BEHAVIOR, COGNITION & INTELLIGENCE

DECISION-MAKING: BIG BRAINS ENABLE COMPLEX BEHAVIORS
CEPHALOPODS ARE RENOWNED FOR BEING “the smartest invertebrate animals” but how do we assess this in such an alien body form and then substantiate it with scientific evidence? One way to judge cognitive adaptability is to test and measure a cephalopod’s capability for rapid and diverse decision-making in a range of complex real-life scenarios. A large brain can facilitate these processes. Unlike most other animals, cephalopods augment much of their behavior and decision-making by changing their appearance—often very dramatically—for defense, feeding, and reproduction. Thus “rapid adaptive coloration” for camouflage and communication is considered a hallmark of soft-bodied cephalopods.
Decision-making is complex: it depends upon the integration of multiple sensory inputs, and cephalopods are well endowed with vision, taste, touch, smell, and hearing. Then a course of action must be determined, and here is where cephalopods excel compared with other invertebrate animals: their agility, speed, and color-changing abilities provide them with multiple options for action. Their huge brain, with 34 lobes and millions of tiny neurons, enables an enormous repertoire of behaviors for all occasions.

Octopus vulgaris is extremely alert all the time as it forages amidst complex environments that have many predators.
FAST LEARNERS
There has been a great deal of laboratory experimentation on visual discrimination learning in Octopus vulgaris. Most of this occurred in the 1950s and 1960s at the Stazione Zoologica in Naples, Italy. Octopuses can accomplish associative learning of some tasks in just a few trials. Octopuses can learn more than one visual discrimination at the same time. Octopus touch learning (via the suckers in their eight arms) is also rapid. Their ability to solve tasks such as serial reversal learning may relate to a flexible feeding strategy (“win–stay but lose–shift”) that depends upon learning and might be useful to a predator searching for patchily distributed prey. There are equivocal results from testing observational learning from conspecifics in octopuses and cuttlefishes, although most authors estimate that octopuses may be able to accomplish some sort of improvement in performance by watching other animals, such as their prey, in the wild. These types of experiments remain to be undertaken. In addition, both short-term (one hour) and long-term (several months) memory are well documented in octopuses and cuttlefishes. Recent studies have provided good evidence for episodic-like memory in cuttlefishes, which were required to recall the “what, where, and when” components of a foraging and feeding event. This is the first behavioral evidence of such complex memory in an invertebrate.
SPATIAL MEMORY OF OCTOPUSES & CUTTLEFISHES
Laboratory and field experiments show clearly that octopuses can learn from spatial cues in their environment. Two impressive field studies—one in the Atlantic and one in the Pacific—show that octopuses can forage many feet during several hours amidst complex habitats such as coral reefs and find their way back to their dens. Sometimes the octopuses even terminate their foraging at a distance and swim on a “bee line” back to their den, indicating a refined sort of visual map that so far defies explanation. In a Pacific study of Octopus cyanea, it was particularly noteworthy that individual octopuses were foraging twice a day, covering an average of 330 ft (100 m) per day, while traversing four distinctly different coral reef habitat types and not foraging in the same areas on successive trips or on successive days. Their navigational skills and apparent spatial memory seem exceptional.
Cuttlefishes in the laboratory demonstrate an ability for spatial learning when tested in a variety of mazes in which escape was the reinforcing “reward.” In some cases such as a wall maze or alley maze they learned within a few trials. However, in T mazes or open-field mazes, they did not learn at all. Cuttlefishes do not depend on home shelters the way that octopuses do, so their needs for spatial learning will be different (and yet to be determined) as these laboratory experiments showed.

Octopus cyanea in a forward swimming posture as it moves across a Pacific coral reef.

