turning over’ in the human brain. There is, however, a very strong program for ‘keeping track of Mom and other moving things’. As the eyes track Mom, the reflexive rotation and extension of the neck initiates the spinal rotation and extension, and the rest of the rolling over movement. When the strength of the muscles and bones and the proportion of the head to torso allow it (remember we are still in the period of
ex utero
gestation and initial ossification), the child does indeed roll over. It may take a few weeks before they can organize turning back the other way by themselves, but in the meantime they are delighted to have you help them return, so that they can turn from back to belly again and again and again.
This repetition of the movement presumably carves some sort of ‘engram’ or trace in the brain, so that the motion is easier to repeat (Berthoz 2000). Repeated enough, the sequence of firing becomes fixed as a habit – easily called up, but less subject to change.
If you were to lie on the floor now and roll from back to front, first to the right and then to the left, you would find that you have a preference, that one side is easier and more ‘natural’ than the other. Before you put down the book to try this experiment for yourself, which you are encouraged to do, keep in mind the following two elements:
1.
Initiate the movement from your eyes, imagining you are tracking something to that side and up, letting your body follow the motion.
2.
Be aware of performing it the same way to both the right and left side – you will see you cannot. Subtle or glaring differences in the two sides abound.
Based on hundreds of iterations of this test with many people, this one-sided preference is not related to your dominant handedness or a dominant eye. It is more likely an expression of which way you were commonly laid in the crib, or the accidental side your mother or the family pet passed you to create the initial stimulus – but in the author’s experience, everyone has a predilection, a preferred side to which to roll. (
Editor’s note
: Consider the information on the Zink common compensatory fascial pattern, as outlined in
Ch. 4
.)
Habits, once firmly established, require a posture to maintain them (
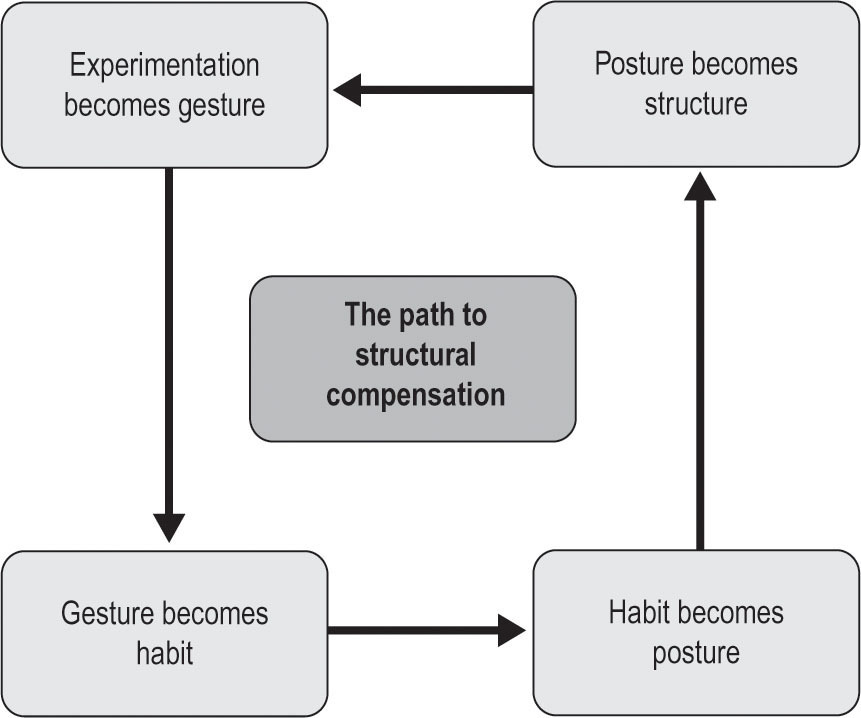
Fig. 3.1
). The preference just outlined will be written visibly into your posture, as well as your movement choices, i.e. which shoulder to look over when you back up your car. Such postural ‘sets’ are maintained in the cerebellum and brainstem, and are thus that much less amenable to ‘voluntary’ control (even though the muscles that these patterns use are all so-called voluntary muscles).
Figure 3.1
The Path to Structural Compensation. Every movement begins as an experiment. An experiment repeated becomes a familiar gesture. Repeat a gesture and it becomes ingrained as a habit. Each habit requires a posture. All of these are neural patterns. Hold a posture for long enough and the fascia takes the pattern. At that point, every new experiment arises from that fascial pattern. In reversing the process, both the fascial patterning and the deep postural habit need to be addressed.
To test this, abduct your arm to horizontal or nearly so and place your forearm on a dresser (or someone else’s shoulder, if you have someone available;
Fig. 3.2
). Press down with your forearm – some combination of the pectorals, latissimus dorsi, teres major, and coracobrachialis will be the active prime movers in this adduction, while the antagonist deltoid will relax. Reach across with your opposite hand to feel (or have your partner feel) the lateral abdominal obliques on the side you are pressing down. Without doubt, you will feel the lateral part of external and internal oblique contract.
These muscles are, of course, acting to stabilize the rib cage on the pelvis. Here is the experiment: try pressing your arm down
without
contracting those muscles, and keep your other hand or your partner’s hand there, to see if you have in fact quelled the tendency to contract these muscles.
If you are like most people – and we have done this experiment with many professional groups in different cultures – the lateral abdominal muscles will pop on automatically when the arm is pressed, and inhibiting this reaction is very difficult. If you are in fact successful in this inhibition, your lower ribs will pull up to the side of the pressing arm (a very ‘silly-looking’ movement); if the insertion cannot go to the origin, the origin will move toward the insertion.
The inability of most people to ‘turn off’ this reaction speaks to the depth of postural patterns in the nervous system. Once established, such patterns as the ‘trunk stabilization program’ we just tested become the unconscious substrate of conscious movement. Our ‘sitting program’ underlies our ‘driving program’ that underlies our ‘this car program’ that underlies the consciously controlled move to reach for the turn signal as we approach the intersection.
Most of the important patterns we wish to change in order to improve our patients’ biomechanics rest in these deeper regions of postural habitus – the postural set that underlies the habit that underlies this particular gesture that is now painful or limited, for whatever reason. While we need to treat the proximate cause to relieve pain or restore function, we often find ourselves entreating our clients to change the habits (perpetuating factors) that led to the failure of the proximate cause, or we will be seeing them soon again for the same symptom.
Figure 3.2
A simple test for understanding levels of neuromuscular programming involves pressing down against resistance with your arm while attempting to inhibit the side belly wall. If you succeed – which requires a trick of the mind – the result is an unusual and not very ‘functional’ exercise.
These patterns are often body-wide, and therefore somewhat confounding. Supraspinatus tendon inflammation above the shoulder joint is caused by improper support from a posteriorly tilted rib cage on top of an anteriorly shifted pelvis and so on, down to shortened hamstrings and a chronically dorsiflexed ankle.
Hold these patterns long enough, and the areas under constant extra load will enjoy more fibroblast activity, which creates more collagenous ‘strapping’ – within or between muscles – to meet the specific strain caused by the repetitive posture (Kjaer et al. 2009).
At this point, we move from a purely neurological pattern to a fascial restriction. From this point on any new experiments, any novel movement the client is learning, will work around that limitation in structure, as opposed to – or more properly, in conjunction with – a limitation in the neuromotor recruitment.
So, as we look at the common patterns that are outlined below, we cannot know without a physical examination whether we are addressing a ‘mere’ neural patterning (although we just saw how deeply that can run) versus structural restriction in non-contractile tissues.
We can have more certainty when the problem follows an injury or surgery that fascial adhesions are involved, as fascial ‘overdoing’ is a common healing response to injury, sustained inflammation, or surgical cutting and sewing (Hyman & Rodeo 2000).
Felt-like adhesions are readily and reliably palpable around old injuries and surgeries in the living, and are often found in dissection in conjunction with surgical intervention or other trauma (
Fig. 3.3
).
The best course of treatment will depend on clinical decisions based on our manual and observational assessments. Heavy-handed manual treatment or deep stretching is usually not necessary, or indeed can be detrimental when trying to enhance neuromotor patterning, but seems well-suited to deep tissue dehydration or adhesion, where neural facilitation trigger-point therapies, proprioceptive neuromuscular facilitation (PNF) or muscle-energy style stretches, or Feldenkrais-like decoupling, would likely need to be repeated many times or sustained over long periods of time to have any salutary effect on true fascial restrictions.
When your hand is a hammer, everything looks like a nail. Practitioners tend to see each problem and pattern through the lens of their own training, in terms of the strategic arrows already in their quiver. This is fine as long as we can admit to ourselves when our techniques are not working and change tack or refer out.
Figure 3.3
Fascial adhesion in dissection. Felt-like fascial adhesions are palpable in the living body and visible in the cadaver. Photo courtesy of Robert Schleip.
Now that we have made this distinction between patterns that are purely held in the neural system versus those that have been there so long (or in response to trauma) that they are now ‘written into’ the fascial system, we can go on to outline some characteristics of the fascial system.
Fascia as a holistic communicating system
Central to this new Anatomy Trains map is the functional unity of the connective tissue system. There are exactly three networks within the body that if magically extracted intact would show us the shape of the whole body, inside and out: the vascular system, the neural net, and the extracellular fibrous web created by the connective tissue cells – the fibroblasts and their cousins.
Large communities of cells need vast infrastructure to maintain their fluid biochemistry in crowded conditions, pressed by gravity, and surrounded by
air. It is an astounding feat of engineering that we take for granted every day:
•
A community of 70 trillion diverse, humming, and semi-autonomous cells, each built for undersea living, organizes itself to get up, walk around, grab a cup of coffee, and answer emails. Most organismic cell-collections, to avoid massive cell death, require a constant flow of fluids, exchange of gases, and a regular trade of food for waste. To preserve their genes, they must ultimately find another similar community of cells and go through a complex dance in order to reproduce.
•
Simultaneously, our body provides each cell with a mechanically stable environment, oceanic conditions of chemical exchange, and the information it needs to participate meaningfully in the day’s work.
•
Every living cell needs to be within four cell layers or so of the fluid exchange provided by the vascular system. Without the ability to deliver chemistry and suck away waste, any underserved area becomes stressed, then distressed, and will finally shrivel or burst and die, as happens in necrotic or gangrenous tissue. Secondly, every cell needs to be within reach of the nervous system’s signaling to regulate its activity with other cells in other areas of the body. And every cell needs to be structurally held in place (or directed in a flow, in the case of blood and other mobile cells) by the connective tissue net.
Any given single cubic centimeter taken from the body would contain elements of all three of these nets – neural, vascular, and fascial. Seen in such a systemic way, the idea of the body as simply a collection of parts begins to lose its luster. We all survive because we are an interwoven set of systems (
Fig. 3.4
).
•
Each of these nets is networked across the entire body from central organizing structures: from its central spinal cord with the bulbous brain plexus at one end, the nervous system
spreads throughout the body in the familiar radiating pattern, to form a sensory simulation of our inner and outer world and a coordinated behavioral response. Dripping through the synapses and interstitia of this system are the messenger molecules of hormones, neurotransmitters, and neuropeptides, regulating neural tone both up and down the system (Pert 1997).
