 Vertebrate and Mammalian Stages in Mind: A Neuro-Theatrical Model Vertebrate and Mammalian Stages in Mind: A Neuro-Theatrical Model
Vertebrate and Mammalian Stages in Mind: A Neuro-Theatrical Model Vertebrate and Mammalian Stages in Mind: A Neuro-Theatrical ModelThis chapter combines insights from neuroscience, psychoanalysis, primatology, and evolutionary biology to show the basic elements of the brain’s inner theater, from its animal heritage to its devilish temptations and angelic aspirations. How does a sense of “Self” develop within the brain, in relation to significant others and “the Other,” the social and (at least for some) spiritual network in which the Self exists? How does this involve empathy and morality, regarding good or evil actions, and interpretations of the other’s motives? How is the inner theater of Self and Other, with conscious and unconscious agents, shaped by early childhood experiences—and our species’ evolution—toward potentially “cathartic” moments in art and everyday life?
As highly social, intelligent, and reflective animals, we make theatrical events to stage potential actions, relationships, and intentions. Not just in “theater” as an art form, but also in many other cultural situations—from childhood play to sports to adult rituals and ceremonies—humans share their inner mental worlds by staging fictional contests and symbolic performances. Such outer theaters relate to the “inner theater” (and cinematic aspects) of each person’s brain. Imaginings, dreams, fantasies, and reality perceptions are staged for the “mind’s eye”—with a sense of Self both acting and watching, along with the social Other. This also involves an inner voice of Self, or “internal monolog,” being staged at each moment, with and for the Other, like a first-or third-person “voice-over” in film.1
Yet the Self is drawn from many potential selves in relation to others in different contexts. Thus, various aspects of Self compete internally to shape one’s identity, supported by different impressions of how others might view such characters in certain situations, with confirmations and challenges coming through outward performances. One’s sense of Self as angelic, bestial, or devilish, and of others as good or evil, as friendly or threatening, depends on an inner theater—before, during, and after acting in the world.
Like but beyond other animals, such as elephants and various primates, which fondle a relative’s bones or carry an infant’s corpse,2 humans have developed an extensive awareness of death, as well as of self and other. Mourning for lost loved ones, along with the growing awareness of one’s own aging and mortality, has led humans, in various cultural ways, to posit a cosmic theater where the dead Self and others continue to exist— as souls or reincarnations, here and elsewhere. Yet, the angels and devils imagined in such a cosmic theater and depicted by artists (with bird or bat wings, on a human or serpent figure) show different super-natural extensions of our vertebrate-reptilian-mammalian or avian relatives, even prior to a scientific theory of evolution.
We usually experience an integrated sense of Self in the body— sometimes considered to be the “soul.” But according to current neuroscience, that Self is built upon the animal heritage of a proto self (assessing internal bodily states) and a core self (interacting with objects), in the homeostatic brainstem and emotional limbic system, plus an autobiographical self in the neocortex, especially in our right-brain imaging and left-brain language areas (Damasio, Self; Decety and Chaminade).3 Indeed, there are various aspects of self in the body-brain nervous system and its social extensions: a body image involving brain circuits in the parietal and frontal lobes, a passionate self with limbic system emotions, a visceral self of “gut reactions” involving the insular cortex (a deep area folded between the temporal, frontal, and parietal lobes), an executive self in the frontal lobes, a mnemonic self of memories via the hippocampus (in the temporal lobes), a vigilant self in the limbic system, thalamus, and brainstem, and a conceptual, social self, unified to some degree and involving many of these areas (Ramachandran and Blakeslee 247–54).
In the brainstem, which evolved in our vertebrate ancestors, a primal sense of Self (or ego) emerges in us, mostly below conscious awareness, through the monitoring of bodily systems and the coordination of movements, also involving the cerebellum at the base of the skull (Damasio, Self). Through emotional contagion in the mammalian core of our brain and mirror neurons in the cortex (the brain’s outer layers),4 a social “manifold” develops, involving “shared circuitry” with other people, through motions and emotions (Gallese, “Roots”; Hurley and Chater 12–13). In extreme cases, some people have “mirror touch synesthesia,” fully feeling the experiences of others, with greater empathy and less grey matter in the temporal parietal junction (TPJ), an area that distinguishes self from others (Banissy and Ward). Thus, the inhibiting of emotional contagion is needed to form distinct ideas of one’s Self, especially for competitive situations, which male primates specialize in more than females (Baron-Cohen 34, 81, 136; de Waal, Age 214–18).
Such “self–other representations” involve the right cortex differently from the left and require inhibitory circuits in the prefrontal (front part of the frontal) cortex to distinguish self from other (Decety and Chaminade; Decety and Somerville). This can also be observed in the empathic behavior of monkeys and apes: from the “state-matching” of shared moods to “concern for others” in consolation acts, to specifically “targeted helping,” which involves understanding the other’s perspective as like and unlike one’s own (de Waal, Age 209). In humans, a further degree of other and self (or group) awareness, enabling or inhibiting compassionate feelings and acts, occurs through the right and left association areas of the cortex, in its parietal and frontal lobes, and in the prefrontal lobes.
Contagious group emotions may also involve reactions “feeding back” between individuals, spreading through particular models and media images (Staub 390). This is sometimes called a “herd instinct” (by Nietzsche and Freud, for example), reflecting our mammalian ancestry, as in Eugene Ionesco’s play, Rhinoceros (1960), which shows humans turning into herd-inclined rhinos. Likewise, with human mobs and riots, destructive group behaviors may develop, involving “both contagion and noncontagion periods” (Staub 390).
And yet, destructive twists of empathic understanding can also be found with individuals. A soldier using rape to torture an enemy onlooker may understand (through cognitive empathy), but not feel (with affective empathy) the pain of the objectified victim and her husband, except as the other’s pain turns into the victimizer’s pleasure (de Waal, Age 21 1–12). Thus, an angelic aspect of the human brain, the ability to fly out of the body, imaginatively, and into the other’s viewpoint, becomes devilish—as in the fall of the Lucifer and his fellow angels in Christian depictions.
Many people today believe in a dualist sense of mind and brain, or of a soul inhabiting the body and yet extending beyond it, in this life and after it. However, most neuroscientists describe the conscious mind as emerging from and fully dependent on the living body—although it might someday be “transferable directly into computers” (Graziano 222).5 Either way, with an independent soul or a dependent mind, the brain’s mechanism for experiencing Self and Other is crucial to our current life, identity, and society.
In certain situations, such as “road rage,” any of us might experience a devilish part of the Self, triggered by physical dangers or ego threats. In more extreme cases, a raging madness might appear like devil possession—requiring exorcism or antipsychotic medication. Yet we can also cultivate the better angels of our nature in ourselves, our children, and others around us. An “empathy circuit” has been found in certain areas of the human brain (Baron-Cohen). Deficits in that circuitry, especially due to childhood traumas shaping neural networks, can produce narcissism, borderline personality disorder, or psychopathic evil. But the empathy circuit can also be developed in the other direction, toward a more compassionate morality, through therapy, artworks, and religious or secular meditation techniques.
Raised in a certain subculture by family and friends, while influenced by the larger culture through schools and mass media, children have their animal inheritance of primal brain circuits “pruned” by those environments (Solms and Turnbull 146–48). Nurturing acts, controlling punishments, tempting pleasures, and painful traumas experienced by the child, at the hands of bigger people, form the basis for beliefs in super-natural figures, as well as reality perceptions throughout life, often unconsciously. Such specific cultural prunings of inner-theater networks shape our later interactions in the theaters of everyday life and our mass social media, involving personal desires, identifications, associations, projections, and after-effects. But how do the primal drives, emotions, memories, and ideas in each of us—about self, other, and the supernatural (or super-animal)—reach a level of conscious awareness, from the brain’s inner theater to the social networks and cosmic reflections of today’s popular media, perpetuating stereotypes or provoking cathartic change?
There is no single Self, as playwright, actor, or spectator, in a central brain area, creating or watching the internal and external show (Damasio, Descartes’). Even the conscious, autobiographical self is continually revised, as a series of “multiple narrative drafts,” composed by an “audience,” driven to ask questions and form tentative answers (Dennett 14, 113, 418). There may be a shifting, functional cluster of neural networks that gives us a feeling of Self, in relation to various emotional signals from the body (Edelman and Tononi). Yet the idea and image of Self, with a will to act, is often illusory—a role played for others, in changing situations, and a mask morphing in the mirror. Experiments by Benjamin Libet in the 1980s showed that the movement of a finger involves the firing of unconscious brain circuits four-tenths of a second prior to the “conscious wanting” of movement (Wegner 52–55). The biological drives and cultural desires of Self are mostly unconscious, prior to choices being rationalized by the conscious mind and interpreted by social relations.
The physical foundations of the Self are also unconscious, through the body’s senses. For example, you do not see directly what your visual system perceives. There is a blind spot in vision where each eyeball connects with its optic nerve, which is automatically filled in by your brain. Each eye has clear vision only at its center, in the fovea, and twitches in continuous saccades to complete the visual field. The eyes’ lenses turn the visual field upside-down, with the brain’s visual system also breaking it apart into components, to then be reconstructed through memories, expectations, and fantasies (Ramachandran and Blakeslee).
Visual signals go from your eyes to the thalamus, at the core of your brain, and then to various primary visual areas at the back of your brain, in the occipital lobe. Area “V1” is sensitive to the contrast of light and dark, “V2” to color and orientation, “V3” to shape, “V4” to color, and “V5” to motion (Baars 65–66). From there, circuits extend to over 40 modules (or network hubs) of “higher” visual processing in the frontal cortex, temporal lobes, and related areas, analyzing objects, locations, faces, and much more, in relation to expectations and memories—as if the visual “actors,” as neural signals, were performing for that “audience,” and thus competing for consciousness (66).
Cognitive psychologist Bernard Baars uses this “theater” model to explain his Global Workspace Theory of consciousness, which he relates to Gerald Edelman’s theory of Neural Darwinism or “neuronal group selection.”6 As Baars argues, “All unified models of cognition today suggest some sort of unconscious audience, including unconscious memory archives and automatic routines that are triggered . . . [by] working memory” (46).7 This does not mean that a proscenium, thrust, or arena stage exists in your brain, like you might experience in an external theater. But consciousness is structured with a limited performance space in the neural circuitry of your mind,8 as an attentional “spotlight,” while supported by fringe consciousness at its edges and working memory as its “stage,” which may hold only about seven random numbers or unrelated verbal items, and fewer mental images (41–44).
The brain’s cognitive “audience” involves many specialized unconscious networks, like a vast society: automatic mechanisms for routine tasks; implicit memories for attitudes, skills, and social interactions; semantic (word) networks representing the world; declarative memories of beliefs and facts; and autobiographical memories for self-identity (Baars 43–46). This audience of memory systems, like a “legislature,” builds coalitions for and against certain players to be on the stage of consciousness, as if applauding or booing them (46–47). It shapes short-term impulses into long-term patterns of external behavior, through “working hierarchies” of conscious significance—as to which ideas and perceptions are valued on the mind’s stage.
More than 90% of brain activity is unconscious, with billions of neurons having trillions of connections (Ramachandran and Blakeslee 152; Solms and Turnbull 84). So there are numerous competing agents within your brain, behind your conscious, imaginary, and symbolic identity. Baars describes Deep Goal and Conceptual Contexts, with an unconscious “self” as agent and knower, plus executive controls (in the frontal lobes), as being like the “director” and “operators” of your brain’s theater, along with its current intentions and expectations, behind the scenes in its staging of consciousness (42–45, 144–46). Certain “players” vie for presence within the limits of the conscious stage (42). These include outer senses (seeing, hearing, feeling, tasting, smelling, warmth, vibration), inner senses (visual imagery, inner speech, dreams, imagined feelings), and ideas (imaginable ideas, verbal ideas, intuitions). Thus, the imagery we see with our eyes closed, while awake or dreaming, appears as if in an inner (movie) theater of the mind—through the brain’s staging and screening of consciousness, with perceptions both presented and blocked.
Indeed, the same brain circuits are used when perceiving reality, dreams, and willful imaginings, but with signals flowing more from top to bottom (from association areas to primary sensory areas) when there are no external stimuli (Baars 74; Kosslyn, “Einstein’s” 263; Solms and Turnbull 210–11).9 The inner theater of imagination also has an oval viewing portal, similar to what we see with our eyes open, about 45 degrees high and 120 degrees wide (Baars 73–74). Your mind’s inner eye, more like a movie camera than your external vision, can zoom in and out when viewing imagined or remembered objects. It can also rotate such objects within the mind’s theater, or pan across a scene (Kosslyn, Ghosts 128–29). Other filmic aspects are part of the brain’s inner theater, too, such as “flashbacks” (quick memory associations), cuts between angles, variable framing with closeups, temporal and spatial “chunks” of perceived scenes (Zacks 206–27), and superimpositions of fantasy and reality, such as animal-human figures.