Octopus cyanea crawling with stealth across a coral reef.
RAPID ADAPTIVE COLORATION
PERHAPS THE MOST REMARKABLE FEATURE OF cephalopods is their complex and beautiful skin. Many species can radically change their appearance within the time it takes to blink your eye (about 200 milliseconds). This instantaneous change in appearance is used for camouflage as well as communication. These remarkable transformations are possible because brain centers send signals to nerves that run throughout the skin to a variety of pigmented chromatophore organs and reflective cells called iridophores. Many cephalopods control coloration by producing bioluminescence in the dark.
OPTICAL MAGIC: DYNAMIC SKIN PATTERNING
Chromatophores are like tiny elastic bags of pigment surrounded by a star-shaped arrangement of muscles. Because these muscles are under nervous control they can contract instantly to expand the pigment bag into a large disk. The color of the pigment may be red, brown, or yellow, depending on development and species. Each of the hundreds to thousands of chromatophores can be controlled individually by the cephalopod, allowing a wide variety of patterns to be either changed rapidly or held for prolonged periods.
Although some deep-sea species do not have functional chromatophores, this system is characteristic and unique to the neocoleoid cephalopods. On some areas of the body of many species the chromatophores overlie reflective tissue composed of iridophores. Light is reflected by layers of special cells called iridocytes. The animal can control color, contrast, brightness, and patterns on their bodies in less than one second by nervous control of the interaction of light with the pigments and reflectors in the skin. Some cephalopods also have permanent white patches, leucophores, just below the iridophores. The iridophores and leucophores can be hidden by expansion of the overlying chromatophores or exposed by their contraction, so producing patterns in their skin.
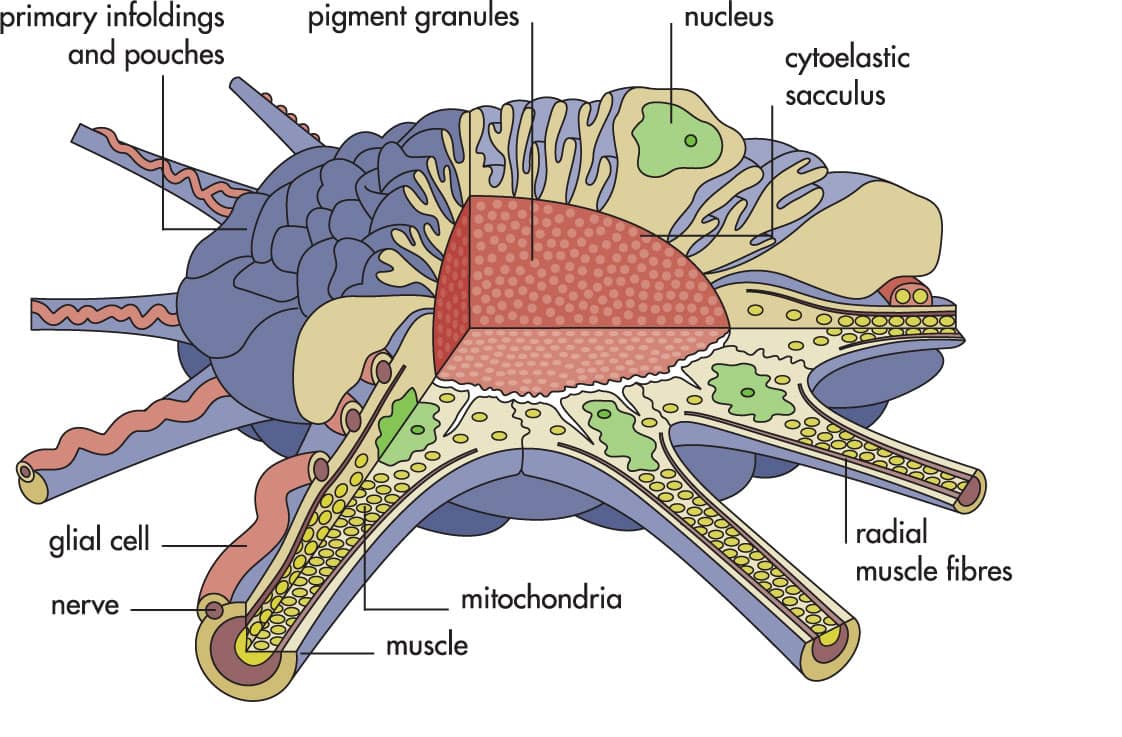
Diagram of the squid chromatophore organ illustrating the complexity of its many cell types that enable opening and closing in one fifth of a second to produce different skin patterns.
SHAPE-SHIFTING SKIN
In addition to the colorful body patterns, octopuses and cuttlefishes can morph their skin into three-dimensional shapes to enhance camouflage when nearby structures also have fine three-dimensional texture. These muscular “papillae” can also be expressed quickly and to any degree—from fully extended down to flat and invisible. Papillae come in a wide variety of shapes, from simply conical to bi-lobed, tri-lobed, or even flat. Some are short, some are long, some are fat, and others are skinny. Some papillae can be quite flamboyant in some octopods and cuttlefishes (see Flamboyant cuttlefish, shown here). The variety is impressive and they have not yet been cataloged and classified properly. No other animal on the planet can do this.
Some cephalopods have permanent skin texture that consists of various-shaped tubercles composed of cartilage or fibrous connective tissue. Distinctive papillae in some locations are sometimes referred to as cirri (superocular cirri, for example, are papillae above the eyes; in some species these are complex and may have embedded tubercles). Some octopods also have a semi-permanent lateral ridge of skin around the mantle.
MAKING LIGHT WITH BIOLUMINESCENCE
Some cephalopods—mainly nocturnal or deep-sea species—have light-producing organs called photophores. There are two fundamentally different types. The first type, intrinsic photophores, produce light biochemically. The second type, the bacterial photophore, uses symbiotic light-producing bacteria that grow in special chambers within the host. These chambers are generally associated with the ink sac. Light can be produced within the bacterial chamber or, in some circumstances, the photogenic bacteria may be expelled with discharged ink, creating a glowing cloud in the water. Bacterial photophores are found in some species of the families Loliginidae and Sepiolidae, which use bioluminescence for counterillumination, the opposite of well-known countershading. Intrinsic photophores are known in many oegopsid squids and in vampires and are thought to be used in signaling as well as camouflage.
Octopods were long thought not to have photophores, but females of some pelagic incirrates are now known to develop large photophores around their mouths when they approach maturity. Also, one species of cirrate octopod (Stauroteuthis syrtensis) has been shown to have photophores associated with its suckers. Their behavioral functions are unknown.

White leucophores in the skin of the California species Octopus rubescens; note the variable shapes and that some of them are obscured by expanded chromatophores.

Close-up photograph of the skin of the cuttlefish Sepia officinalis showing the white leucophore spots surrounded by expanded red and brown chromatophores.

Octopus tetricus with extended skin papillae that produce a very textured skin to aid camouflage.
The light produced by both types of photophore is generally blue-green in color but this can be altered by structures associated with the photophore. As with the iridophores, flat platelets composed of the protein reflectin are formed to reflect and direct the light produced by the photophores. In some cases, the photophore lenses absorb selected wavelengths of the light, allowing other wavelengths to pass through. As a result, some photophores are blue, while others are green, and some are even red. In addition to structures that reflect and refract light, opaque structures around the photophores absorb light going in unwanted directions or at unwanted times. Some photophores have opaque lid-like structures that can be blinked open and shut to emit brief flashes of light.

The squid Sepioteuthis lessoniana at night showing a disruptive skin pattern. Note the broad fin around the mantle and the precise postures of the eight arms and two tentacles.
THE FAST COLORATION CHANGE PROCESS
It was recently established experimentally that cephalopods control their body patterning visually. For camouflage, they view their surroundings and quickly process that visual information, and then the central brain sends neural signals throughout the skin to millions of chromatophores and iridophores to produce the appropriate body pattern. Astonishingly this entire process can occur in as little as 200 milliseconds (one fifth of a second)! The diagram opposite illustrates this sophisticated process in cuttlefish.

NATURE’S BEST CAMOUFLAGE
THE MASTERS OF CAMOUFLAGE ON THIS PLANET are universally accepted to be cephalopods, which can create optical illusions in the blink of an eye. An octopus or cuttlefish on a coral reef can produce effective camouflage on any of the hundreds of backgrounds that it encounters in this most complex of nature’s habitats. The selective pressure for this unique capability emanates from the extraordinary variety of visual systems of multiple predators, particularly fishes, diving birds, and marine mammals.
Camouflage is all about visual perception. Primarily it centers around the visual capabilities of various predators, but in the case of cephalopods it is also about their own visual capabilities and decisions about which camouflage tactic to deploy. Thus, having the ability to change camouflage brings with it the burden of developing a method to swiftly analyze a visual scene, extract its key features, and then deploy an effective camouflage pattern. This is a remarkable cognitive process that involves decision-making at a high degree. It begs three basic questions: what are the fundamental tactics of camouflage that deceive such a wide range of predators? How many camouflage patterns does an individual cuttlefish or octopus have the ability to produce? How quickly can they change their camouflage?
DYNAMIC CHANGES TO AVOID DETECTION OR RECOGNITION
Octopus cyanea on Pacific coral reefs change their appearance more than 150 times per hour while foraging each day! Fast change is needed because cephalopods are mobile and move into different visual scenes constantly when foraging. During their stop-and-go foraging trek, they change their camouflage pattern each time they stop and it is tailored to that particular background. The speed of change is remarkable. By studying video of naturally foraging octopuses and cuttlefishes, it is known that they change in a fraction of a second. That is, they can create patterns in their skin in as little as 200 milliseconds; complex patterns may take up to 2,000 milliseconds (2 seconds) but usually it is in the order of 200–300 milliseconds, which is the speed of a human eyeblink.
Camouflage is something that cephalopods do almost all of the time because they are soft bodied and their primary defense is to not be detected or recognized in the first place. This presents different challenges when they are swimming in the water column or down near the bottom, where there can be a great deal of structure and color, or there can be open sand or mud plains that create different survival challenges.
The fundamental tactics are: 1) remain undetected; 2) disrupt body form so that the cephalopod is not recognizable as a distinctive octopus, cuttlefish, or squid; or 3) look like an uninteresting or distasteful object. Most animals in other phyla use one of these tricks, but cephalopods use all three with variations on each for unparalleled diversity.
How many camouflage patterns does a cuttlefish have? The European cuttlefish Sepia officinalis has been studied in the greatest detail both in its natural habitats and in the laboratory. Those extensive studies have revealed an unexpected finding: this species appears to have just three basic camouflage pattern templates (each with variations on the theme) to utilize on the wide range of backgrounds that it encounters over its very broad range. These patterns—called Uniform, Mottle, and Disruptive—are illustrated above right.
BACKGROUND MATCHING & DISRUPTIVE COLORATION
Resembling or matching the visual background is an effective method of remaining undetected and cephalopods excel at this. Uniform and Mottle patterns are shown by a wide variety of cephalopods such as the common European cuttlefish, which can swiftly change from conspicuous to Uniform or Mottle camouflage.

