four
SCANNING FOR GODOT
ONE OF THE GREATEST ADVANCES IN RECENT MEDICAL technology has been the development of neuroimaging, the set of procedures that lets doctors and scientists scan the living contents of the skull without surgery. These procedures have had huge scientific impact, garnered two Nobel Prizes, and probably done more than any other aspect of contemporary neuroscience to shape popular conceptions of the brain. Judging from its prevalence and influence, neuroimaging seems to have a mystique all to itself. Over ten thousand medical research articles involving neuroimaging are now published each year. Brain scanning also figures in fields as far-flung as economics and the law. You have almost certainly seen brain images, perhaps spinning in 3-D to show you the location of a tumor, perhaps sparkling with colors that show you how the brain is affected by a task or treatment. If you have been to the hospital with any sort of neurological complaint, you have probably had an X-ray computed tomography (CT) scan or a magnetic resonance imaging (MRI) examination; the spinning brain or sparkling colors might have been yours. Neuroimaging is how many people get to know their brains.
The most exciting form of neuroimaging is so-called functional brain scanning—the use of imaging to measure the brain in action, rather than merely its structure. This is most commonly performed with a functional MRI or fMRI, a technique I have spent much of my own career working with. In the 1990s, fMRI emerged as the most powerful method for mapping human brain activity, and it has since become a mainstay of neuroscience programs everywhere. To perform an fMRI experiment, researchers obtain a movie-like series of brain scans of a person lying in an imaging machine for a period of time. They then analyze the image series for time-varying changes that correlate with whatever the subject was doing or experiencing. These changes indicate how different parts of the brain are involved in the subject’s behavior. Using fMRI, researchers have been able to find regions of the brain that process shapes, colors, smells, tastes, mistakes, actions, emotions, calculations, and much, much more. More edgy studies have sought to find brain areas whose activity depends on thinking like a lawyer or preferring Pepsi versus Coke. In the clinic, doctors and scientists apply fMRI and related methods to discern brain activity abnormalities associated with diseases like autism or schizophrenia. “The influence of fMRI-based research has changed our world,” says Bruce Rosen, one of the inventors of the technique.
Befitting its importance in science and medicine, functional imaging is one of the most visible facets of brain research in the media and popular culture. Hundreds of newspaper articles related to fMRI are published each year, turning imaging into a flagship of modern neurobiology. Readers get hooked with catchy headlines like “This Is Your Brain on Politics” or “Watching New Love As It Sears the Brain.” Particular attention goes to audacious suggestions that brain imaging can read minds, expose lies, and help marketers advertise their goods. Commenting on the public enthusiasm for such stories, psychiatrist Sally Satel and psychologist Scott Lilienfeld bemoan the fact that functional brain mapping has supplanted other valid ways to analyze mental and behavioral phenomena. At the same time, they admit that “it is easy to see why brain imaging would beguile almost anyone interested in pulling back the curtain on the mental lives of others.”
By offering us face-to-face encounters with living human brains, one might also imagine that functional neuroimaging could provide an antidote to the cerebral mystique. What better way to grapple with the biological reality behind our minds than by seeing our own brains at work inside our heads, with the aid of techniques like fMRI? In a much discussed 2008 article, psychologists David McCabe and Alan Castel proposed that brain images fascinate laypeople precisely because they “provide a physical basis for abstract cognitive processes.” But we will see in this chapter that the evidence in support of this view is sparse. Instead, brain imaging results are prone to contradictory interpretations that leave people free to choose among totally disparate concepts of the mind and brain. Even the most important and scientifically meaningful contribution of functional neuroimaging—the identification of brain areas specialized for distinct cognitive tasks—paradoxically reinforces the same kind of dualist perspectives we saw in the previous chapters. I will argue here that we must look well beyond the capabilities of today’s brain imaging technology for a truer appreciation of the brain’s place in human nature.
Let’s start by testing your reaction to some real functional brain imaging data. A pair of mottled gray-on-black ellipsoids ogle you from the page (see Figure 6). The one on the right is dull and bluish, but the one on the left twinkles with hot sparks of orange and yellow. If you had been born yesterday, you might see these forms as nothing more than embellished Rorschach blots, ready to accept whatever significance you ascribe to them. But today, trained by countless similar images in the media, you probably know that they are brain images.
As it happens, the left and right ovals represent functional brain imaging data from two separate sets of subjects. The dull brain on the right side is labeled “obese group.” The flashy one on the left is “nonobese.” Its glint of color, absent on the other one, denotes activation of a brain region called the prefrontal cortex. Below them sits a caption: “When viewing images of food, obese participants had reduced activation in brain areas associated with self-control.” The obvious implication of this vignette is that brain studies help explain how obese people react to food.
Can you believe these results? Do you find them intriguing, or perhaps surprising? Would you feel the same way if the caption was not paired with brain images? Do your beliefs about the mind or the soul influence your reactions? For instance, if you are religious, does your faith make you view the brain images or accompanying caption with lesser or greater skepticism?

