But it’s a desolate place like the Valley of the Moon in northwestern Argentina where Bonaparte and his dinosaurs are likely to be found, anytime from September to April. The conditions are harsh, the equipment primitive. Bonaparte and Sereno drove a rickety Renault with a broken fuel pump during their dig in the Valley of the Moon in 1988. An assistant had to perch on the roof much of the ride, dangling the fuel line. Bonaparte is accustomed to sleeping outdoors or in the sheep-shearing room of remote estanzias, the vast Argentine ranches. He explored Argentina’s Patagonian mountains on horseback, at least until he was turned back by fierce summer snow squalls.
—DON LESSEM, OMNI MAGAZINE, 1993
WHAT IS A DINOSAUR?
Often the first thing a paleontologist has to deal with when talking to the public is their misconceptions about dinosaurs. Many people think any large extinct animal is a dinosaur. Those bags of plastic dinosaur toys mixed with nondinosaurs such as the saber-toothed cat and mammoth reinforce this misconception. Likewise, those same plastic “dinosaur toy” sets often include the fin-backed protomammal Dimetrodon because it is also large and prehistoric—even though it is part of the mammal lineage. In movies like Jurassic World and TV shows like Walking with Dinosaurs, and in many other media, people see images of marine reptiles such as mosasaurs, long-necked plesiosaurs, and dolphin-like ichthyosaurs and assume that these large reptiles of the Age of Dinosaurs must be dinosaurs too. Another common misconception is that all dinosaurs are large, but a great many were small, including a lot that were the size of small birds. A little more excusable is the common misconception that pterosaurs are dinosaurs. The public ignorance is so annoying to paleontologists that Mark Norell of the American Museum of Natural History in New York wrote a cleverly illustrated children’s book. Every page shows a nondinosaur that the public thinks is a dinosaur, reinforcing the same point over and over again. On the other hand, if you show an image of a bird, most of the public will not call it a dinosaur—but it is.
If none of these creatures are dinosaurs, what do paleontologists mean by the term? When Richard Owen had only Megalosaurus and Iguanodon and a few others to go on, he defined dinosaurs as a group of huge extinct reptiles with a number of distinctive features (see chapter 2). As the number of new dinosaurs (and tiny dinosaurs) increased rapidly in the late 1800s and early 1900s, that definition was modified. By the time of Edward Drinker Cope’s work in the 1880s, and especially when Samuel Wendell Williston published Osteology of the Reptiles in 1925, the dinosaurs had been separated from other groups of reptiles, such as the marine reptiles, pterosaurs, and others such as the protomammals (still called “mammal-like reptiles” then). In 1878, Othniel Charles Marsh recognized four groups of dinosaurs that are still valid today: sauropods, theropods, ornithopods, and stegosaurs (figure 5.1). But few of these authors provided an exact anatomical definition of what constitutes a dinosaur.

Figure 5.1
Simplified family tree of the major groups of dinosaurs, showing the hip structure diagnostic of each group. The dark-colored bone on the lower left of each hip diagram is the pubic bone, which points backward in ornithischians (lower right corner diagram). (Courtesy of Wikimedia Commons)
In 1888, British paleontologist Harry Govier Seeley recognized two groups of dinosaurs: the “lizard-hipped” dinosaurs, or Saurischia (which included the theropods and sauropods), and the “bird-hipped” dinosaurs, or Ornithischia (which include most of the herbivorous dinosaurs except sauropods). These ideas gained widespread acceptance, and for the next 130 years most paleontologists agreed that to be a dinosaur a fossil had to be a member of one of these two groups. In 1974, Robert Bakker and Peter Galton described a number of unique anatomical features that established dinosaurs as a natural group.
In 2017, British paleontologists Matthew Baron, David Norman, and Paul Barrett published a detailed analysis of dinosaur interrelationships. Looking at the most primitive theropods, sauropods, and ornithischians, they found no unique anatomical specializations that could be used to define the Saurischia. In addition, other evidence suggests that the theropods and ornithischians are closely related rather than theropods plus sauropods together as Saurischia (figure 5.2). This idea remains controversial, and a recent response by a group of leading researchers led by Max Langer questioned many of the interpretations. These arguments are highly technical and beyond the scope of this book, so we will focus on well-defined groups of genera and families.

