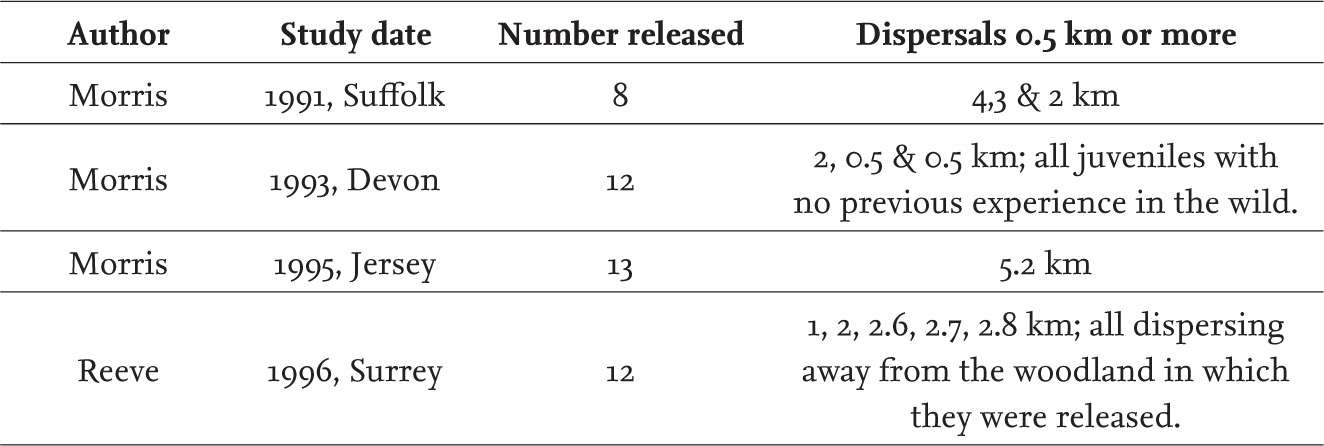
AS A HIBERNATOR, ONE OF the few that we have in Britain apart from bats, the hedgehog shows a very strong seasonal pattern in its behaviour. Few are active before the end of March and most have gone into hibernation by late October (see Chapter 6). This pattern of activity is very clear from records of road casualties, and consistent from year to year when records are collected annually from the same area (Massey, 1972). Whenever regular surveys have been undertaken, road casualty counts also confirm that hedgehogs can be active even in January during temporary periods of arousal from hibernation, but their main period of activity is from April until September (Fig. 101). In many studies, including my own, samples of hedgehogs collected seasonally also often show a very skewed sex ratio, again reflecting changing activity patterns. Males predominate in spring and early summer, females later in the year just prior to hibernation. This corresponds to changes in behaviour, with the males being more active (and therefore more likely to be trapped, seen or run over) in early summer. By late summer, males will be preparing for hibernation, or may have already become inactive, but females will still need to be fully out and about well into the autumn in order to replenish fat reserves lost during lactation. Some females may be still raising offspring in September, actually losing fat at a time when they need to be gaining it ready for the oncoming winter. Females will therefore typically be more active in the autumn than males, and for longer, so there is a preponderance of them in samples collected at this time. There is no evidence to suggest that an apparent skewed sex ratio is anything more than a reflection of this difference in behaviour.
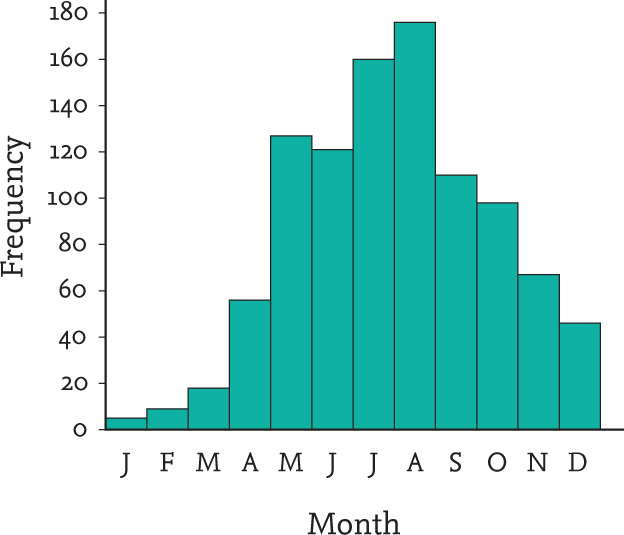
FIG 101. The number of hedgehogs reported for each month of the year in London reflects their annual pattern of activity. These 992 observations were made over half a century ago, long before climate change became a topic of discussion, yet they show winter activity is normal, although at a very reduced level.
