![]()
The smut and bunt fungi (the Ustomycetes) produce very dark powdery resting spores (ustilospores) which, en masse, are reminiscent of the black smuts that were one of the perils of travelling by rail in the days of steam. Like the rusts they are biotrophic members of the Basidiomycota, but differ in that most have a long invisible phase when they are systemic within the host. The hyphae grow between and through the living cells but do not produce haustoria. They have simpler life cycles than the rusts, with no alternate hosts and generally only two spore types. They are fairly readily grown in artificial culture and have therefore been extensively used as models for genetic research. Some are referred to as bunts; bunt is an old word of unknown etymology, sometimes used to describe pouter pigeons. Cereal grains packed with the ustilospores of smut fungi in the genus Tilletia are rounded, with a fat, pouter-like appearance, hence the use of the term in plant pathology, perhaps. We wonder whether Billy Bunter’s name comes from the same root.
SMUTS AS PATHOGENS
To the farmer smuts and bunts were at one time catastrophic diseases, especially of cereals. So effective are modern control measures, however, that they are now rarely found in agriculture, although they remain an ever-present threat. Fortunately for the naturalist, they are easily found on wild and garden plants.
In Europe most of us could walk onto any piece of rough ground in midsummer and find plants of the oat grass (Arrhenatherum elatius) with seeds transformed into masses of the black smut spores of a forma specialis of Ustilago avenae, the oat smut (Fig. 10.1). In infected plants every floret in a flowerhead and every flowerhead from a single plant will carry the disease. These black ustilospores are the equivalent of the teliospores of the rust fungi. They are sometimes also referred to as teliospores, brand spores or chlamydospores. In the USA, U. bullata is an equally common grass smut, appearing in the florets and seeds of over 50 species of wild and cultivated grasses. This species is also found in Europe, where it is confined to bromes (Bromus spp.).
But not all smuts form their ustilospores in the flowering parts of plants. The common stem smut of grasses (U. hypodytes) forms its ustilospores on the stem surface, sometimes affecting almost the whole stem. There is no protecting membrane covering the spores, but at first they are enclosed by the leaf sheaths. In Europe the disease occurs on common couch grasses (Agropyron spp.), some bromes and fescue, but is most spectacular on the seashore grasses, especially marram and sea lyme. The same or a closely similar fungus is found on Canadian lyme grass (Elymus canadensis) and ryegrasses in North America.
FIG. 10.1
Black ustilospores of Ustilago avenae filling the florets of oat grass (Arrhenatherum elatius) at Aberlady, East Lothian, Scotland. (D.S. Ingram.)

FIG. 10.2
Black ustilospores of Ustilago tragopogonis destroying the florets of goat’s-beard (Tragopogon pratensis) at Gullane, East Lothian, Scotland. (Debbie White, Royal Botanic Garden Edinburgh.
Equally striking is Ustilago longissima, found in Europe on the leaves of reed grass (Glyceria maxima) and flote grass (G. fluitans) in slow-flowing water. It forms its ustilospore masses within the tissues as narrow stripes in the parts of the leaf not occupied by vascular bundles. They can easily be seen by holding an infected leaf up to the light or shining a torch through it. As the ustilospores mature and are blown away, narrow furrows are left in the leaves. The superficially similar but unrelated species Urocystis agropyri forms elongated blisters on the leaves of couch grasses in both Europe and North America. A similar pattern may be seen in the worldwide species complex U. tritici, the flag smut of wheat. Urocystis differs from Ustilago in having ‘spore balls’ made up of a small number of fused ustilospores with a covering of sterile cells.
There are over 1,100 known species of smuts and more than 60% of them occur on grasses or cereals. Another 10% or so occur on rushes and sedges and the remaining 30% are widely scattered over a number of different flowering plant families. Ustilago vaillantii, for example, is often found in the anthers of a number of spring-flowering liliaceous plants, including squill (Scilla sp.), glory of the snow (Chionodoxa sp.) and grape hyacinth (Muscari sp.). The dark violet ustilospores replace the pollen and dust and stain the mouth of the flower tube, making the infection very visible. A similar smut, Ustilago violacea, occurs in the anthers of members of the campion family (Caryophyllaceae). It is most easily seen in red campion (Silene dioica) and white campion (S. album), where once more the dusting of dark ustilospores readily catches the eye (Plate 7). It has also been known to cause economic loss in cultivated relatives such as glasshouse carnations (Dianthus caryophyllus). Ustilago tragopogonis-pratensis may often be seen on flowers of Jack go to bed at noon or goat’s-beard, the black smut spores destroying the florets (Fig. 10.2).