Even with our basic perception of reality, there are theatrical and cinematic elements in the brain. Along with filling in the eyes’ blind spots (as mentioned earlier), the brain backdates and rearranges our experiences, “like an editor cutting a film,” because of a half second time-lag between perception and consciousness (Carter 26). In this sense, the brain has multiple stages moving toward the staging and screening of consciousness, “switching rapidly back and forth several times per second” (Baars 73). Cooperating and competing networks of perceptions, images, and ideas produce what you experience in each conscious moment. What appears on the main stage (or screen), through the editing cuts between substages, is based on “biological values,” involving the cerebellum and hippocampus, for example, in movement coordination, emotion modulation, and the making of new memories (Damasio, Self 72, 74). But we also “fill in” our memories and future imaginings, through the “belief-transmission game” of contagious cultural values (Gilbert 78–81, 91, 216–22).
There can be interference or augmentation between the brain’s substages. For example, when we watch a movie with the video and audio out of synch, this dissonance can jar our brain’s acceptance or “suspension of disbelief” about the screen’s fiction. Likewise, there can be conflicts between neural networks in their various sensory perceptions, imagined and remembered expectations, and verbal frameworks for conscious experience. Or they may conjoin to create dominant percepts and concepts, repressing others in the shifting stream of consciousness—as when we tap a foot while hearing music, or sing or dance with other people. Jeff Smith makes a similar point about movie music, with reference to cognitive experiments by Annabel Cohen, showing the emotional associations of a “polarizing” or “congruent” soundtrack, conflicting with or supporting abstract animated films (160). The tempo, pitch, and melody of the soundtrack (in a major or minor key) affect the viewer’s rating of the films as “calm/ agitated, sad/cheerful, villainous/heroic, or serious/humorous.” This relates to our mammalian evolution of a unique auditory system with middle-ear bones that enable greater social and emotional communication, especially between mother and offspring (Carter et al. 172).
As with theater and filmmaking, rehearsals and editing are involved in the brain’s staging/screening of percepts, concepts, and behaviors. Even with ordinary dialogue, the flow of what is heard must be cut by the brain into decipherable words. And then, turns must be taken in speaking, with ideas held back at times, in working memory, and tied to what others have just said, involving multisensory cues such as gestural mirroring and facial expressions—key theatrical elements in our “social intelligence” (Goleman, Social).10 Indeed, the human evolution of less facial hair and more facial muscles than other apes shows the value of emotional signaling with the face, as well as other body parts, to our ancestors’ mating and survival success (Greenspan and Shanker 144).
Within the brain’s theater and throughout the external theaters of everyday life, networks of interactive agents collaborate and compete, augmenting or blocking what’s conscious, individually and collectively, with unconscious “stagehands” signaling between brains, too. Consistency is usually enforced, with a sense of unity in one mind or between minds in a group. But there are, at the same time, a great variety of percepts, associations, and ideas—in the individual or social unconscious. For example, each of us has a “how” pathway in the brain, for unconscious vision and primal orienting behavior, as well as a conscious “what” pathway (Ramachandran and Blakeslee 74–79).11 Some people, with neural damage to the “what” pathway, are consciously blind, yet can see unconsciously, even when putting a letter into a narrow mail slot. Likewise, there are numerous sensory stimuli that each of our brains gets at any given moment, but only a few, if any, reach the level of consciousness. And there are many networks of inner memories, associations, fantasies, and ideas that inhibit or excite one another—to produce the players onstage that we are conscious of.
People with Charles Bonnet syndrome, including millions worldwide with black spots in vision (scotomas), due to migraines, glaucoma, cataracts, diabetic retinopathy, or macular degeneration, have brains that automatically fill in such spots with fantastic or hyperreal figures (Ramachandran and Blakeslee 87, 104–12). Likewise, amputees with “phantom limb” pain have brains that project sensations onto a missing limb, from memory networks and personal associations, while lacking the return signals from that absent part of the body image (21–58). In this way, according to neurologist V.S. Ramachandran, all of us may be hallucinating, to some degree, all the time, but also getting sensory stimuli from our external contexts that inhibit the internally generated imagery (58–62)—until we sleep and dream, or reach another state of altered consciousness through sensory deprivation or other means.
Our social networks filter or “screen” what is experienced, and then accept it or reinterpret it as meaningful, in each brain’s staging/screening of consciousness, especially through the mirror-neuron systems in and between brains. Even when we are alone, we carry that social context, or various superegos, within us. But we also have inflated, yet insecure egos that fight to maintain consistency and unity, with or against the social mirror. And we have id energies that drive us to conform or rebel. With an emotional id and a rationalizing ego at the heart of consciousness, especially through right and left cortical networks, various brain actors vie for its spotlight, as desires and fantasies of ego identity become pressured by social, superego demands.
Baars uses cognitive experiments to demonstrate the theatrical staging of consciousness, which involves unconscious contexts, goals, expectations, beliefs, intentions, and memory systems directing the percepts and concepts that vie like “players” for the “spotlight.” This relates, too, in my view, to the Freudian notion of “ego,” as conscious (and unconscious) Self, pressured by the “superego,” as absorbed social contexts, and “id,” as primal, survival, and sexual goals. The ego, or at least its conscious tip, would be a collection of the lead actors in the spotlight: the dominant ideas and sensations of the present moment, making up the Self, in relation to the audience of memories (as unconscious ego traces) and the director and operators of deep goals and conceptual contexts (as id and superego). “I am” what I think and feel now, regarding who I was and what I want in my environment—with my ego desires pressured by id needs and superego demands from the internalized Other.
According to Baars, conscious processes have a great range of contents, relating often to one another and to unconscious contexts, yet with a high priority on internal consistency and with limited processing capacity, while working in a serial fashion over time (182). Unconscious processes each have a limited range of contents, are relatively isolated and context free, and yet are much more diverse, operating in parallel with much greater capacities. Thus, in Freudian and cognitive terms, the conscious tip of the “ego,” with its priority on consistency and serial unity, yet a limited capacity, resists (and is sometimes disrupted by) the isolated, diverse, and powerful anomalies that pop up from alternative networks, as they contest the current Self’s ideas and perceptions.
Baars refers to Freud’s influence on popular thought, art, and literature of the past century, with his exploration of “unconscious processes,” but notes that Freud’s claims “could not be tested in a persuasive way” (16). And yet, in the two decades since Baars wrote this, many neuroscience experiments have confirmed, corrected, and refined Freud’s initial theories about unconscious processes—especially through the Centre for Neuro-Psychoanalysis, based in London, and its academic journal.12
Neurologist Antonio Damasio matches Freud’s final, 1938 view of the unconscious with his own today, as the unrevealed aspects of the human mind, known only through a “narrow window of consciousness” (Self 177). For Damasio, the self is a “broker” of that conscious experience, at the surface of the vast unconscious mind: “an internal and imperfectly constructed informer rather than an external, reliable observer.” Ramachandran goes further, relating his scientific observations and experiments to various Freudian defense mechanisms: denial, confabulation, repression, reaction formation, rationalization, humor, and projection, especially with the left cortex inhibiting the right (Ramachandran and Blakeslee 131–56; McGilchrist). Likewise, neuroscientists Mark Solms and Oliver Turnbull relate many psychoanalytic theories to neuroscience research, including Freud’s notion of the “ego,” as partly conscious but mostly unconscious, with rational, reality-constrained, executive functions inhibiting the animalistic drive energies of the id (286–87). They argue that each cortical hemisphere involves different aspects of ego functioning—with the bilateral prefrontal lobes also bearing the inhibitory capacity of the Freudian ego against id drives and basic emotions, which they locate in “sub-cortical structures” (104, 271–72).13
Solms thus relates the ventromedial prefrontal cortex (VMPFC) to the Freudian superego, as a “stimulus barrier” against emotional drives from the subcortical and limbic id, protecting the ego “from the incessant demands of instinctual life” (Kaplan-Solms and Solms 275–76).14 Other researchers, without using Freud’s terms, point to the VMPFC, with its ties to the emotional limbic system (MacLean’s “paleo-mammalian brain”), as a key area for linking moral cognition and moral affect (Roskies 192, 198). Damage to this area prevents the formation of new moral motives, although long-term moral knowledge is retained. The VMPFC is needed for developing “a moral sense,” with the left VMPFC most active “in people who are compassionate and caring” (Changeux et al. x–xi).15 Yet, damage to the right VMPFC, increasing the left’s dominance, produces self-protective, broadly based denials, even of obvious things, as an ego defense mechanism (Ramachandran and Blakeslee 142–43). Researchers have also found that the VMPFC is crucial for emotional associations, self-representation, reward systems, and goals, including beliefs about both religious statements and ordinary facts (Harris et al.). With VMPFC damage, there are problems in moral motivation and social decision-making, including “social exchange” issues: impulsiveness, abnormal emotions, and faux pas (Damasio et al.; Kennett and Fine 178, 185; Prinz 266–67). This suggests that the animal legacies of the human brain involve subcortical drives and limbic emotions that are inhibited by the VMPFC, as it mediates between moral cognition and affect, to shape moral motivation and compassion.
Baars also suggests such animal legacies when he adapts MacLean’s notion of the tripartite brain for his theatrical model of consciousness. He focuses on the human brainstem’s reticular formation, in the midbrain, as the “Reptilian Brain” (common to all vertebrates), which is needed for wakefulness (31–32). He describes the intralaminar nuclei, including the thalamus, at the top of the brainstem, as the “Mammalian Brain,” needed for maintaining consciousness. (In MacLean’s model, this would also include the limbic system, surrounding the top of the brainstem.) Baars views the sensory areas of the neocortex (in the top rear of your head) as the “Primate Brain,” shaping mental contents, and our greatly expanded frontal areas (in the forehead), as the “Human Frontal Cortex,” used for long-term planning, abstract thought, and voluntary executive functions (32–33). By combining Baars’s model with Solms and Turnbull’s neuro-psychoanalytic theory, plus the research mentioned earlier, we can locate the Freudian “superego” in the VMPFC, which will be explored in Chapter 5 as an inner “stage manager” or cinematographer. It is the key hub of a neural network where advanced, distinctively human areas of the frontal cortex meet the subcortical structures of the vertebrate (fish/reptilian) and mammalian brains. Here, consciousness is staged in a particular way, involving moral cognition and compassion, as crucial elements in the social dimension of ego consciousness, which also involves thalamocortical feedback loops that mediate sensory signals from the primate brain.
Baars argues that aspects of “self” are deeply rooted in the brain, involving mostly unconscious intentions and expectations, as contextual frameworks behind the scenes of our self-awareness (145). Likewise, Damasio describes the “protoself,” as unconscious existential subjectivity, “the state of the organism,” emerging from the human brainstem (Feeling 174). He locates consciousness in the brain’s “composite ‘performance space,’” which involves the “image space of the early cortical regions and upper brain stem” (Self 241).16 He maps these early cortical regions in somatosensory, occipital, and temporal areas (152). As with Baars’s director and operators of deep goal and conceptual contexts, and audience of long-term memories, Damasio finds that the brain’s conscious performance space is “continuously engineered by interactions with the dispositional space that spontaneously organizes images as a function of ongoing perception and past memories.” Damasio says that his account “compliments” Baars’s Global Workspace model and he locates the “puppeteers” of the brain’s dispositional space “in the association cortices of the frontal, temporal, and parietal sectors” (189–90). From here, the “puppet masters organize the show” in the brain’s image spaces, involving other areas of cortex and the upper brainstem. To recall an experience, or perceive a new one in relation to memory categories, the dispositional puppeteers reconstruct sensory and ideational “maps” in the image networks (140–41). Thus, the elements of an experience are not stored and accessed directly, but are reconstructed as “explicit representations” on the mind’s stage—each time the fragments of an event are re-membered.
Damasio mentions that the human fronto-insular, anterior cingulate, and dorsolateral prefrontal cortical areas have Von Economo neurons, also known as spindle cells or intuition neurons (signaling rapidly across long distances in the brain)—like the large brains of other, highly social mammals: dolphins, whales, elephants, and various apes (Self 242; Gazzaniga, Who’s 39–40). Bottlenose dolphins, killer whales, elephants, and apes also recognize themselves in a mirror, to varying degrees, demonstrating self-awareness akin to humans (de Waal, Our 193–94; Rochat 87).17 Of course, we show a much higher degree of self-reflection by creating images, words, and theatrical artworks. We also have Von Economo or “intuitive social decision making” neurons in much larger numbers, about 30 times as many as the average nonhuman ape (Gazzaniga, Who’s 39–40). And yet, Damasio defines the sensation of “self” as more basic, speculating that even reptiles and birds may have conscious minds, from proto to core self, and that various mammals may have an autobiographical self: wolves and dogs, cats, elephants, and marine mammals, as well as apes (Self 26).