Figure 5.2
The new family tree of dinosaurs according to the analysis by Matthew Baron, David Norman, and Paul Barrett in 2017. Ornithischia remain a natural group, but “Saurischia” is broken up, placing theropods closer to Ornithischia than to sauropods. (From M. Baron, D. Norman, and P. Barrett, “A New Hypothesis of Dinosaur Relationships and Early Dinosaur Evolution,” Nature 543 [2017]: 501–506, fig. 1; courtesy of Nature)
As more and more fossils were found, the differences between the two groups (based on hip structure) seemed to be consistent, and the individual groups (Sauropoda, Theropoda, and so on) continued to work well. The Saurischia were the lizard-hipped dinosaurs, with the pubic bone of the hip region pointing forward (figure 5.3A and C). The Ornithischia, or “bird-hipped dinosaurs,” had at least part of the pubic bone shifted backward, parallel to the rear bone of the hip region, the ischium (figure 5.3A–B).

Figure 5.3
(A) The hip socket or acetabulum of dinosaurs is an open hole between their three hip bones where they join the middle. The “ball” or head of the thighbone (femur) inserts in this hole. On the top left is an ornithischian pelvis, with part of the pubic bone running backward parallel to the ischium. On the top right is a saurischian pelvis, with the pubic bone pointed forward. The head is to the right and the tail is to the left. (B) Close-up of the hip structure of a typical ornithischian, showing the pubic bone running beneath and parallel to the ischium (to the left of the thighbone). The head is to the right and the tail is to the left. (C) Close-up of the hips of a saurischian, showing the large pubic bone to the right of the thighbone, and the smaller ischium to the left of the thighbone. ([A] Redrawn from several sources; [B–C] photographs by the author)
However, the major suborders defined by Seeley did not answer the question of how the groups within Saurischia and Ornithischia are interrelated. I remember attending lectures from the early 1970s in which paleontologists were not sure that Saurischia and Ornithischia could be combined into the Dinosauria. In his 1955 textbook, Evolution of the Vertebrates, Edwin Colbert wrote that “the term includes two distinct reptilian orders. Consequently, the word dinosaur is now a convenient vernacular name but not a systematic one.” This sad misunderstanding might have been typical of thinking in 1955 and 1969, when the first two editions of Colbert’s book were published, but sadly it remained even in the last edition in 2001, by which time this idea had been resoundingly debunked.
However, at that very time these doubts were expressed, a revolutionary new way of thinking about animal classification called “cladistics,” or “phylogenetic systematics,” came along. First developed by the German entomologist Willi Hennig in 1950, but not translated into English until 1966, it swept through nearly every area of animal and plant systematics in the late 1960s and 1970s and has helped solve many troubling and confusing problems in animal classification. Hennig’s ideas were complex, but his principal insight was that we must base classification systems strictly on their evolutionary history, or phylogeny (hence the name “phylogenetic systematics”). For example, if birds are descended from dinosaurs, they must be classified as a subgroup of dinosaurs, not set up in their own “Class Aves” parallel to and distinct from their ancestors among “Class Reptilia.”
One of his key insights was that natural groups in classification schemes are defined based on unique evolutionary specializations, or “shared derived characters.” For example, our anatomy with four limbs goes all the way back to the earliest tetrapods (“amphibians” in the old sense) that crawled out on land, so we wouldn’t use “four limbs” as an anatomical specialization to define mammals or humans because four limbs are primitive features of all nonfish vertebrates (tetrapods). Mammals can be defined by unique evolutionary novelties such as having hair or fur and the presence of milk in mammary glands in females to nurse their young. But we wouldn’t use the presence of fur or mammary glands to define a group within mammals, such as our own order Primates. Likewise, the “lizard-hipped” condition of the Saurischia is primitive for all reptiles and doesn’t serve well to define a natural group.
Once we discarded all the “wastebasket” groups and focused on shared evolutionary specializations as the key to understanding relationships, many previously insoluble problems in classification could be resolved. This also helped redefine what it meant to be a dinosaur in the strict sense. As Jacques Gauthier first documented in 1986, one of the unique evolutionary specializations of all dinosaurs is that there is a hole right through the hip socket (“acetabulum” in anatomical lingo) rather than a pocket in the bone that holds the head of the thighbone as it moves (figure 5.3A). No other animals have this characteristic; it a unique feature that defines Dinosauria.