FIGURE 6. A vignette including functional imaging data from Hook and Farah’s study of the effects of neuroimaging on beliefs. Areas of activation are denoted by thin dotted lines; positive = light gray, negative = dark gray. Adapted with permission from C. J. Hook and M. J. Farah, “Look again: Effects of brain images and mind-brain dualism on lay evaluations of research,” Journal of Cognitive Neuroscience 25 (2013): 1397–1405, © 2013 by the Massachusetts Institute of Technology.
Cognitive neuroscientists Cayce Hook and Martha Farah asked these questions in a large 2013 survey of how people respond to brain imaging. The researchers wanted in particular to test the McCabe and Castel hypothesis that brain imaging makes its splash by showing that mental processes have a physical realization. If the McCabe-Castel hypothesis were true, Hook and Farah reasoned, then people who believe in an incorporeal soul might tend to be more surprised and less accepting of neuroimaging results than others. Showing fMRI data to these dualists could be like introducing space aliens to someone who denies the possibility of extraterrestrial life. On the other hand, people who believe the mind is completely material should be unfazed by functional brain imaging. These people—often referred to as physicalists—probably already think the brain is where the magic happens and would not be surprised by more news of a relationship between brain and behavior.
Remarkably, Hook and Farah found that dualists and physicalists reacted similarly to the functional brain imaging data they saw. Vignettes that provided evidence for a physical manifestation of the mind in the brain did not appear to surprise or interest dualists more than physicalists, or vice versa. Moreover, the inclusion of actual brain images in the vignettes did not make much difference to how the participants responded, other than piquing both groups’ interest a little more. “Across 988 participants,” the researchers concluded, “we found little evidence of neuroimaging’s… relation to self-professed dualistic beliefs.” If brain imaging provides evidence of a biological basis for the mind, shouldn’t it be otherwise?
It turns out that far from being shocked or disbelieving of neuroimaging data, some committed dualists actually embrace functional brain imaging as a tool for studying the disembodied mind they believe in. The Dalai Lama is one. Over the past decade, the spiritual leader of Tibetan Buddhism has worked with cognitive scientist Richard Davidson at the University of Wisconsin to organize a set of experiments in which the brains of Tibetan monks are scanned while they meditate. According to Buddhist dogma, meditation is one of eight steps along the spiritual path to attaining nirvana, a permanent escape from the endless cycle of birth, death, and reincarnation. But Davidson and his colleagues found plainly physical differences between the brain activation patterns detected from monks versus novices while they meditated. The results suggested that the monks’ Buddhist exercises correlate with how their brains behave during meditation, and also seem quite compatible with the Dalai Lama’s own dualist take. While physicalists might say that Davidson’s brain imaging results show something about how brain activity underlies the act of meditation, and the mind more generally, the Dalai Lama simply turns this scenario on its head. He says he is interested “in the extent to which the mind itself, and specific subtle thoughts, may have an influence upon the brain.”
Davidson’s work with the meditating Tibetans is part of a research area sometimes called neurotheology, in which the neuroimaging and associated techniques are applied to analyze brain function during a variety of spiritual and religious activities. The very existence of this field depends on the compatibility between functional brain imaging and religion. Neurotheology laboratories have compared brain activity in religious believers and nonbelievers as they reason, moralize, or pray. These studies are sustained by cohorts of religious volunteers who presumably find that the experiments do little to challenge their traditional concepts of the soul. Andrew Newberg of the University of Pennsylvania has been a major promoter of neurotheology and has gained recognition for some of his functional neuroimaging work. In one study, Newberg’s team recruited a group of devout Charismatic and Pentecostal Christians to engage in the euphoric practice of glossolalia (speaking in tongues) while being scanned. “I don’t think faith has anything to be afraid of from science,” explained pastor Gerry Stoltzfoos, who participated in the experiments. Stoltzfoos’s attitude is very much like the Dalai Lama’s: “Science validates faith,” he insists.
A leading neurotheologist named Mario Beauregard goes beyond just studying spiritual phenomena in the brain. He has authored several books advocating nonmaterialist views of the mind, while at the same time using brain imaging to document neural correlates of mystical experiences. A 2007 Scientific American article characterizes Beauregard’s work as “Searching for God in the Brain.” Like the Dalai Lama, Beauregard fully accepts the brain’s importance but views the brain as a servant to the disembodied mind. “A wealth of scientific studies,” he argues, “indicate that our thoughts, beliefs, and emotions influence what is happening in our brains.” In Beauregard’s eyes, the fMRI machine is a tool for detecting spiritual influences on the matter of the brain, rather than for explaining the spirit itself in terms of matter.