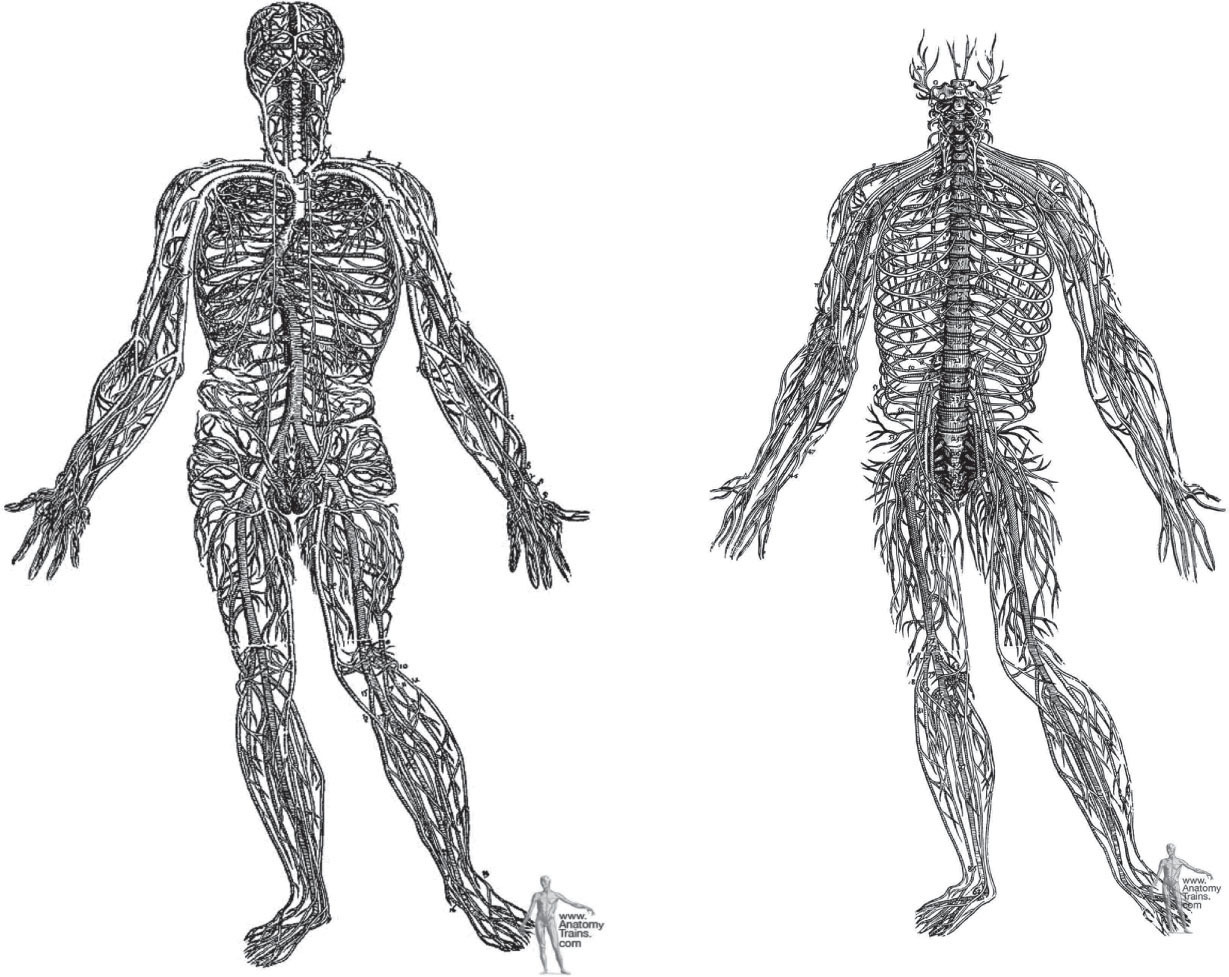
Figure 3.4
Vesalius nervous system and circulatory system. Vesalius detailed the neurological and vascular systems in 1548, a miraculous feat of dissection given the methods of the time. We have continued this model more or less unchanged for the ensuing 450 years. Reprinted courtesy of Dover Publications.
•
The circulatory system, from the axis of the aorta and the muscular heart, links the thousands of miles of capillaries and lymphatics in a circle that serves the chemical needs of the cellular community.
•
Thirdly, from the central armature of the notochord (the embryological form of the vertebral bodies and discs), connective tissue spreads out to create protective sacs and nets around all the cells, structures, and systems of the body, organizing stable mechanical relationships, allowing certain movements, discouraging others and providing the organization for all movements: voluntary, autonomic and physiological.
All three networks communicate throughout the body. The nerves carry sensory data in order to construct a second-to-second picture of the world, and send signals out to the muscles and glands, at speeds between 7 and 170 mph (~10–250 kph; Williams 1995). The fluid systems circulate chemistry around the body every few minutes, though many chemical rhythms fluctuate in hourly, daily, or as women know, monthly cycles.
The fibrous system communicates mechanical information – tension and compression – via the intercellular matrix of fascia, tendon, ligament and bone. This information is a vibration that travels at the speed of sound, about 700 mph – slower than light, but definitely faster than the nervous system. The speed of plastic deformation and remodeling in the connective tissue system, however, is measured in weeks, months, and even years. Thus, the fibrous system is both the fastest (in communicating) and the slowest (in responding) of the three.
Unlike the neural and vascular systems, this connective tissue net has yet to be well-mapped, because it has long been considered to be the ‘dead’ material that we need to remove to see the ‘interesting’ neural, vascular, muscular and other local systems. Because the connective tissue provides the divisions along which the scalpel runs to parse out other systems, the connective tissue has also been studied less as a system than other, more familiar systems.
So, as a thought experiment: what if, instead of dividing the body into individual identifiable structures, we were to dip it into a solvent that stripped away all the cellular material but left the entire extracellular matrix (ECM) intact (
Fig. 3.5
)? This is being done now in the creation of new artificial organs where the cellular material (of a healthy heart, say) is ‘shampooed’
out, and the remaining ECM is seeded with cardiac muscle cells to ‘grow’ a new heart – or, in a similar way, a kidney, or lung or an ear.
Figure 3.5
The endomysium gives us the clearest picture of what a body-wide extraction of the connective tissue matrix would look like. Photo courtesy of Peter Purslow. Reproduced from Purslow 1994, with kind permission from Springer Science and Business Media.
The beginning of form
This system of the connective tissue matrix can be seen as our ‘organ of form’ (Verela & Frenk 1987). From the moment of the first division of the ovum, the intercellular matrix of the connective tissue exists as a secreted glycosaminoglycan (mucous) gel that acts to glue the cells together (Moore & Persaud 1999). Around the end of the second week of embryological development, the first fibrous version of this net appears; a web of fine reticulin spun by specialized mesodermal cells on either side of the developing notochord (spine).
This net is the origin of our fascial web – our ‘metamembrane’ – the singular container that shapes our form and directs the flow of all our biochemical processes (Juhan 1987). This ECM, taken as a whole, not only unites the various elements of the body, it unites the many branches of medicine (Snyder 1975).
The ability of the connective tissue cells to alter and mix the three elements of the intercellular space – the water, the fibers, and the gluey ground-substance gel of glycosaminoglycans – produces on demand the wide spectrum of familiar building materials in the body (Williams 1995). Bone, cartilage, ligament, tendon, areolar, and adipose networks are all examples of this biological fabric. The body’s joints, the organ system of movement, are almost entirely composed of ECMs constructed by the various connective tissue cells.
The fiber/gel matrix becomes an immediate part of the environment of every cell, like cellulose in plants, or the coral’s limestone ‘apartment building’. The animal (and human) ECM, however, is very responsive to change – a pliable or limiting passive response to outside forces, as well as in active cellular response to damage or need (Schleip et al. 2012). Given that the matrix is a liquid crystal capable of storing and transmitting information, and that it is so intimately married to the lives of our cells, it can be seen as a living part of the body, not a dead packing material (Pirschinger 2007).
Fascial signaling and remodeling is part of our adaptive response to the needs of practical continuance; it is possibly part of the underlying substrate of our consciousness (Koob 2009).
Before we veer any farther into the realm of speculation, let us return to view the development of this network that supports the cellular community in a dynamically upright position.
Whole systems engineering
Thinking in ‘wholes’, attractive as it is to contemporary therapists, must in the end lead to useful maps. While ‘everything is connected to everything else’ as a philosophy is technically accurate, it leaves the practitioner adrift in a sea of connections, unsure as to whether that frozen shoulder will respond to work in the elbow, the contralateral hip, or to a reflex point on the ipsilateral foot. While any of these might work, useful maps are necessary to organize our therapeutic choices into a better strategy than ‘press and pray’.
The idea of ‘body as assembled machine’, inherited from Descartes and Newton, is so pervasive, and maps based on the ‘origin-insertion’ isolation of single muscles working as levers around fulcrums are so easily understood and useful, that it is difficult to think outside these parameters.
What can we learn when we shift from a ‘symptom-oriented’ view of the body to a ‘system-oriented’ one?
From the initial blastosphere, we see the involution of gastrulation and the subsequent lateral and sagittal folds are followed by literally thousands of others. This developmental origami takes the original simple three-dimensional spider’s web of surrounding and investing fascia and folds it into more
than a thousand compartments and divisions that we subsequently cut out and identify as separate ‘parts’. Initial folds create the dorsal cavity for the brain and ventral cavity for the organs and the coelomic bags surround each organ with a double-layered fascial sac.
One of the final folds brings the two halves of the palate together, which explains why a cleft palate is such a common birth defect.
With this image, we can get away from the shibboleth that there are some 600 muscles in the body. There is, in fact, only one muscle. One mind, and one muscle – contractile tissue is just folded into 600 pockets within the unitary fascial bag. We are so thoroughly imbued with the idea of origins, insertions, and actions of each individual muscle, and it is a mental struggle to rise above this concept, but the view from the mountain of wholeness is breathtaking.
One other holistic image useful in jumping out of this ‘machine made out of parts’ image so ingrained in our systems, is tensegrity geometry. The normal geometric picture of our anatomy is that the skeleton is a continuous compression framework, like a construction crane’s frame, or a stack of blocks, from which the muscles hang like cables.
This leads to the single muscle theory again – the skeleton is stable but segmentally moveable, and we parse out what each muscle does to that framework on its own across those segments, adding them together to analyze functional movement. A little thought, however, soon puts this idea out to pasture. Take the muscles away, and the long skeleton on tiny feet is anything but stable; take all the soft tissue away and the bones would clatter to the floor, as they do not stack in any kind of locked way.
If we can get away from the idea that bones are like girders, and muscles are the cables that move the girders, we are led to a class of structures called ‘tensegrity’ (a portmanteau signifying that the structural integrity lies in the balance of continuous tension). Originated by Kenneth Snelson and developed by Buckminster Fuller, tensegrity geometry more closely approximates the body as we live and feel it than does the old ‘crane’ model (Heartney 2009, Fuller 1975).
In the dance of stability and mobility that is a human moving, the bones and cartilage are clearly compression-resisting struts that push outward against the myofascial net. The net, in turn, is always tensional, always trying to pull inward toward the center. Both elements are necessary for stability, and both contribute to practical mobility.
In this new orthopedic model, the bony struts ‘float’ within the sea of tension provided by the soft tissues (
Fig. 3.6
). The position of the bones is thus
dependent on the tensional balance among these soft-tissue elements. This model is of great importance in seeing the larger potential of soft-tissue approaches to structure, in that bony position and posture is far more dependent on soft-tissue balance than on any high-velocity thrusting of bones back into ‘alignment’.