Additional characteristics include that all dinosaurs held their limbs straight beneath their bodies in an upright posture, just like mammals. Nearly all dinosaurs have just three or fewer fully developed fingers in their hands, with the ring finger and pinky highly reduced or missing. Dinosaurs nearly all walk on the tips of their fingers and toes, with only three toes in their feet (the fourth toe and little toe are reduced or lost). Finally, there are just three vertebrae fused to the upper part of the hip bones, connecting the spine to the hind legs and forming a sacrum. Other animals have fewer or more vertebrae in their hips. Baron, Norman, and Barrett demonstrated that a few of these features show up in the close relatives of dinosaurs, but no other group of animals combine all these features.
RULING REPTILES
Early paleontologists such as Cope and Williston first noticed that a natural cluster of reptiles included crocodiles and dinosaurs, which Cope called the Archosauria, or “ruling reptiles” in Greek. Some of the groups Cope placed in his Archosauria in 1869 have since moved to other parts of the reptilian family tree, but Gauthier’s 1986 detailed cladistic analysis of archosaurs found characteristics that define all archosaurs. The most obvious feature is that their teeth (which in most groups are simple conical pegs) sit in sockets in the jaw, whereas the teeth of lizards and snakes emerge from a continuous groove along the inside wall of the jaw. This teeth-in-sockets pattern is known as the “thecodont” condition, and for many years paleontologists used the wastebasket group “Thecodontia” to refer to all archosaurs that were not crocodiles, pterosaurs, dinosaurs, or birds.
If you look at almost any archosaur skull (figure 5.4A), you will find an opening before the eye socket called the antorbital (“ante” means “in front” of the “orbit”) fenestra, and in the lower jaw, or mandible, there is an additional hole called the mandibular fenestra. In many archosaurs, these extra holes in the skull serve as points for muscle attachment or allow jaw muscles to bulge or may reduce the bony weight of the skull. Finally, all archosaurs have an additional ridge on the thighbone called the fourth trochanter, which provides an additional anchor point for the muscles that pull the thigh backward. Thanks to these leg muscles, many (but not all) archosaurs have moved away from the completely sprawling posture of lizards, who rest on their bellies when not running, to a semi-upright posture seen in crocodilians, or the fully upright posture of dinosaurs. There are other differences as well, but these are the easiest to see on the skeleton.
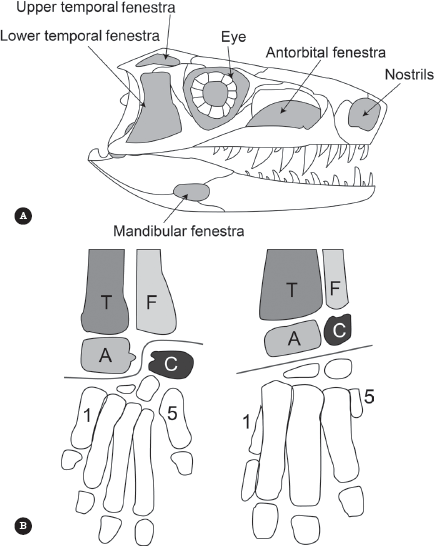
Figure 5.4
(A) Archosaur skulls all have distinctive windows, or “fenestrae,” in them that lighten the skull and add points for attachment and expansion of muscles, especially jaw muscles. The antorbital (“in front of the orbit”) and mandibular fenestrae are unique to archosaurs. (B) The ankle joints of archosaurs come in two main types. The crocodile branch (Pseudosuchia) mostly has a hinge that runs between the first row of ankle bones, the astragalus (labeled A in the figure) and the calcaneum (labeled C). The dinosaur-pterosaur branch (Ornithodira or Avemetatarsalia) has fused the astragalus and calcaneum to the tip of the shinbone (T and F, or tibia and fibula), and the hinge runs between them and the second row of ankle bones (mesotarsal joint). The number “1” is digit 1 (the big toe) and “5” is the pinky toe. (Redrawn from several sources)

Figure 5.5
Family tree of the major groups of archosaurs, showing the crocodile branch (Pseudosuchia or Crurotarsi) with the armored aetosaurs, the crocodile-mimic phytosaurs, the huge four-legged predatory rauisuchids, and the true crocodylians. The other half of the archosaurs are Avemetatarsalia or Ornithodira, the dinosaur-pterosaur branch. (From S. Brusatte, M. J. Benton, J. B. Desojo, and M. C. Langer, “The Higher-level Phylogeny of Archosauria [Tetrapoda: Diapsida],” Journal of Systematic Palaeontology, no. 8 [2010]: 3–47; courtesy of S. Brusatte)
The archosaurs include not only the living crocodilians and birds and the extinct pterosaurs and dinosaurs but a wide spectrum of other reptilian groups (figure 5.5). These were dominant in the Triassic, pushing the protomammals that had dominated the Permian and Early Triassic aside as they became the ruling reptiles. At one time, all of these extinct creatures were called by the wastebasket name “thecodonts,” but now we must discuss each one individually because there is no name for archosaurs that aren’t crocodiles or dinosaur and birds. Instead, they come in two main branches (figure 5.5). The Pseudosuchia or Crurotarsi include not only the crocodilians but also their extinct relatives, such as the armored aetosaurs and the crocodile-like phytosaurs. In addition, the largest Triassic land predators were not dinosaurs but a group known as erythrosuchids (“bloody crocodiles” in Greek), rauisuchians or sphenosuchians, which looked a bit like Komodo dragons only larger. The other branch, the Avemetatarsalia or Ornithosuchia, includes the pterosaurus, dinosaurs, and birds. These are known from a handful of Triassic fossils that then radiated quickly to produce the dinosaurs and pterosaurs in the Late Triassic.