Examples from the Hook and Farah survey to Beauregard’s work show how easily functional brain imaging studies can be reconciled with supernatural beliefs about the mind or soul. It seems that if we use fMRI to pull back the curtain on our mental lives, we can see more or less what we want to. Although some people think the brain gives rise to the mind, and others think the mind controls the brain, nobody is surprised by the fact that the brain is involved. Even René Descartes, the man whose name became synonymous with dualism, posited that the spirit interacts with the body through a tiny brain structure called the pineal gland. Neuroimaging has not provided the kind of information that would rule in or rule out such mind-brain interactions, and it therefore cannot serve as a basis for distinguishing among dualist or physicalist worldviews. To figure out why this is so, let us pull back the curtain on brain imaging itself, and take a closer look at the kinds of knowledge it actually produces.
Modern brain imaging was born in a drab hospital room in Wimbledon, near the site of the famous tennis tournaments. On October 1, 1971, a middleaged woman lay supine on an elevated stretcher, knees raised. Her head disappeared into a chunky square block, about a meter per side and twenty-five centimeters thick, held up edgewise by a heavy gantry. A cylindrical capsule attached to one edge of the square ran smoothly from corner to corner, like the lure at a dog track. After the capsule’s run, the square rotated with a quick jerk about the patient’s head. The capsule ran and the square rotated again and again, continuing rhythmically back and forth like cumbersome clockwork. After five minutes, the square had made half a revolution around the woman’s head. In a room next door, stuffed with space-age electronic instruments, a picture flickered onto a computer screen: a white oval on a black background. The oval’s dark, faintly textured core was bisected by a hazy light streak, but to one side a small black patch rudely disrupted the symmetry. The picture looked like a Miró painting, but it was actually the first clinical brain scan, obtained by Godfrey Hounsfield and his collaborators using their prototype X-ray CT scanner. The lady in the portrait had a brain tumor—the black patch in the oval. It was later successfully operated on, a coup brilliantly enabled by the newfangled neuroimaging know-how.
Although this experiment took place almost half a century ago, it debuted features of neuroimaging that persist to this day, such as the scanner configuration and the alliance of imaging hardware with computer processing. The basic idea behind CT is to measure X-ray transmission from every possible angle and position through a subject; in Hounsfield’s setup this was accomplished with the dog lure and the rotating square, which contained an X-ray source and detector. A mathematical algorithm is then applied to reconstruct the image. CT images are static, however. In some cases, CT can help explain a cognitive problem by discovering the location of disruption, but it cannot indicate what the brain is doing during the scan.
The first brain scans to look dynamically at the brain’s biological processes were methods sensitive to radioactive tracers (radiotracers for short). These substances are close analogs of natural biological or pharmacological molecules; when injected or ingested into the body, they go to the same places and do the same things as their nonradioactive counterparts. Radiotracers also emit gamma photons, which pass easily through biological tissue. Because this radiation can be detected noninvasively even with extraordinarily low tracer doses, there is minimal risk of side effects. Tracers called positron emitters yield two gamma photons simultaneously, enabling particularly sensitive and spatially accurate detection. Imaging these molecules in three dimensions became possible using the positron emission tomography (PET) scanner introduced in 1975 by Michel Ter-Pogossian, Michael Phelps, and their coworkers at Washington University in St. Louis.
PET imaging quickly formed the basis of several strategies for mapping aspects of brain function. In one approach, a positron-emitting version of the blood sugar glucose—the body’s main energy source—is used to image cerebral metabolism. The radioactive agent 18F-fluorodeoxyglucose (FDG) accumulates in proportion to the brain’s glucose utilization. The buildup of FDG radioactivity can be monitored to see which areas of the brain are most active, at least according to their level of “fuel consumption.” A second PET functional imaging method uses blood-borne radiotracers such as 15O-water or 13N-ammonia to measure changes in cerebral blood flow. Blood flow increases are evoked by neural activity and thereby result in greater delivery of the tracer to activated brain regions. Blood flow changes are harder to interpret in terms of neural mechanisms, but they take place faster than measurable changes in metabolic rate. In a further suite of applications, specific neurochemical processes are studied using radiotracers designed to interact with enzymes or receptors involved in those processes.
Many of the original PET brain imaging techniques are still in common use, and they have been joined by methods that use new tracers. A recent breakthrough, for instance, has been the development by the University of Pittsburgh’s William Klunk and others of PET tracers that reveal the pathology of Alzheimer’s disease. PET has proved limited for many studies of brain activity, however. One problem is that PET scans offer relatively coarse-grained spatial detail, or resolution. Pixel sizes of several millimeters are typical, meaning that every point in a PET scan corresponds to tens of thousands of cells and sometimes even more than one brain region. More importantly, PET scans are ponderously slow compared with brain processes like perceiving and thinking. Even the fastest functional PET experiments require scan times of about a minute—almost a thousand times longer than it takes to recognize a person’s face, and about five times longer than it took world champion Magnus Carlsen to defeat Bill Gates in an entire game of speed chess.