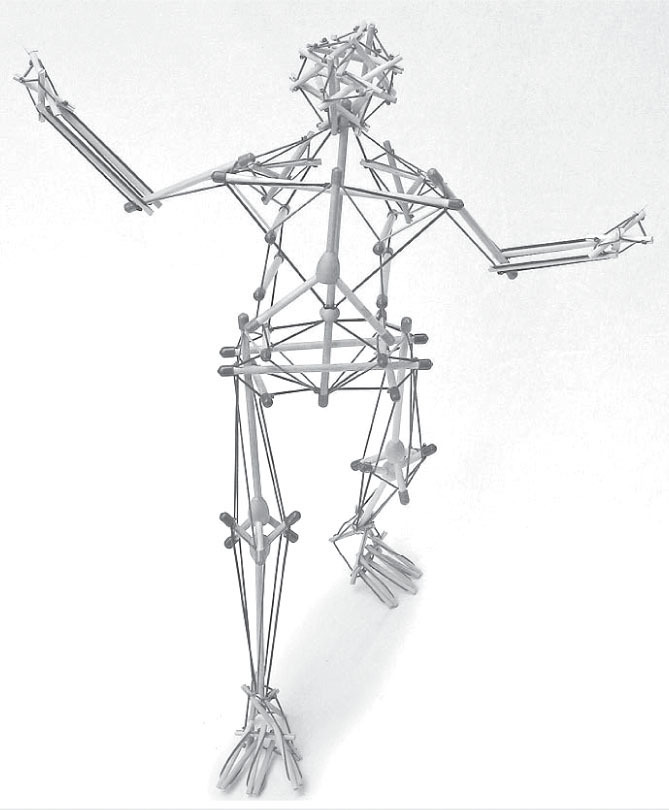
Figure 3.6
The tensegrity models created by Intension Designs mimic biological movement in a way that fulcrum and lever models do not. The interplay of tension and compression gives ‘proportional stability’ in a way that rigid structures cannot accommodate.
Photo courtesy of Tom Flemons. (For descriptions, short films, and the models themselves, contact designer Tom Flemons at
intensiondesigns.com
.)
The tendency for these structures to store energy, to ‘give’ and then restore themselves in an elastic way is very different from the rigidity of the levers and fulcrums model. This property commends tensegrity as a model of the body, which also gives and returns in this elastic way. When your shoulder is pressed to the wheel, the shoulder gives, the rib cage adjusts and the spine bends, the pelvis rotates, the legs set – there are a thousand small adaptive reactions all over the body, not a single setting of a rigidly locked system. Although difficult to show on a printed page, tensegrities show this kind of behavior, just like bodies.
These patterns of slight ‘give’ in multiple segments are, when held into the posture over a long period of time, the very constellation we need to see to make an effective fascial release treatment that will make structural sense to both the mind and the joints. Research supports the idea of tensegrity geometry, ruling mechanical transmission from the cellular level upwards, and macro-level models are becoming more anatomically accurate with each passing year (Turvey & Fonseca 2014).
How we move and are moved during our lives shapes this web, and the shape of the web, in turn, helps to determine our experience of living in our bodies.
Introduction to the Anatomy Trains
The Anatomy Trains’ myofascial meridians map the global lines of tension that traverse the body’s muscular surface, acting to keep the skeleton in shape, guide the available tracks for movement, and coordinate global postural patterns (
Fig. 3.7
).
Anatomy Trains describes an intermediate level between these overarching global considerations and useful detailed anatomy. These ‘myofascial meridians’ provide a practical transition between the individual parts and the whole of a human being – the gestalt of physics, physiology, stored experience, and current awareness that often defies mapping. The Anatomy Trains view of the body’s locomotor fabric opens new avenues to treatment consideration, particularly for stubborn chronic conditions and global postural effects.
At its simplest, myofascial meridians simply track the warp and weft through the parietal ‘myofascialature’. The basic rule is to follow muscular and fascial fibers in a relatively straight line without abrupt changes in depth or intervening fascial walls.
Where does this webbing run continuously in straight lines – lines that can transmit tensional forces that travel out from their local areas to create global effects via the interconnectedness of this overall network? The answer to this question provides a map for tracking myofascial strain transmission and the slight ‘give’ that allows this system to adapt as a whole.
Let us introduce the
dramatis personae
of the Anatomy Trains before going on to analyze some common postural compensations. Those already familiar with the system can proceed to the ‘BodyReading’ section that follows. For a more in-depth exploration of these concepts and their application, see the book
Anatomy Trains
(Myers 2014), or visit the site at
www.AnatomyTrains.com
for books, webinars, and DVD self-study courses on this skill.
Anatomy Trains provides a traceable basis for effective treatment at some distance from the site of dysfunction or pain. This new view of structural patterning has far-reaching implications for treatment strategies, especially for longstanding postural imbalances, unsound body usage, and sequelae from injury or insult.
Figure 3.7
(A,B) Anatomy Trains’ myofascial meridians map continuous lines of myofascial tension that generally run longitudinally along the body’s musculature.
The concept is very simple: if we follow the grain of the fascial fabric, we can see where muscles link up longitudinally. When this is done, there are 12 or so major myofascial meridians that appear, forming clear lines, or tracks, that traverse the body. A few rules and terminological considerations apply to their construction:
1.
Myofascial continuities must run in straight lines.
Since an Anatomy Train is a line of tensional pull, not compressional push, it must therefore travel in a mostly straight line. So, the first and major rule is that the fibers in adjacent myofascial structures must ‘line up’ in the fascia, without major changes of direction or depth. While the hamstrings and gluteus maximus might be functionally connected in running or climbing stairs, the change in direction and change of level between the two prevent them from being a fascial continuity. The hamstrings and erector spinae, however, are clearly connected in a mostly straight line via the sacrotuberous ligament.
2.
Fascia is continuous, while muscles are discrete – ‘tracks’ and ‘stations’.
The stripes of muscles and fasciae are termed ‘tracks’, and what are commonly known as muscle
attachments are termed ‘stations’ to emphasize the continuous nature of the fascial fabric. Muscles themselves may be discrete, but the fascia that contains them is continuous and communicates to the next structure up or down the line. The external oblique muscle and the serratus anterior muscle may be separate and have separate functions, but the sinew that envelops each of them is part of the same fascial plane, which communicates across their attachment points and beyond, in an overall functional continuity called the Spiral Line.
3.
Fascial planes can divide or blend – ‘switches’ and ‘roundhouses’.
Fascial planes sometime split into two planes or, conversely, two planes blend into one. We call these dividing places ‘switches’, and the physics of the situation will determine which plane takes how much of the force involved. The rhomboids thus communicate fascially with both the serratus anterior and the rotator cuff. Which line is employed will depend on the position, load, and orientation of these and surrounding structures. Places where many muscles meet and provide competitive directions for where a bony structure might be pulled – the ASIS, for instance, or the scapula – are termed ‘roundhouses’. See notes on another ‘roundhouse’ – the lumbar interfascial triangle (LIFT) – in
Chapter 1
.
4.
Deeper, single-joint muscles hold posture – ‘expresses’ and ‘locals’.
Monoarticular, or one-joint, muscles are termed ‘locals’, whereas multiarticular muscles are termed ‘expresses’. Posture is more often held in the deeper single-joint locals, not in the coordinating expresses that often overlie them. Thus, one looks to relieve a chronically flexed hip via the iliacus or pectineus myofascia, more often than the rectus femoris or sartorius, both of which also cross the knee and are therefore not capable of maintaining an angle in any one joint.
5.
When the rules get bent – ‘derailments’.
And finally, we sometimes encounter ‘derailments’, where the myofascial meridian does not utterly conform to the above rules, but works under particular conditions or positions. For instance, the line of fascia along the back of the body is a continuous string of fascia when the knee is straight, but ‘de-links’ into two pieces – one above and one below – when the knee is significantly flexed. This explains why nearly every classic yoga stretch for the Superficial Back Line, as we term it, has the knees in the extended position, and why it is easier to pick up your dropped keys with even slightly flexed knees than with straight legs.
Anatomy Trains lines
Each of these individual myofascial meridians can be viewed as:
1.
One-dimensional tensional lines that pass from attachment point to attachment point, from one end to the other
2.
Two-dimensional fascial planes that encompass larger areas of superficial fasciae, or
3.
Three-dimensional sets of muscles and connective tissues, which taken together comprise more or less the entire volume of the musculoskeletal system.
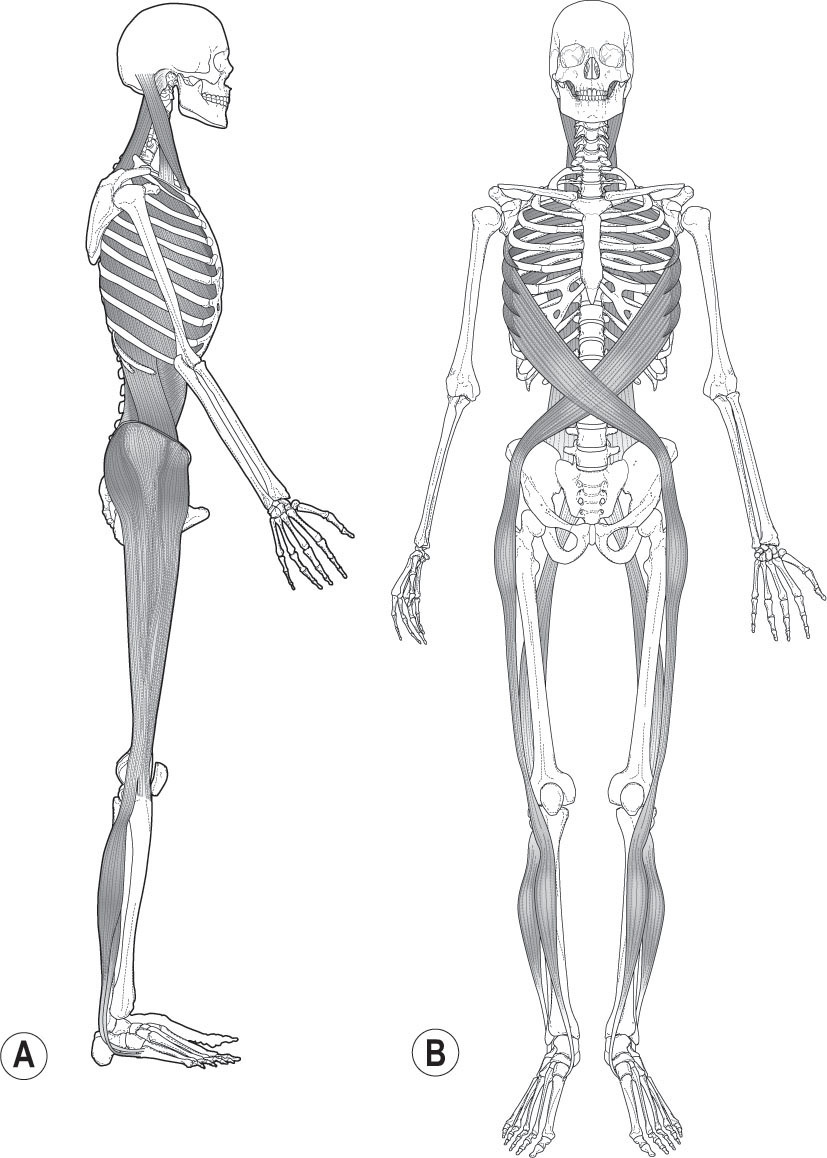
Superficial Front Line
The Superficial Front Line (SFL –
Fig. 3.8A
) runs on both the right and left sides of the body from the top of the foot to the skull, including the muscles and associated fascia of the anterior compartment of the shin, the quadriceps, the rectus abdominis, sternal fascia, and sternocleidomastoid muscle up onto the galea aponeurotica of the skull. In terms of muscles and tensional forces, the SFL runs in two pieces – toes to pelvis, and pelvis to head, which function as one piece when the hip is extended, as in standing.