The performance space of the human brain’s proto, core, and autobiographical selves, from its fish/reptilian (vertebrate) brainstem to mammalian thalamus and limbic system to advanced neocortical networks, produces a conscious “ego,” with wide-ranging access to unconscious traces, yet a limited, serial capacity for staging ideas and perceptions. It struggles to maintain an internal consistency, in relation to its outer, social networks, and its internal “id” instincts and emotions, with a “superego” function in the VMPFC as a stimulus filter, involving various defense mechanisms. Thus, the lower-level vertebrates in our evolutionary heritage, with their successful erotic and life/death drives (of reproduction and survival) have given us the legacy of remnant instincts that percolate through our brainstem and thalamus—toward the mammalian limbic system.
The mammalian drives for nurturing young and for social networking, in relation to reproduction and survival, become expressed as the “primary emotions” of joy, surprise, disgust, sadness, fear, and rage (Damasio, Feeling 50). To these Damasio adds sympathy, pride, indignation, contempt, embarrassment, shame, envy, guilt, and admiration, as “secondary emotions,” or “social emotions,” involving higher levels of social interaction in humans, along with “background emotions” or moods, on a scale from wellbeing to malaise, and calm to tense (51; Looking 45). Emotions “use the body as their theater,” through the internal milieu of visceral, vestibular, and musculoskeletal systems, and then emerge, across the threshold of consciousness, as feelings in the theater of the mind ( Feeling 51; Descartes’ 28)—as with percepts and concepts competing for the spotlight in Baars’s model.
Sensory association and higher-order cortices appraise any external, “emotionally competent stimulus” (Damasio, “Neurobiological” 51). The amygdala, VMPFC, and anterior insula also become involved in appraisal—and then trigger an emotional state. The basal forebrain, hypothalamus, brainstem nuclei, and anterior cingulate execute the emotional state in the body proper, through the central nervous system. Higher-level, social emotions, perceived as feelings, evoke innate and learned “scripts” (54). Such feelings and scripts involve intuition, reasoning, action tendencies, and the learning of stimulus-outcome links for future use. Regarding moral decisions, intuition often comes first and then is rationalized as “moral reasoning” (53). There can also be feedback to the body’s somatic markers of emotion, from the mind’s imaginary and conceptual theater of “as if” feelings, as a top-down pathway, according to Damasio. I would add that these are practiced by actors, onstage or onscreen, and spectators identifying with them as stars or characters, through alignment and allegiance (according to recent film theories).18
Psychologist Jonathan Haidt categorizes four “families” of moral emotions, which relate to Damasio’s list and his use, along with Baars, of theater metaphors. Haidt lists the “other-condemning” family: contempt, anger, and disgust (with further children, such as indignation and loathing)—which we might view as playing between the body and mind, as primary and secondary (“Moral” 855). These emotions are instinctually based with survival triggers, such as goal blockage (anger) or food danger (disgust), and yet culturally refined with aspects of reciprocal altruism, including honor or ostracism (855–58). Haidt also lists the “self-conscious” family: shame, embarrassment, and guilt—which we can see as moral feelings playing in the mind’s theater, through the brain’s staging/screening of Self, regarding bodily emotions of survival, mating, and territory (involving proprioception, property, and propriety). Haidt lists, too, the “other-suffering” family of compassion and the “other-praising” family of gratitude and elevation—which are crucial, along with the primary emotions of anger and disgust, to the “as if” feelings evoked by stage and screen performances, especially with tragicomic rasa -catharsis, as explored later in this book. (Haidt’s “moral foundations” theory will also be considered in Chapter 7.)
According to Damasio, the mind’s feelings relate, in a top-down and bottom-up way, to social, primary, and background emotions in the body, to deeper drives and motivations, to pain and pleasure reactions in the dopamine neurotransmitter system, and to the deepest homeostatic (body-state monitoring) mechanisms: immune responses, basic reflexes, and metabolic regulation (Looking 37–45). Feelings “map” bodily states, especially through the right somatosensory cortex and insula (112–17). Some of this involves mirroring the emotional expressions of others. For example, facial expressions are mirrored automatically, with signals to mimic them sent to the viewer’s facial muscles, at a level below conscious awareness (117). “False” body states can also be produced by as-if or filtered feelings, through the prefrontal lobes, as well as “actual” body states—with the brain’s body-sensing (somatosensory) areas becoming “a sort of theater” (117–18). Thus, the internal homeostasis of bodily functions extends outward, too, in another feedback loop, through pain/pleasure mechanisms, instincts, emotions, and feelings expressed theatrically by the body (by the face, gesture, and vocal tone), interacting to fit with and against others, as a “sociocultural homeostasis” (Self 27).
Figure 1.1 combines MacLean’s evolutionary triune brain, Baars’s remodeling of that with the “staging” of consciousness (into four animal-human areas), Solms and Turnbull’s Freudian terms, and Damasio’s notion of unconscious emotions of the body playing as feelings in the more conscious mind, involving dispositional “puppeteers” and sensory “image spaces,” which combine into a composite performance space. Some of the theater terms in Figure 1.1 will be explained further in Chapter 5, with other details given in Tables 5.1, 5.2, and 7.2. This also involves sociocultural homeostasis, as a cognitive, emotional, and bodily “hall of mirrors” between brains, explored in Chapter 3.
Many mammals, including apes, monkeys, wolves, and bats (plus birds), “demonstrate action programs that can be assimilated to those of human social emotions” (Damasio, “Neurobiology” 50). Neurologist Joseph LeDoux has explored specific emotional systems in the brain, especially the circuitry of fear, as a primal mammalian emotion that affects various social emotions in humans. He cites “basic emotions theorists” who argue that there are universal emotions across human cultures, which are also shared by “lower animals” (Emotional 114). But he also cites “social constructionists” who stress the different permutations of social emotions across human cultures (115). He mentions the work of Paul Ekman, which distinguishes certain facial expressions that are common worldwide from the bodily “emblems” (gestures with verbal meanings) and “illustrators” (gestures punctuating speech) that differ between cultures (LeDoux, Emotional 117).
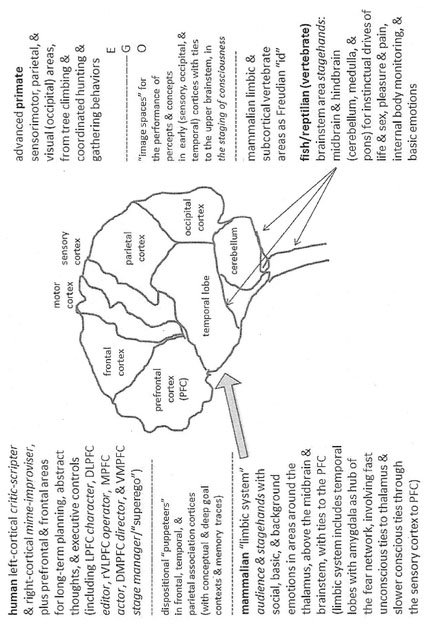
Figure 1.1 Levels, Stages, and Feedback Loops of the Animal Legacies in the Human Brain
But even “universal facial expressions” are regulated by social learning.19 They may be interrupted, diminished, amplified, or masked—through “display rules” as automatic behaviors or conscious choices (117–18).
LeDoux finds that fear and defense behaviors are similar across mammalian species, even similar to those of birds and reptiles, arguing against MacLean that early evolved brainstem (hindbrain and midbrain) mechanisms are key to our basic emotions, not just limbic and cortical structures (Emotional 97, 102–3, 171). LeDoux has explored two distinct fear circuits in the human (and mammalian) brain: a fast, unconscious “low road” from the stimulus pathways through the sensory thalamus directly to the amygdala with its signals for emotional responses and a slower “high road” through the thalamus to the sensory and prefrontal cortices, with conscious interpretation and potential control of feelings, and then to the amygdala and its signaling (164).
For example, if I am walking my dog and suddenly hear another dog bark at us, my body reacts with a startle response, which my dog might feel through the leash, increasing her reaction as well. Fear, for her, also activates an aggression circuit and she barks. But my initial shudder, with a reflexive pulling in of my arms and spinning around to see where the other dog is, turns into a reassuring tone (“It’s OK”) to my dog—when I see that the big vicious beast is behind a fence and cannot harm us. The first reaction went through the fast circuit, in my brain and my dog’s, even before I was conscious of the bark. The second went along the slower route, through my sensory association cortex and memory systems, evaluating the danger, through images and words (in the right and left hemispheres): bark, dog, big, but behind fence, not able to jump over.
The slower route, which LeDoux calls the “high road,” evolved in mammals also (and perhaps, in different ways, in birds and reptiles). But only in humans are distinctive left-cortical language areas and more complex frontal-lobe interpretations and controls involved. Thus, the primary emotion of fear, in the thalamus-amygdala circuit, engages human social emotions through the cortex, about 200 milliseconds later.20 Regarding the Baars/Damasio theatrical model, a perception of fear reaches my consciousness and has a spot-lit presence onstage, driven by somatic markers that have already occurred unconsciously: the muscle reflexes, adrenalin rush, increased heart rate, and gasp of my shudder response.
Such signals compete with my other thoughts and feelings while walking the dog. The higher, cortical route may also inhibit my body’s emotional reaction (to flee from or fight with the strange dog) like the leash and soothing tone that I use as connections to my dog and her mammalian brain. This inner feedback loop, between lower and higher amygdala circuits—tied to the social feedback loop between my brain, my dog’s, and our environment—involves deep goal and conceptual contexts, changing expectations, persistent beliefs, and long-term memories. The dispositional puppeteers (in Damasio’s terms) coordinate how I perceive, conceptualize, feel about, and consciously react to the auditory and visual stimuli, as images in my brain play in its limited space of conscious awareness. Regarding the dog-barking example, I might feel alienated as I walk on that street, think about why I am walking my dog there, recall whether I have been barked at there before, and yet persist in believing I have the right to be there—as I continue walking further and then turn around, to pass by the hostile dog again (involving other people’s potential perceptions of my race, class, and gender as well).
This example also shows how territorial drives are involved, as social and emotional expressions of biological, life and sex instincts, in humans and our fellow mammals, even when they are tamed as pets. I feel fear, and a bit of rage, then joy at survival—in relation to the more complex social emotions and territorial priorities of my cultural awareness—as I consciously realize that I am on a public street, yet near a private backyard where a big dog is barking at me from behind a fence. If a pretty lady is in that yard, too, sunning her body in a bikini, then my sexual instincts and further social emotions might be involved as well. The pleasurable lure of that perception, in my consciousness, might upstage any images of territorial danger, evoking feelings and thoughts about social fitness at that point, in the changing cultural environment, regarding my primal, core, and autobiographical selves. If her boyfriend comes to the fence, not just her dog, and we engage in a vocal or physical rivalry (like ungulate males, ramming heads in a rutting display), then the inner competition of ideas and senses in our brains will be externalized, as a conflict between us, over the rights of space and gazing. We will show our mammalian instincts toward her—as object of desire and yet spectator of us—in nature’s theater of survival fitness and sexual selection.
As highly social animals, our human ancestors evolved memory systems with triggers for unconscious fears. These affect how we interact in various social contexts, through the feedback loop of our internal theater and external performances. But emotions also create unconscious (implicit) memories, through the “amygdala system,” a different circuit than conscious (declarative or explicit) memory, which involves the limbic hippocampus and related cortical areas (LeDoux, Emotional 202). Traumatic events, coursing through the “low road” of fear via the amygdala, construct very stable networks that can be triggered later in life, as shown by animal studies of “conditioned response”—which may increase in potency due to repeated activation, like an “incubation of fear” (203). Such amygdala-system memories, involving deeper mammalian networks, affect the body as well, causing muscles to tense up, hormones to be released, and blood pressure and heart rate to change (202), as an “amygdala hijack” (Goleman, The Brain 30). While drawing on instinctual, automatic reactions, conditioned responses for fear, even in primates, are learned through family and social relationships, especially in the theater of infancy. For example, infant monkeys raised in a laboratory are not afraid of snakes—unless the mother is present when the snake appears (LeDoux, Emotional 237). But they learn more quickly to fear snakes, as ancient survival threats, than other objects.
According to Oehman’s “preparedness theory,” humans today may have a tendency to fear things that threatened our mammalian ancestors, such as snakes and insects, although with genetic variations causing some humans to be “super-prepared” for specific phobias (Oehman and Mineka; LeDoux, Emotional 237). Research has found that phobias for such primordial threats are stronger, or more resistant to “extinction” (desensitization), than a conditioned fear of flowers or modern dangers, such as guns and knives (238). Snakes and insects also prove to be stronger fear stimuli when masked from conscious awareness.