The hedgehog is a nocturnal species whose detailed behaviour is difficult to observe in the dark. Any attempt to use a torch or approach closely typically results in a wary response. The animal will stop still and raise the spines on the crown of its head. If it becomes more suspicious, it will hunch its head down between the shoulders and raise more spines on its back. Loud sounds – the click of a camera shutter, for example – or close approach will also prompt this kind of reaction, exactly as Shakespeare described in The Tempest, Act 2, Scene 2, Line 10: ‘Then like tumbling hedgehogs in my barefoot way and mount their pricks at my footfall.’
The hedgehog will remain motionless for long periods waiting for the perceived danger to go away and frustrating any attempt to study its behaviour. If it is touched or provoked, it may well roll up, completely encased in its spiny covering, and stay that way, obstinately waiting to be left alone. This highly characteristic action, so long identified with the hedgehog, is a form of passive defence that will deter most predators (see Chapter 9) but results in many being killed on the roads. Despite all the difficulties of direct observation, it is possible to follow hedgehogs and watch what they do. Some will behave in a very cautious manner, while others can be quite bold and even aggressively bad-tempered. On the other hand, some pet hedgehogs become very tame and will even come when they are called.
GENERAL BEHAVIOUR
Hedgehogs normally begin their activities about an hour after sunset, probably later in areas where there is a lot of disturbance. Its effective spiny defence might make us wonder why the hedgehog is nocturnal at all when it could manage just as well being active during the day. We regard that as a normal behaviour pattern because we humans are primates, most of which are diurnal: daytime activity is the norm for them and for ourselves. However, the hedgehog comes from an ancient line of mammals and the earliest species were nocturnal. Its lineage has remained so, probably because most of the things upon which hedgehogs feed also emerge at night (Fig. 102). That is because creatures like worms and slugs are liable to lose water through their permeable skin when exposed to warm sunshine. They avoid this fate by coming out at night when cooler air results in moist surroundings and even dewfall. Many creatures that are potential hedgehog food are also nocturnal because in the dark they are more likely to escape predation by birds, most of which are active during the day. During dry spells, hedgehogs will usually not set out until after the dew has fallen, probably because there is little to eat on dry ground.

FIG 102. Most of the hedgehog’s normal prey are active at night, like this grey slug (Limax sp.), snails, worms and many species of beetle.
Activity often follows a bimodal pattern, with much moving about soon after emerging at dusk and during the early part of the night. Later, there will be another bout of activity once the animal’s stomach has emptied and it is time to go foraging again. Lactating females will need more food than non-breeders and may have to be more continuously busy. Blood pressure rises as the animal gets going for the night and is highest just after the second period of general activity towards the end of the night. Hedgehogs sleep during the day and are said to dream a lot, more than sloths anyway, but it’s hard to know what about (Grist, 1983).
During its nocturnal ramblings, a hedgehog will normally potter about taking a minute or two to cover little more than a metre, meticulously searching for food and frequently sniffing quite loudly as though using scent to locate edible items. Its progress is often surprisingly noisy, especially among dry leaves, almost as though its spiny coat gives the animal a feeling of confidence and invulnerability, reducing the need for caution and wariness that characterise the movements of most other mammals. A hedgehog’s progression seemingly involves ‘dithering about’, often doubling back on its tracks as though a navigational error had just been realised. Then suddenly the animal may pick up a scent trail and scoot off in a different direction. In normal foraging the hedgehog will snuffle about, poking its nose into the ground, lifting dead leaves and occasionally pausing to sniff the air (Fig. 103). Shaking, yawning, stretching and arching the back are all common behaviours.