In the SFL, fast-twitch muscle fibers predominate. The SFL functions in movement to flex the trunk and hips, to extend the knee, and to dorsiflex the foot. In standing posture, the SFL flexes the lower neck but hyperextends the upper neck. Posturally, the SFL also maintains knee and ankle extension, protects the soft organs of the ventral cavity, and provides tensile support to lift those parts of the skeleton that extend forward of the gravity line – the pubis, the rib cage, and the face. And, of course, it provides a balance to the pull of the superficial back line.
A common human response to shock or attack, the startle response, can be seen as a shortening of the SFL. Chronic contraction of this line – so very common after trauma or the slings and arrows of even a normal childhood – creates many postural patterns that lead to pain patterns.
Superficial Back Line
The Superficial Back Line (SBL –
Fig. 3.8B
) runs from the bottom of the toes around the heel and up the back of the body, crossing over the head to its
terminus at the frontal ridge at the eyebrows. Like the SFL, it also has two pieces, toes to knees and knees to head, which function as one when the knee is extended. It includes the plantar tissues, the triceps surae, the hamstrings and sacrotuberous ligament, lumbosacral fascia, the erector spinae, and the epicranial fascia.
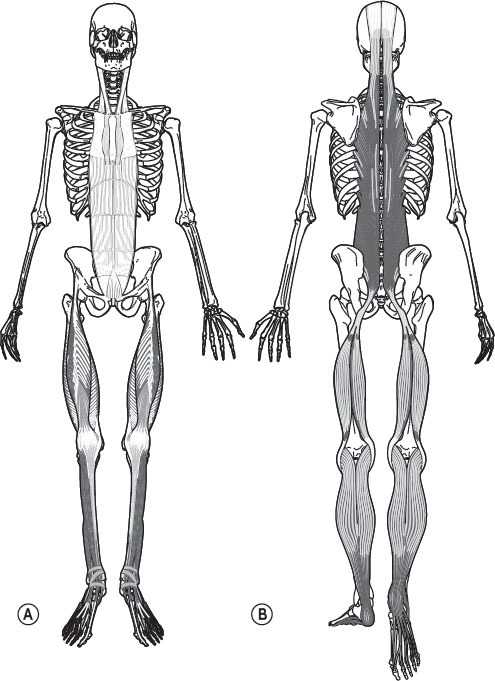
Figure 3.8
Two of the Anatomy Trains’ myofascial meridians, the Superficial Front Line (A) and Superficial Back Line (B), counterbalance each other on either side of the skeleton. Reproduced with the kind permission of Lotus Books.
The SBL functions in movement to extend the spine and hips, but also to flex the knee and ankle. The SBL lifts the baby’s eyes from primary embryological flexion, progressively lifting the body to standing. In psychomotor terms, dysfunction in the SBL is often linked to difficulties in attaining full maturity.
Posturally, the SBL maintains the body in standing, spanning the series of primary and secondary curves of the skeleton (including the cranium and heel in the catalogue of primary curves, and knee and foot arches in the list of secondary curves). In very basic terms, the primary curves – heel, sacrum, mid-back, and occiput – are designed to align over each other, and the secondary curves – neck, low back, knees, and arches – should align over each other.
Because of its ‘all-day’ postural function, the SBL is a more densely fascial line than the SFL, with strong fascial bands along its length, and a predominance of slow-twitch fibers in the muscular portion.
Lateral Line
The Lateral Line (LL –
Fig. 3.9A
) traverses each side of the body from the medial and lateral midpoints of the foot around the fibular malleolus and up the lateral aspects of the leg and thigh, passing along the trunk in a woven pattern that extends to the skull’s mastoid process.
Figure 3.9
Lateral and Spiral Line. (A) The Lateral Line runs from the outer arch to the ear up the lateral side of the body and acts primarily to stabilize movement. (B) The Spiral Line acts to create or stabilize rotational and oblique motions. Reproduced with the kind permission of Lotus Books.
In movement, the LL creates lateral flexion in the spine, abduction at the hip, and eversion at the foot, and operates as an adjustable ‘brake’ for lateral and rotational movements of the trunk.
The LL acts posturally like guy-wires of a tent, to balance the left and right sides of the body. In this way, the LL restrains more movement than it creates in the human, maintaining a stable base for the flexion–extension that characterizes our direction through the world, restricting side-to-side movement that would otherwise be energetically wasteful.
Many of our side-to-side asymmetries are held in the LL, so that complementary parts of the LL can maintain the compensation. In the short leg pattern, for instance, the LL will be held short from hip to ankle on the short leg side, but held short hip to shoulder on the contralateral side.
Spiral Line
The Spiral Line (SL –
Fig. 3.9
B) winds through the three cardinal lines, looping around the trunk in a helix, with a long jump rope in the legs from hip to arch and back again. It joins one side of the skull across the midline of the back to the opposite shoulder, and then returns to the side we started on, running across the front of the torso to the hip, knee, and arch of the foot. The line then returns up the back of the body to the head.
In movement, the SL creates and mediates rotations and oblique movements in the body. The SL interacts with the other cardinal lines in a multiplicity of functions from the watchspring-like rotations of walking to the multiple spirals of a golf swing.
In posture, the SL wraps the torso in a double helix that helps to maintain spinal length and balance in all planes. The SL connects the foot arches with tracking of the knee and pelvic position. The SL often compensates for deeper rotations in the spine or pelvic core.
Since side bends are almost always accompanied by spinal rotations, any strong side-to-side asymmetries will involve compensation in the SL. People with these patterns will often look a bit ‘wonky’ temporarily as they move out of these compensatory patterns toward health.
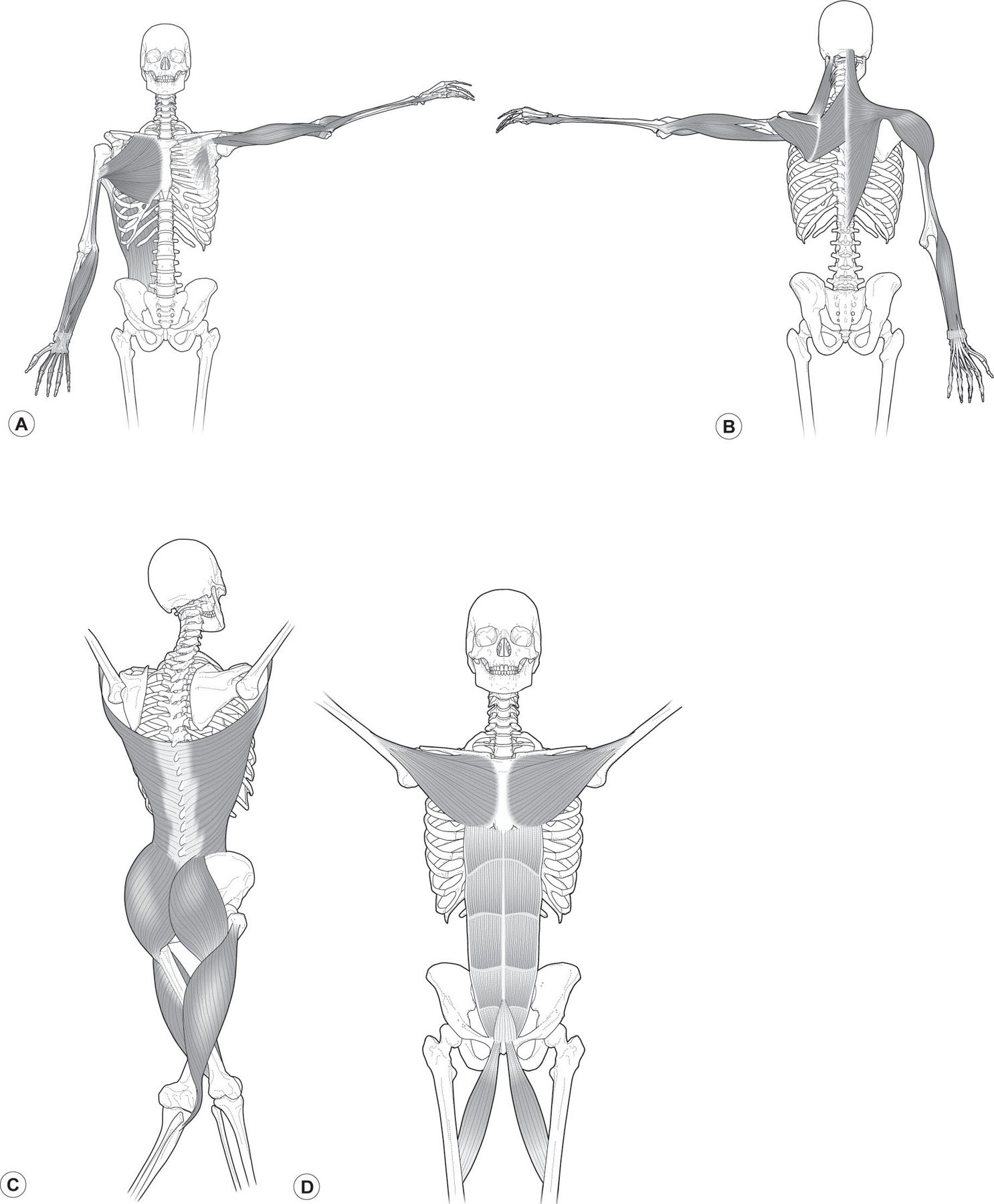
Arm Lines
The four Arm Lines (ALs –
Fig. 3.10A,B
) run from the front and back of the axial torso to the tips of the fingers. They are named for their planar relation in the axilla, and they are roughly parallel to the four lines in the leg. These lines connect seamlessly into the other lines, particularly the Lateral, Functional, Spiral, and Superficial Front Lines.
In movement, the ALs place the hand in appropriate positions for the tasks before us – examining, manipulating, or responding to the environment. The ALs act across 10 or so joint levels in the arm to bring things to us or to push them away; to push, pull, or stabilize our own bodies; or simply to hold some part of the world still for our perusal or modification.
The ALs affect posture indirectly, since they are not part of the structural column. Given the weight of the shoulder and arm assembly, however, displacement of the shoulders in posture or in a movement strategy will affect other lines. Conversely, structural displacement of the trunk in turn affects the arms’ effectiveness in specific tasks and may predispose them to injury.
Functional Lines
Two of the Functional Lines (FLs –
Fig. 3.10
C,D) join the contralateral girdles across the front and back of the body, running from one humerus to the opposite femur and vice versa. A third line, the Ipsilateral Functional Line (not pictured), runs from the lateral edge of the latissimus to the inferior edge of the external abdominal oblique over the ASIS to the sartorius, running to the medial tibial condyle and the pes anserinus.
Figure 3.10
Arm Lines and Functional Lines. (A,B) The four Arm Lines go to the four corners of the hands from the axial skeleton. (C,D) The Functional Lines extend the arm lines to the contralateral hip. Reproduced with the kind permission of Lotus Books.
The FLs are used in innumerable active movements, from walking to the most extreme sports. They act to extend the levers of the arms to the opposite leg, as in a kayak paddle, a baseball throw, or a cricket pitch (or leg to opposite shoulder in the case of a football kick). Like the SL, the FLs are helical, and thus help create strong rotational movement. Their postural function is minimal; their role in coordinating limb movements to the trunk is invaluable.