In relation to our animal heritage, LeDoux finds experimental evidence for the Freudian theory that some unconscious memories may never be accessible to conscious awareness (Emotional 239–46). The experience of stress triggers the release of hormones (adrenal steroids) that enable the mammal’s body to react quickly to threats. Mild stressors, with adrenalin effects, actually enhance memory, in a “flashbulb” effect (243). But intense stress causes the shriveling of dendrites (neural connections) in the hippocampus, impairing conscious memory functions—due to a repeated feedback loop, in which the amygdala increases hormonal output (of the pituitary and adrenal glands) while the hippocampus decreases it. With prolonged stress, the hippocampus falters and damage can be permanent. This is shown with studies of stress in mice, rats, and even tree shrews (a mammal species related to early primate evolution), when subordinates are exposed to a dominant male in the social hierarchy. With monkeys in the wild, autopsies of subordinate males reveal stomach ulcers from stress and degeneration of the hippocampus. Human survivors of trauma, such as abused children and war veterans with PTSD, have a “shrunken” hippocampus and memory deficits (242). Excess stress in humans can cause depression, which is linked to poor memory, too. Thus, traumas, prolonged stress, and depression can cause physical damage to the hippocampal system, impairing the brain’s conscious staging of emotional memories.21
Emotions are powerful, with or without ties to conscious memories. The networks of emotion from amygdala to cortex, producing feelings on the conscious stage, are much stronger than circuits in the other direction that inhibit emotions.22 LeDoux suggests that psychoanalysis “may involve the control of the amygdala by explicit knowledge through the temporal lobe memory system and other cortical areas involved in conscious awareness” (Emotional 265). But the stronger emotive players in the brain’s theater, competing for stage space against weaker, inhibitory controls in the temporal-lobe audience, show “why it is so easy for emotional information to invade our conscious thoughts.” And yet, the weaker, cortex-to-amygdala connections are still “far greater in primates than in other mammals” (303). This suggests, according to LeDoux, that we are evolving toward a greater “control” of emotions by advanced frontal lobes, or toward a better “balance” between frontal and limbic systems, with “a more harmonious integration of reason and passion.” This also relates, in my view, to theatrical notions of “catharsis” as the clarifying of emotional identifications between the audience and artwork.
Such clarification, as a fronto-limbic balance, is especially needed because our powerful mammalian amygdala (in the temporal lobes) projects to sensory areas from which it does not get input, increasing the inner imagery and effects of emotional experiences (LeDoux, Emotional 284). Extreme examples might be seen in temporal-lobe epilepsy, with a kindling of perceptual circuits making everything rich with meaning (related to orgasms and mystical visions), and in Charles Bonnet syndrome, when blind spots are filled in with hyper-real figures (Ramachandran and Blakeslee). Unconscious, emotional traces in the amygdala, projecting to current sensory areas, involve fears that extend far back into the past, even to the womb. For the amygdala reaches a high degree of maturity in the eighth month of gestation, allowing it “to associate a fear response to a stimulus prior to birth” (Cozolino, Human 56–57).
The amygdala, as limbic stagehand, also affects working memory, the stage platform for consciousness (Baars), with “strong connections to the anterior cingulate,” a partner in the frontal lobe’s “executive circuitry” (LeDoux, Emotional 285), which might be called the mind’s proscenium frame. With various ties, behind the scenes of consciousness, to the sensory cortex, hippocampus, and prefrontal cortex, emotions from the amygdala affect ongoing perception, mental imagery, attention, short-term memory, working memory, long-term memory, and higher-order thought processes (287). Mediating pleasure and pain drives, from the earlier evolved brainstem, the amygdala has ties to the orbital lobes in the prefrontal cortex just above your eyes (i.e., the VMPFC stage manager), which “may be especially involved in working memories about rewards and punishments” (285). Specifically, ties between the amygdala and orbital medial prefrontal cortex (OMPFC) have evolved in humans beyond direct danger signals, to “evaluate the reward or punishment value of highly complex social interactions” (Cozolino, Human 319). Such neural connections, revised by our species, exemplify why we strive for meta-meanings, through politics, religion, and art. With the gift and curse of our brain’s theater, with its hyper-awareness of self, Other, and mortality, we need a purpose to life—based on, yet evolving beyond the pleasures of survival and reproduction.
In humans, the basic vertebrate (fish/reptilian) brainstem instincts— toward pleasure and away from pain—take many permutations. Pleasure or its anticipation, in the dopaminergic system (with specific circuits excited by the neurotransmitter dopamine), may even unite with pain pathways, through primary and social emotions. Thus, painful punishment becomes a pleasurable reward, as with various masochistic rituals in covert places or the theaters of everyday life. With other animals, pleasure fuels healthy survival and reproductive behaviors, while pain helps with avoiding danger and learning hierarchical contexts. Animals aim for a balance between pleasure seeking and pain avoidance (Leknes and Tracey 318). But humans have evolved to be prematurely born, in comparison with other apes, due to our big brains and the narrow birth canal in females that upright walking necessitates. Humans are normally born at less than half the proportional gestation period of other apes. We should be in the womb for 18–24 months, to reach their level of physical maturity at birth (Cozolino, Human 21; Shlain 8). In our species, the theaters of family, friends, and mass media powerfully shape the infant’s developing brain outside the womb, beyond prewired instinctual behaviors for pleasure seeking and pain avoidance.23 In its first few years, the human infant has billions more neurons than later in life (Solms and Turnbull 146–47). Its brain is gradually “pruned” through cultural interactions, with nonuse causing neuronal death and activation causing growth. This rewires the brain’s circuitry through feedback from others, in the lifelong staging of desires, fantasies, and repeated behaviors—in hedonistic or masochistic directions that far outweigh basic survival needs.
Such repetition, pruning the brain circuits, occurs early in life through interactions with the child’s primary caretaker—in the neuro-aesthetic “ritualization” of baby talk (Brown and Dissanayake 46–48). With humans and other animals, ritualized behaviors alter specific actions, in a new context, to make them simplified, repeated, exaggerated, and elaborated, sometimes by manipulating expectations (47). Thus, rituals take ordinary actions and give them new meanings—as in dance, poetry, music, visual art, and theater. But primal ritualized behaviors also arise “naturally” between a mother and child, according to culturally evolved patterns. The “vocalizations, facial expressions, and head/body movements of adult caretakers” trigger an infant’s instinctual responses and vice versa. The mother communicates feelings by her “smiling, open eyes, eyebrow flash, head bob, head nod, soft undulant vocalization, touching, patting, [and] kissing” (48). This initially involves the limbic system and right cortex, more than left, in “attunement” between mother and child, which shapes the infant’s ego through the mirroring of conscious and unconscious desires (Cozolino, Human 191–96). It also involves “instrumental parenting” with adults challenging the child to gradually develop new skills, while acting like an external frontal cortex—with memories and future goals for the child (Wexler 108).
But such artistic rites in child to adult development, eventually involving left-cortex language areas and today’s virtual technologies, create an alienation from nature, as the cultural sense of self develops through the Other’s desires and demands. Often, the ritual art of family life, as an extended womb pruning neural circuitry, is not enough to make up for the loss of instinctual rules for existence, which “wild” animals still have. Although our basic needs are securely met, the animal in us wants more— as pleasure and purpose—through consumer passions, media fashions, political movements, religious ideals, and artistic transcendence.
Influencing the new field of “neuro-aesthetics,” Ramachandran has defined nine trans-cultural “laws” of art, based on the animal heritage of our neural structures: grouping, peak shift, contrast, isolation, peekaboo (perceptual problem solving), abhorrence of coincidences, orderliness, symmetry, and metaphor (200). These neuro-aesthetic laws are defined in relation to visual art, yet they also suggest aspects of perception and meaning in other art forms, including theater and film. Ramachandran argues that grouping, as meaningful in historical paintings, developed from a much earlier evolved advantage for predator and prey animals, perceiving one another despite camouflage and “cluttered scenes” (203). Fragments of an enemy, barely visible through tree leaves, but grouped into a threatening form by the brain of our ancestors, gave them an evolutionary advantage, passed on to us. Indeed, the firing of neurons in monkey brains become “synchronized” when fragments are grouped into a whole object, producing a collective effect on “the emotional core of the brain” in humans as well—to give attention, identify the form as positive or negative, and then take action (205).
Likewise, caricatures are appealing to us today because they capture “the essence” of a significant face, a function that Ramachandran relates to the Indian notion of rasa (from the ancient theater manual, Natya-Shastra), evoking a particular emotional flavor and spirit in the viewer (Ramachandran 206). Exaggerations of form and movement, as with an Indian sculpture of a goddess or nymph, “hyperactivate” mirror neurons in the superior temporal sulcus, moving the viewer with beauty and awe (208). This peak shift effect (Ramachandran’s second law) can be found with seagull chicks, too, which peck instinctively at the red spot on the mother’s yellow beak (to make her regurgitate food), but will peck even more at several red lines on a yellow stick as an abstract “superbeak” (210–11). Ramachandran argues that modernist abstract art taps into “the figural primitives of our perceptual grammar” (212). Yet, personal quirks of aesthetic preference may vary greatly. Ramachandran agrees with Freud that they are based, especially when involving sexual attractions, on “templates” that develop from family experiences and later “accidental encounters” (213).
Attention to contrast (Ramachandran’s third law) is part of the human brain’s universal grammar for art because it helps in an animal’s survival to perceive edges and boundaries, or figures against a background (219–20). Artistic isolation (fourth law) also helps the brain, especially the right parietal lobe, to focus on meaningful, emotional items in the perceptual landscape (221–23)—for the spotlight of the brain’s inner theater, in Baars’s terms. The aesthetic joy of the peekaboo effect (fifth law) stimulates the brain’s feedback networks between higher and lower visual areas in various stages of perceptual “problem solving.” This involves signals to “limbic reward structures”—with art becoming a “foreplay for the grand climax of object recognition”—when the elements in view produce cognitive and emotional meaning (228–29). Ramachandran suggests how aesthetic beauty may become different from, yet still be related to erotic pleasure, as the peekaboo effect of collective meaning becomes a more “multidimensional experience” with art, rather than just a primal reward for individual survival and sexual reproduction (230–31).
According to Ramachandran’s sixth law, abhorrence of coincidences, aesthetic interpretation “feels good” when alignments in the artwork make sense, or do not seem too coincidental if depicting a natural scene (231–33). Orderliness (his seventh law), through repetition and rhythm, also becomes pleasing through predictability (233–34). Indeed, the firing of neurons at a certain rhythm called a “gamma” range (about 40 Hz), in the brain’s theater of remembered, imagined, and perceptual representations, becomes crucial to the “binding” of neural networks—especially in relation to rhythmic actions and social networks: “rituals, chanting, dancing, drumming, and so on” (Fost 37–39). This feels valuable and meaningful through serotonin modulation of the dopamine reward system, with ties to transcendent experiences in art, religion, or temporal lobe epilepsy (40–48, 102–4; Ramachandran and Blakeslee 179–88).
And yet, art balances at times between regularity and chaos (Ramachandran 234). It moves from “familiarity” to a novel “uncertainty and ambiguity,” and then to the “resolution” of that tension (Fitch et al. 66). Fauvist paintings, for example, present a “conflict to resolve” between color and object, against conventional expectations, activating certain brain areas (Kirk 320–22). Surrealist paintings do that with object and context (321–23). In music, studies show a brain response, even without conscious awareness, to “mismatch negativity,” when a repeated note is suddenly replaced by a different note, or by “sounds deviating from a majority of tones in their timbre,” or by notes in a melody played out of tune or out of key (Tervaniemi 222–23). Thus, the conscious experience of artistic pleasure involves perceptual analyses, implicit memory integration, and cognitive mastering, “the moment at which interpretation or meaning is imposed on the artwork” (Vartanian 266–67)—or when new meanings are found in it and through it, rather than imposed upon it.24 I would call this “cognitive remastering” (akin to cognitive reappraisal): a change in human-animal, fronto-limbic networks, with a catharsis of primal and social emotions, as meaning emerges from the artwork, in the visual and acoustic experience of movies as well.