WHENCE DINOSAURS?
Once the concept of what defines a dinosaur and an archosaur became clearer, we could look for fossils that matched this definition. The search to find actual fossils that most closely resemble the likely ancestors of dinosaurs has been ongoing for more than a century.
All the major groups of dinosaurs—the early relatives of sauropods (Plateosaurus; see chapter 6) and the earliest theropods (Coelophysis; see chapter 11) and ornithischians—are found in Upper Triassic beds in many places around the world. So the likely place to look for these fossils would be Upper Triassic or even upper Middle Triassic beds. Unfortunately, beds of this age are not exposed in most places, and most of those outcrops do not yield good terrestrial fossils. Nevertheless, a number of fossils have been suggested as close relatives of dinosaurs over the years. Primitive theropods (like Coelophysis) and ornithischians were small bipedal animals, and the early sauropods (like the prosauropods) were at least partially bipedal as well, so the likeliest ancestor of the dinosaurs was a small bipedal predator.
One of the early candidates was the small bipedal archosaur Saltopus (figure 5.6A), whose name means “hopping foot.” It was only about 80–100 centimeters (2.5–3 feet) long (counting the long tail) and weighed about 1 kilogram (2.2 pounds). Described by Friedrich von Huene in 1910 (see chapter 6), it was based on fossils from Upper Triassic Lossiemouth Quarries in the Elgin Formation of northern Scotland. It was frequently featured on the big murals that showed Triassic archosaurs and early dinosaurs by paleoartists such as Rudolph Zallinger, and it appeared in many children’s dinosaur books before the 1970s as well. More recent studies, however, have shown it to be a very primitive relative of dinosaurs, and it is not within the Dinosauria.

Figure 5.6
Early candidates for “dinosaur ancestor”: (A) Saltopus, a small bipedal predator from the Late Triassic of Germany and Scotland; (B) Marasuchus (formerly called Lagosuchus), a tiny hopping bipedal predator from the Late Triassic of Argentina; (C) Silesaurus, a larger, more quadrupedal form from the Late Triassic of Poland. ([A] and [C] Courtesy of Wikimedia Commons; [B] courtesy of N. Tamura)
In 1971, Alfred Sherwood Romer described the little fossil Lagosuchus (bunny crocodile) from the Upper Triassic Chañares Formation of Argentina (figure 5.6B). Based on the extremely incomplete remains (just a hind leg, pelvis, shoulder blade, many vertebrae, and part of the skull), it was a lightly build archosaur with very long hind legs, suggesting that it could hop like a rabbit as well as run on both four legs and two. Later researchers declared the name Lagosuchus invalid and stated that the dinosaurian features of the specimen belonged to a different taxon called Marasuchus. However, in 1975 José Bonaparte (discussed below) showed that all aspects of its anatomy are dinosaurian, and now it is considered the nearest relative of dinosaurs. A similar fossil from the same formation was described by Romer in the same 1971 paper as Lagerpeton (bunny reptile). It too is only known from scraps of the hips, hind limb, and back and tail vertebrae, but many paleontologists pointed at both the “bunny croc” and the “bunny reptile” as likely dinosaurian relatives. Today they are classified as the nearest relatives of dinosaurs (Dinosauromorpha) but are not within the Dinosauria.