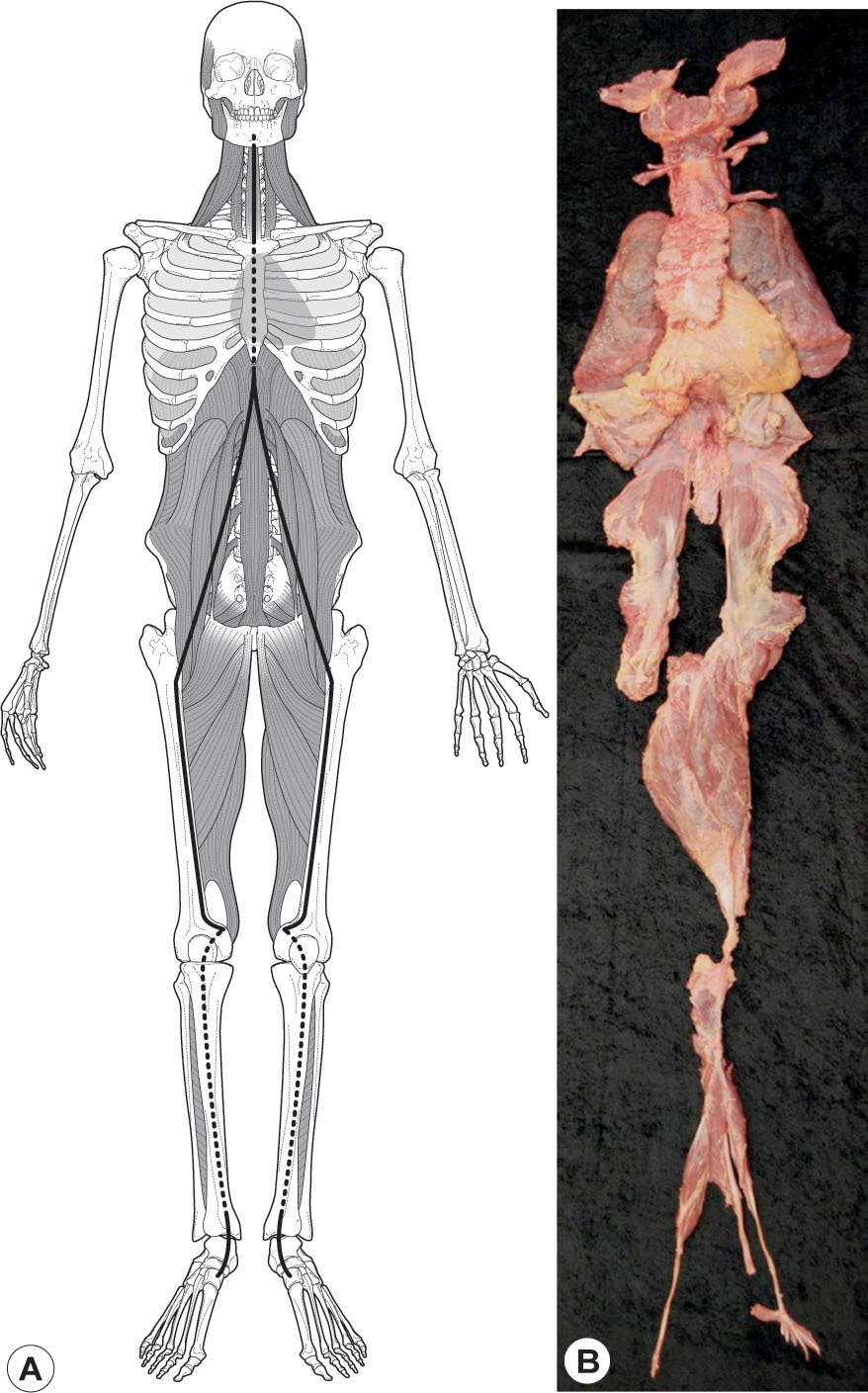
Deep Front Line
The Deep Front Line (DFL –
Fig. 3.11
) forms a complex core volume from the inner arch of the foot, up the inseam of the leg, into the pelvis before and behind the hip, and up the front of the spine to the bottom of the skull and the jaw. This ‘core’ line lies between the Front and Back Lines in the sagittal plane, between the two LLs coronally, and is wrapped circumferentially by the Spiral and Functional Lines. This line contains many of the more obscure supporting muscles of our anatomy, and because of its internal position, it has a strong fascial density around the muscular tissue.
Structurally, this line has an intimate connection with the arches, the hip joint, lumbar support, and neck balance. Functionally, it connects the ebb and flow of breathing, dictated by the diaphragm, to the rhythm of walking, organized by the psoas. In the trunk, the DFL is intimately linked with the front of the spine and the autonomic ganglia, and thus uniquely involved in the sympathetic/parasympathetic balance between our neuromotor ‘chassis’ and the ancient organs of cell-support in the ventral cavity.
The importance of the DFL to posture, movement, and attitude cannot be over-emphasized. A dimensional understanding of the DFL is necessary for successful application of nearly any method of manual or movement therapy. Because many of the movement functions of the DFL are redundant to the superficial lines, lack of participation in the DFL can be barely visible in the outset, but these small dysfunctions will gradually lead to larger problems. Restoration of proper DFL functioning is by far the best preventive and treatment-sustaining measure for structural and movement therapies.
Figure 3.11
Deep Front Line. (A) The Deep Front Line forms the body’s core, from the inner arch to the inner hip, up around the ventral cavity to the skull. (B) The Deep Front Line dissected as one myofascial meridian (at least for the left leg) including the hyoid and jaw bone as ‘sesamoids’ within the line.
BodyReading the lines
In order to describe the anomalies in the lines and the relationships among them, we will use a simple set of four terms that describe skeletal relationships. These readily understandable terms can be used to create a sketch of the generalized body pattern (which is what we will be doing here), but they can also bear the weight of a detailed inter-segmental analysis or argument.
Tilt
describes simple deviations from vertical or horizontal; in other words, a body part or skeletal element that is higher on one side than on another. We are using this word with its readily understood common meaning.
The term is modified by the direction to which the top of the structure is tilted. Thus:
•
In a left side tilt of the shoulder girdle, the client’s right acromion would be higher than the left, and the clavicles would lean to the client’s left
•
A posterior tilt of the rib cage would involve the upper ribs going back relative to the lower
•
In a right side tilt of the head, the left ear would be higher than the right, and the planes of the face would tilt to the right
•
In a posterior tilt of the head, the eyes would look up, the back of the head approaches the spinous processes of the neck, and the top of the head moves posteriorly.
For clarity in communication and accuracy in translating this language into soft-tissue strategy, it is very important to understand ‘relative to what’: an ‘anterior pelvic tilt relative to the femur’ is a useful observation, a simple ‘anterior pelvic tilt’ opens the door to confusion – to the floor? To the femur? To the rib cage?
Bend
is a series of tilts resulting in a curve, usually applied to the spine. The normal lumbar curve thus has a back bend, and the normal thoracic spine a forward bend. If the lumbar spine is side bent, this could be described as a series of tilts between each of the lumbar vertebrae, which we usually summarize as a bend, which can, again, be to either side, forward or back, and is named for where the top of the bend is pointing relative to the bottom.
Ergo, in the common situation where the pelvis is tilted to the left, and the lumbars correct that tilt with a bend so that the rib cage can be straight up, those lumbars have a right bend, as the top lumbar points to the right compared to the lower.
Tilts and bends are modified by left, right, anterior, or posterior.
Rotation
occurs around a vertical axis in the horizontal plane, and thus often applies to the femur, tibia, pelvis, spine, head, humerus, or rib cage.
Rotation is named for the direction in which the front of the named structure is pointing – right or left. For instance, in a right rotation of the head (relative to the rib cage), the nose or chin would face to the right of the sternum. Notice that, if the rib cage were left rotated relative to the pelvis, the head could be right rotated relative to the rib cage and still be neutral relative to the pelvis or feet.
Thus relativity is very important in assessing postural rotations.
Also notice that if the rib cage is left rotated, the accompanying spinous processes would be moving to the right, viewed from behind, but would still involve a leftward rotation of the vertebrae, since the front of the vertebrae are still turning left.
In paired structures, we use medial or lateral rotation. While this is in common use as regards femoral or humeral rotation, we extend this vocabulary to all structures. What is commonly called a ‘protracted’ scapula would, in our vocabulary, be a ‘laterally shifted and medially rotated’ scapula, since the anterior surface of the scapula turns to face the midline.
(One might question: Why use that long phrase when the simple term ‘protracted’ is available? While ‘protracted’ is fine in general conversation, we must note that it is not clear enough to specify treatment. Each case of protraction involves differing degrees of lateral shift, medial rotation, and anterior tilt. Additionally, there may be upward rotation involved (which we would term a medial tilt). Each of these designations specifies a piece of anatomy for examination and treatment – i.e. we would look to the 5th rib slip of the pectoralis minor for the anterior tilt, but turn to the 2nd and 3rd rib slip for more medial rotation. Likewise, different sections of the serratus anterior would be indicated in medial tilt or medial rotation. In other words, generalized words like protraction lead to generalized treatment strategies, whereas we can use this method to generate individualized protocols.)
Shift
(translate, shunt) is an additional useful term for displacements (right–left, anterior–posterior, or superior–inferior) of the center of gravity of one structure relative to another. Balinese dance involves a lot of head shifting – side-to-side movement while the eyes stay horizontal. The rib cage of fashion models is commonly shifted posteriorly while staying relatively vertical relative to the ground.
Such shifts, of course, commonly co-occur with tilts, bends, and rotations. We can use the terminology to specify these particular relationships when called for, but using ‘left side shifted rib cage’ or ‘head shifted to the right relative to the pelvis’ is a useful shorthand when making an initial evaluation.
The mobile scapula is commonly shifted in any of the six modifying directions. The pelvis is commonly described as being anteriorly or posteriorly shifted relative to the malleoli, with the understanding that some tilts must occur along the way for that to happen.
We may employ these terms and the Anatomy Trains lines to understand global postural patterning. The following examples are designed to demonstrate common postural problems. The reader is advised to see them in terms of fascial patterning and myofascial meridians, and to outline strategies for creating sturdier, self-sustaining structural balance.