Watching a hedgehog moving around over open ground, it seems to have short legs. Actually they are quite long and enable the animal to run surprisingly fast. Averaged over half an hour or more, a foraging hedgehog travels at about 2–4 m per minute, but when it wants to it can scuttle along about as fast as a human can walk without breaking into a run. Nigel Reeve timed some of his male hedgehogs travelling at average speeds of 3.2–3.7 m per minute, females and youngsters around 2.2 m per minute. In short bursts, some hedgehogs managed speeds in excess of 30 m per minute for several minutes at a stretch, although these speedy individuals were all males. One was observed crossing the golf course fairway, covering at least 60 m in one minute, but this was the fastest that any hedgehog seemed to run.

FIG 103. When out foraging, a hedgehog will typically pause to sniff the air at frequent intervals. (Rose Goodwin/BHPS)
Evidence is sometimes offered to suggest that hedgehogs may use a regular route on their nocturnal forays. For example, the German biologist Walter Lindeman used to take two of his pet hedgehogs for a walk and reported that ‘Eri’ followed her favourite path on 47 per cent of her outings and ‘Jossy’ 52 per cent (Lindeman, 1951), but observations like this may be just a case of the animal following familiar linear features like a fence line or the edge of a lawn. Hedgehogs are also often found run over at the same point on a road, again suggesting a regular pathway or crossing point, but overall it seems that activity is not confined to particular routes, at least not for any significant distance. If it rains heavily, the animal will curtail its business and retire to cover, normally by the most direct route rather than follow a particular track back to its nest.
Even at mating time, hedgehogs seem to be mainly solitary. They do not call to each other or use sound for aggressive or social purposes, although young ones may occasionally emit a high-pitched chirping noise and very occasionally a distressed hedgehog can give vent to a loud squeal that sounds just like a pig. On the other hand, although non-vocal, hedgehogs are surprisingly noisy creatures in other ways, trampling and rustling leaves, pushing aside twigs and other obstacles. They wander hither and thither, their nose sniffing and poking at the ground, forever searching and investigating until they retire to their nest about an hour before dawn. Sometimes a hedgehog will make a short dash to grab a fleeing beetle or maybe seize upon a worm. Some have learned not to pull too hard or the animal will snap and half of it will be lost. Instead, the worm will be gripped with the teeth and in a gentle rocking motion eased out of its burrow so the whole thing can be consumed (Fig. 104). One forefoot may be used to wipe off the gritty earth that will be sticking to it, but paws are otherwise not used to grasp food as a squirrel does when holding nuts. Occasionally the hedgehog will pause to groom itself, particularly soon after leaving its nest. It uses the long claw on one of its hind toes, lifting the foot and balancing on the other three. The animal can reach its flanks and also the back and middle of the body and behind its ears by twisting itself sideways to scratch between the spines. It can also lick the forelegs and the abdominal fur, aided by the extraordinary mobility of the skin, which can be pulled around backwards or sideways by the underlying muscles.

FIG 104. Some hedgehogs become adept at coaxing worms out of their burrow. Pulling too hard or too soon will cause the worm to break, reducing its value as food. (Brian Bevan/Ardea)
Scent-marking, to leave a social message or demarcate territory, is an important activity among many species of mammals, but not hedgehogs. While dogs mark lamp posts with their urine very frequently, the hedgehog appears to urinate at random. Otters (Lutra lutra) use their faeces to dispose of undigested food remains, but as these ‘spraints’ are carefully deposited by the otter they are augmented with secretions from scent glands to leave a signal to others of their species. Badgers stake out their clan territories with small dung pits around the territorial boundaries. The hedgehog does none of this and its droppings appear to be left at random, and not as part of an elaborate olfactory dimension to their social behaviour. It is rare to find more than a single faecal pellet in one place and other hedgehogs do not add further contributions as they pass by. This makes it all the more peculiar that accumulations of 50 or more hedgehog faecal pellets have been reported in New Zealand (Jones & Jackson, 2009). It is unclear whether more than one animal contributed, or why they should have been deposited in the particular locations recorded, and it seems not to have been observed elsewhere.