Symmetry in art (Ramachandran’s eighth universal law) may relate to evolved values of beauty, signaling health, because facial asymmetry can be a sign of parasitic infection, which reduces fertility in a potential mate (235). Even babies, a week after birth, look longer at attractive faces (Chatterjee 304). Symmetrical beauty may also be involved in collective rituals, from baby talk to public performances, along with exaggeration, repetition, and embellishment, to signify and reinforce social cohesion. It has been found that symmetry “enhances processing fluency,” helping the viewer to makes sense of the stimulus, through reward circuits involving the orbital frontal cortex (Chenier and Winkielman 281, 285). Yet the brain’s preference for symmetry applies to objects more than large-scale scenes. This may be due to different visual networks: the “what” pathway through the temporal lobes for discerning objects and features within objects, which also detects symmetry, versus the “how” pathway through the parietal lobes for relations between objects and surroundings (Ramachandran 236).
For his ninth law, metaphor, Ramachandran notices how “multiple metaphors amplify each other” in classical Indian sculpture—perhaps bridging the “translation barrier” between the left brain’s language logic and the right’s more dreamlike intuition, for a “communion” between them (237, 243). This relates also to different tendencies of male and female brains, and of Western and Eastern cultures. Left and right brain functions, through frontal and parietal ties to the limbic system, may help to explain certain aspects of “catharsis,” with complex artworks purifying the viewer’s sympathy and fear, as suggested by Aristotle about Greek tragedies, over two thousand years ago.
Catharsis has developed a bad reputation in psychology because it is often understood as simply “venting” emotions in the hope of purging them, which may increase aggressive circuitry in the brain, through what I call “cathartic backfire.” This is a risk in film, TV, and videogame viewing/ playing, as many studies have shown—with the habitual learning of violence, from screen to life, along with an addictive desensitization toward it onscreen, increasing the viewer’s appetite (Zacks 113–24). Especially through the melodramatic formula of a sympathetic hero avenging a victim (usually female) by battling an evil villain, stereotypes are confirmed and vigilante violence encouraged. Sometimes, copycat violence is triggered by such a film, even if it has an ironic anti-hero, through many factors in a spectator’s brain—such as John Hinckley’s shooting of President Reagan and others, inspired by Taxi Driver and an obsession with actress Jodie Foster, who played the avenged female victim in that film.25
And yet, the Aristotelian sense of tragic catharsis, as clarifying or purifying emotions, not simply venting or purging, might be experienced at the tragicomic edges of melodramatic movies, regarding the ancient Indian notion of rasa (refined emotional flavor), which Ramachandran mentions. Primary and social emotions may be refined in the spectator not just through the peak-shift effect, but also with metaphoric twists of left-cortical logic and right-cortical intuition, involving Ramachandran’s other laws as well. For example, symmetry may trigger erotic desire or nurturing care systems in the viewer. Yet asymmetric, bestial elements of the same character, like vampire fangs in a beautiful face, or a human body transforming into a wolf’s (with peekaboo and isolation effects also), might evoke fear and disgust. Ironic twists in the brain’s emotional systems may challenge assumptions of orderliness, involving binary stereotypes, through the scene and story.
Certain “motion pictures” will be explored in detail in the chapters ahead—in order to discover such tragicomic edges in beast-people depictions and their potential rasa effects. (Spoiler alert: endings will be given away.) One might explore each film deeply, almost frame by frame, to consider how visual compositions, rhythms, and editing cuts affect the biological universals of spectators’ inner theaters, according to Ramachandran’s laws. And his theories will be applied to prehistoric cave paintings in the next chapter. But because this book offers a comparison of many films in several genres, it will then focus mostly on narrative and dramatic elements, character imagery, and tragicomic twists in melodramatic frameworks—especially regarding how violent emotions, with the potential for cathartic backfire, might turn toward rasa-catharsis. In many of the films, a specific mix of emotional flavors will be investigated, as a rasa recipe, eliciting inner-theater networks. Examples will show ironic twists of conflicting emotions, which might evoke a shift in spectators’ brains: from right-cortical Artaudian passions to left-cortical Brechtian critiques (and back)—for more attentiveness to one’s own emotional impulses, as a rasa re finement of cognitive reappraisal and remastering.
Although this neuro-theatrical theory of ironic twists, with passionate empathy and yet critical distance, involves the modern views of Antonin Artaud and Bertolt Brecht, it is also suggested by the ancient Indian texts, the Natya-Shastra (by Bharata Muni, dated between the 400s BCE and 200s CE) and the later commentary, Abhinavabharati (by Abhinavagupta, c. 1000). According to Keijo Virtanen, rasa involves a “generalizing process of emotions, in which a spectator can at the same time participate in enduring feelings. . . and still preserve the necessary distance for understanding” (64). This includes three stages: first a fluid “melting,” then “enlargement and folding,” and then “expansion” of consciousness. Virtanen thus explains the affective to cognitive development of a rasa experience, relevant to my theory of catharsis as emotional purification and mindfulness, through left and right, prefrontal and limbic areas: “the ‘tasting’ . . . [as] the perception of one’s own awareness” (67).26 Likewise, Ramendra Kumar Sen relates both Aristotelian catharsis (with a balance of pity and fear) and traditional rasa theory to the “cure by opposites,” as a dominant technique in ancient Greek and Indian medicine (92–97), 27 akin to my neuro-theatrical theory of ironic emotional twists, with film illustrations described in the chapters ahead.
But this also requires a spectator in “the right frame of mind,” who is “budha, ‘wise,’ . . . with all senses undistracted and capable of critical observation” (Virtanen 67). The rasas involve unconscious archetypes (vasanas), which are “universal,” like Ramachandran’s neuro-aesthetic laws, as “potential functions of the mind”—evoked through the causes, effects, and transient emotions shown onstage (65–66). Savoring the rasa includes both resonance, “in the same way that the strings of a guitar will respond, untouched, with similar vibrations to those produced on another guitar,” and aesthetic distance (with painful emotions becoming pleasurable) through depersonalization and detachment, toward “Pure Consciousness” (Maillard 35–37; Virtanen 64, 70). Rasa is also defined as a skillful blend onstage of a particular emotion’s determinants (immediate causes in real life), consequents (direct effects), and transitory states (fleeting or secondary manifestations)—like the flavors in a cooked dish (Maillard 33–34). This paradoxical progress of rasa, through sensual pleasure to its resonant source and yet also toward transcendence of it, 28 sounds more like Artaud’s theater of cruelty, with actor-audience communion, than Brecht’s epic theater (influenced by European Enlightenment ideals), with his goal of social change through historical parallels in the plot, involving an actor’s gestures onstage to provoke ironic questions about character and context. Yet I would include both of these modern views in a neuro-theatrical theory of rasa catharsis, with potential, personal and social effects today. The degree of Artaudian or Brechtian results in the audience would depend on the plot, character, and emotional twists onscreen (or onstage) and the spectators’ various inner-theater attitudes and associations—toward a particular form of shared awareness with changes occurring then in the performances of everyday life.
The eight emotions (bhavas) of ancient Indian theater, with their related rasas, include Aristotle’s two cathartic feelings, tragic sympathy (or sorrow) and fear, but add six more, variously translated as love/happiness/ delight, humor/mirth, anger/wrath, heroic vigor/courage, disgust, and wonder/awe. 29 According to the Abhinavabharati, the goal of Sanskrit drama is to display a certain dominant bhava (emotion or mental state), as well as secondary ones, through specific facial expressions, hand and body gestures, and the story (Goodwin 177–82). This evokes the corresponding rasa in spectators 30 and moves them toward union with Brahman (the Oversoul), through tanmayibhavana (universal communion). Influenced by Buddhism, with calmness as the goal of Enlightenment, beyond sensual attachments, the Abhinavabharati names a ninth rasa, “peace” (shanta), as the source and destination of the eight (Byrski 176–77, 186).31
Plays were defined in ancient Greece as a tragedy, comedy, or farce (satyr play), but in India through the eight bhavas and their corresponding rasas—with peaceful Enlightenment, beyond emotional ego pleasures, as the cathartic goal.32 Specific facial expressions, poses, and gestures were used in ancient India to convey each bhava and evoke its rasa in the viewers (along with other ideas through a coded sign language), a tradition that continues with various dance-dramas today, such as Kutiyattam, Bharata-natyam, and Kathakali, which have also influenced Bollywood movie musicals. A recent experiment with American and Indian college students showed that recognition of such coded facial expressions for the bhavas may be transhistorical and cross-cultural (Hejmadi et al.).
Current neuroscience focuses on a similar array of six primal emotions: sadness, fear, anger, happiness, surprise, and disgust, according to Damasio’s list mentioned earlier (comparable to Panksepp’s list considered later). Damasio’s further list of social emotions—sympathy, pride, indignation, shame, envy, guilt, and admiration—also relates to the bhavas of courage and awe, as well as to Aristotle’s example of Oedipus evoking admiration and yet sympathetic shame, through the tragic flaw of hubris in Sophocles’ play. (Recall that awe is involved, too, in Ramachandran’s peak shift effect, with the hyperactivation of mirror neurons in the upper temporal lobes.) Like the ancient Greek practice of performing three tragedies and then a satirical satyr play, all in one day, Indian Sanskrit dramas, even if focusing on sorrow, fear, or disgust, were always striving “to end on a note of fulfillment and happiness” (Bhat 32–33).33 But the same might be said of most movies today, which may have tragicomic twists but rarely end as tragedies, whether from Hollywood, Bollywood, or elsewhere. That final note of fulfillment and happiness, along with a refinement of emotional tastes throughout the show, creates a potential catharsis of spectators’ inner theaters, altering brainstem-limbic-parietal-frontal, right and left cortical, animal-human networks, including personal and cultural memories, within and between brains.
Most humans around the globe, especially in the “First World,” have developed technologies of survival that make the animal necessities of food, water, and safety easy to obtain. Ironically, however, our higher-order consciousness, language, and technology makes us more aware of death’s inevitability, more alienated and vulnerable to depression, while stressed by the demands and distractions of modern life and mass-media marketing. The human ego extends our survival and reproduction drives far beyond biological concerns. Our premature birth, even as normal humans, along with our self-awareness, means that our ego is often at risk—as it performs for others. The inflated, insecure, human ego draws on pleasure/pain mechanisms and emotions, from the proto to core to autobiographical self, in the brainstem, limbic, and cortical systems. The amygdala’s fear circuits, fast and slow, unconscious and conscious, may be engaged even when our bodies are not in danger—but our egos are. Social fitness becomes more crucial than environmental adaptation, because culture is our dominant environment. We need to belong and have purpose in our lives, not just physically survive. Reproducing parts of our identities, even without offspring, we influence others and leave symbolic yet material traces of our egos with them, through monuments, artworks, and daily interactions with their neural networks, beyond our body’s mortality.34 Thus, the current cultural evolution of consumer desires, artistic challenges, and virtual media powers builds upon the neural scripts of our vertebrate and mammalian, as well as later human ancestors. But what climax do they seek, and what cathartic effects do our animal scripts produce, in and between our brain theaters?
As members of a complex social group, young mammals play, in a “rough and tumble” way, practicing competitive and collaborative skills (Panksepp, Affective 284–86). Humans go much farther with childhood and adult sports, theater, cinema, television, videogames, and other mass media, fashioning their fear and rage drives in socially acceptable ways. Even our dreams, with a mechanism and perhaps some contents inherited from our mammalian ancestors, may be a threat perception and avoidance “rehearsal” system for relating primal survival networks in the brain to the current cultural environment (Revonsuo). This helps to explain the prevalence of wild animals in children’s dreams today (or the wolf I saw, as a child, in my bedroom window while awake), as inherited threat figures in the brain’s imaginary theater, even if they are not encountered in everyday life, except in zoos or nature shows onscreen.35 Such dream figures eventually shift toward modern threats as the child matures. Research also shows that dreams consolidate new skills and emotional experiences from recent daytime events, integrating them with long-term memories, although in a world of much greater imaginative flexibility, where “the normal waking rules don’t apply” (Linden 210–20). This encourages creativity in waking life, especially in the realms of art and entertainment, yet that also hinges upon basic survival and reproduction scripts.
Today, we have multiple, proliferating, live and screened, fictional and “news” media. Each of these dreamlike theaters stages a particular collective consciousness—with extensive social networks, yet a limited spotlight of attention, like the staging/screening of consciousness in our brains. Through these theaters, including online social media, we present idealized and demonized ego-images of ourselves and others, as beautiful or grotesque, proper or perverse characters, in pleasurable or threatening scenes. The “superego” that we absorb through such media may demand enjoyment, even more than moral obedience, in perverse twists of pleasure and pain, to perpetuate endless desires and marketing lures (Zizek, Enjoy and For). “Be patriotic and go shopping,” Americans were told by President Bush after 9/11. But the pressure of “shop till you drop,” like the War on Terror, never seems to end—and increases during certain seasons as a super-natural rite.