The three basic pattern types that cuttlefish use for camouflage: Uniform, Mottle, and Disruptive.Each individual can show these patterns.

This octopus is resembling the dark sand in pattern, color, brightness, and even the texture of its skin. The small white leucophore spots also represent a random sample of other small white pebbles in the background.
Disruptive patterning is a less obvious way to attain camouflage. Some disruptive patterns of cuttlefishes tend to blend into the background while other disruptive patterns—even shown by the same animal—are clearly detectable yet they render the cuttlefishes very difficult to recognize because they create false edges and put on light and dark patches of variable size, shape, and orientation so that it is hard to distinguish head from mantle, front from back.
Octopuses are very adept at background matching. However, due to their very flexible arms and non-rigid body mantle, they can contort themselves to cause visual disruption without creating as many skin patches as are needed by cuttlefishes or squids.
MIMICRY & MASQUERADE
Mimicry is looking like another animal, while masquerade is looking like an uninteresting object. Both tactics confuse recognition and both can be found in certain cephalopods. Mimicry is commonly understood to imply the resemblance of one animal (the mimic) to another animal (the model) such that a third animal (a predator) is deceived by their physical similarity into confusing the two. Only three cases of defensive mimicry are authenticated in cephalopods and all three are octopuses. The so-called “mimic octopus” Thaumoctopus mimicus (here) and the undescribed “white V octopus” of tropical Indonesia, and the Caribbean long-arm octopus Macrotritopus defilippi mimic the shape, coloration, and swimming speed of local flatfish when they are moving. All three species live in open sandy flats where they would be detected by predators, so the guise is to look like some other animal.
Masquerade is widespread among shallow-water octopuses, cuttlefishes, and squids. To a human observer, cephalopods can sometimes look like stones, algae, seagrasses, and other objects such as soft and hard corals. Octopuses, cuttlefishes, and squids all use arm postures to enhance masquerade matching, and they are very good at using their vision to decide when and how to position their arms to help match nearby objects.

Sepia officinalis in a conspicuous zebra pattern while swimming, then note the rapid change to camouflage in the next image.

The cuttlefish settled and in a fraction of a second deployed a Mottle pattern to match the background.
MOTION CAMOUFLAGE
Motion camouflage is moving in a fashion that decreases the probability of detection by a predator. Octopuses are good at this—they can move with stealth across open areas without being recognized. The cognitive aspects of this behavior are noteworthy because the speed of the octopus is generally similar to the speed of rippling light in that environment, suggesting that the octopus consciously regulates its stealth speed in accordance with ambient motion such as dappled sunlight from waves. Moreover, they can change their overall body shape and skin texture according to other three-dimensional objects in the distant background as shown in the illustration below, based on Octopus vulgaris in the Caribbean. That is, they can perform the “moving algae” trick when spiky algae are nearby, or the “moving rock” trick when smooth coral heads or rocks are nearby.

When Sepia officinalis settle on pebbles or rocks with high-contrast light objects, they will respond with a Disruptive pattern to obscure their body outline.