Another candidate was Silesaurus and its nearest relatives (figure 5.6C), making up the family Silesauridae. The original specimen was found in the early Middle Triassic of Poland. Described by Polish paleontologist Jerzy Dzik in 2003 based on some 20 skeletons (it was one of the best known of the primitive dinosaur relatives), it was the size of a Great Dane, about 2.3 meters (7.5 feet) long because of its long tail, but not as high at the shoulders as a Great Dane. However, it was not primarily bipedal like the other candidates but quadrupedal, and its teeth suggested it was a herbivore, possibly with a beak on the toothless front part of the lower jaw. Most recent analyses place Silesaurus and its relatives (Sacisaurus from Brazil, Eucoelophysis from New Mexico, Asilisaurus from Tanzania, and Lewisuchus from the Chañares Formation of Argentina) as the nearest kin of the dinosaurs (Dinosauromorpha) but not within Dinosauria itself. So the past 40 years have produced many different fossils that are very close to being true dinosaurs, but none have all the key features that define Dinosauria.
THE VALLEY OF THE MOON
For over a century, paleontologists searched for the earliest, most primitive fossil that could truly be called a dinosaur. They looked almost every place there were good exposures of Upper Triassic beds: North America, Africa, Europe, and Asia. Yet until about 40 years ago almost nothing was known of the dinosaurs of South America. There were Jurassic and Cretaceous badlands exposures in many places in the dry eastern foothills of the Andes in Patagonia. Thanks to the clouds off the Pacific rising on the Chilean side and raining or snowing out all their moisture, by the time the Pacific winds came down the eastern side, they were stripped of their moisture by the Andean rain shadow, forming miles and miles of desert badlands largely unexplored by paleontologists. The early pioneers of South American paleontology, such as the brothers Florentino and Carlos Ameghino, focused almost exclusively on the bizarre and amazing mammals from the Cenozoic of South America and only rarely found fossils in the Mesozoic beds.
The discovery and explosive growth of research in Argentinian dinosaurs can be attributed mainly to one man, José Bonaparte. He is a distant relative of Napoleon III, but he was certainly the emperor of Argentinian paleontology for many years. Robert Bakker called him “Master of the Mesozoic.” Bonaparte is widely considered to be the founder of dinosaurian paleontology in South America. Nearly all the modern dinosaur paleontologists in Argentina were trained and mentored by him.
Bonaparte found and named more than two dozen Mesozoic genera, and 90 percent of these are still considered valid. (This contrasts with people such as Cope, who coined more names but nearly all turned out to be invalid.) His finds include predatory dinosaurs such as the bull-horned Carnotaurus and its close relative Abelisaurus, plus other predatory dinosaurs including Ligabueino, Noasaurus, Piatnitzkysaurus, and Velocisaurus. Most of the South American sauropods were found by Bonaparte (see chapter 10), including Argentinosaurus (possibly the biggest of all land animals), Andesaurus (another giant close to Argentinosaurus in size), Amargasaurus (the sauropod with spikes on its back), Agustinia (a brachiosaur), Coloradisaurus and Dinheirosaurus (from Portugal), Lapparentosaurus (from Madagascar), Mussaurus (a Triassic prosauropod ancestral to later giants), and Rayososaurus, Riojasaurus, Saltasaurus (an armored titanosaur), Volkheimeria, and Ligabuesaurus.
While finding an amazing diversity of dinosaurs, Bonaparte also discovered and documented many animals that lived alongside them. These include the peculiar pterosaur with the flamingo-like comb of delicate teeth on its lower jaw known as Pterodaustro, and the primitive flightless bird Patagopteryx. Despite his reputation for finding dinosaurs, Bonaparte prefers to study Mesozoic mammals. He found some of the only Cretaceous mammals from South America belonging to strange groups known as gondwanatheres (unique to South America and Madagascar), plus many other Mesozoic groups. Some of these mammals were previously known only in the Jurassic of the Northern Hemisphere until Bonaparte found them in Argentina.
Bonaparte was born on June 14, 1928, so he is past his ninetieth birthday as I write this chapter. I was lucky to meet him several times during my career. He was interested in my research on Jurassic mammals called dryolestoids, one of his favorite research topics. In an article in Omni from 1993, Don Lessem described him this way: “Distinguished, sixtyish, he is a man of modest proportions. His large glasses and thinning pate, polished manners, neat attire, and scholarly parlance lend him the air of an academic, which he is, by practice if not training, ten months a year.”