Several of the weaknesses of PET were addressed by a radically different imaging technology developed by Paul Lauterbur at the State University of New York in 1973. Lauterbur was a chemist who specialized in an analytical technique called nuclear magnetic resonance (NMR) spectroscopy. NMR is an effect whereby the nuclei of certain atoms—most commonly the hydrogen atoms in water—absorb radio waves at specific transmission frequencies when placed in a strong magnetic field. Lauterbur discovered a way to use NMR to reveal the spatial positions of absorbing nuclei. Because biological tissues are largely transparent to radio waves (and undamaged by them), this new NMR-based imaging was perfect for visualizing living soft tissue in three dimensions. As NMR imaging gained steam in the medical community, the threatening N for nuclear was lost, and the method became best known as MRI. MRI was quickly recognized for its superior rendering of anatomical detail in soft tissues like the brain.
In the early 1990s, scientists discovered ways to perform functional brain imaging using MRI. In the first published fMRI study, Jack Belliveau, Bruce Rosen, and their collaborators at the Massachusetts General Hospital in Boston emulated earlier PET experiments by injecting an MRI contrast agent into the bloodstream of volunteer subjects while they were being scanned. The scientists were then able to map brain activity by seeing where the contrast agent accumulated during visual stimulation. At about the same time, another group of researchers led by Seiji Ogawa at Bell Labs showed that blood itself can act as an intrinsic contrast agent for fMRI. Because of the weak magnetism of both oxygen and iron in blood, small changes in blood flow and oxygenation can be detected without the need for injections. Such effects occur within seconds of elevated brain activity and are the basis of most current functional imaging experiments.
Not surprisingly, limitations of fMRI arise from its dependence on blood. The spatial resolution of fMRI is fundamentally limited by the spacing between blood vessels in the brain. This is somewhere around a tenth of a millimeter, much larger than brain cell sizes. Most fMRI signals are likely to arise from contributions involving many different types of neurons and glia, as well as possible changes in blood circulation unrelated to local brain activity. The hundreds of chemical messengers we considered in Chapter 2, as well as the synapses and connectivity of Chapter 3, are all lost in translation. Berkeley neuroimager Jack Gallant comments that “fMRI is like measuring the total electricity usage in your office at specific times to figure out what’s going on at everyone’s desk.” Researchers also bemoan the sluggishness of fMRI compared with neural activity. Imagine watching a movie that has been blurred in time over several seconds per frame. Our favorite action heroes—Rocky and Ivan, Rosa and James, Obi-wan and Vader—would all be reduced to uninterpretable streaks of color. This is what blood flow effects detected by fMRI are like. For this reason, scientists sometimes complement fMRI with measurements from faster recording techniques called electroencephalography (EEG) and magnetoencephalography (MEG). But although MEG and EEG respond quickly to the brain’s electromagnetic activity, they cannot localize the activity nearly as precisely or reliably as fMRI.
The signals detected by fMRI and other functional imaging approaches are also tiny—brain activity typically elicits ripples of at most a few percent in image intensity. These subtle changes play against a background of fluctuations resulting from the subject’s movements, instability of the scanning equipment, and physiological processes unrelated to the study. Scientists must try hard to separate out image changes that are genuinely related to whatever stimulus or phenomenon they are trying to learn about. This usually involves expansive computational analysis of dozens of repeated trials, multiple subjects, and varied experimental conditions. The results of such calculations are usually depicted as colored blobs of inferred brain activity, superimposed on black and white anatomical images as in Figure 6. Such pictures provide the best information we can currently obtain about human brain function, but they do not really show what the brain is doing at any point in time, and they rarely come from anybody’s brain in particular. Instead, functional brain maps are highly processed, statistical aggregations of image data that are sometimes as distant from underlying biological processes as bologna is from a pig.
The sophisticated computational tricks used in analyzing neuroimaging data come with equally sophisticated pitfalls. A University of California, Santa Barbara, postdoc named Craig Bennett shockingly illustrated this point by using apparently innocent fMRI methods to reveal brain activation in a dead salmon. Bennett and his collaborators scanned the late fish while it “saw” a series of photographs. A few pixels in the brain showed what looked like responses to the photos based on statistical testing, resulting in a very fishy version of a typical fMRI activation map. In reality, the apparent brain responses resulted from random fluctuations in the images, which were ineffectively screened out by commonplace analysis methods. Bennett had a difficult time publishing his satirical study but was aptly awarded an Ig Nobel prize, given to “honor achievements that first make people laugh, and then make them think.” A second damning study, led by an MIT student named Ed Vul, revealed that high-profile brain imaging papers were frequently reporting statistically impossible results, the equivalent of claiming odds of better than 50-50 for a coin toss. These mistakes led authors of the offending studies to report correlations between brain areas and complex stimuli that were literally too good to be true. Although the types of errors that Bennett, Vul, and their colleagues exposed are neither inherent nor specific to brain imaging research, imaging studies are particularly prone to such problems because of the small signals and large datasets involved.
The indirectness and coarse resolution of current brain imaging techniques clearly provide wide latitude for disparate influences and interpretations. Examples like the salmon study also show us how easy it is to bias results simply by carelessly designing or analyzing experiments. We have seen that different people can look at imaging data and see the mind at work on the brain or the brain busy carrying out functions of the mind—brain images make as much sense to dualists as to physicalists. After learning about what functional neuroimaging really tells us, we can begin to understand this contradiction. Today’s brain activity maps are so indistinct that we can imagine almost anything going on behind the scenes.