Here the “mimic octopus” Thaumoctopus mimicus is mimicking a swimming flounder.
Octopus vulgaris can fine-tune its motion camouflage to resemble the shape of nearby algae (left), corals, or rocks (right) while it is moving.
WHEN CAMOUFLAGE FAILS
WHEN THE PRIMARY DEFENSE OF CAMOUFLAGE FAILS, as it sometimes does, cephalopods have a two-stage response of secondary defenses that startle then confuse predators. The first stage—termed deimatic behavior—is meant to make the predator hesitate in its attack sequence. The second stage—termed protean behavior—involves complexes of erratic unpredictable escape maneuvers, color changes, and behaviors. Together, these constitute a formidable defense strategy suitable for the soft-bodied cephalopods, which are otherwise quite defenseless.
MULTIPLE ESCAPE TRICKS
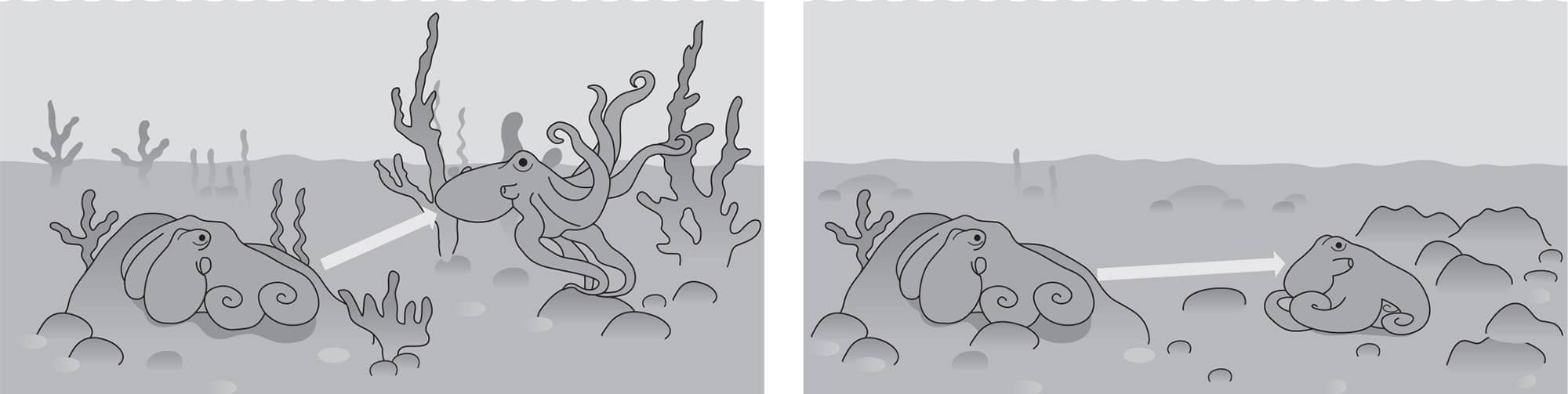
To interrupt the first stage of attack, cephalopods first use either a Stay or a Go tactic. The “Stay” tactic is a conspicuous deimatic display in which an octopus spreads the web between its arms to look bigger than it is, and makes a large dark spot around the eyes to startle the prey to make it hesitate just temporarily. Conversely they may immediately ink and jet away, which is a “Go” tactic.
This is immediately followed by erratic unpredictable escape maneuvers. This phase of secondary defense is aptly called protean behavior after Proteus—the Greek god of unpredictability. These behaviors can be very complex—some octopuses can mix signals, maneuvers, and camouflage up to ten times in just 13 seconds. One such segment is the “fake right and go left” maneuver shown below. That one segment, extracted from video, took only 400 milliseconds.
TAILORED ESCAPE TO DIFFERENT PREDATORS
In filmed sequences underwater, as well as during laboratory experiments, it was found that cuttlefishes are capable of recognizing different predators and they modified their secondary defenses accordingly. This was true even for hatchling cuttlefishes that had no experience with predators, so this initial recognition represents an innate ability. Thereafter, however, the cuttlefishes were continually making decisions about how to implement protean behavior and they did so according to the microhabitat they were in at that time. This sort of swift sensory processing and decision-making is characteristic of high-level cognitive processing that large brains can enable.
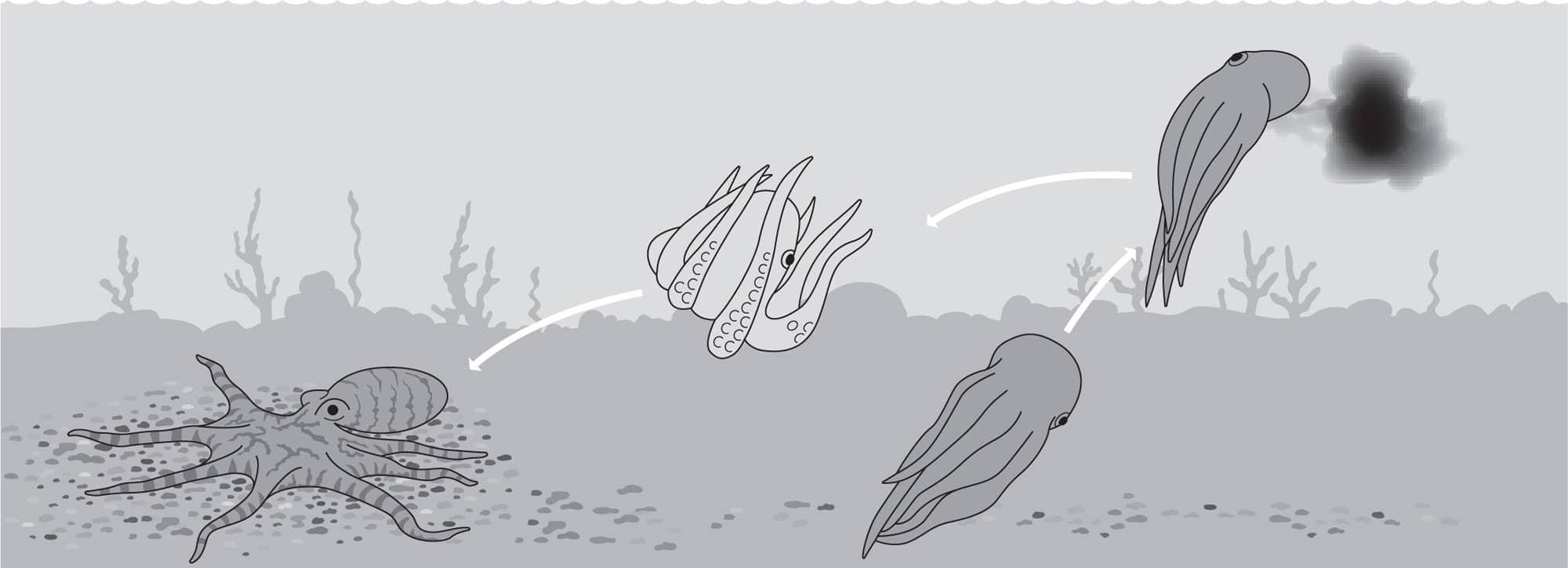
“Fake right, go left” is the trick being used here to confuse a predator, whose eye is drawn to the black ink puffed to the right while the octopus blanches, quickly descends to the bottom and camouflages itself.

This “deimatic display” of Octopus vulgaris is shown briefly to startle predators that have approached very close; the octopus is making itself look larger and different.

Octopus vulgaris performing the “Blanch—Ink—Jet Away” maneuver at high speed as secondary defense after its primary defense of camouflage fails.
SUPER FIGHTS FOR MATES
BECAUSE MANY CEPHALOPODS LIVE FOR ONLY ONE year and reproduce once, males have forged a variety of tactics to acquire a female mate. The male–male aggression is intensive and there are long sequences of fighting bouts in which multiple visual signals and postures are used. Some of these displays rival those of birds and mammals, and some fights escalate to pushing and biting, but not to death. Contestants continually decide whether to withdraw or escalate fights depending on the opponent’s actions.
ELABORATE GIANT AUSTRALIAN CUTTLEFISH DISPLAYS
Sepia apama in southern Australia aggregate once per year for spawning. Throughout the mass spawning season (May to August) there are about 180,000 cuttlefishes right at the shoreline in just a 21/2 mile (4 km) stretch of rocky reef coastline. Some males are large, and others are small. Large males fight each other to gain some advantage in pairing temporarily with a female mate. Intense male–male aggression results in elaborate fighting displays, sometimes with roving dark bands moving over the body. Parts of these displays are very stereotyped and sometimes the males are locked in battle for many minutes. Recent experiments and field trials have shown that each male is continually assessing the other during these fights and adjusting his fighting tactic. This level of cognition compares with birds and mammals.
CARIBBEAN REEF SQUID DUETS
Sepioteuthis sepioidea is among the most sophisticated of squids and has both courtship and fighting displays. Females incite males with a bright white pied pattern that leads to male–male fights as elaborate as those of giant cuttlefishes. These fights are very short by comparison (a few seconds each) and the pattern on the top of the squid is different from that on the bottom of the squid, and each conveys a different level of aggression. Males can then show a silver pattern on one side to repel approaching males while showing the female a brown courtship pattern on the other side of its body.

Large male Sepia apama are fighting one another with flared arms, dark head and fins, and moving dark bands on the mantle to convey different levels of aggression.

The female Sepioteuthis sepioidea (top) shows a white “pied pattern” that incites males to compete for her. The male (bottom) has begun to pair with her and will fight other males as they approach.