Hedgehogs can be quite aggressive and one will often attempt to bite another’s legs where the skin is thin and unprotected by spines. The hind leg is particularly vulnerable as it is exposed from under the spiny umbrella when the animal tries to run away. Another form of belligerence is to use the spines as weapons in active attacks, the hedgehog attempting to butt rivals with its spine-covered forehead and trying to bowl over any animal whose presence is unwelcome. In response, the victim will move more of its skin and spines over towards the attacker, presenting an increasingly robust and challenging target. One animal may take the hint and swiftly depart or they might both give up and go their own way. Many people who put out food for hedgehogs in the garden often describe something resembling a peck order, even suggesting that the animals may form a sort of queue at the food bowl, headed by the most dominant individuals. Sometimes a particularly large and aggressive hedgehog will briskly dispatch others from the food. On other occasions, several hedgehogs may feed together amicably. Overall there seems to be little evidence of a regular or structured social system among wild hedgehogs, although a dominance hierarchy has been described by several authors, based on observations of hedgehogs in captivity where the physical constraints on space and access to food may distort patterns of natural activity. Fighting and the appearance of dominant behaviour have encouraged speculation that hedgehogs may be territorial, a matter discussed later in this chapter.

FIG 105. Hedgehogs can swim and will readily take to the water, but they easily become exhausted and drown, even in a small garden pond, if they cannot get back on to dry land.

FIG 106. Although they cannot grip with their paws the way monkeys or squirrels do, hedgehogs are capable of climbing walls and fences.
When necessary, a hedgehog can swim moderately well and there are reports of them crossing rivers in this way, even a wide one like the River Thames at Henley (Fig. 105). But if they cannot get ashore, or climb out of an artificial pond, they will become exhausted and drown just like any other animal. Hedgehogs can also dig and climb, easily scaling wire mesh fences and occasionally brick or stone walls. In this, and in tree climbing, they are aided if the obstacle lies at a slope rather than being fully vertical (Fig. 106). There are stories of hedgehogs climbing ivy-clad walls and even nesting in a thatched roof. I have been told several times about them climbing upstairs in houses, although it is hard to see what motive there would be for scaling an indoor staircase. A climbing hedgehog seems to have no fear of falling, probably because it can roll over and have a hard landing cushioned by its springy spines.
SELF-ANOINTING
Unlike many mammals, hedgehogs seem to spend no time in play, not even the young ones. They may interrupt normal activity for occasional stops to groom or indulge in sexual activity, but their active hours are normally devoted almost entirely to the serious business of seeking food. It is therefore all the more remarkable when an animal stops what it is doing and begins to indulge in what appears to be a very energetic, but completely pointless, activity that we now call ‘self anointing’. This bizarre and somewhat alarming, almost manic, behaviour on the part of hedgehogs seems to have been first described in 1912 by Ludwig Heck, a German zoologist who named it Selbstbespucken, meaning ‘self-spitting’. Konrad Herter later dealt with the subject only briefly and other German, French and Danish authors also subsequently described this behaviour without elaborating further. It was Maurice Burton who wrote the first detailed account (Burton, 1957), and coined the term ‘self anointing’ to describe it.
What happens is that a hedgehog will stop its normal activity and begin to salivate copiously, often stimulated by chewing at a variety of objects. When it has a frothy mass of saliva on its tongue, it twists the body and throws its head over its shoulder sufficiently far to flick the froth onto its spines. Sometimes this involves lifting one foot off the ground or leaning so far to one side that the animal may actually fall over. Undeterred, it continues with this extraordinary performance, spreading saliva on both its flanks and on its back. Its eyes are wide open and the creature appears deeply engrossed in what it is doing and totally disengaged from its surroundings (Fig. 107). Burton expressed surprise that this startling and conspicuous behaviour had gone unreported anywhere until 1912, yet when he published his own observations in the 1950s there was an immediate flurry of response from many other people who had seen the same thing, usually with pet hedgehogs. The distracted behaviour and violent contortions caused some observers to fear that the animal was suffering from epilepsy or having a fit. Burton described a hedgehog called ‘Rufus’ who spent a lot of time self-anointing and seemed to be stimulated by contact with almost anything. The carpet, wool, rugs, glazed tiles, human sweat, the varnish on a wooden bookcase, garden plants, insulating tape, glue, the bristles of a brush, earthworms and dog excrement would all set him going. As Rufus continued self-anointing, he seemed to develop a frenzy and after a while would use his teeth, biting as well as licking. This frantic behaviour could go on for 20 minutes or more, although other hedgehogs self-anointed only briefly and most did not do it at all.