The brain is like a Swiss Army knife, argues my MIT colleague Nancy Kanwisher, a pioneer in applying fMRI to problems in cognitive science. Despite limitations of the technique, imaging studies including hers have delineated a once unanticipated array of distinct brain regions that respond during specialized tasks, from recognizing faces to thinking about thinking (see Figure 7). Each of these brain areas seems to be specialized for its task, like different tools on the knife. Almost half of published neuroimaging studies are localization studies, and many of the rest deal with further characterization of defined brain regions. Localization results are the most obvious take-home lessons from functional neuroimaging research. When carefully performed and interpreted, they inform us about how the brain and mind are constructed, but when regarded superficially, the localization of cognitive functions can be a distraction from efforts to understand how our brains and minds really work.
Hard evidence for the specialization of brain regions has been around for a long time. Before PET and fMRI, data came largely from small numbers of neurological patients in whom specific cognitive or behavioral defects could be traced to focal brain injuries. Perhaps the best-known example was the case of a patient named Louis Leborgne, who was studied by the French physician Paul Broca in 1861. Leborgne had suffered from epilepsy since childhood, and as an adult lost his ability to speak; he was hospitalized and could utter only one syllable, “tan.” Despite his speech loss, Leborgne’s comprehension and general cognitive capabilities were unharmed—a set of conditions now known as Broca’s aphasia. In a postmortem brain examination, Broca found that Leborgne had a lesion in the left frontal lobe of his cerebral cortex, and he went on to discover injuries at the same location in patients with similar language defects. Discovery of the correspondence between language production and the brain region called Broca’s area represented a striking validation of Franz Gall’s theory of functional localization (see Chapter 1). The underlying idea behind phrenology was at least partially correct, even if Gall’s maps of specific regions and corresponding skull features were entirely wrongheaded.
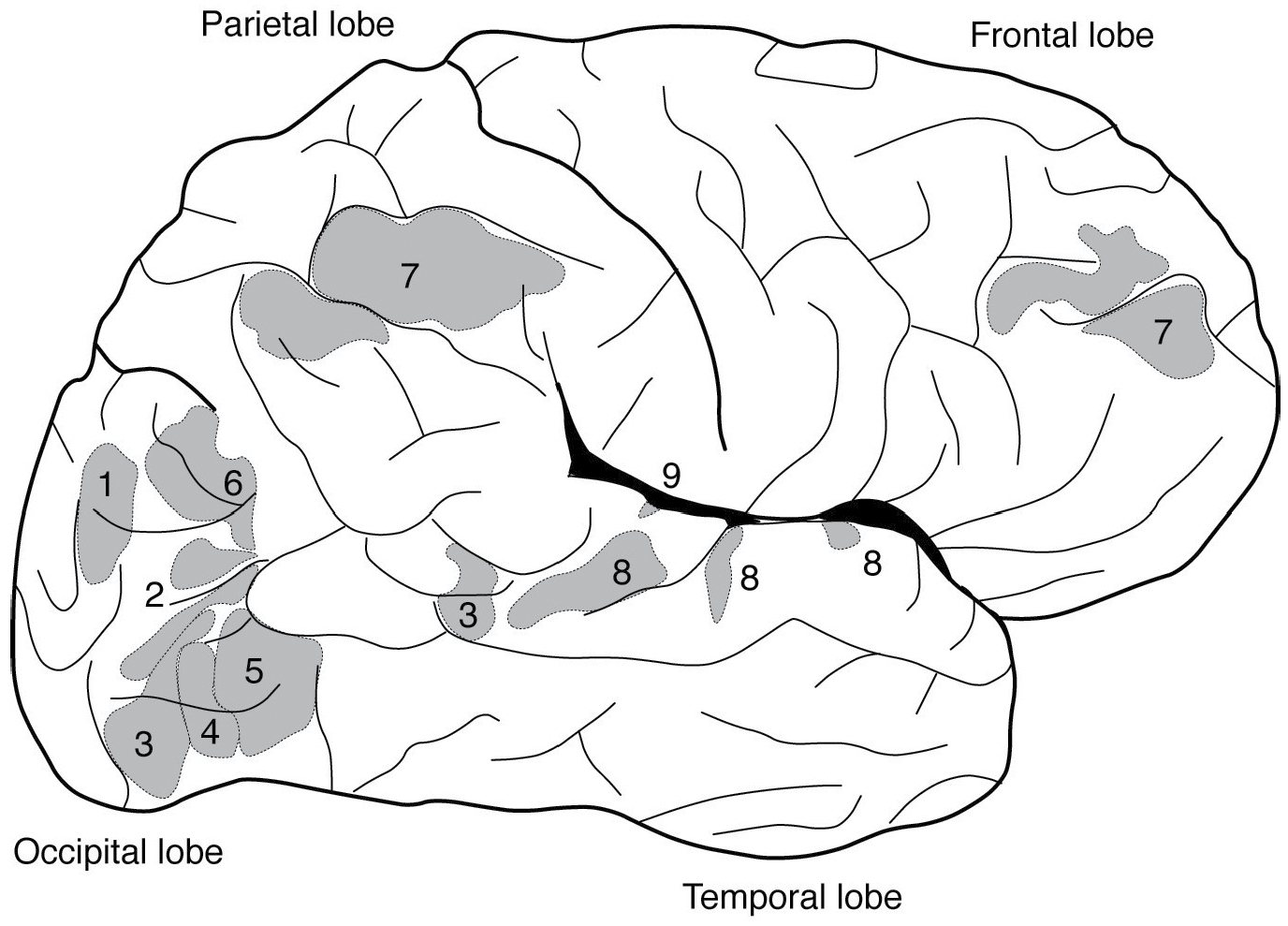
FIGURE 7. Human cerebral cortex showing lobes and areas that according to neuroimaging studies respond specifically to (1) places, (2) body parts, (3) faces, (4) faces and motion, (5) motion alone, (6) thinking about other people’s thinking, (7) difficult cognitive tasks, (8) speech sounds, and (9) sounds with pitch.