There are several Urocystis spp. that may easily be seen on non-grass species. U. eranthidis, for example, infects winter aconite (Eranthis hyemalis) (Fig. 10.3). In old established colonies of the plant you may find on the leaves and petioles silvery swellings which open to reveal the dark spore masses of the fungus. Often the infected tissues are swollen and distorted. U. primulicola, a much less obvious pathogen, produces its spore balls in Europe in the ovaries of the wild primroses Primula vulgaris and P. farinosa, and is consequently not easily seen except by a special search. However, secondary spores (called sporidia) are formed in the anthers and appear as white mealy masses replacing the pollen in both thrum- and pin-eyed forms of the plant (Plate 7). They are apparently carried from plant to plant by pollinating insects. The details of the life cycle are fragmentary and need further elucidation. Urocystis cepulae, the onion smut, which moved from Europe to the USA in the 1880s, is a serious disease of sown onions and leeks in cool temperate climates worldwide. The ustilospore balls lie in the soil and germinate with the seed to bring about infection, resulting in swollen areas which open to release the masses of spore balls, permanently damaging the plant. The spore balls do not germinate well above 22°C, which is why the disease does not occur in warmer climates.
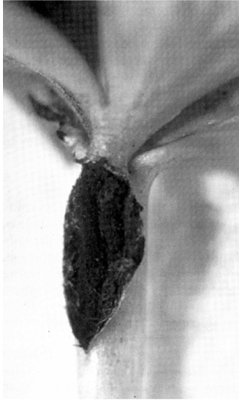
FIG. 10.3
Ustilospores of Urocystis eranthidis bursting out from a lesion on a petiole of winter aconite (Eranthis hyemalis) in Cambridge, England. (D.S. Ingram.)
To complete this brief overview of easily-seen smuts on wild and garden plants, mention should be made of the genus Entyloma. The species most frequently seen is E. calendulae on marigolds (Calendula spp.) and daisies (Bellis spp.). Another form of this species, E. calendulae f.sp. dahliae, which originated in South Africa, is now common on Dahlia spp. throughout Europe. Unusually the ustilospores are colourless or pale brown and embedded in the leaf, where they cause some discoloration of the tissues. They germinate in the lesion and sporidia appear on the surface of the leaf; the process of germination is not unlike that of Tilletia spp, which will be described below.
THE SMUTS OF CEREALS (INCLUDING BUNT OF WHEAT)
Many smuts are associated with the reproductive organs of the plants they infect – sometimes they occupy the ovary, sometimes the anthers and sometimes both. Cereal smuts frequently replace the starchy endosperm of the grain with inedible spore masses, destroying a human food resource. Because of their importance they have been studied extensively and the knowledge so gained provides us with a deeper insight into the smuts in general.
There are two main families of Ustomycetes that attack cereals, the Tilletiaceae and the Ustilaginaceae, separated by the behaviour of the germinating ustilospores. In the family Tilletiaceae the ustilospores are large and frequently ornamented with a net-like pattern. They germinate to produce a non-septate mycelium (the promycelium, equivalent to a basidium), which is negatively geotrophic and therefore bends upwards. On the apex of this promycelium primary sporidia are produced (equivalent to basidiospores). These germinate, fuse in pairs and then produce dikaryotic secondary sporidia which infect the plant. The genus Tilletia and two genera already mentioned, Urocystis and Entyloma, are included in this family. In the family Ustilaginaceae the ustilospores are smaller than in the Tilletiaceae and the promycelium is septate, as in the rust basidium, and produces the sporidia laterally, like rust basidiospores. Most cereals are attacked by different species of the principal genus, Ustilago, which has already been mentioned.
Bunt of wheat (Tilletia tritici and T. laevis) continues to be potentially the most damaging disease of wheat worldwide. Significant annual losses occurred in all countries in the early years of the twentieth century, but were progressively reduced with the advent of constantly improving chemical control. Exceptions were areas such as Pacific northwest America, where soil contamination took place on a massive scale and conditions were particularly favourable to the survival of ustilospores.
The characteristic symptoms of bunt come not only from the replacement of the grain by the ustilospores, giving them in consequence a swollen appearance, but also from the associated fishy smell, caused by the presence of trimethylamine. This has given rise to the alternative common name ‘stinking smut’. The yield of an infected crop is significantly reduced, and contaminated healthy grain is discounted on the market because of the odour. There is also a threat to human health from respiratory and skin allergic reactions to the ustilospores, which may on occasion form dense clouds during harvest. Moreover, these clouds of spores are easily ignited and were once the cause of spectacular explosions and fires on the prairies before steps were taken to design safer harvesting, threshing and milling machinery.