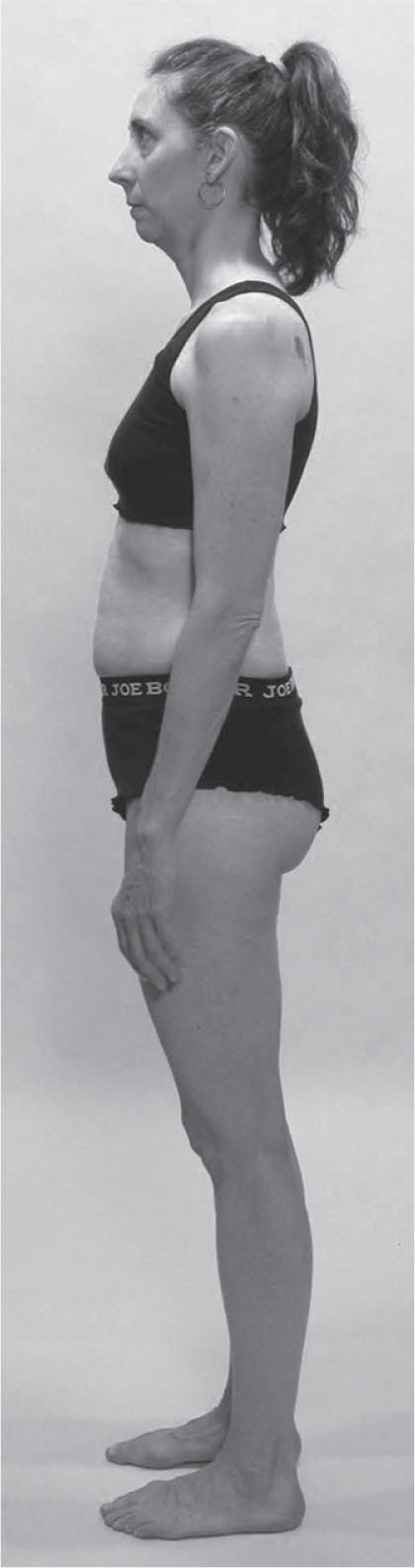
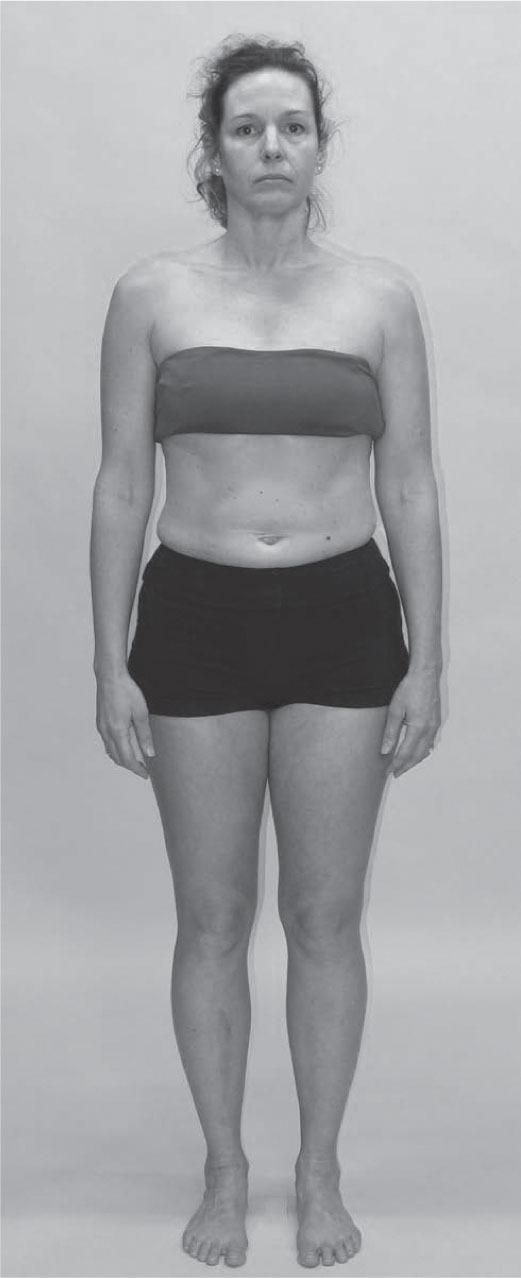
Head forward posture and front–back balance
No one is typical but the woman in
Figure 3.12
typifies a very common posture in both the Western and Asian world, and serves as a point of discussion about the fascial fabric relations between the Superficial Back, Superficial Front, and Deep Front Lines.
In terms of the language just outlined, this person demonstrates a number of tilts and shifts:
•
The femur is anteriorly tilted relative to the tibia, a common correlate of posteriorly shifted (hyperextended) knees.
•
The pelvis is posteriorly tilted relative to the femur.
•
The rib cage is posteriorly tilted relative to the pelvis.
•
The lower cervicals are anteriorly tilted relative to the rib cage.
•
The mid-cervicals have a posterior bend that allows the head to be horizontal.
•
The head is anteriorly shifted relative to the upper ribs and shoulders, and the pelvis has shifted forward over the forefeet. The result is that the SBL is acting like a bowstring from shoulder to ankle. (In this case, since the knees are shifted back into hyperextension, the SBL is more accurately short here between T6 and the knee.)
•
The tightening of the SBL puts much of the SFL under eccentric strain, which often shows up, naturally enough, in the abdomen/anterior lumbar area. This eccentric loading does not, however, extend to the whole line. The top and
bottom portion of the SFL are often both concentrically shortened in this pattern.
•
The lowest track of the SFL, the anterior crural compartment of the tibialis anterior and long toe extensors, will be held short and often pinned down the retinacula and tibia along the way.
Freedom and de-adhesion for this compartment is crucial to undoing the pattern above.
•
At the top, the section of the line from the mastoid process to the mid-sternum is also shortened and will need to be lengthened. Under the general rule of ‘lengthen the short before strengthening the weak’, toning the abdomen will make no sense and may have either little positive, or an actual negative, effect until 1) the middle part of the SBL is lengthened (thoracolumbar fascia, the erector spinae, and fasciae in the region of the outer edge of the quadratus lumborum); and 2) both ends of the SFL (sternocleidomastoid and the anterior crural compartment) are eased.
•
In the legs as a whole, we have a very common situation whose solution requires the unique perspective of the fascial fabric. Regardless of the tonus of the various individual muscles, the superficial, profundis, and myofascial tissue layers all along the front of the leg need to be lifted (allowing them to de-adhere and ‘glide’ on underlying layers), and the corresponding layers along the back of the leg need to be de-adhered and dropped toward the heel.
•
We manual therapists so often think in terms of ‘what’s short?’. In this case, we need to think about the relationship between these fascial planes, and, by freeing fascial glide among layers, ‘redrape the toga’ of the myofascia as a whole over the skeleton beneath so that it hangs properly. We have found no other strategy that so reliably and sustainably allows the client to recenter the pelvis over the ankle and (in this case) allow the locked knees to ease.
•
The LL will be part of the treatment plan here, where the middle ribs are pulled inferiorly and posteriorly by the internal intercostals and internal oblique. Because of the hyperextended knees, the lowest track of the LL, the fibularis muscles, will be locked to maintain plantar flexion against the pull of the iliotibial tract above.
•
Any of the strategies outlined above will be for naught if the DFL is not also included in the treatment plan. Although obscured by the outer lines in this photo, the adductor section of the DFL will rotate forward inside the femur, putting most of the adductor magnus in concentric loading, like the hamstrings, and the anterior adductors into eccentric strain, like the hip flexors.
•
Although this woman will likely report her hip flexors as ‘tight’, the last thing she needs is lengthening in the psoas complex (pectineus, psoas, iliacus, all originating from the lesser trochanter). As the mid-SBL eases in the back and the pelvis can ease posteriorly to center over the ankles, she will be able to ‘gather’ and balance her hip flexors. Central to this change will be an ease in the deep lateral rotators (which in this case are acting as ‘short hip extensors’, helping to keep the pelvis simultaneously pushed forward and held into posterior tilt on the femur).
•
Once the upper SFL is freed from sternum to mastoid, the final, and essential, part of the DFL that needs to be released runs up the visceral cavity from the diaphragm to about C4. This will definitely involve the anterior scalene and attendant fascia, and possibly the cupola of the lungs as well. In this case, the transversus thoracis and pericardium can be approached with visceral or osteopathic indirect manipulation.
•
Unhooking this deep downward pull in the DFL is crucial to correcting the head forward
posture and restoring a sturdy and free front–back balance. Despite its centrality, freeing these tissues (in the author’s experience) should follow the strategies outlined above for the more superficial lines; it is not advisable to go directly to this primary problem without freeing the outer fascial pieces of the puzzle first.
•
Although individuals vary, here are a few common palpation and strategy tips for these pattern-types:

In hyperextended knees, palpate all the plantar flexors to see which one(s) are holding the ankle into plantarflexion. Often the fibularii of the LL or the deep posterior compartment (DFL) will be the culprits, although the deeper part of the soleus is involved in the stronger patterns.
In hyperextended knees, palpate all the plantar flexors to see which one(s) are holding the ankle into plantarflexion. Often the fibularii of the LL or the deep posterior compartment (DFL) will be the culprits, although the deeper part of the soleus is involved in the stronger patterns.
With the anterior shift/posterior tilt of the pelvis so common in today’s ‘gym-trained’ people, the hip flexors will palpate as very ‘tight’, but this will be eccentric tension, and will not respond to stretching or manipulation until the ‘short hip extensors’ (the deep lateral rotators) are functionally lengthened.
The lower ribs are held forward and will palpate as sluggish or unmoving in the breath movement. Any number of fascial release, trigger point or facilitated stretching techniques will help their buried ribs to participate in breathing again.
Corresponding to the lower ribs, palpate the anterior scalene under the clavicular head of the sternocleidomastoid to find a steel strap linking the lower neck to the upper ribs. Use the breath, stretch, or manipulation to release it.
The woman shown in
Figure 3.13
presents a similar pattern, so we will point only to the instructive differences.
The SBL bowstring goes all the way from shoulder to ankle here, so the ankle is plantarflexed rather than dorsiflexed under hyperextended knees, as in the previous example. Thus, the tilt that brings the pelvis over the forefoot involves the whole leg. The strategy of ‘up the front, down the back’, in terms of
tissue relation, still applies. Look at the knee – can you see how the front of the knee tissue is down relative to the tissue behind the knee?
The pelvis is more anteriorly tilted in this case, and we can imagine that we will find a good amount of tension in both the hip flexors (including the inferior-medial fibers of the psoas) and the hip extensors, the deep lateral rotators (notably piriformis), which will show a combination of concentrically and eccentrically loaded tissue in this pattern. The rib cage is not as collapsed onto the pelvis; the ‘core’ of the DFL is more intact.
This neck does not have such a strong posterior bend, but nevertheless the lower neck is similarly pulled into the upper ribs and chest cavity. The lower ribs are shoved forward to lift the head above the body in what looks to be an ‘I am strong’ attitude. Opening up the posterior portion of the lower ribs and the lumbar portion of the erectors is insufficient to the task, as it will leave her with her head jutting forward in a way that will make no sense to her. The complementary compensation requires opening up the deep front of the neck and first two ribs for the body to find a new stability that ‘makes sense’ kinesthetically.
The last of our front–back comparisons (
Fig. 3.14
) presents other variations on this common theme. In this case:
•
The anterior tilt that creates the (milder, here) anterior shift of the pelvis occurs in the tibia, creating dorsiflexed ankles but a vertical femur. The knees are flexed, not locked back.
•
The pelvis is anteriorly tilted relative to both the femur and the rib cage.
•
The rib cage shows a posterior shift with little tilt either forward or back. There is a bit of posterior tilt, somewhat disguised by the protraction (medial rotation in the language outlined above) of the scapulae.
•
The head forward posture is more evident as exactly that – the center of the head is literally forward of any other gravitational center other than the lower arm.
Strategically, this posture has commonalities with the others, but to add another element that would be of help when we see this ‘slumped’ pattern: the SLs are both slack to allow this pattern. The SLs (see
Fig 3.9
) cross between the shoulder blades in back and across the umbilicus in front. By toning the SLs – boxing or kayaking are both excellent ways to do this – the belly would be brought back and the mid-back and medial scapulae supported forward, correcting much of the low-energy stance we see here.
Aside from points mentioned above, palpate the line between the pectoralis major and the rectus abdominis along the 6th and 7th ribs, to find a fascial strap that extends horizontally from slips of the serratus anterior across the front of the chest. Creating ‘glide’ between the planes of this fascial ‘bra line’ will be key to adaptive structural change in this client, to improve the upward range of movement for the shoulder–arm complex, as well as setting the stage for re-orientation of the head.
Now we turn our attention to right–left differences, which can be most easily seen from the front or back views. The LL differences will be most obvious, of course, though these are often underpinned by differences in the DFL sandwiched between the two LLs. (We have already noted that it is sandwiched sagittally between the SBL and SFL.)
The practical reality of human postural compensation is that side-to-side differences most often involve rotations as well, though these are difficult to parse in two-dimensional photographs.
The woman photographed in
Figure 3.15
presents some clear differences from right to left.