Neuroimaging makes the same point but sharpens it. Using today’s techniques, observations are uncoupled from the rare conjunction of serendipity and personal misfortune that led to breakthroughs like Broca’s. Willing volunteers in generous numbers can be scanned during many different stimuli or tasks in single or multiple sittings. Researchers can see the results almost immediately after each experiment, rather than waiting for their subjects to die and undergo autopsy. The brains of healthy neuroimaging subjects are not distorted by damage or disease, as lesioned cerebra can be. Brain imaging results therefore usually reflect normal physiology. Most importantly, imaging, unlike injury, probes the entire brain at once. PET or fMRI can reveal if multiple structures are engaged in experimental paradigms and can characterize the extent and magnitude of responses in each region. The roles of structures involved in language, for instance, can all be examined in a single experiment: Broca’s area, important for articulation; Wernicke’s area, important for comprehension; auditory and motor cortices, important for generic aspects of hearing and movement; and numerous functionally important subdivisions of each of these regions are all involved and can be seen working in parallel during a speech task.
Discovery of specialized brain regions has indubitable biological significance. Just as forces at work throughout the earth’s geological evolution gave rise to the mountain ranges, oceans, and rivers we see today, so we imagine that the factors that shaped human evolution patterned our brains in the ways we can now detect. The presence of brain areas or groups of areas whose activities correlate strongly with mental functions like speech and socializing suggests in particular that these functions are distinct adaptations with dedicated neural hardware, an interpretation now shared by many neuroscientists. “What’s important… is not the particular locations of [the corresponding] brain regions,” Kanwisher explains, “but the simple fact that we have selective, specific components of mind and brain in the first place.”
But the importance neuroimaging studies place on associating mental faculties with physical spots in the brain has also led many to disparage it as a reincarnation of phrenological pseudoscience. “Critics feel that fMRI overlooks the networked or distributed nature of the brain’s workings, emphasizing localized activity when it is the communication among regions that is most critical to mental function,” writes David Dobbs in a Scientific American piece entitled “Fact or Phrenology?” Psychologist Russell Poldrack went so far as to compile a list of published fMRI studies that implicitly lend support to phrenological mental categories, illustrating the surprising compatibility between modern science and outmoded ideas. For each example, Poldrack related old phrenological classifications to thematically similar fMRI experiments, and then to the specific brain regions the imaging studies had identified. “One can be almost certain that Gall and his contemporaries would have taken these neuroimaging results as evidence for the biological reality of his proposed faculties,” Poldrack comments. The way neuroimaging results are reported often strengthens Poldrack’s point. Titles such as “Neural Correlates of Giving Support to a Loved One,” “The Neural Substrate of Human Empathy,” or “Neural Correlates of Superior Intelligence” give the impression that complex character traits can be boiled down to blotches of territory in the brain. It is easy to imagine some of the neural correlates and substrates—each corresponding to one or more localized fMRI activations—nestling alongside phrenological regions for attributes like “amativeness” and “acquisitiveness” on one of Lorenzo Fowler’s ceramic heads.
Careless interpretation of brain localization results invites further criticism by implying that regional activity, like labeled bumps on the head, actually stands for particular cognitive processes. Advertising expert Martin Lindstrom claimed in a 2011 New York Times op-ed that people literally love their Apple iPhones because photographs of iPhones activate a brain region called the insular cortex, which is also among brain regions that respond when people view pictures of their romantic partners. Here Lindstrom treats insular cortex fMRI signals as denoting love, even though the truth is that this region responds during both negative and positive emotions. In his book Imagine, author Jonah Lehrer summarizes an experiment that found an association between problem solving and a brain area called the anterior superior temporal gyrus (aSTG). He writes that “the aSTG is able to discover” the answers to word puzzles—the aSTG itself becomes a problem solver. Even the Nobel Prize–winning biologist Francis Crick appears to slip into the same “brain area equals cognitive function” fallacy when he references lesion studies to argue that “Free Will is located in or near the anterior cingulate sulcus,” a small fold near the midline of the brain.