The male (left) is showing a courtship pattern to the female while showing an aggressive “Unilateral Silver” on the other side to ward off approaching male rivals. When the female switches side the male immediately switches his pattern so that she does not see the aggressive pattern.
SNEAKY MALES & SNEAKIER FEMALES
SEX GAMES IN CEPHALOPODS ARE NOT A ONE-WAY street. For males, there are significant challenges to obtaining a mate because in some systems there are more males than females at the time and place of reproduction. Strong male competition has resulted in various sneaker tactics; that is, methods to obtain a mate that do not require fighting another male. For females, the challenge is to choose among many male suitors, and they can do this “cryptically” after mating whether the males are there or not.
CLEVER MALE TACTICS TO ACQUIRE A MATE
The ratio of males to females in the mating aggregation of Sepia apama in South Australia ranges from 4:1 to as high as 11:1, creating a strong bias that forces strong competition among males. Large males fight each other to become consorts to females, but smaller males are numerous and have only one spawning season to mate. They use fast dashes towards the female to obtain a quick copulation; when four or five small males are doing this all the time they distract the large consort male who cannot fend all of them off. If this does not work, small males will surreptitiously sneak under the rock where the female is laying her eggs and mate her out of sight of the large guarding consort male.
In the most extraordinary tactic of all, small Sepia apama males will “morph” into a female by hiding their fourth arms, which in males are very long and elaborate (here). They then bulge their arms at the base as if holding a large egg as females do, and with this “sexual mimicry” they swim right past the large male, who thinks he is acquiring another female mate, and the sneaker male can then mate with the female.
Small males seem “pretty smart” because they are making many decisions during these active episodes. In fact, they are very flexible in their behaviors and are aware when the behavioral context changes. For example, if the large consort males leave for some reason, a small sneaker will move right up to a female and he will now act as a consort, not a sneaker, and he will fight off other sneakers. Squids can do this too. Such conditional mating strategies are characteristic of vertebrates including fishes and mammals.
On the other hand, the large consort male seems “pretty dumb” in these encounters because we know cuttlefishes have short-term and long-term memories, and he has just been chasing away that sneaker male for tens of minutes, and suddenly it morphs into a female and waltzes right in without the large male figuring it out. Not only that, but in a few cases the large consort male also tries to mate with the female mimic. Hence the visual deception on behalf of the small sneaker is highly effective.
CRYPTIC FEMALE CHOICE
Female cuttlefishes are also deciding with whom to mate. They reject 70 percent of mating attempts overall but sneakers gain many copulations, so sneaking is a successful male tactic, although it does not necessarily mean they gain fertilizations. Females on breeding grounds often have stored spermatophores from five to eight males and so have multiple sources of sperm available when they fertilize and lay each large egg. Large consort males that are guarding females as they lay eggs do not interfere with the female. This is the stage at which females are sneakier than males, because DNA fingerprinting shows that females are probably manipulating sperm in some unknown manner and “cryptically” choosing some sperm sources over others. Males are ignorant of these processes!

A typical mating pair of the giant Australian cuttlefish, Sepia apama. The female is on the left. These pairs are very temporary, rarely lasting more than a day and sometimes only an hour.

Sexual mimicry by a small male Sepia apama. The large consort male is reaching upwards to attempt to mate a small male that is imitating a female by hiding his dimorphic fourth arms, bulging his arms as if holding an egg, and displaying the female signal of white rim around his mantle. The female is below the large consort male, who is guarding her.
EVALUATING INTELLIGENCE IN SUCH BIZARRE ANIMALS
POPULAR OPINION IS THAT CEPHALOPODS ARE SMART or intelligent, yet it is very difficult to assess such qualities given our human biases. We have to immerse ourselves in the sensory world of an octopus, squid, or cuttlefish before we can begin to analyze its range of abilities. This has not yet been accomplished in an unequivocal scientific manner. Also, some behaviors of some cephalopods appear to human observers to be quite stupid and inappropriate. Let us review this.
Cephalopods certainly process many features that we equate with smartness or intelligence in vertebrate animals with which we, as humans, are more familiar: think of birds, rodents, and dogs. Intelligence, in most people’s understanding, involves combining pieces of evidence to reach non-obvious conclusions. Cephalopod learning and memory capabilities are substantial, and most importantly it is clear from field and laboratory work on cephalopods that their behavioral repertoire is very wide by any standard, and they are making decisions constantly for a wide variety of interactions with predators, prey, and members of their own species. They have keen well-developed senses, a large complex brain, and a vast network of nerves throughout their skin and other body organs. It has often been postulated that cephalopods live a difficult life in the behavior space dominated by vertebrates, and that these selective pressures have forged the ecologically successful cephalopods we know today.
EMBODIED COGNITION
It is difficult to judge cognitive sophistication in an animal group with such a bizarre body organization. After all, these animals have their head on their foot (cephalo pod), they have a brain built on a molluscan plan, and they live in marine environments—with which most humans are unfamiliar. Perhaps it is more appropriate to judge their performance relative to the specific brain and body organization that they possess. This concept of “embodied cognition” has gained recent acceptance among neuroscientists and behavioral ecologists because it draws attention to the adaptive qualities that the animals possess. Anyone who has spent a lot of time underwater immersed in the sensory world of an octopus and has observed its wide range of behaviors over long time spans would agree that these animals clearly possess a brand of what humans call “intelligence."
ADVANCED INVERTEBRATES
In the terrestrial world—with which we humans are familiar—intelligence or smartness is calculated partly on behaviors such as exploration, play, tool use, or even consciousness. Moreover, social interactions also play a large part in such human assessments. However, except for certain squid species, cephalopods are not social. Efforts to demonstrate play and tool use in octopuses are just beginning and the results are not yet convincing; more experimental and field studies with several species are needed. There are no data available on consciousness although speculation continues.
However—and this is perhaps the most striking fact to emerge from the last 20 years of research—cephalopods are “advanced” invertebrates: almost every system that has been studied recently is extraordinarily sophisticated and unique, the result of over 200 million years of evolution. Finally, there is the intriguing possibility that cephalopods have evolved complex behaviors with cortical structure and neural pathways that differ from the vertebrate lineage. This idea deserves attention given that cephalopods are the only group to have diverged from the vertebrate line and evolved large nervous systems and complex behaviors.