Her right Lateral Line is shorter from hip to lateral arch than its complement on the left. The left LL is shorter between hip and shoulder than the right, and then it switches again in the neck, where the right is shorter. This alternation between upper (trunk) and lower (leg) LLs is a common compensation pattern, but how the neck responds is variable and must be assessed individually.)
Strategically, lengthening the shorter myofascial lines is a good preparation for training tone in the antagonist eccentrically loaded portions. (‘Lengthen the short before you strengthen the weak.’) As stated, this type of pattern when released in this way will almost certainly reveal some rotations, which we can see in nascent form here: the pubis points to her left, the shoulder girdle comparatively to the right, and the neck around to the left again. The prominent sternocleidomastoid on the left is a giveaway here, required to give head stability when this posture goes into motion.
Her right SL, from the right side of the head around the left shoulder to the right hip, is significantly shorter than its left complement, which is another cue to the involvement of rotations in this apparent side-to-side compensatory pattern.
In this case, the DFL is maintaining its integrity pretty well, despite the multiple compensations going on around it. The core support on the inner line of the leg and up the front of the spine is fairly well intact.
In cases where right–left differences predominate, palpation of both sides to notice differences is essential to precise treatment. In this model, you would find the right splenii and scalenes shorter and denser on her right, the quadratus lumborum shorter on the left, and fascial density differences between the hip abductors on either side.
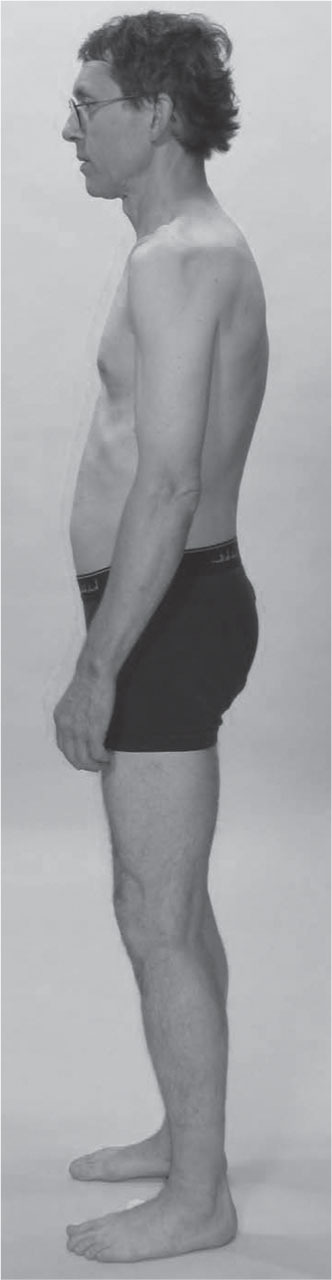
The trained dancer photographed in
Figure 3.16
shows us another common Lateral and Spiral Line pattern.
The most evident aspect of this pattern is that the rib cage is shifted to her left relative to the pelvis. To take this measure quickly in practice, measure the horizontal difference between the waist and the greater trochanter. (Using the space between the arms and the body is an unreliable measure.) If it is significantly different on the two sides, as it clearly is here, then look for a lateral shift of the ribs.
In this case, the left upper Spiral Line is shorter than the right, pulling the ribs into a right rotation.
This woman, despite the length in her waist, will have a very complex pattern in the DFL, quadratus lumborum and surrounding thoracolumbar fascia, as well as psoas major, which will present differing challenges on each side. The spine will have a significant two-bend with concomitant rotations pattern between the sacrum and mid-thoracic region. The head is following the ribs in being shifted to the left, so we can surmise that the neck is less strained than in the previous model.
In this case, however, much of the weight of the upper body falls on the left leg, which is compensated in the lower SL. The right anterior SL in the left leg is pulled or collapsed down into the inner arch, and the corresponding posterior line – the fibularis longus and biceps femoris pulled upward to the sacroiliac joint. The short head of the biceps appears to be maintaining a lateral rotation of the tibia on the femur. Her right leg shows hints of the same pattern but to a much lesser degree.
Although myofascial tonus in this young dancer is likely to be quite even across her body, she is built around the radical differences in the psoas complex and deep spinal muscles due to the lateral shift and rotation. We could anticipate that her superficial balance will be disturbed during the rebalancing of her lumbopelvic core, so it is best to undertake this work outside her performance season.
It would also be instructive to palpate the inside and outside of the knee on both sides, to feel how the ‘toga’ of the fascia profundis (fascia lata above, crural fascia below) is being pulled up on the outside and down on the inside of her left knee, compared to the right.
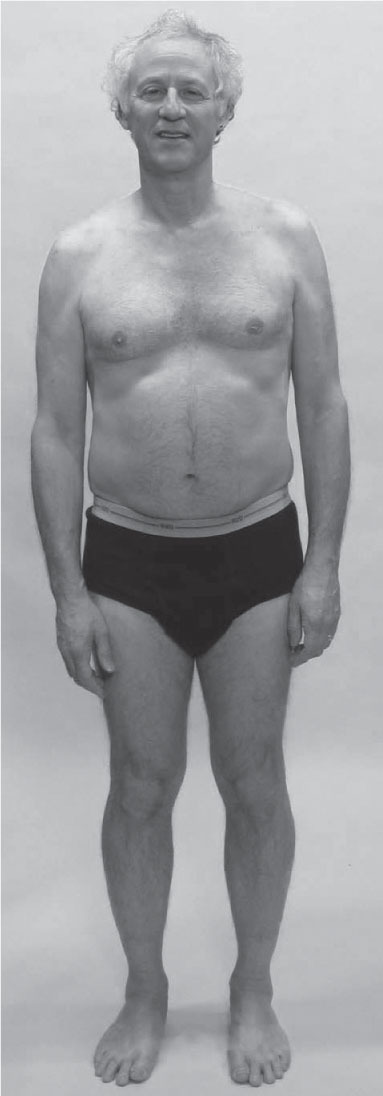
The man shown in
Figure 3.17
presents a different set of challenges.
The collapse of the entire upper body into the right arch is very dramatic – perhaps a true anatomical leg-length difference. The inner line of the right leg – the DFL – is pulling from the arch to the groin.
The pelvis, rib cage, and shoulders are all tilted to the right. The neck must perforce have a left bend to retain the eyes and vestibular system on a horizontal plane, albeit shifted to his right compared with the rest of the body. The amount of eccentric/isometric strain in the left side of the neck must be considerable.
The entire right LL requires a lift, and concomitant lowering of the left LL. The DFL on the right side needs to be lifted and stabilized through the groin. The left SL is shorter than the right, drawing him around into a left thoracic rotation.
•
Palpation of the two sides of the neck would be very instructive here, as all the deeper muscles would be arranged quite differently.
•
The left upper shoulder muscles will palpate as short and thick, as they are attempting to hold his head on.
•
Palpation of the hip flexors on both sides would be instructive to see exactly where each hip joint is positioned.
The young woman illustrated in
Figure 3.18
presents an odd pattern, variants of which can nevertheless be seen fairly often. Like the dancer shown in
Figure 3.16
, we can see the significant left shift of the rib cage as evidenced by the waist-to-trochanter horizontal measurement. The hips and legs themselves have very little compensation (as of yet, she is young).
The rib cage is, however, also tilted to the right, so that the shoulder girdle is tilted to the right along with it. Resist the temptation to fault the shoulder girdle; many people look first at the shoulders and start working. The shoulders are often not at fault; they are doing the best they can with the rib cage they have to sit on.
Interestingly, both the neck and the head continue the rightward tilt, instead of leveling out like the man in
Figure 3.17
.
Like the dancer in
Figure 3.16
, she will have complex tissue disposition on both sides of the lumbar spine – quadratus lumborum and psoas – which will need to be approached differently on each side.
The young woman looks oddly different on her left and right sides in the upper body, despite the symmetry in her lower body. Obscure the upper left side and imagine it doubled over; then do the same with the right. The left side is square, a swimmer’s upper body, whereas the right is sloped and less assertive.
Specifically, palpate the LLs and SBLs to feel the fascial tonus differences between the left and right support for the trunk.
Like the man in
Figure 3.17
, this woman will probably have dense, solid upper shoulder muscles on her left but the solution is not to work on those muscles, but on the shortness deep in her right waist. Relieve that shortness and the shoulder muscles will soften of their own accord.
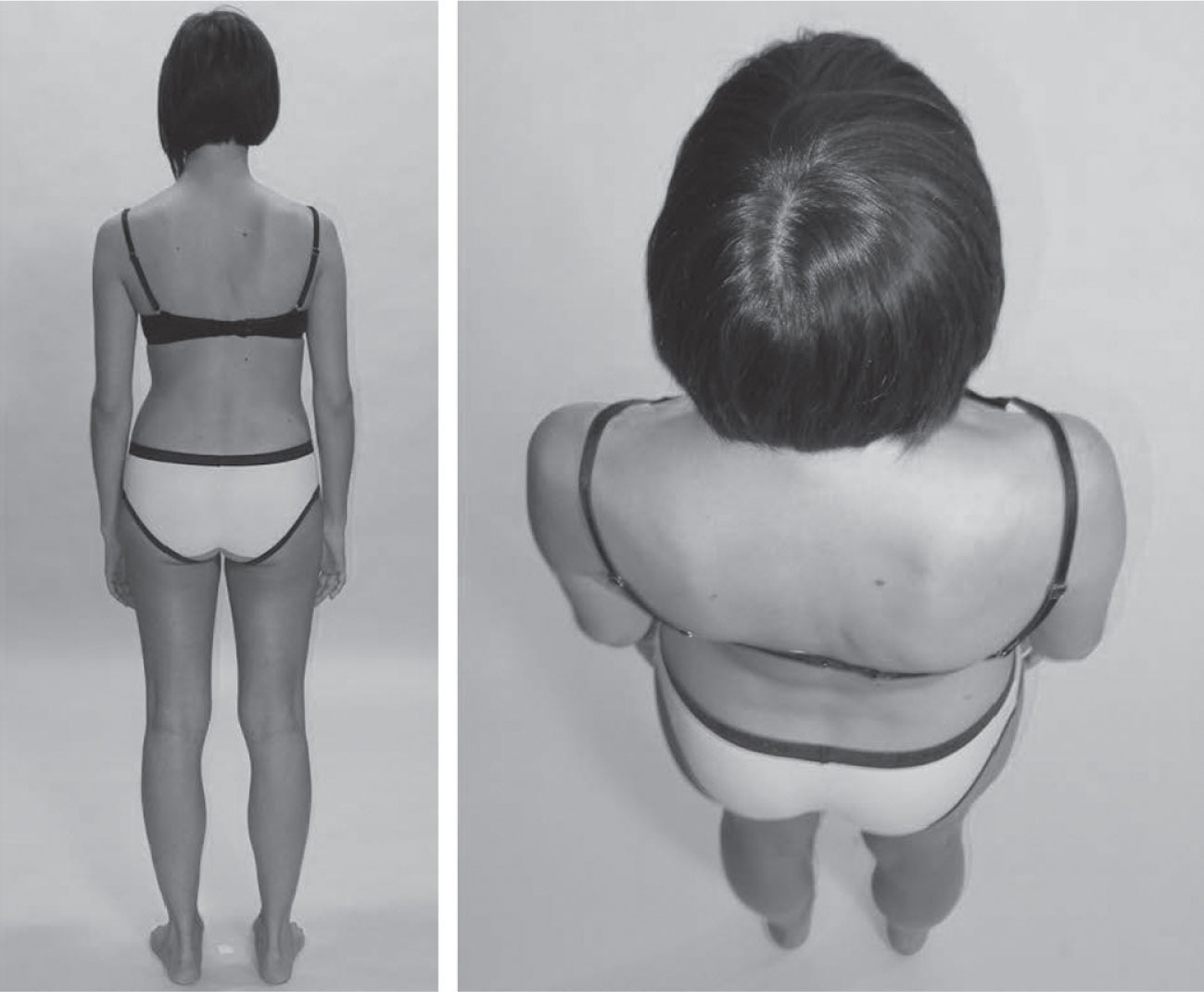
The woman photographed in
Figure 3.19
, seen from behind, shows the same kind of displacement of the ribs over the hips as seen in
Figure 3.18
and in the dancer (see
Fig. 3.16
), this time to the right, using the waist-to-trochanter measure.