This type of thinking is mistaken on both technical and theoretical grounds. The technical critique takes off from the limitations of brain imaging itself. Each blotch of activation represents thousands if not millions of cells, synapses, and neurochemicals, all contributing together to brain function like myriad voices engaged in a chaotic debate. Neuroimagers currently have no good way to parse or distinguish among the voices, so instead they tend to do the easiest thing: they listen to the loudest opinions! The loudest opinions could arise from unanimous stimulus responses of cells in a brain region, but more likely the viewpoints represent a majority, a plurality, or even just a vocal minority of cells overshadowing a silent majority. As far as fMRI and PET are concerned, the loudest voices are also those that produce the biggest blood flow changes, not necessarily the voices most important for actual brain function. A further complication is that brain activation maps are almost always determined by comparing imaging responses to a test condition with responses under one or more reference conditions, so that the brain regions identified are actually those that display greater activity under the test condition, rather than regions that are turned on only by the test. The upshot is that the fact that a particular brain region “lights up” most during a particular mental task does not mean that the region as a whole specializes in that task to the exclusion of other functions.
On the flip side, the regions that do light up in imaging experiments rarely include all of the brain areas involved in any given cognitive process. A key reason for this is the “tip of the iceberg” problem. Icebergs are known for being much larger than the parts you can see; 90 percent of each floating mass hides menacingly under the ocean’s surface. In functional imaging analysis, the analog of the iceberg is a map of fMRI signal changes correlated with whatever task or stimulus was used in the experiment. The map is directly computed from the raw images. But although the map in principle covers the whole brain, the only parts you see as activation are areas where the reliability or magnitude of the imaging signals exceeds a cutoff value set by the experimenter. If the cutoff is set too low, too many peaks become visible, and the likelihood is high that some are caused by random nonneural fluctuations, like false activation in a dead salmon’s brain. For typical conservative cutoff values, however, some of what remains hidden beneath the cutoff—the submerged part of the iceberg—is still due to task-specific brain function. This genuine brain activation is lost in the data analysis and usually never discussed. Because of this problem, most functional imaging studies systematically overstate the degree to which brain responses are localized to a few small areas.
James Haxby of Dartmouth College has argued that the whole iceberg, including imaging signals that are usually ignored, should be considered when interpreting brain scanning experiments. In an influential 2001 paper, Haxby and his colleagues strayed from the standard practice of focusing only on brain areas with the largest responses to experimental stimuli. By doing so, they were able to observe that neural responses reach across “a wide expanse of cortex in which both large- and small-amplitude responses carry information” about visual stimuli. This type of approach favors a picture of brain activity in which mental processes are distributed over much of the brain, rather than being compartmentalized in specific structures.
At a fundamental level, all neuroscientists know that a picture like this must be accurate. Even if a particular patch of brain displays highly specialized activity patterns, the activity has to come from somewhere. If a brain region is activated by faces, for instance, face stimuli have to percolate from the retina through multiple levels of the brain’s visual system to generate signals in the brain area that responds most prominently to faces. If there were nothing face-like about the responses in other parts of the brain, then the face area would have no way to distinguish a face stimulus from any other. It could even be the case that negative fMRI signals outside the main face area—likely indicating reduced neural activity—help distinguish between faces and other stimuli in the brain. Such negative responses would be a bit like Arthur Conan Doyle’s famous guard dog who didn’t bark at an intruder in the night, revealing to Sherlock Holmes that the dog knew the intruder. But the variegated patterns of brain activity that might display such characteristics are necessarily overlooked by imaging analyses that focus only on peaks of brain activation and maximal responses.
This gets at the more theoretical problem with analyses that emphasize the location of cognitive processes: they tend to black-box the question of how the processes are actually performed. “Even if we could associate precisely defined cognitive functions in particular areas of the brain… it would tell us very little if anything about how the brain computes, represents, encodes, or instantiates psychological processes,” writes psychologist William Uttal in his 2003 book The New Phrenology. In a similar vein, philosopher Daniel Dennett derides the idea that specific brain regions could account for one particular cognitive process: the phenomenon of human consciousness. Localizing the process merely recasts the problem of deciphering how the brain functions to deciphering how the particular brain region in question functions. Dennett sarcastically analogizes this approach to a theatrical performance in which the brain’s embodiment of consciousness gets to “watch” all the mind’s happenings and be conscious of them, a scenario reminiscent again of Descartes’s mind-body dualism, even if it all takes place in the brain. Dennett regards this Cartesian theater as absurd because the brain region or regions that implement consciousness are as inscrutable as consciousness itself was to begin with, and the boundary between parts of the brain that are conscious and parts that aren’t can be nothing but arbitrary.