Bonaparte was born in the provincial town of Rosario, Argentina, and grew up in Mercedes, 100 kilometers west of Buenos Aires. His father was an Italian sailor. Most South American pioneers in paleontology had no formal training because there were no paleontologists in the country at that time, and this is true of Bonaparte. He is mostly self-trained and achieved his goals through incredible hard work. Bonaparte was inspired to collect fossils at a young age when a local retired fossil collector showed him some of his own finds. Bonaparte made collecting fossils his obsession and his passion. His house was soon full of fossils that he found in nearby riverbanks, and he created a museum in his town to house them. (They are now in the collection of the University of Tucuman.) By the late 1970s, he was awarded a job at the National Museum of Natural Science in Buenos Aires, the largest museum in Argentina, where he continues to work long after he officially retired.
Almost all the pioneers in paleontology had to work hard with very limited resources because rich donors and government agencies were not willing to fund research on something as impractical as dinosaurs. Bonaparte was incredibly focused on finding fossils no matter what the suffering and inconvenience for him and his crew. Many of his students complain that he is a hard taskmaster and can be extremely difficult. Bonaparte admits this, saying, “Yes, I can be tough, I suppose, but I work hard.” He is famous for being a workaholic, nose to the grindstone 16 hours a day, six days a week. He drives his crews just as relentlessly. It is not surprising that many of his former students have complicated feelings about their hard times with him, yet he helped launch the careers of most of Argentina’s current generation of dinosaur paleontologists.
Dinosaur paleontologist Peter Dodson said of Bonaparte, “almost single-handedly he’s responsible for Argentina becoming the sixth country in the world in kinds of dinosaurs. The United States is still first, but Bonaparte’s shown that Argentina is so rich in dinosaurs from so many time periods that it may yet top us one day.” Bob Bakker said, “We wouldn’t know anything about South America’s dinosaurs without him.” Dinosaur encyclopedia author George Olshevsky wrote, “His discoveries are fantastic. On a scale of one to ten of how strange a dinosaur could be, with a ten being the first dinosaur with wings, some of Bonaparte’s finds are a nine.”
Professional paleontologists appreciate him and his hard work. Not only has received numerous awards and honors in his native Argentina, but in 2008 he was awarded the Romer-Simpson Medal of the Society of Vertebrate Paleontology, the highest award any vertebrate paleontologist can receive (figure 5.7). It is named after Alfred S. Romer and George Gaylord Simpson, two of the greatest paleontologists of the twentieth century. In addition, both of them collected many fossils in Argentina during their careers and knew Bonaparte well during the 1930s, 1940s, and 1950s.

Figure 5.7
José Bonaparte receiving the Romer-Simpson Medal for lifetime achievement at the Society of Vertebrate Paleontology meeting in 2008. From left to right: Dr. Louis Jacobs of Southern Methodist University; Chuck Schaff of Harvard’s Museum of Comparative Zoology; Bonaparte; and Lou Taylor of the Denver Museum of Natural and Science. (Courtesy of L. H. Taylor)
THE ORIGIN OF THE DINOSAURS
In 1959, an Andean goatherd by the name of Victorino Herrera was following his flocks when he spotted fossil bones eroding out of the path. He brought in paleontologist Osvaldo Reig, who collected the specimen and described it in 1963 as Herrerasaurus ischigualastensis, in honor of the man who found it; its species name comes from the lower Upper Triassic Ischigualasto Formation where it was found. The original specimen had such a weird mixture of sauropod and theropod features that for decades no one was quite sure what kind of creature it was. Reig thought it might be a primitive allosaur or megalosaur, but in 1964 Alick Walker thought it might be a prosauropod. In 1985, Alan Charig noted similarities to both prosauropods and theropods, but Romer’s 1966 textbook, Vertebrate Paleontology, put Herrerasaurus in the prosauropods. Edwin Colbert suggested that it was related to theropods, which was supported by Bonaparte in 1970, and by some later authors. Yet Don Brinkman and Hans-Dieter Sues argued in 1987 that it has features that are primitive for all dinosaurs, including theropods, sauropods, and ornithischians; this was confirmed by Fernando Novas in 1992.