This octopod is swimming in the water column, which is very rare for these bottom-dwelling creatures.
COMMON OCTOPUS
Octopus vulgaris
THE MOST COSMOPOLITAN SHALLOW-WATER octopus in the Atlantic is a master of camouflage in habitats as diverse as coral reefs, coral rubble, seagrass beds, rock reefs, and sometimes open mud and sand plains. It is found on both sides of the Atlantic from tropical to temperate, throughout the Mediterranean and Caribbean Seas, and is fished commercially off the Sahara Bank of Africa.
A COGNITIVELY ADVANCED CREATURE
The large complex brain of this octopus enables it to carry out a wide range of behaviors in complex habitats with many predators. Learning and memory are particularly well established in these bottom-dwelling cephalopods. This and other species conduct daily forages spanning dozens of feet and different microhabitats, yet they always find their way back to the den, presumably by spatial memory. Their soft pliable body enables them to squeeze into small holes or dens for shelter during most of the day and night. They use keen vision to forage on likely food spots and then feed in a tactile manner by enveloping coral heads and rocks that harbor shellfish and crabs. Some scientists think octopuses are capable of tool use and play although this is controversial.
FAMILY |
Octopodidae |
OTHER NAMES |
None |
TYPICAL HABITAT |
Seagrass beds, shallow coral reefs, rock reefs |
SIZE |
Mantle length to 8 in (20.5 cm) |
FEEDING HABITS |
Forages daily for mussels and small crabs; rarely eats fish |
KEY BEHAVIORS |
Day-active, sophisticated camouflage, spatial memory to find its way back to den |

STARTLING BEHAVIOR
This conspicuous deimatic display requires the octopus to “stand” on four of its eight arms and to spread its flexible arm webs maximally to look larger. It then produces dark high-contrast markings that make the eye appear larger as well. The octopus maneuvers so that it is always presenting its largest apparent size to the threat (usually a predator).
DAY OCTOPUS
Octopus cyanea
THIS LARGE, DAY-ACTIVE OCTOPUS IS VERY COMMON on coral reefs throughout the tropical Pacific Ocean. This is one of the largest octopus species and is among the most beautiful and colorful. Its camouflage is superb—it has to be since Pacific coral reefs have such a huge diversity of corals, sponges, and algae and there are dozens of rapacious predators looking for this high-protein meal. This graceful octopus is well known to divers and is also the target of small artisanal fisheries (mainly through spear fishing) throughout many of the Indo-Pacific islands.
KEYSTONE REEF PREDATOR
Octopus cyanea lives in large dens on the reef top and forages several hundred feet per day in search of mussels, clams, and crabs hiding among the corals and crevices. When these octopuses forage, small groupers follow them and feed on other small shrimps and fishes that escape as the octopus “parachute attacks” small rock promontories. Hence the octopus is grazing the filter-feeding molluscs and indirectly feeding small groupers. At the same time, the larger predators on the reef such as snappers and barracuda are preying upon the octopuses, particularly when they are small. The octopus is in the middle of the food chain on a coral reef and this key ecological niche that they occupy deserves conservation efforts to protect this keystone reef species.
FAMILY |
Octopodidae |
OTHER NAMES |
Big blue octopus, Cyane’s octopus |
TYPICAL HABITAT |
Shallow coral reefs throughout Indo-Pacific Oceans |
SIZE |
Mantle length 8–10 in (20–25 cm) |
FEEDING HABITS |
Preys on live crabs, mussels, and fish |
KEY BEHAVIORS |
Day-active, sophisticated camouflage ability, parachute attacks on likely food spots |
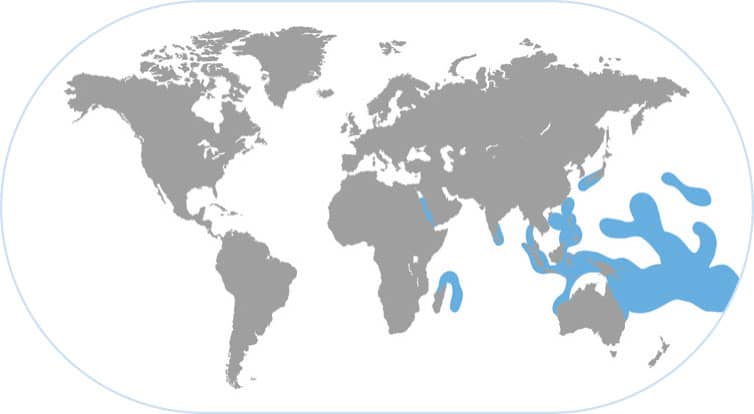

KING OF CAMOUFLAGE
The day octopus may represent the pinnacle of sophistication among octopuses because it lives in the most complex of habitats: Pacific coral reefs with great structural diversity and biodiversity. During a typical 4-hour forage they may change their skin patterning more than 150 times to accomplish camouflage amidst different corals.
MIMIC OCTOPUS
Thaumoctopus mimicus
THIS SPECIES IS FROM A CLADE OF LONG-ARMED octopuses that have adapted to the bizarre underwater seascapes of open sand. These habitats are known to marine biologists as “muck dive sites” because they are devoid of coral or other structures, and are mostly characterized by seemingly featureless sand or mud bottoms. A shrimp, eel, fish, or cephalopod found in these habitats always has some specialized behavior enabling it to thrive in this strange environment.
ADAPTIVE IMITATION OF DIFFERENT ANIMALS
Both the Pacific and Atlantic mimic octopus species imitate swimming flounders and soles most of the time when they need to move swiftly and swim longer distances. The Indo-Pacific species has been reported to imitate up to a dozen animals such as shrimp, jellyfish, and snake eels, but there are few data to support these observations. Clearly this species can contort its body to look conspicuously like a variety of objects, many of which appear to a human to be other local species. Only recently discovered, it is not clear who the common predators of the mimic octopus are. Moreover, it would be informative to determine if these behaviors are learned or innate, and whether they are different in different locations across the wide range of these octopuses.
FAMILY |
Octopodidae |
OTHER NAMES |
None |
TYPICAL HABITAT |
Sand and mud adjacent to coral reefs |
SIZE |
Mantle length about 2 in (5 cm), total length including the very long arms about 24 in (60 cm) |
FEEDING HABITS |
Preys on live crabs and shrimps |
KEY BEHAVIORS |
Aggressive imitation of a variety of marine species |
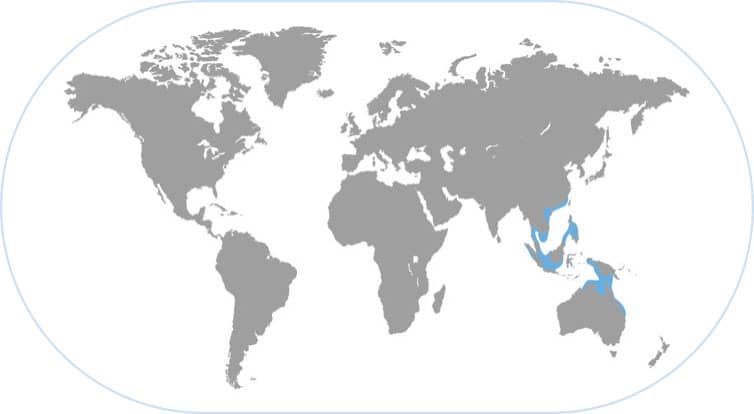