The infection cycle in bunt
As one of the most damaging diseases of cereals in the eighteenth and nineteenth centuries, bunt attracted the attention of the early plant pathologists, and became in consequence a classic disease in the development of the subject. The ustilospores, which are threshed out with the grain of bunted wheat, coat the surfaces of otherwise healthy seed and after sowing infect the young plant as it germinates. The mycelium then becomes systemic in the plant, growing between the living cells, ultimately entering the developing ovary as the flowers form. As these mature further ustilospores are formed within the grain to begin the cycle again. This sequence of events was first described in broad terms by Mathieu Tillet (hence Tilletia) in France in 1755, in the first attempt to demonstrate scientifically that a fungus was the cause of a plant disease. He compared the ears of plants grown from ‘clean’ grain with those grown from the same seed contaminated with bunt spores. The clean grain gave rise to only a few bunted ears, whereas at least half of the ears grown from contaminated seed were bunted. As a demonstration it was a success, but Tillet was lucky to have so little chance contamination by bunt in the plots grown from healthy seed. Further proof of Tillet’s hypothesis came with the work of the Rev. Miles Berkeley a century later. Berkeley’s first attempt to prove that fungi caused disease was made with Phytophthora infestans, the cause of potato blight (see here), but he found bunt of wheat an easier organism to deal with. His studies went some way to establish the connection between the presence of ustilospores on the seed and the development of disease. In addition he appreciated the importance of ustilospore contamination of the maturing grain, and studied ustilospore germination.
Tillet’s work was also extended by another pioneer of plant pathology, Isaac Benedict Prévost from Switzerland. Working in France in 1807, Prévost not only confirmed Tillet’s findings but also germinated bunt ustilospores in the laboratory and guessed that they infected the young plant. The nature of this ustilospore germination was beautifully illustrated by the brothers Tulasne in 1847. A germ tube (the promycelium) is first produced, bearing at the tip uninucleate needle-shaped structures, the primary sporidia (Fig. 10.4). These fuse and produce secondary binucleate sporidia which then infect the seedling in the region of the root node. Further knowledge of the bunt fungus, apart from a reference by Berkeley to bunt mycelium within the tissues of the leaf, came as a result of studies by Oscar Brefeld in Germany in 1888, and the subsequent development of twentieth-century methods of microscopy.

FIG. 10.4
Tilletia caries, cause of bunt or stinking smut of cereals: (a) an ustilospore germinating to produce a promycelium bearing primary sporidia; (b) fusion of pairs of primary sporidia; (c) germination of a primary sporidium to form a secondary sporidium. (Drawings by Mary Bates, after A. De Bary and based on drawings of Tulasne.)
Understanding this life cycle gave the key to control. If antifungal chemicals were applied to the seeds of the host before sowing, the ustilospores would be killed at germination and the infection cycle broken. This identification of the ‘Achilles heel’ of the fungus led to the virtual eradication of bunt from agriculture in the twentieth century and established the principle that recognition of the weak point in a pathogen’s life cycle underlies all successful control measures for plant diseases.
The cereal smuts
The members of the family Ustilaginaceae appear to occupy the same ecological niche as bunt, producing ustilospores in place of grain, and at first sight with the same infection mechanism. The most damaging pathogens are Ustilago nuda, the loose smut of wheat and barley and U. hordei, the covered smut of barley and oats. U. avenae, already mentioned as a pathogen of oat grass (Arrhenatherum elatius), has a special form that causes a loose smut of oats.
When the experimental infection of wheat by bunt had been achieved it seemed obvious that the smuts of wheat, barley and oats would have the same pattern of infection; they did not, and therein lay a mystery. Dusting the seed with ustilospores before sowing led to as many failed as successful infections. Similarly with the entirely separate corn smut (U. maydis) of maize, dusting the grain prior to sowing produced no infection. After much frustrating work it gradually came to be understood that there are four main types of infection pattern for smuts, depending on the time in the growth cycle of the plant at which the fungus infects.
The pattern described for the bunts holds only for the covered smut of barley and oats, U. hordei. All the grains of an ear are usually converted into ustilospores, but these are not blown by the wind because the chaffy scales, although modified, remain as a semi-transparent covering. The ustilospores are released when this covering is broken during harvest or threshing. Seed dusted with ustilospores, or clean seed sown in soil contaminated with ustilospores, gives rise to infected seedlings. The fungus enters at the coleoptile stage, before the true leaves open, and grows systemically in the developing plant, eventually forming ustilospore masses in the grain to complete the life cycle. The actual infection is probably initiated by mycelium growing from the binucleate secondary sporidia produced by fusion of the basidiospores (primary sporidia).