The confusion about Herrerasaurus is largely due to the incomplete nature of the original specimens. In 1988, Paul Sereno and his University of Chicago crew were working in the Ischigualasto Formation with José Bonaparte, learning the lay of the land with the master (figure 5.8). Sereno’s crew found the first complete skull and skeleton of Herrerasaurus, and finally a lot of key features that were missing from the debate were understood. One of the most famous and publicized dinosaur experts alive today, Paul and I were students together at Columbia University and the American Museum of Natural History in the early 1980s. He did one of his first research projects with me working on dwarf rhinoceroses before moving on to dinosaurs. Most authors since then have considered Herrerasaurus to be a very primitive saurischian dinosaur, with some favoring closer relationships to the sauropods, and others to the theropods.

Figure 5.8
The surreal hoodoos and deeply eroded landscape of the Ischigualasto Formation in the Valley of the Moon, Argentina. (Courtesy of Wikimedia Commons)
For one of the most primitive dinosaurs known, Herrerasaurus was clearly bipedal, but not the tiny creature that many paleontologists had expected. Adults were up to 4 meters (16 feet) long, including their very long neck and tail, and weighed about 350 kilograms (770 pounds). However, some adult specimens were only half this size, so there was enormous variability in their adult body sizes, possibly due to the differences between males and females.
Unlike many of the close relatives of dinosaurs we have just discussed, Herrerasaurus was completely bipedal with small front limbs and long, powerful running hind limbs. Like many running animals, the thighbones are relatively short and the toes elongated, with some loss of the side toes. There are stiffening features in the tail, showing that it was held out straight behind it to improve balance during running. Its hip bones did not have a large hole through the hips, but instead a bony hip socket with only a small opening (the beginning of the open hip socket seen in all other dinosaurs). Other features of the hip seem to be more like theropods.
The skull of Herrerasaurus was long and narrow, with long, recurved teeth with serrated edges, suitable for slashing prey. It had an odd flexible joint in the lower jaw that let the animal slide its jaw back and forth to give it a grasping bite, pulling the prey back after the initial biting motion. However, the skull was missing nearly all the specializations that are found in nearly all later dinosaurs, another reason that it keeps being considered a primitive relative of all the dinosaurian groups and not a true member of any one of them (figure 5.9A).

Figure 5.9
(A) The reconstruction of Herrerasaurus. (B) Skeleton of the larger Herrerasaurus and the smaller Eoraptor side by side. ([A] Courtesy of N. Tamura; [B] courtesy of Wikimedia Commons)
Herrerasaurus was a large predator by Triassic standards, and it probably could prey on most of the smaller animals found in the Ischigualasto Formation. A few skulls show marks consistent with the bites of another Herrerasaurus, so there were definitely fights among the animals in the group. However, it was not the largest predator in its time. One of the skulls shows bite marks from an animal with teeth very different from Herrerasaurus, so it was probably bitten by the huge rauisuchian archosaur Saurosuchus.
For all its confusing features, Herrerasaurus seems to have a lot of specializations and does not closely resemble the likely ancestor of all the dinosaurs. Three years after working with José Bonaparte when they found the first complete specimens of Herrerasaurus, Paul Sereno and his crew from the University of Chicago were back in the “Valley of the Moon” in the Ischigualasto beds in the austral summer of 1991. Ricardo Martinez, a University of San Juan paleontologist working with Sereno, found some tiny bones sticking out of the rocks. The specimen was very delicate, so Martinez, Sereno, and crew spent more than 12 hours carefully excavating it from its burial site and sealing it in a plaster jacket. It then spent over a year at the Field Museum in Chicago, where my good friend preparator Bill Simpson expertly removed it from its stony tomb and recovered all the bones that were there. The original specimen was on display in Chicago for a while, then returned to Argentina, and there are casts of it on display in many museums.
Finally, in 1993, Paul and his coauthors Cathy Forster, Ray Rogers, and A. Moneto published the first description of the fossil. They named it Eoraptor lunensis, “dawn raptor of the moon,” in reference to the Valley of the Moon (figure 5.9B). It made the cover of National Geographic and all the news media as the oldest and most primitive known dinosaur. It certainly is very primitive, and it is a tiny bipedal animal as well. It was only about 1 meter (3.3 feet) long, about the size of a turkey, and probably weighed about 10 kilograms (22 pounds). The long bones all have hollow shafts, so Eoraptor was very lightly built compared to the much heavier build of other primitive dinosaur relatives. The skull has relatively large eye sockets and a short snout, so it had great vision but was not built with the vicious jaws and teeth of Herrerasaurus. It lacked the sliding jaw joint seen in Herrerasaurus and many other theropod dinosaurs, which makes it more primitive in this aspect. Unlike Herrerasaurus and later theropods, only its upper teeth curved backward, another primitive feature. Its lower teeth were simple leaf-shaped structures never seen in any theropod, or in most other dinosaurs either.