SHAPE SHIFTER
The Indo-Pacific mimic octopus lives on open sand plains between coral reefs. In addition to camouflage for defense, it often mimics flounders and other sand-dwelling animals to confuse some of its predators while it is foraging to find food in these open flat habitats that do not provide hiding places.
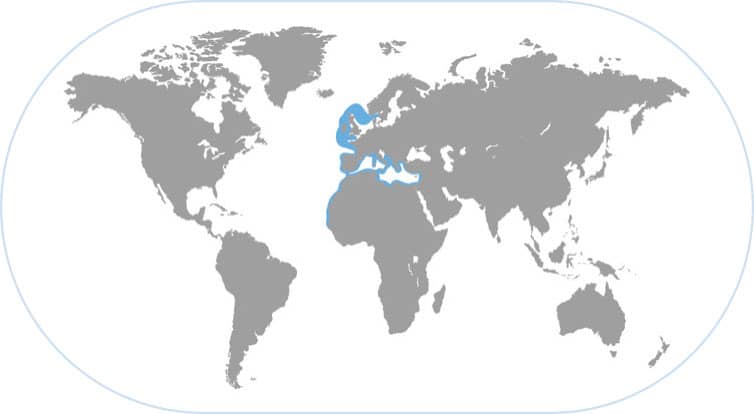
COMMON EUROPEAN CUTTLEFISH
Sepia officinalis
THIS FABLED SPECIES WAS FIRST WRITTEN ABOUT BY Aristotle, who marveled at its color-changing capabilities and the clouds of ink it emitted when alarmed. It has a wide geographic distribution in the eastern Atlantic Ocean from the North Sea southward throughout the Mediterranean and all the way to equatorial West Africa.
A WILY PREDATOR
Along with Octopus vulgaris, Sepia officinalis is one of the best-studied cephalopods in biology because it occurs in reasonable abundance near many marine laboratories throughout Europe. This cuttlefish is easy to keep or even culture in the laboratory. Hence, it has been very useful in elucidating how vision controls changeable camouflage, because any visual background you present it with will be quickly analyzed and answered with an appropriate body pattern. In nature, its camouflage capabilities are excellent, not only to deter predators, but to hide from unwary shrimps and fishes that it stalks slowly then captures with its lightning-fast strike using its two long tentacles. Sepia officinalis can also partially bury in the sand and act as a “lie and wait” predator. This species is fished commercially on a large scale on the Sahara Bank of northwest Africa.
FAMILY |
Sepiidae |
OTHER NAMES |
Common cuttlefish |
TYPICAL HABITAT |
Sandy seagrass beds, shallow rock reefs |
SIZE |
Mantle length about 12 in (30 cm) |
FEEDING HABITS |
Lie and wait predator of small fishes; eats large crabs on occasion; hunts day and night |
KEY BEHAVIORS |
Superb camouflage, sometimes buries in sand; aggressive attack with two long tentacles |

RAPID COLOR-CHANGER
This “Zebra display” is typically shown by large males during intensive fights with other males. Each zebra skin pattern is unique, like a fingerprint. This pattern and others can be switched off or on in less than one-third of a second so that they can conduct rapid behavioral sequences with visual impact.
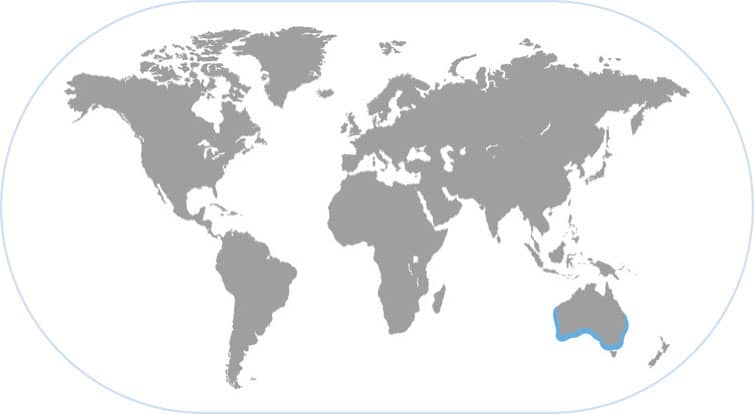
GIANT AUSTRALIAN CUTTLEFISH
Sepia apama
THIS NATIVE OF SOUTHERN AUSTRALIA GROWS UP to 26 lb (12 kg) and is the largest known cuttlefish species. Males have the long floppy and colorful fourth arm that is used as a sexual signal of male-ness. At the beginning of each austral winter (May/June), approximately 200,000 cuttlefishes migrate to a small spawning ground in northern Spencer Gulf, north of Adelaide.
ONLY CUTTLEFISH THAT FORMS HUGE SPAWNING AGGREGATIONS
This unique mass spawning is a “cuttlefish wonder of the world” and SCUBA divers can immediately see hundreds of cuttlefishes during a typical dive right next to shore. There are four to eleven males for every female on the spawning grounds, and this has led to some remarkable and sophisticated tactics by males to obtain a mate, including sneaker males and even sexual mimicry. Males conduct elaborate fights with one another to gain short-term consortships with females, who lay eggs under flat rocks as the males guard them against rivals. When the cuttlefishes are not competing for mates, they are camouflaging themselves in this rocky reef habitat. This is perhaps the best-studied and most complex mating system known in cephalopods and it rivals those of many vertebrate animals, including mammals.
FAMILY |
Sepiidae |
OTHER NAMES |
Giant cuttlefish |
TYPICAL HABITAT |
Rock reefs, open sandy and seagrass beds |
SIZE |
Mantle length up to 20 in (51 cm) |
FEEDING HABITS |
Forages daily for small crabs and fishes |
KEY BEHAVIORS |
Solitary most of life cycle; day-active, sophisticated camouflage, complex reproductive behaviors |

DISTINCTIVE MALE ARM
This large male cuttlefish is showing the elaborate green iridescence on its fourth right arm, which is very large compared with the other three arms on that side. This is the only visible dimorphic character that distinguishes males from females. Males also use this arm in fighting displays against other males when they are competing for female mates.
BROADCLUB CUTTLEFISH
Sepia latimanus
THE BROADCLUB OVERLAPS WITH THE DAY OCTOPUS on Pacific reefs and is day-active in these complex environments. This large and colorful species ranges widely in the western Pacific Ocean and lives exclusively on coral reefs, where it usually hovers motionless and undetectable like a mysterious flying saucer as it scans the scene for predators and prey.
CLEVER QUEEN OF PACIFIC REEFS
Despite their relatively large size and the clear water of coral reefs, these cuttlefishes are hard to find because they use so many camouflage tactics. While Octopus cyanea is the king of camouflage for reef bottom dwellers, Sepia latimanus might be considered the queen of camouflage on the same reefs: it does not sit on the bottom but hovers in and around corals and other reef structures. With its superb vision it determines where to forage in a stealth mode (that is, slowly and with changeable camouflage) and stalks small fishes. Occasionally it encounters a large crab, and then produces a dazzling visual display known as the “Passing cloud” in which fast-moving dark bands traverse its body—seeming to mesmerize the crab visually, which it then attacks. The cuttlefishes and octopuses can cohabit the same reef because they use different vertical space, forage differently, and eat different prey.
FAMILY |
Sepiidae |
OTHER NAMES |
None |
TYPICAL HABITAT |
Coral reefs throughout the western Pacific |
SIZE |
Mantle length about 12 in (30 cm) |
FEEDING HABITS |
Forages daily for small fishes and large crabs |
KEY BEHAVIORS |
Solitary most of life cycle; day-active, sophisticated camouflage, complex reproductive behaviors |
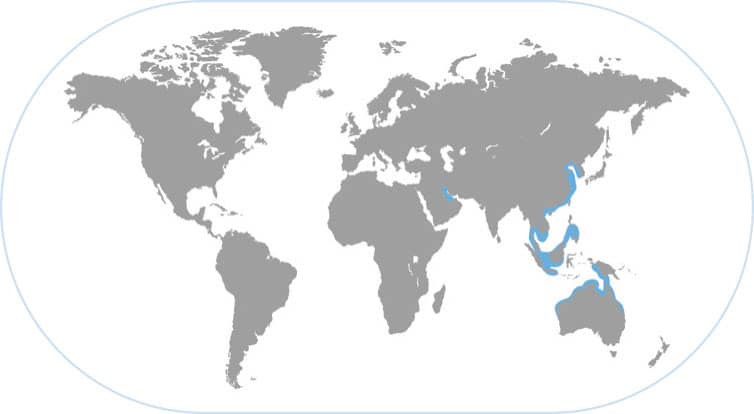