Behavioral economists have demonstrated that people will bid up the price of a product, such as a coffee mug, higher than they initially offered for it, once they have been given it for free and owned it for an hour. This increases its value to them, as a gift and possession, along with the desire of others for it. People will even bid $28 for a $20 bill, in an auction of it, competing with others’ desires. And experimental subjects will overbid on a stock in a collective computer game—even when told that its value will automatically drop, at each step in time, toward the game’s end. Thus, our inflated, insecure egos, drawing on primal emotions of fear and rage, through competitive feelings of identity and greed, create “bubbles” in the marketplace, far beyond survival needs (or sexual reproduction instincts). Yet the collapse of such a bubble causes real survival problems, as well as ego crises and economic depressions.
“Panic” in the stock market, like the infectious threat of terrorism or an individual’s symptomatic anxiety, draws on a particular circuitry in the brain that we have inherited from our mammalian and earlier ancestors. Experiments with rat pups have shown that, when separated from their mother for a few minutes in the first two weeks of life, their vocalizing increases until she returns, triggering her nurturing behavior of licking, grooming, and carrying them (Kandel, “Biology” 514). Such consistent maternal care in infancy lowers the stress response (measured by the production of glucocorticoid hormones) in the pup’s brain, throughout its life, reducing fearfulness and stress-related disease. Panic in the pup, or “separation-distress,” with instinctual calls for mom, increases the likelihood of the mother’s better “attachment behavior” and the pup’s future survival, even with fearful situations. Likewise, the panic system in our brains serves an important purpose, as mammals that are helpless at birth and very dependent on nurturing throughout infancy.
Newborn monkeys that are isolated from their mother for the first six to twelve months of life act “like severely disturbed or autistic children,” even when returned to the company of other monkeys, though the same procedure with older monkeys has little effect (Kandel, “Biology” 513). And as humans, we may have a higher susceptibility to panic than other primates, given our normal prematurity at birth, especially those of us who have suffered the absence of maternal care at critical points in development. The death of a parent, for example, places human children “at higher short-term risk of major depression, anxiety disorder, PTSD, conduct problems, and impaired academic and social competence,” plus long-term risk of major depression and anxiety in adulthood, although quality care giving after the loss may reduce that risk (Luecken et al. 310). Attachment trauma in childhood, “from abuse, neglect, or profound misattunement between parent and child,” may also produce “borderline personality disorder” later in life, with terror triggered by real or imagined abandonment, “similar to what any young primate experiences when physically abandoned by its mother” (Cozolino, Human 256–57). On the other hand, a study showed that infants in a neonatal ward, born more prematurely than normal, who were touched and stroked by caregivers (like the rat pups being groomed and licked) for just 15 minutes, three times a day, grew 50% faster, were more active and alert, and matured faster in behaviors than those who got the same medical care but were not touched (Sapolsky 95–96).
Prolonged separation of the rat mother and pup (three to six hours per day for two weeks) causes her to ignore the pup, increasing the pup’s glucocorticoid response to stress throughout its life. During such separation, after calling to the mother for a period of time, the pup will then withdraw into a physically inert state, which appears like human depression (Solms and Turnbull 131).36 Instinctually, this may help it survive, by hiding from predators, until the mother finds it. The panic or “separation-distress” system in human brains operates in a similar way (129–30). It is centered in the anterior cingulate (at the top of the limbic system) with ties to other areas involved in maternal and sexual behavior in mammals. Thus, it is “intimately connected with social bonding,” as well as par enting. When the human infant is separated from its mother, it first expresses anxiety and anger, shown by clinging, searching, and crying, like the rat pup. It then shifts to sadness and despair, instinctively “conserving energy and withdrawing from danger” (Kandel, “Biology” 514).
Individual anxiety in adult humans, or collective panic with the stock market or terrorism, draws on the same mammalian panic system, but adds the “bubble” effect of a particular ego and culture. Deep survival goals direct the amygdala’s quick, unconscious fear responses, amplifying phobic objects. This produces images and ideas on the cortical stage of consciousness (the higher fear route) through offstage contexts: long-term beliefs and prejudices, immediate expectations, and personal memory traces—anticipating further threats in the media’s collective dream theater toward the bubble’s eventual collapse.37
Jaak Panksepp has researched the neural circuits of primary emotions in rats and other mammals, to find the corresponding circuits in the human brain for its panic/grief, fear, rage, and seeking systems, including lust, care, and play subsystems.38 The panic/grief circuits are concentrated in the anterior cingulate,39 fear and rage circuits in the lateral and medial temporal lobes, and seeking in the ventromedial frontal lobes ( Affective 316). Yet fear and rage are tied to our fish/reptilian heritage and seeking relates us to invertebrates (Davis and Panksepp 1948; Huber et al.). Indeed, all of these networks converge on emotional and self-representation zones in the human brainstem: the periaqueductal gray (PAG) area of the midbrain. Thus, Panksepp offers neural correlates for the rasas of sorrow, fear, and rage, as well as the play impulse of theater and movie making. He also helps to distinguish the lust and care networks of the brain, in relation to primal reproductive drives, which are combined in the rasa of romantic love.
Solms and Turnbull relate Panksepp’s four “primitive instinctual mechanisms” (panic/grief, fear, rage, and seeking) to the Freudian “id,”
Table 1.1
Modern Social Neuroscience and Ancient Theatre Theories of Emotions
| Jaak Panksepp's research on emotion networks in human brains, as related to animals: |
| • seeking (from invertebrate to fish and reptilian brains) |
| • fear, rage, and lust (in reptiles and mammals) |
| • sadness/grief/panic, caring, and joyful play (just in mammals as highly social emotions) |
| [These involve stimulus-memory-prediction patterns that become hyper-theatrical in us.] |
| Antonio Damasio's list (including some of Panksepp's terms, shown in bold): |
| • background emotions (moods): well-being to malaise, calm to stress |
| • primal or primary emotions: joy, surprise, disgust, sadness, fear, and rage |
| • social or secondary feelings: sympathy, pride, indignation, shame, envy, guilt, & admiration |
| Jonathan Haidt's "families" of moral emotions (with Panksepp's terms in bold, Damasio's in italics): |
| • the "other-condemning" family: contempt, anger (rage), and disgust |
| • the "self-conscious" family: shame, embarrassment, and guilt |
| • the "other-suffering" family: compassion (related to sympathy and caring) |
| • the "other-praising" family: gratitude and elevation (admiration) |
| [These are from Haidt, "Moral." But see also Table 7.3 in chapter 7.] |
| Aristotle's ancient Greek theory of katharsis as purifying certain emotions (through plot, character, ideas, language, sound effects, and spectacle): |
| • tragic sympathy/caring and fear in the audience, through the hero's seeking and conflicts |
| eight ancient Indian bhavas (emotions), each with a specific rasa (a flavor evoked by art): |
| • tragic sadness/sympathy/caring (as in Aristotle's theory) |
| • fear (as in Aristotle's theory) |
| • anger (or rage) |
| • erotic/romantic love or happiness (involving lust & caring) |
| • humor (joyful play) |
| • courage or heroic vigor (with obstacles to the hero's seeking, in Aristotle's model) |
| • awe (suggested by Aristotle with the audience's admiration for the tragic hero) |
| • disgust (indicated, too, by Aristotle's notion of the hero's tragic flaw or hamartia) |
| & the ninth rasa of peace (shanta), as the goal of mindful attentiveness to such feelings [union with Brahman (the Over-soul), through tanmayibhavana (universal communion), as influenced by Buddhism, with calmness as the goal of Enlightenment, beyond sensual attachments, according to the Abhinavabharati, regarding the earlier Natya-Shastra] |
involving vertebrate to mammalian levels of self and emotions (137). They relate frontal-lobe mechanisms to the Freudian “ego,” especially in the ventromedial and orbitofrontal areas, involving “voluntary programming, regulation, and verification of action,” which distinguish humans from other animals, and adults from children (136–37). In their model, the ego of the frontal lobes is in a continual balancing act with the id of the limbic system and brainstem (especially the seeking, rage, fear, and panic circuits), mediated by the superego of the ventromedial prefrontal cortex (VMPFC). They associate the id with Damasio’s notion of the “core self” and ego with the “autobiographical self,” yet remind us that only a small part of the ego is conscious (97).40 They say the “aim” of psychotherapy (and of artistic “catharsis,” I would add) is to “ extend the functional sphere of influence of the prefrontal lobes ” (287–88). Such a material change in the brain, in precisely that area, has been demonstrated with functional-imaging studies (288). Studies also show that depressed patients, successfully treated with psychotherapy, have achieved “metabolic normalization in their frontal and temporal lobes” (Cozolino, Human 337). Patients with anxiety disorders, thus treated, “have reduced activation in cortical and subcortical areas.” Therapeutic and theatrical catharsis might change not just our conscious ideas, but also how consciousness itself is staged by unconscious agents, including our mammalian, emotional networks— thus increasing the reflective awareness of certain feelings.
Following Damasio, Solms and Turnbull locate the “content” of consciousness in posterior cortical association areas (the advanced primate brain within us), monitoring the outside world, and the “state” of consciousness in “the ascending activating system of the brainstem” (our vertebrate, fish/reptilian inheritance), which monitors the body’s interior (90). The evolution from animal to human consciousness, with the brain representing thoughts, not just inner and outer sensations, through complex, abstract, and internally generated images, means that our conscious state “is generated by a virtual body.”41 Human consciousness, with its mammalian heritage, “consists of feelings (evaluations) projected onto what is happening around us” (92). Such projections are increased by “repression,” in the Freudian sense, which removes certain experiences from conscious memory access, but allows primary “id” processes, with “compulsive, stereotyped” emotional networks, to return with a force beyond our secondary, rational, ego controls (286–87), as LeDoux shows with fear circuits. Here again, the inner extension of prefrontal networks, through therapeutic awareness and artistic catharsis, may help humans to better integrate the brain’s various animal and cultural mechanisms—across the staging of proto-self to core-self to autobiographical consciousness.
Such a catharsis (as the tragicomic clarifying of emotions) may provide an antidote for addictive ideals of inflated ego power in our mass media. These power ideals often rationalize compulsive stereotypes and justify the quick reactions of fear and rage circuits (or the bhavas of fear, anger, and heroic vigor) in film, television, videogame, and news-story melodramas of heroes versus villains. Extending subjective awareness about our animal emotions, by questioning particular cultural fantasies, could counter the ecstatic seeking of money and fame, by investors and self-promoters, from Wall Street to “reality TV,” Facebook, Twitter, and YouTube. For these drives repeatedly produce economic and social-media bubbles, which become like Pandora’s Boxes, spreading panic and formerly repressed feelings when the bubbles collapse.
Akin to the cultural seeking of money and fame, the biological “seeking system” that Panksepp maps, in rat and human brains, is also known as a “foraging/expectancy system,” and more generally as an “incentive or appetitive motivational system,” which involves “‘wanting’ as opposed to ‘liking’” (Affective 145). It is associated with curiosity, interest, and expectations—or the anticipation of a reward, even before getting it (Solms and Turnbull 115). Reward anticipation is also related to trust, especially through activity in the caudate nucleus (in the striatum, near the top of the brainstem), as shown in a neuro-economic study, involving an investment game (Lone Frank 196–97). Also, women tend to have a longer activation of expectation-reward areas than men (198), perhaps through their evolutionary role in child raising, with long-term trust and delayed rewards in the seeking of a mate and security.
Like Freud’s notion of “libido” and “libidinal drive,” in the animal heritage of our brains, the seeking system involves survival and reproduction instincts: foraging for food and water, plus sexual interests (Solms and Turnbull 116–17). There are internal “need-detector mechanisms” in the hypothalamus (in the brainstem, just under the thalamus), sampling for hunger, thirst, and sexual needs (117). These, as well as perceptual and cognitive triggers, activate the seeking system and its bodily behaviors (118). But in humans, the drive for food, water, or sex is not fully regulated by instincts, or restricted to healthy limits, as in other animals.42
The brain’s seeking system operates in either an “on” or “off” mode, as part of the mesocortical-mesolimbic dopamine system (Solms and Turnbull 116–19).43 It relies on memory networks to stage specific desires and actions. Dopaminergic synapses perform like “gatekeepers,” fueling desires from unconscious stages to the conscious or obsessive spotlight (Panksepp, Affective 144). When abundantly active, they make a person feel “as if he or she can do anything.” Over-activation of the seeking system also produces psychotic delusions, which can be addictively triggered by cocaine and amphetamines. Antipsychotic medication thus lowers the dopamine level. But L-dopa raises it, in the treatment of Parkinson’s disease, which can also produce psychotic symptoms. So dopamine at the right level is crucial for “people and animals to operate smoothly and efficiently in all of their day-to-day pursuits.”