When we emphasize the localization of other cognitive functions, we are setting the stage for analogously absurd theaters, in which the brain regions that perceive colors, sentences, regions of space, and so on become unnaturally empowered and separated from the parts that don’t. Although no neuroscientist would endorse this burlesque view, it is exactly this impression that we get from simplistic portrayals of functional neuroimaging results. To many researchers, the main value of localization studies is not that they identify regions of the brain with apparently discrete functions, but that they provide clues as to how to begin more incisive experimental investigations, often using invasive experimental methods that directly probe cellular activity in animals.
Dennett’s Cartesian theater analogy and its extension to other cognitive processes evokes the real-life theater of modernist playwrights like Samuel Beckett. In Beckett’s absurdist masterpiece Waiting for Godot, the two vagabonds Vladimir and Estragon loiter by the side of the same road each day, hoping that they will eventually encounter the eponymous Godot, who never arrives. “You’re sure it was here?” asks Estragon, wondering if they are even looking in the right place for their man. Befitting a play that is often taken as a commentary about the meaninglessness of existence, Beckett and his critics never agreed on a single interpretation of the drama or characters. Like a colored blob of brain activation in an fMRI map, Godot in particular remains a mystery; we never learn who he is, what he represents, or whether he even exists. When we try to localize cognitive functions using brain imaging techniques, are we perhaps scanning for Godot? Are we searching for an enigma sometimes defined more by our expectations than by reality, with little promise of enlightenment at the end of our wait?
Contemporary neuroimaging strengthens the mystique of the brain with a combination of scientific luster, media hype, simple but sometimes simplistic findings, and compatibility with a wide variety of belief systems. With techniques like fMRI, we can learn interesting facts about brain activity without any pressure to revise our deeply held attitudes. Those who hope that cognition can be demystified using today’s human brain imaging technology are out of luck, however. Even when allied with the most sophisticated analysis methods, functional neuroimaging simply does not have the resolution or specificity to figure out what brain activation patterns really mean, how they are established, or how they are connected to the rest of the brain. “Claims that computational methods and non-invasive neuroimaging… should be sufficient to understand brain function and disorders are… naive and utterly incorrect,” writes neuroscientist and fMRI expert Nikos Logothetis.
Functional neuroimaging results today are somewhat like cartography before the days of authoritative atlases, settled borders, and satellite imagery. Like fMRI-based brain maps, ancient physical maps are often wonky, inaccurate depictions, limited by the technology available at the time they were made. Early cartographers found space for monsters as well as for the landmasses we know today, while modern brain interpreters find areas for free will as well as face perception. Some apparently specialized brain regions will stand the tests of further exploration; for instance, the existence of dedicated areas involved in face recognition has been backed up by electrode recordings, lesions, and stimulation studies in people and monkeys. But some regions, and even the cognitive concepts that define them, may prove as ephemeral as the lost lands of Thule and Atlantis. Whether validated or not, the association of mental functions with circumscribed places in the brain by its very nature will continue to foster a neurosegregation that keeps the biological bases of our mental processes bounded, separated from each other and the rest of the world. If we want to explain and understand at a deep level how mental functions work, we must therefore look beyond today’s human neuroimaging techniques.
Imagine a brain imaging method in which no cell’s activity, no pathway or connection, and no swirl of neurochemicals go undetected. With each touch, sound, or glimmer of light, a cascade of brain events takes place, each one open to the all-seeing eyes of our “total neuroimaging” technique. This is not a distant fantasy—it is at least approximate reality in neuroscience laboratories that study small transparent organisms. By combining cutting-edge optical microscopy techniques with fluorescent biochemical indicators of neural activity, researchers like Misha Ahrens of the Howard Hughes Medical Institute have already been able to record signals in almost every neuron of a baby zebrafish brain simultaneously. In experiments like his, there is vastly less room for uncertainty about cause-effect relationships and the organization of neural activity than there is in human brain imaging. Some scientists suggest that it may one day be possible to adapt techniques analogous to Ahrens’s for use in humans. Some of my own laboratory’s research aims, for instance, to create biochemical neural activity indicators detectable by fMRI; these could be a step toward total neuroimaging by allowing chemical and cellular-level signals to be detected noninvasively.
Total neuroimaging is the kind of technology that in the future could dramatically accelerate our ability to learn how the brain functions as an integrated, multifunctional organ. Although we are a long way off from having methods like this for studying people, progress is being made. Even this would not enable us to understand how mental processes really work, however. Researchers have been using high-resolution whole-brain optical imaging in worms and zebrafish for some time now, but as we saw in the last chapter, supposedly comprehensive information from simple nervous systems still falls short of explaining behavior. One of the reasons for this is that the brain and nervous system do not perform cognition on their own. Just as the brain regions discussed in this chapter cannot be viewed as acting in isolation, so the brain as a whole cannot be considered in isolation either; it must be viewed in the context of the body and the environment. The next chapters will further explore the continuum that unites the brain with its surroundings.