RAPID PATTERN CHANGES
The broadclub often deploys a characteristic white bar pattern (above) that disrupts its recognizable shape to predators. It can instantly change its body pattern to a uniform coloration (below) to help it resemble light-colored backgrounds such as coral or sand.

LONG-FINNED INSHORE SQUID
Doryteuthis pealeii
THIS SQUID LIVES IN LARGE SCHOOLS FOR MUCH OF its short life span of 6–12 months and it supports a large and valuable fishery off the eastern USA. It is a sleek predator capable of very fast jet propulsion (both backward and forward). Off the coast of New England, these squids migrate offshore to deeper water for the winter, then return to shallow nearshore spawning grounds in the summer.
THE SQUID NERVOUS SYSTEM STUDIED FOR NEUROSCIENCE
The long-finned inshore squid schools generally disperse at night to feed and aggregate in schools as a defense against predators during the day. Their large eyes are capable of seeing polarized light, which enables them to see and consume transparent prey organisms. When a large predatory fish approaches a school of squids, some of them will swiftly descend to the bottom and camouflage themselves by producing dark transverse bars across their body. Many aspects of the biology of this species are known because it has been studied for so long in Woods Hole, Massachusetts, where scientists from around the world gather during the summer. The nervous system, especially the giant axon, is a famous biomedical model.
FAMILY |
Loliginidae |
OTHER NAMES |
Summer squid, Woods Hole squid |
TYPICAL HABITAT |
In summer, open sandy and seagrass beds near shore; in winter, in deep canyons offshore |
SIZE |
Adult males about 12 in (30 cm) long, females a bit smaller |
FEEDING HABITS |
Forages nightly for fish; sometimes cannibalistic |
KEY BEHAVIORS |
Schooling near the bottom; dispersing at night to feed over large areas; complex reproductive behaviors |
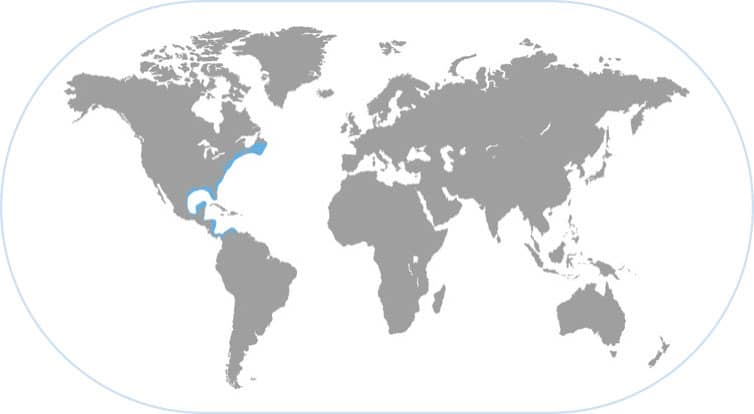

MOVING IN FOR THE KILL
This sleek nearshore squid can jet very fast backward to avoid predators or forward to capture prey. The two long tentacles can just be seen in this photograph at the tip of the eight arms as the squid is about to attack a shrimp.
CARIBBEAN REEF SQUID
Sepioteuthis sepioidea
SQUIDS ARE SCHOOLING ANIMALS THAT HAVE social interactions, particularly with respect to sexual selection. The Caribbean reef squid has an elaborate mating system in which females show body patterns that incite the males to begin competing for them. The males then engage in action-packed fights among themselves using extravagant visual displays. Females are choosy and reject many male suitors. When a male pairs temporarily with a female, he will sometimes show a courtship pattern on the side of his body towards the female, and a fighting pattern on the other side of his body where rival males approach. This level of cognition is unknown among other invertebrates and compares to the sophisticated mating systems of fishes, birds, and mammals.
COLORFUL DISPLAYS FOR SEX AND THREAT
The reef squid lives in shallow, clear, bright water and produces the full rainbow of colors in its skin when signaling. Fighting and courtship patterns tend to be high-contrast to create unambiguous signals; examples are the “Pied” pattern of females and the “Zebra display” of males. When fish predators approach, the squids put on colorful patterns and postures that are tuned to the visual perception of those predators, for example, these squids display bright yellow in reaction to certain predators that approach closely.
FAMILY |
Loliginidae |
OTHER NAMES |
Reef squid |
TYPICAL HABITAT |
Shallow coral reefs |
SIZE |
Total length, including arms, to 8 in (20 cm) |
FEEDING HABITS |
Preys on small live fishes mainly at night |
KEY BEHAVIORS |
Huge signal repertoire, schooling, elaborate mating system |
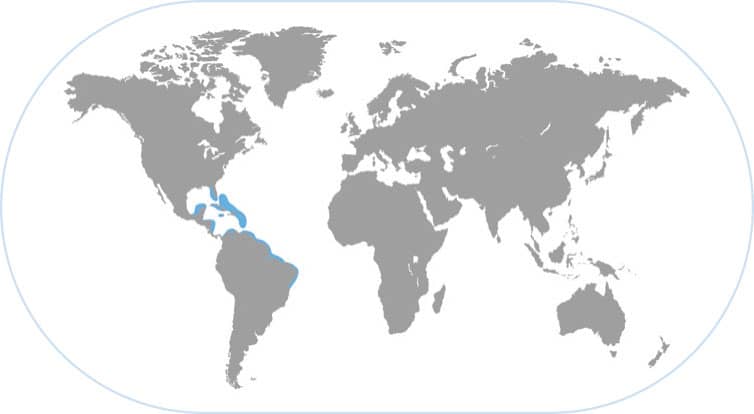

GLITTERING COLORS
The Caribbean reef squid is one of the most beautifully colored cephalopods with skin that glitters from iridescence and pigments. They are commonly seen during the day in small schools comprising a wide size range of individuals. They have different body patterns for different behaviors.