Like Herrerasaurus and many of the other bipedal archosaurs close to dinosaurs, it had short thighbones and long toes, specializations for rapid running. It had large claws on its three main toes, but apparently the fourth and fifth toes were tiny or lost, as in many dinosaurs. The spool-like centra in the spine were hollow, like many close relatives of dinosaurs. However, it was more advanced than Herrerasaurus in having three vertebrae in the sacral region attached to the hip, whereas Herrerasaurus has only two as in more primitive archosaurs.
The place of Eoraptor in the dinosaur family tree is controversial. It is clearly more primitive than Herrerasaurus and many other very primitive dinosaurs, but it still has features found in theropods. When Sereno and colleagues first described it in 1993, and again in 1995, they pointed out that it was one of the most primitive dinosaurs known, but they assigned it to the Theropoda, like they did Herrerasaurus; in their words, it is closer to “the hypothetical dinosaurian condition than any other dinosaurian subgroup.” Phil Currie in 1997 thought it was closer to the common ancestor of all the dinosaurs rather than a primitive theropod. But in 2011, Ricardo Martinez, Paul Sereno, and coauthors described another primitive dinosaur from Argentina, Eodromeus, and argued that Eoraptor came out closer to prosauropods. This was disputed by Michael Benton, yet confirmed by a study by Alpadetti and coauthors in 2011. Then in 2011 Hans-Dieter Sues, Sterling Nesbitt, David Berman, and Amy Henrici argued that Eoraptor is still within the theropods and is not a primitive relative of all the dinosaurs. Two years later Sereno and coauthors reanalyzed the complete skeleton of Eoraptor and returned to the idea that it was related to sauropods. Finally, in 2017, the controversial study by Baron, Norman, and Barrett (which broke up the “Saurischia”) placed Eoraptor at the very base of the theropods.
In short, the confusing mix of features in Eoraptor means that it is very close to the ancestral condition of dinosaurs and does not clearly fall within the theropods or sauropods. Paleontologists do not expect to find a fossil that is perfectly ancestral to any group, but Eoraptor comes as close as we have to an approximation of how dinosaurs started out.
FOR FURTHER READING
Baron, Matthew G., David B. Norman, and Paul M. Barrett. “A New Hypothesis of Dinosaur Relationships and Early Dinosaur Evolution.” Nature 543 (2017): 501–506.
Benton, Michael J. “Origin and Early Evolution of Dinosaurs.” In The Complete Dinosaur, ed. James O. Farlow and M. K. Brett-Surman, 204–215. Bloomington: Indiana University Press, 1999.
Fastovsky, David, and David Weishampel. Dinosaurs: A Concise Natural History, 3rd ed. Cambridge: Cambridge University Press, 2016.
Holtz, Thomas R., Jr. Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. New York: Random House, 2011.
Lach, Will, Jonny Lambert, and Mark Norell. I Am NOT a Dinosaur! New York: Sterling Books, 2016.
Langer, Max C. “Basal Saurischia.” In The Dinosauria, 2nd ed., ed. David B. Weishampel, Peter Dodson, and Halszka Osmólska, 25–46. Berkeley: University of California Press, 2004.
Langer, Max C., Martin D. Ezcurra, Oliver W. M. Rauhut, Michael J. Benton, Fabien Knoll, Blair W. McPhee, Fernando E. Novas, Diego Pol, and Stephen L. Brusatte. “Untangling the Dinosaur Family Tree.” Nature 551 (2017): E1–E3.
Naish, Darren, and Paul M. Barrett. Dinosaurs: How They Lived and Evolved. Washington, D.C.: Smithsonian Books, 2016.
Parrish, J. Michael. “Evolution of the Archosaurs.” In The Complete Dinosaur, ed. James O. Farlow and M. K. Brett-Surman, 191–203. Bloomington: Indiana University Press, 1999.
Sues, Hans-Dieter. “Staurikosaurus and Herrerasauridae.” In The Dinosauria, 2nd ed., ed. David B. Weishampel, Peter Dodson, and Halszka Osmólska, 143–147. Berkeley: University of California Press, 1990.