As experiments with rats show, dopamine levels increase in the brain with its correct anticipation of a pleasurable reward, even more than with the reward itself, but dopamine activity also increases with an unexpected reward (Politser 88–89). As learning progresses and the stimulus-reward association becomes reliable, there is a “fast-acting subthalamic loop connecting to dopamine neurons in the striatum” (in the basal ganglia around the top of the brainstem), producing the pleasurable “expectation component of the predicted reward” (94).44 This triggers excitement, through connections to the hypothalamus (just under the thalamus at the top of the brainstem). But that is also inhibited by another circuit when the reward becomes more predictable (95). Thus, the seeking system has an expectation-reward component, with a “plastic” (highly flexible) learning mode, yet it also connects with a “hardwired” habit network, “engaged by repeated rewards” (124). With this network engaged, certain rites of seeking and consuming become driven by habit, even when involving pain or “repeated punishment,” since “cells in the dorsal striatum that modulate action often resist adaptation.” This may still serve an evolutionary purpose, since “habits can be precise, reliable, and economical choice mechanisms. Yet, they are prone to bias.”
As in the neuro-aesthetic models considered above, a tension between familiarity and novelty is shown in this neuro-economic view, which may increase the brain’s dopaminergic engagement in learning, or be countered by the neural rutting of habit. And yet, through artistic catharsis, the artist’s and viewer’s inner theaters might break the habitual networks of unconscious fantasies that produce repeated desires and actions. Such reevaluation and dopaminergic excitement, as adaptive cultural learning, also involves the brain’s prefrontal lobes with memory networks in the orbital, ventromedial, frontomedial, and dorsolateral areas (Politser 96–97).
Likewise, in the neuro-economic “actor-critic” model, a critic (or stage manager) in the brain “updates the expectations of reward and reevaluates policies according to experience,” which may relate to “learning signals encoded in dopamine neurons” (Politser 100). The actor in this system then “binds the dopamine-mediated predictions of the hedonic [pleasure] value of stimuli to the sequences of acts needed to obtain them.” Again, the prefrontal cortex is involved with representing and perhaps changing current goals. Serotonin may also play a role in focusing this actor-critic, learning mechanism, with dopamine involved at the “wanting” level. Expectations and goals, from critic to actor, then connect with a “comparator” in the brain, which functions like “an ‘audience reaction’ to the ‘actor’s’ play”—producing a sense of “liking” when the desired object is consumed or obtained (101). And yet, habitual actions may result when the brain’s “automatic pilot” overrides the actor-critic-audience’s appetite for novelty (102).
I would argue that today’s marketing lures, with ego-survival and sex-drive displays, from magazine ads and billboards to TV commercials and Web interactions, may stimulate our dopamine levels and seeking systems to high levels.45 This may have effects in our brain like cocaine and amphetamines, to some degree, which “produce a great burst of energy and increased interest in objects in the world” (Solms and Turnbull 206). Events in the world have a special significance and yet arouse suspicions (206–7). At higher doses, these drugs (or the internally produced dopamine levels) cause a “stimulant psychosis,” with patients becoming paranoid and experiencing auditory hallucinations. As teenagers, we are especially susceptible to this, since our prefrontal networks do not mature until our mid-twenties (282; Cozolino, Human 69–72).
At all ages, we may become habitually addicted to the hallucinatory images and energies in our evermore virtual, paranoid-schizoid, and mass-mediated world. Our fear and seeking systems may need more and more stimulation for excitement to be felt—and we may suffer withdrawal pains if we turn away from the movie, TV, laptop, and handheld screens. Yet, that appears to be the collective environment for most of us in the “developed world.” It has become normal for us to consume mass-media rewards, and sacrifice our survival energies to get them, for our dopamine fix, with other medications prescribed to modify the side effects. Our brains, for better or worse, are changing in this hyper-screen environment. But a re-evaluation is still possible, through traditional art forms and these newer media, involving our brain’s panic, fear, rage, and seeking/lust systems (and perhaps other bhavas) toward a greater awareness of emotional triggers and habitual practices—with cathartic insights about the current cultural twists in our animal natures.
We inherit a “cold” type of aggression from our predatory ancestors, aligned with the seeking system and its drive for food and sex (Solms and Turnbull 124). This is distinct from the “hot” aggression or rage system, but both may be evoked by the competitive marketplace of our consumer culture and geopolitical news media. These animal systems in our brains also relate to a third type of aggression (in the lust system), involving male dominance behavior, territoriality, and social emotions, which are significant, too, in our mass-mediated environment. In many mammal species, males are larger than females and they fight for dominance, especially for mating rights during breeding times, when testosterone levels are high. But with humans, unlike most animals, females have evolved a hidden ovulation—not displaying when they are in heat, ready to breed.46 Humans compete for breeding possibilities beyond seasonal, instinctual rules, using cultural paradigms that sublimate such aggression, selection, and pleasure principles into many other fields. With fluctuating hormonal levels, humans compete for dominance, or to be desired objects, in numerous arenas and rites, such as sports, politics, popular entertainment, marketing, and even religion.
The rage system, involving hot aggression, is triggered when a goal is frustrated. This may be specific as in fighting for an object, such as food, or abstract as in fighting for an idea, like “freedom.” Even a human baby will express rage, struggling for freedom without knowing the word, when its arms are held at its sides. When a goal of the seeking system is thwarted, “neural patterns of frustration” in the frontal cortex send downward signals to rage circuits, in the temporal lobes, amydgala, and brainstem (Panksepp, Affective 189). Our brains are thus prepared by evolution to project “the cause of anger” upon an external source, to blame others and fight with them, unless higher cognitive appraisals are made (189–90).
This is often expressed in stage and screen melodramas (especially action, horror, and sci-fi/fantasy movies with monsters and slayers) with a “villain” blocking the hero’s goal and thus becoming the object of the hero’s righteous rage. In horror films, there is also a shift from panic/grief, with paranoid uncertainty about danger, to focused fear and courage. The cold, rational aggressiveness of goal seeking turns into hot wrath, with vengeful battles for dominance and territory, evoking similar rage and pleasure-lust networks in spectators’ inner theaters as they cheer the hero’s violence. Projecting the cause of the hero’s and spectator’s fear/anger onto the villain, while a great formula for selling tickets, along with violent spectacle and an emotional soundtrack, may confirm stereotypes represented by that monstrous antagonist. This may encourage preemptive strikes or vengeful acts, by vigilantes and governments, against “terrorists” in real life—as threats like vampires, werewolves, or beast-people to the hero’s allies and loved ones. But when heroes and villains are presented with a tragicomic mix of intimacy and distancing effects, shifting audience networks between cold, hot, and lustful aggression, or between paradoxical bhavas of fear, grief/compassion, and humor,47 heroic vigor and disgust, or (especially with vampires) awe, love, and disgust, a more complex catharsis might result, toward the peaceful rasa of reflective emotional awareness—as examples in Chapters 4 and 6 may show. This potentially positive cathartic effect of violent movies, involving slayers and monsters, also comes with a contagious danger, however, of spectators extending melodramatic identifications and emotions toward real-life stressors as fear and rage triggers, drawing on memory networks of personal and cultural traumas.
Studies show that persistent stress activates rage in rats, with an increase of the neurotransmitters dopamine and norepinephrine (which triggers the hormone adrenaline), especially through a “trauma-sensitized” alarm system (Niehoff 126–33). The rage circuit extends, like fear, from the amygdala through the hypothalamus to the PAG (Solms and Turnbull 124–26).48 Rage also involves hormone signals between the body and brain, through the HPA (hypothalamus-pituitary-adrenal) axis, particularly testosterone in male rodent, primate, or human status seeking, which can lead to dominance battles, and estrogen in both men and women (Niehoff 157–59, 171–73). The difference between rage and fear, triggering fight or flight instincts, is determined by interactions between the medial and lateral central parts of the amygdala (Solms and Turnbull 126).
Even at low levels, activation of the rage system can cause irritability, priming us for aggressive outbursts—with our mammalian body shifting from chronic scowling and muscle tension to grimacing with teeth bared, mouth growling, and fists (or claws) extended. This benefitted our animal ancestors with automatic action routines for surviving violent encounters. But the frequent frustrations of modern life, with multiple consumer desires and work anxieties, may prime us toward quick anger, like with the quick fear route, between sensory areas and the amygdala. This is shown, for example, in “road rage” or with holiday mobs in shopping malls. And yet, if we realize the signs of our body changing, with feelings rising in us, such as fear or anger, we can activate prefrontal networks inhibiting automatic responses—even when faced with the emotional contagion of someone else’s rage: “Is my survival really threatened or just my ego?”
Neuropsychologist Louis Cozolino encourages us to see such choices in the long-range view of evolution, especially regarding two tendencies that we inherit as a species. We are primed by Mother Nature to project our flaws: “to attribute our own unconscious processes, feelings, and motives to those around us” (Human 341), thus battling inner demons by seeing them in others. Also, our brain’s power for “abstraction, conceptualization, and imagining the future”—along with the stressors of our individualistic, competitive culture and advanced, multitasking technologies—increases our anxieties today, giving us chronic stress and irritability. Anxiety and stress are evolved mechanisms for survival, yet they have dangerous consequences in our brains and bodies when chronically activated.
Ironically, through the success of our species, we often seek such chronic experiences in daily life, or in dreams and the mass media, as if continuing to rehearse modes of bestial survival, through “omnivorous consumerism and thrill-seeking behavior” (Cozolino, Human 342).49 And yet, we may now be on an “evolutionary frontier,” as we change our neural wiring and social networks (341)—perhaps through the rasa-catharsis of mindful attentiveness, practiced in theater and film viewing. Such cultural and neural changes involve the primal emotions of panic, fear, rage, lust, and seeking, plus the social emotions of care, sadness, and joyful play. But these also connect with the left and right cortical hemispheres in different ways, relating to the evolved specialties of males and females, from our hunter-gatherer ancestry, for hundreds of thousands of years—and the primate, mammalian, and vertebrate structures of our inner theaters, which took millions of years to evolve.
There was a bottleneck in human evolution 70,000 years ago, possibly due to climate swings (after eruptions of the Toba volcano, in Sumatra, Indonesia), which could have reduced the human population to 10,000 adults, or as low as 1,000 breeding pairs, and selected for a more flexible brain to survive the extended periods of cold and drought (Ambrose; “Supervolcano”). Although there were many earlier migrations of hominins out of Africa, humans on other continents today are descended from a relatively small group that left around that time, 70,000–60,000 years ago. They spread across the world, even to the northern climates of Eurasia during the Ice Age. (Neanderthals evolved in northern Europe and Asia separately from humans, from a common African ancestor, and survived there until 30,000 years ago, yet also mixed genetically with our species.) Something changed in our ancestors’ brains—enabling their survival through the climate bottleneck and their reproduction across the globe, into numerous cultures in vastly different environments.
That change probably involved a higher degree of flexibility in adapting to new environments, with expanded frontal lobe and left-brain networks, conducive to creativity, and yet also to selfish impulses, especially as humans evolved from hunter-gatherer groups toward increasingly larger, settled societies (Gazzaniga, Who’s 31–36, 151, 157–58). Eventually, humans developed different cultural attitudes, regarding self-agency or collective harmony, as in the individualistic Euro-American or more communal Asian and African traditions (183). But already, 32,000–11,000 years ago, when humans created cave art in Europe, they had bigger brains on average than ours today,50 with larger frontal lobes than those of Neanderthals. How did the connections in those large brains become specialized for greater adaptability—leading to higher levels of artistic self-awareness and technological transformations of the environment, radically escalating in the last hundred years?
Chimpanzees and orangutans, but usually not gorillas, show self-recognition in mirror tests (with a red dot on the face). According to Daniel Povinelli’s arboreal environment theory, apes that continued to dwell in trees despite their large size evolved a “kinesthetic self-concept,” through their dangerous interactions with arboreal branches, which gorillas lost or never developed because they spent most of their lives on the ground (334). Our common ancestor with these apes, “a large-bodied, highly arboreal species,” may have seen itself in a three-dimensional (3D) “skein of highly variable fragility, compliance, and space,” while swinging and bouncing on tree branches, six million years ago.
Regarding a later stage of evolution, Merlin Donald theorizes that our hominin ancestor, Homo erectus, developed a “mimetic” culture, about two million years ago, involving a “kinematic” imagination, gestural and tonal communication, and playacting (273). Such self-awareness in humans today, with facial recognition and mimetic expression in the mirror, is centered in the right prefrontal lobe (Keenan 134–40). Prosody in human language is also a right-cortex function: “deliberately raising and lowering the voice, and producing imitations of emotional sounds” (Donald 261). Thus, our Homo erectus ancestors, as big apes that moved to the ground and walked erect, brought their self-aware, poetic, right-brain functions with them and developed an early form of theater—through mimetic interactions.