For this model, we include a picture from the ‘breathing down her neck’ perspective; a position we recommend the practitioner observe, as it foreshortens the spine and makes the assessment of spinal rotations easier. Below, there is an obvious rotation to the right between the pelvis and the feet, which we can assess by drawing a line from heel to heel, and comparing it to the line from posterior superior iliac spine (PSIS) to PSIS – clearly a turn to the right. This will be held, in our experience, primarily in the muscles and intermuscular septa of the adductors, with the compartment drawn back on the right and forward on the left.
Note that the two hips, when seen from above, are resting at markedly different states of rotation, with the left hip in lateral rotation and the right in medial rotation. This is not readily discernible in any of the horizontal views, so use this vertical view to assess rotations more easily.
If we compare the line between the two PSISs to the line along the bottom of the scapulae (represented in this photo essentially by the bra line), we can see that there is only a limited amount of additional right rotation between the pelvis and the lower thoracic spine. Above this line, however, the rotation goes to the left, strongly, such that by the top of the ribs the manubrium is essentially facing straight ahead, and only a small additional left rotation is required in the shoulders and neck to face fully forward.
While the right SL is clearly shorter than the left here, it is probably countering a short left psoas complex that is pulling the lumbars forward on the left, and pectineus on the right, which will be pulling the pubis to the right.
The state of the spine in terms of the idiopathic scoliosis is unknown without an X-ray or MRI, so we are limited to this soft-tissue analysis.
•
Palpate each psoas to feel a marked difference in fascial tone. Different portions of the psoas will assess short or relaxed in these rotoscoliotic patterns.
•
Precise palpation of the spinal muscles is required in every scoliotic pattern, as the soft-tissue patterns are highly individual.
Our final model (
Fig. 3.20
) demonstrates a common fascial pattern type that is not easily amenable to a myofascial meridian analysis, but is nevertheless commonly seen, useful to notice, and subject to manual correction.
While some of this pattern is similar to models above, the essential characteristic can be expressed as ‘cylinders’. The two legs are essentially two cylinders, which can easily be either medially rotated or laterally rotated. Carry that idea up into the trunk, as if the two cylinders carried up through the rib cage and shoulders on either side of the spine in the middle. You can see the two cylinders in the trunk are laterally rotated, while the hips and legs are overall medially rotated. Trouble often occurs where cylinders change orientation from medial to lateral.
If we imagine that this concept of cylinders extended up through the torso to the shoulders, one cylinder on either side of the spine, and further imagine that these cylinders can rotate on each other medially or laterally like the legs, we can arrive at a strategy for the pattern we see here.
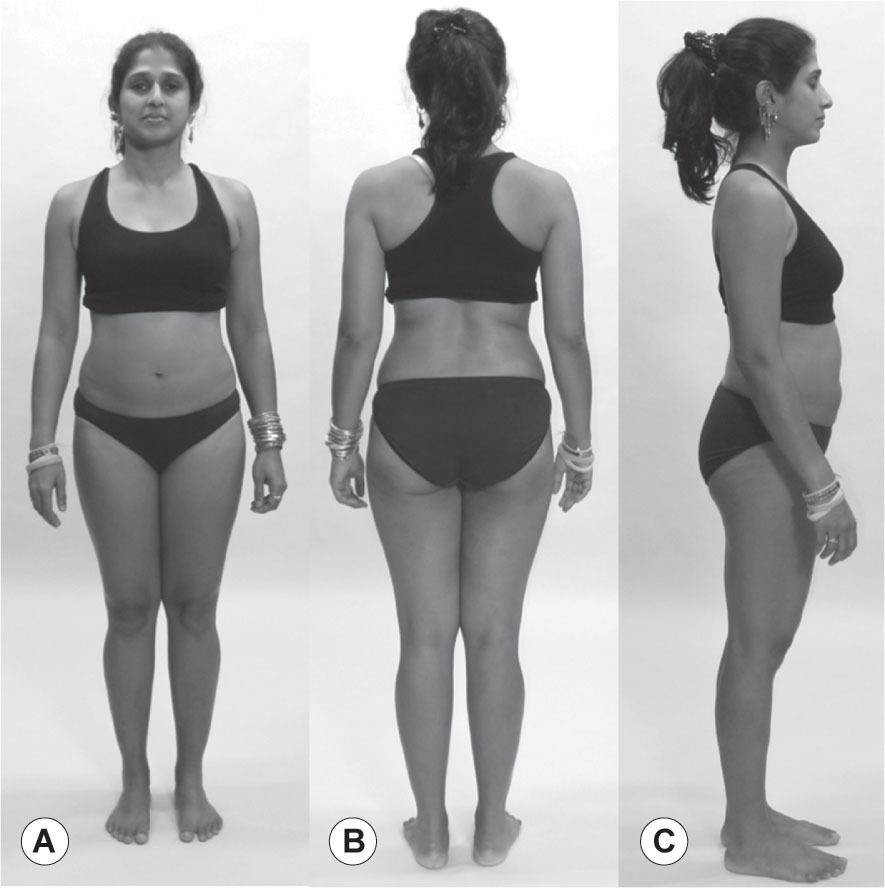
Her lower legs are more laterally rotated than her thighs, and the hips follow the medial rotation pattern: she looks narrower in the front across the pubic bone than she does across the ischia at the back of the pelvis. At the lumbar section, however, the situation reverses and the lumbars are narrow at the back and the belly is wide at the front. This pattern of the cylinders – open at the front, closed at the back – continues up her body through the shoulders. (Rarely, and only when the pattern has been strongly held for a long time, this pattern enters the cranium and can be seen or felt in the cranial respiration. Also rarely, and usually only in response to a more extreme trauma or scoliosis, one can find trunk cylinders where one side is rotated medially and the other laterally. This pattern involves significant strain and usually chronic pain in the system.)
Loading this structure significantly with weights or performance stress would not be a good plan. These patterns are especially vulnerable in the place where the medial and lateral rotations interface – in this case at the SI joints or L5–S1. When observed, this pattern calls for fascial opening across the closed parts of the cylinders before attempting a strengthening program.
In this case, this would mean widening the myofascial and fascial tissues along the front of the pelvis, meaning the attachments along the ischiopubic ramus and the pectineus on the iliopectineal ridge. The medial rotators of the hip – the tensor fasciae latae and the anterior gluteus medius and minimus – would also need attention. The deep lateral rotators are most probably ‘locked long’ in eccentric loading, and would thus require some cross-fiber work to help them reset their tone.
In the lumbars, the cylinders change to a lateral rotation. This takes the lumbars forward, and as the spinous processes move forward they take the fascial
planes with them, narrowing the tissue in the back and opening it in the front.
While the widening in the front in the belly will be more amenable to a training–toning–strengthening program, such as Pilates or physical therapy, the tendency of the fascial layers in the back to narrow as the lumbars move forward is best dealt with by strong and specific fascial stretching. Putting the client over a physioball to encourage widening is a good start, but is usually insufficient or requires a great deal of time.
More efficient is a protocol – whether over a ball, seated, or prone with a bolster supporting the belly, in order not to push the lumbars more anterior – of widening the thoracolumbar fascia away from the midline. Because of the number of layers of thoracolumbar fascia and fascially dense erector spinae and multifidus, and the long expanse of this cylinder rotation along her spine, we should not expect this to happen all in one session. Progressive work from layer to layer and section to section is often necessary, along with the educational/homework piece, to effect a sustained change of these ingrained patterns.
•
Palpate the anterior adductors – pectineus, brevis, and longus – to find the restraining guy-wires that hold the pelvis down.
•
Palpate along either side of the lumbar spine about 1” (2 cm) from the midline to find the thick and slick vertical fascial bands running with the erectors and the thoracolumbar fascia. Palpating this tissue will convince you of the necessity for sustained and effective work to change this pattern.
Conclusion
Numerous other examples in clinical application are offered in the book, Anatomy Trains
(Myers 2014), and its supporting video programs and courses.
Every therapist has seen shoulders drop away from the ears when the feet and legs are worked, low back pain melt away from work in the groin, or a client’s breathing open from work on the forearms. The Anatomy Trains map offers one way of understanding and managing these effects in terms of mechanical or energetic communication across our ‘sinew channels’ of the fascial connections.
Once the relationships within each line are understood, the interactions among the lines open new possibilities for resolving longstanding postural and movement patterns that will not yield to ‘single part’ attempts to remedy a problem. Progressive work with the lines can create dynamic shifts in these patterns, resulting in the reintroduction of ‘poise’ – an integral balance and length in body structure. As we move beyond ‘problems’ to performance enhancement, this same bodyreading process can be used to ‘tune’ myofascial balance toward a more resilient, responsive tensegrity where the whole body participates and accommodates to the required activity – athletic or artistic.
An additional ‘global’ assessment protocol, together with a number of local palpation approaches are detailed in
Chapter 4
– a companion to the exploration in this chapter of clinical methods of evaluation of global fascial function and dysfunction.
Next:
Global postural considerations are also the focus of
Chapters 16
(
Rolfing
®
structural integration
), and
17
(
Global Postural Re-education
).
References
Berthoz A 2000 The body sense of movement. Harvard University Press, Cambridge, MA
Fuller B 1975 Synergetics. Macmillan, New York, Ch 7
Heartney E 2009 Kenneth Snelson: forces made visible. Hudson Hills, Easthampton, MA
Hyman J, Rodeo SA 2000 Injury and repair of tendons and ligaments. Phys Med Rehabil Clin N Am 11(2):267–268
Juhan D 1987 Job’s body. Station Hill Press, Tarrytown, NY
Kjaer M et al 2009 From mechanical loading to collagen synthesis, structural changes and function in human tendon. Scand J Med Sci Sports 19(4):500–510
Koob A 2009 The root of thought. Pearson Education, Upper Saddle River, NJ
Moore K, Persaud T 1999 The developing human, 6th edn. WB Saunders, London
Myers T 2014 Anatomy Trains, 3rd edn. Churchill Livingstone Elsevier, Edinburgh
Pert C 1997 Molecules of emotion. Scribner, New York
Pirschinger A 2007 The extracellular matrix and ground regulation. North Atlantic, Berkeley
Purslow P 1994 The morphology and mechanical properties of endomysium in series-fibred muscles: variations with muscle length. J Muscle Res Cell Motil 15(3):299–308
Schleip R, Findley T, Chaitow L, Huijing P 2012 Fascia: the tensional network of the human body. Churchill Livingstone Elsevier, Edinburgh
Snyder G 1975 Fasciae: applied anatomy and physiology. Kirksville College of Osteopathy yearbook, Kirksville, MO
Turvey M, Fonseca S 2014 The medium of haptic perception: a tensegrity hypothesis. J Motor Behav 46(3):143–187
Verela F, Frenk S 1987 The organ of form. J Social Biological Structure 10:73–83
Williams PL 1995 Gray’s Anatomy, 38th edn. Churchill Livingstone, Edinburgh pp 75, 906