The corn smut of maize (U. maydis), which was rare in Britain until the hot summers of 1975 and 1976, forms large galls on leaves, stems or flower spikes (Fig. 10.5) and it is in these that the ustilospores are formed. In smutted plants the male spike may bear female or hermaphrodite flowers in its lower part. Infection takes place in any growing part of the plant and can apparently arise from direct entry of an ustilospore germ tube (promycelium) or by the entry of a germ tube derived from a sporidium. This type of infection always remains localised and never becomes systemic.
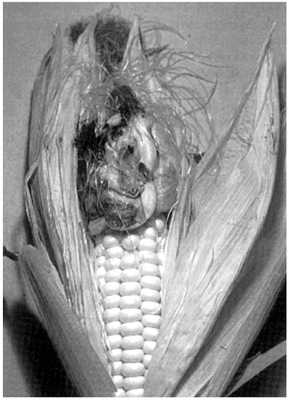
FIG. 10.5
Ustilospores of Ustilago maydis in the swollen and distorted ovaries of corn (maize) (Zea mays). (B. Goddard.)
Shoot infection was first demonstrated experimentally not with a cereal smut but with U. violacea on white campion. Infection takes place through the base of the emerging stem or the buds in the leaf axils and the fungus then becomes systemic and grows with the elongating shoot, eventually forming ustilospores in the flowers. There is a suggestion that ustilospores transferred, perhaps by insects, from an infected flower to a flower on an uninfected plant can initiate systemic infection in that plant. Similar shoot infection leading to the production of ustilospores in leaves and stems occurs in the sugar cane (Saccharum officinarum) smut (U. scitaminea), which periodically becomes a serious disease, mostly in response to faulty breeding programmes that lose the generalised resistance of the adapted crop.
This type of infection occurs in loose smut of wheat and barley (U. nuda) and of oats (U. avenae). Ustilospores released from the uncovered spore masses formed in the ears at about the time of anther maturation are blown to flowers on other plants by wind, where they establish an infection. It was once thought that the ustilospores behaved like pollen grains and were trapped by the stigmas, infecting the ovary by that route. It is now clear that they lodge between the glumes (bracts enclosing the flower) around the growing ovary, germinate before the grain is ripe and put out a promycelium which forms a mycelium (without first forming sporidia) in the scutellum, the layer of secretory cells between the storage part of the grain and the embryonic plant. When the grain ripens and becomes dormant, the smut mycelium, still confined to the scutellum, also becomes dormant. At germination the mycelium comes to life again and grows systemically in the plant, keeping up with the growing apex; eventually it grows into the young developing ear. With U. avenae the pattern is slightly different. The fungus does not penetrate the young ovary, but instead forms a plate of resting mycelium on the surface; this becomes dormant but is reactivated when the grain starts to grow early in spring.
Control of cereal smuts
Serious attempts to control smuts and bunts of cereals date from the early experimental demonstration of the importance of ustilospores adhering to the grain in the transmission of Tilletia caries. Because cereal grains in the dormant state were fairly resistant to mild chemical treatment, the practice developed of ‘pickling’ them in various concoctions, including copper salts. Copper sulphate was first used as a steep, and a lime bath was found to reduce toxicity to the seed. Later copper carbonate was used and, to overcome the uncertainties of wet treatment, dry dusting became standard practice for a time. Spraying dilute solutions of formalin on to oat grains infected with loose smut, followed by a period under a cover, was an effective pickling method used in the early part of the twentieth century.
But all these procedures were imprecise and the parasites might escape control, or the grain might be put at risk by too great an exposure to the chemicals; moreover their effectiveness against different smuts was very variable. A breakthrough came with the development of organo-mercury materials, which were effective against a range of smuts and other seed-borne fungi such as Pyrenophora avenae and P. teres, destructive leaf diseases of oats and barley. But mercury is very poisonous and the dusts were difficult to apply without harming the operative. Slurries reduced the respiratory risk, but the overall toxicity of mercury produced pressure for its removal from the environment. Currently a variety of less damaging fungitoxic organic compounds is used and all quality seed is now supplied ready dressed at time of sowing. Many of these modern fungicides are ‘systemic’, being taken up by the cells of the seedling plant and translocated within it.