About a half million years ago, according to Donald, humans evolved a “mythic” culture with verbal language, which now involves distinct areas in the left hemisphere (Broca and Wernicke’s areas).51 Donald also finds the start of today’s “theoretic” culture of technology and art 40,000 years ago—with complex tool use, art objects, and art on cave walls, externalizing human memory and retrieving cultural knowledge through symbolic devices (262). Thus, humans’ right-brain self-awareness, as a visuo-spatial, kinesthetic, imaginary skill, extended into a left-brain, verbal, narrative, symbolic dimension of self/other recognition, communication, and theatricality, with devices that transformed the environment. This may have occurred through the hardships of climate change and migration, selecting for a much more mutable human consciousness—to interact collaboratively and creatively in new settings.
However, such successful adaptations in human brains and cultures came at a cost: the loss of instinctual patterns and purposes for life in a stable environment—or the “loss of being” (Lacan’s manque-à-être ). This produces our human “want to be,” our endless desire for more, to fill our hollow, insecure egos, which evolved as kinesthetic, narrative, and technological extensions of Self for survival and reproduction. Prematurely born and then powerfully transformed by the external womb of a particular culture, our vertebrate drives, mammalian emotions, and ape egos became twisted in extraordinary ways, especially through left and right cortical interactions, within and between brains.
Humans are amazingly collaborative, living in populations far larger than other apes. But we are also highly territorial. Our collective egos draw neighborhood, class, ethnic, racial, religious, gender, and national borders—idealizing our allies (or exotic others) and stereotyping our enemies, to commune with or battle. Some of these categories encourage us to cross the lines of conflict and join hands, when we see others as different and yet our kin. But we often find the need to demonize enemies, as outside threats, in order to bind ourselves within the group more cooperatively— with mimetic and mythic identities inherited across many generations (Volkan). Or we make a scapegoat to focus reciprocal violence in the community on a certain sacrificial object, a person or group that can be vilified, punished, or exterminated (Girard).
Wild male chimpanzees use their collective skills not only to hunt and kill monkeys for food, but also to patrol the borders of their territory and kill, trample, tear the skin off, and drink the blood of a lone chimp, which might have been a former family member and playmate (Wrangham and Peterson 5–17). Yet, human groups have gone much further, with the mass murder of neighbors, many times in history, using right-brain kinesthetic and left-brain symbolic stagings of identity and desire. Even the best cultures, with peaks of “enlightenment” in ancient to modern Europe, Asia, Africa, and the Americas (or Australia and Oceania), bear tragic flaws and legacies, with trails of destructive errors, as they continue to revise their neural and social networks, in each new generation.
Psychological tests show that Westerners today, especially those of northern European heritage, are more individualistic, analytical, and linear thinking, whereas East Asians are more holistic, relational, and cyclical thinking (Nisbett). For example, Americans focus more on foreground objects in pictures and the shapes of sculptures (with arbitrary names), while Japanese attend more to background scenes and the sculpture’s materiality, even at the age of two (81–95).52 Americans interpret abstract moving figures on a video screen as having more internal causal factors like shape and weight; Asians see them affected more by context, such as gravity and friction (116–19). Americans believe more in fixed personality types and consistent trends, Koreans in situational influences and changing fortunes (119–20). In random association tests, Americans often infer a false rule, whereas Chinese did not (95–96). Yet Americans are better at judging the position of an object (a rod) without being influenced by its field (a box) than Chinese, who are more contextually oriented. Surveys also show that Asians feel less in control of their lives than Westerners, and instead of trying to control situations, they are more likely to adjust to them (97). Feeling “in control” of their lives is associated with health more by European Americans than by Asians or Asian Americans. And Asians express more well-being than Americans when they are part of a group that might provide control. Women are also found to be more holistic and group oriented than men in both Eastern and Western cultures, yet the differences are greater between cultures than between genders (99–100).
I would extend such findings to argue that Western cultures have shaped the human brain toward left-hemisphere, egoistic, analytical, cause-and-effect functions, stressing individual freedom and greed, especially with American “free market” ideals involving “less government.” East Asian cultures have shaped brain networks toward right-hemisphere (or more bilateral), communal, relational, holistic processes, emphasizing group harmony, with individuals submitting to systematic organization. 53 These current inclinations, exemplified by Western capitalism and Eastern communism (or the different forms of capitalism in various Asian countries), relate to traditional belief systems, especially Greco-Christian and Daoist-Buddhist/Shinto-Zen (Nisbett). Of course, such traditions are mixing today, more and more, with increasing cross-cultural influences. Research shows that an individual Asian-American or Hong Kong resident (influenced by English culture) may be primed in either direction with Eastern or Western cues, such as a dragon or Mickey Mouse, calligraphy or a cowboy, or with personal memories—toward contextual or individualistic, self-motivated perceptions (118–19).
In the last century, Nazi Germany and Imperial Japan demonstrated that Westerners and Easterners are both susceptible to the dangers of emotional contagion and ideological rationalization (as right and left brain processes) in a herd mentality inherited from our mammalian ancestors, which may also produce predatory groups and leaders in a pack or troop system, like wolves and apes, but more wildly and systematically destructive. The recent American development of PACs (Political Action Committees) raising huge funds for campaigning politicians who are then beholden to donors and corporations may be another sign of a herd-pack-troop culture, even in the individualistic West. The contagion of fears and desires, in a herd of antelope (or school of fish), where each member influences others in rippling waves of mostly unconscious psycho-physical gestures, to eat grass, migrate for water, or flee from danger, may shift in humans from a “grassroots,” bottom-up democracy toward a pack or troop hierarchy, with brutally competitive leaders (or investment bankers) operating beyond laws and ethics. This happens also when the business interests and “global security” of nations or religious groups involve violent tactics, with troops or suicide bombers, causing civilian sacrifices, which then call for more and more. Yet such collective, self-perpetuating stress, with the sacrificial demands of bio-cultural survival, draws on mechanisms in our prey and predator, herd and troop heritage that might be altered—if we become more aware of them.
While investigating our brain structures and nervous systems, as inherited from mammalian relatives, Robert Sapolsky found that “zebras don’t get ulcers” but primates do. When alerted to danger as a herd, through emotional and physical contagion, zebras’ stress systems are activated and they flee from predators. One of their weaker or younger members might be lost, sacrificed as a meal to the lions. Yet the surviving zebras, once in safety again, return to munching grass. Their stress system is easily deactivated. But baboons, which Sapolsky studied directly in the wild, forage for grass, tubers, and fruit only about four hours per day (257). They spend much of the rest of the day in stressful competition about the shifting social hierarchy of the troop. And they get ulcers like us.
One might think that the least stressed baboon would be the alpha male. But glucocorticoid hormone levels show that alphas are highly stressed, because baboon leadership changes every seven to eight months, or weekly at other levels of rank and mating rights (Cheney and Seyfarth 57–58, 116–17). The least stressed males are those who gain social control by initiating fights, who best assess when threats are real and predict the outcome, and who use others as outlets for frustration (Sapolsky 263). These baboons “give rather than get ulcers.” But another, more peaceful type of male also has low stress damage, through patterns of “social affiliation.” They spend the most time grooming females who are not in heat, and are groomed by them, while also playing with the young. With other primates, too, it has been found that both genetics and mothering style shape the infant’s behavioral personality in later life (264). Offspring of mothers who are “less anxious and more permissive. . . grow up to be more confident and exploratory.”
Sapolsky argues that humans are likewise “ecologically buffered and privileged enough to get stressed mainly over social and psychological matters”(257). But feelings of predictability or control can reduce stress. Indeed, a nursing home study showed that when visits by college students were predictable or controlled by the elderly, their health benefitted more than when visits were not (270). Yet, Sapolsky warns that feeling in control about a disastrous event might increase the stress afterward, causing guilt because “it’s all my fault.” In relation to Nisbett’s research, this shows a significant danger to the Western, left-cortical bias of self-control.
The staging and screening of Self and Other in our inner (movie) theater involves various levels of our animal heritage: fish/reptilian brainstem areas, mammalian limbic system, primate parietal awareness, and distinctively human right- and left-cortical functions, especially in the executive control areas of the frontal lobes, as “director” and light/ sound “operators.” Such animal levels in our brains produce proto, core, and autobiographical selves. These involve dispositional “puppeteers” and other director/operators of deep goal and conceptual contexts, as well as the “audience” of long-term (automatic, semantic, declarative, and autobiographical) memory systems. They shape the competition of neural actors to reach the “spotlight” (and silver screen) of consciousness, as percepts and concepts, regarding the platform of working memory.
The backstage (or film production) elements of our inner theater include primal drives of survival, reproduction, and territoriality. These remnant animal instincts are radically altered by human cultural environments, especially with right- and then left-cortical attunement to caretakers at an early age, in home rituals as our first theatrical experiences. Our animal drives become expressed through pleasure/pain signals, primary emotions (seeking, fear, rage, panic/grief, and lust), background moods, social feelings, and collective cognitive norms.
Along with visual and auditory cues, smell may trigger collective fear between humans through “alarm pheromones,” which activate the right amygdala (Radulescu and Mujica-Parodi).54 Thus, emotional contagion, akin to that of our mammalian relatives, may contribute, through right-and left-cortical networks, to moral intuition and reason, regarding dangers and errors in the social environment. Emotional contagion with visual and auditory cues from the screen might also convey “as if” dangers and tragicomic flaws in a movie theater, with pheromone cues from other viewers as well. (Similar effects might have occurred in prehistoric cave theaters with rock images and resonant chambers.) The mirror neuron systems involved in such contagious signaling of unconscious backstage agents between brains also relate to the “peak shift” and other neuroaesthetic effects in visual art and music (as key cinematic elements), plus a potential cathartic clarification and rasa re finement in prefrontal areas of conscious mindfulness, with tragicomic twists and various emotional ironies in particular films. But a quicker signaling may occur between our subcortical stagehands, through emotional contagion with screen images and sounds, even as we experience a slower cognitive empathy with more cortical awareness (Eisenberg and Eggum 73). This depends, too, on the “emotional content of pain,” but not the pain itself, mirrored in our inner theaters when we watch scenes of someone suffering, through “deliberate acts of imagination” that increase mirror-neuron activity, drawing on the audience of memory systems resonating with the “context” onscreen (Watson and Greenberg 127–29).
The lure of personal control over stressful situations helps to explain the appeal of horror movies, action films, or farce onscreen and related shows onstage. Identifying with the hero in danger from a monster, villain, or banana peel arouses survival drives through primary and social emotions, in a dreamlike threat rehearsal, where pain becomes pleasurable. With the viewer’s safe distance from the scene and subsequent sense of relief, in the victim’s escape and the hero’s victory, or in collective laughter, the theatrical threat also gives a dose of individual control and communal belonging. Like religious believers, who find relief from anxiety and mourning by imagining how God watches over them and decides what is best—theater, cinema, and television viewers, or videogame players, find hope in a godlike role, while watching characters akin to themselves struggle under stress. The right cortex and limbic system may catch contagious horror, rage, panic/distress, or other bhavas, but the left brain’s belief and control networks can practice their circuitry of being in charge. This may also involve distinct fear circuits: the quicker “low road” through the amygdala with immediate bodily reactions, such as the startle effect, shifting toward the slower, more reflective “high road” through frontal and left cortical networks (LeDoux, Emotional 164). Thus, a further catharsis of complex, tragicomic rasas might challenge left-cortical binaries of good and evil, through right-cortical openness to new perspectives, altering VMPFC prejudices. But virtual stress also demands the real sacrifice of emotion, time, and money for its pleasure and practice rewards, confirming or challenging stereotypes—while drawing on survival and erotic drives in communal and individual ways.
As many Eastern, Western, and other traditions mix in a jet airline and Web-linked world, the animal legacies in our brain theaters continue to evolve, on earth and in the screen cosmos. But traditional ideals of the divine are still attractive to many. They are symptomatic of the current “clash of cultures” between “Matter First” secularists and “Mind First” believers (Richards 260–67)—as well as East and West, or “developed” and “developing” worlds, involving the Party, Buddha, Allah, Yahweh, God, and various other ideals of human transcendence. Figures of the divine mostly have human attributes today. Yet, they emerge from a worldwide tradition, which extends to prehistory, of seeing the animal, human, and divine as a continuum, not just a binary of man versus animal or god versus man. In the next chapter, we will consider how prehistoric cave art, tens of thousands of years ago, shows a critical point in our cultural evolution: a crucial shift in self and Other identity, from natural to supernatural and technological, through the animal within the human. This suggests, too, how a catharsis (and rasa re finement) of cultures might evolve today—not as a battle for survival or an ethnic cleansing of scapegoats, but an awareness, acceptance, and renegotiating of the animal drives in each of us, even perhaps an extended kinship with those who seem to threaten “our way of life.”