Because the loose smuts overwinter as mycelium in the grain a quite distinct method of control was developed for these fungi by J.L. Jensen, a self-taught Danish plant pathologist. He began life as a teacher and magazine editor, then became a seed merchant and from this base became interested in the role of hot water treatment for the control of seed-borne diseases. In 1888 he perfected a procedure for treating barley and wheat seed with hot water, to destroy the smut mycelium without killing the grain. Nowadays hot water treatment, sometimes with systemic fungicides, is used where necessary to treat nuclear stocks from which seed stocks are multiplied. Those requiring treatment can be identified by macerating grain samples in potassium hydroxide, then squashing and staining them to reveal the presence or absence of infecting mycelium in the embryo; where the percentage of infected seeds in a batch is low treatment is not necessary.
Of course seed treatment is not suitable for all plants and all situations, and resistance to smut diseases has been widely bred into cultivated cereals. Both major gene and more complex forms of resistance have been exploited. Also, there is evidence that tolerance is found naturally in ‘adapted crop varieties’ which have been selected, perhaps unknowingly, for survival in situations where smuts are limiting. These can coexist with the disease, resulting in only limited damage. However, such is the capacity for variation in the smuts and bunt that constant attention is required to select varieties that maintain the balance between virulence and resistance. Variation is exacerbated by the capacity of many smut species to hybridise with one another.
THE SECRET LIVES OF THE SMUTS
A feature of so many of the smuts and bunts is their capacity to become systemic in their hosts without any visible antagonistic response by the tissues they are inhabiting. To the casual observer, cereal plants infected with any of the common smut or bunt diseases appear little different from their uninfected neighbours. Farmers, plant pathologists and keen-eyed naturalists, however, who know the plants well, can point to differences in stature, tillering (production of axillary grain-bearing shoots) and vigour. These are not caused by resistance reactions to exclude the fungus, but by interference with the growth control systems in the host and the diversion of nutrients to the developing pathogen. Haustoria are not formed, as they are in the rusts, but the mycelium grows between and through the living cells. When a cell is penetrated the membrane is not broken, but encloses the hypha as it would a haustorium. Layers of new material are deposited between the membrane and the hyphal wall and may have a role in communication between host and pathogen. Presumably nutrients are taken up through all parts of the mycelium, but especially through the hyphae growing within the cells. The relationship between fungus and host is a very cosy one until the moment of spore formation and the replacement of plant tissue by ustilospores.
As is often the case with biotrophs, infection sometimes causes the plant hormones controlling growth to get out of balance. Tissue overgrowth then occurs, as with the maize smut Ustilago maydis. In this disease, when the reproductive tissue is attacked, a whole variety of responses can be observed: sometimes the female or male flowers are replaced by leaf-like structures; sometimes the control of male and female structures appears to break down and male and female inflorescence parts follow one another on the stem axis; sometimes male inflorescences become bisexual, with the development of both male and female entities; or individual grains in the female inflorescence may become greatly enlarged.
Perhaps the most famous example of a smut affecting the sex of flowers is the change brought about in white and red campions infected with Ustilago violacea. When the female flowers of these normally unisexual plants are attacked they become changed, with the development of stamens (with anthers converted to smut spore masses) as well as the normal ovary.
Finally, there are bunts that induce dwarfism in their hosts. In the cereals most bunted plants are slightly smaller than their neighbours, and some strains of Tilletia spp. induce even smaller plants. The most extreme case has come to be recognised as a dwarf bunt, caused by T. controversa, which is known in both Europe and the USA. It came to be called ‘stubble bunt’ because it was missed by the normal settings of the combine harvester. Indeed, it has been suggested that it came into prominence as a result of the advent of new harvesting methods.
SMUTS AND HUMAN BEINGS
Apart from their direct effects in reducing crop production worldwide, marring the beauty of wild and garden plants and providing excitement for plant pathologists, the smuts impinge on other aspects of human life. In a severe epidemic great quantities of spores are released by cereal crops. Breathed in by humans, they cause allergies of the respiratory tract. Even today, when present at comparatively low levels, they contribute to the population of spores over cereal fields, bringing on hay fever in susceptible people. Field and laboratory workers sorting cereal samples are particularly vulnerable, and may also develop severe dermatitis from skin contact with the spores.
Reports of humans and livestock being poisoned by eating spores of smuts have generally not been corroborated. On the contrary, some smuts are recognised as foods. Ustilago esculenta, which infects the wild rice plant Zizania aquatica in Taiwan, China and Japan, causes buds in the axils of the leaves to grow and become swollen and deformed. These swollen shoots, which contain a compact mass of white fungal mycelium, make good eating, we are told. Similarly, the young galls of U. maydis, gathered and eaten before the spores form, are said to be delicious. In Mexico they are given the name ‘huitlacoche’ and in the USA are sold as ‘smoky maize mushroom’. Why not try some? But beware – being expensive they can seriously damage the bank balance.