FIG 107. A hedgehog in the act of self-anointing seems almost manic in its efforts to spread saliva over its body, ignoring all else, including the flash of a camera.

FIG 108. The hedgehog spreads frothy saliva over its fur and spines, twisting the body to an extent that seems barely possible.
Maurice Burton’s correspondence files revealed a wide variety of items that had set off a hedgehog’s bout of self-anointing and the list continued to lengthen as still more observers contributed comments. The reported stimulants included the skin of a dead toad, a clean handkerchief, cigar butt and an empty cigarette packet, as well as leather goods and at least 20 other different and completely unrelated things. The list includes various astringent or aromatic substances that might well stimulate salivary flow, but self-anointing has also been initiated by contact with nothing more exciting than distilled water. It was frequently reported that a hedgehog would sniff assiduously at the surface of an object before starting to lick it and at intervals would often return to exactly the same spot to lick it again. Among pet hedgehogs, there are some that would only self-anoint on the same few things, whereas for others there was an almost unlimited list of substances and objects that turned them on. A few hedgehogs were reported to perform only occasionally, but the majority seemed never to engage in self anointing at all. The action seems to occur in both sexes, possibly slightly more often by males. It has even been reported to occur among hand-reared nestlings and juveniles (Figs. 109).

FIG 109. It’s not only the adults that engage in self-anointing; this hand-reared baby is doing so with food mixed with its saliva. (Melissa Ilston/BHPS)
This behaviour is so bizarre that it would be easy to assume that it was an artefact brought on by the unfamiliar and perhaps stressful conditions of captivity. But Burton observed that dried spittle also occurred on the spines of wild hedgehogs. Bob Brockie inspected 929 hedgehogs during his field studies in New Zealand and observed saliva on 19 of them (Brockie, 1976). He said they had (to him) a distinct smell and suggested that self-anointing might have some sort of sexual function, but offered no evidence in support of that idea. In three years, Nigel Reeve found only nine wild hedgehogs that showed clear evidence of recent self-anointing, confirming that it is a normal but infrequent behaviour. Three of his animals were sub-adults and two had covered themselves in dog faeces. Nigel also made the first direct observation of wild hedgehogs actually engaged in self-anointing. He found evidence of it outside the breeding season and not associated with courtship behaviour. ‘Emily’, his tame hedgehog, self-anointed at all times of the year, confounding Brockie’s speculation. A study based on a larger sample of 194 wild hedgehogs in Belgium found a higher proportion of animals indulging (at least 11 per cent) and confirmed that self-anointing occurred among both young and old animals. Indeed the sub-adults seem to have been much more active in this behaviour than the older generations. Males engaged in the behaviour significantly more often than females and there was a peak of activity in July among the adults, but not in the younger animals (D’Have et al., 2005).
The long list of potential stimulants is matched by the diversity of speculative explanations for this extraordinary activity. Almost every suggestion that matches one aspect of this behaviour is ruled out by another observation. For example, one idea was that it was some form of greeting, but hedgehogs normally self-anoint alone. Another suggestion was that it makes the hedgehog unpalatable to its predators, although these are few and observations show them more likely to be deterred by the sharp spines. One of Burton’s correspondents mentioned a hedgehog that self-anointed only with boiled beetroot and went about coloured purple, but this cannot be a universal explanation, for the behaviour especially as colours are meaningless in the dark. Some people have suggested that it might be a means of getting rid of fleas and ticks, but if so it is remarkably ineffective. Anyway why concentrate on adding saliva to the spines when many of the skin parasites live on the legs and belly, which are easier for the tongue to reach? Another suggestion was that self-anointing was a means of keeping the spines supple, but why? Spines are better protection if they are stiff and firm. Yet another idea, prompted by the use of toad skin (Brodie, 1977), is that saliva contaminated with the toad’s natural skin poison placed on the spines would make them more effective against enemies. But hedgehogs don’t normally find or seek out a convenient toad and the spines are usually so sharp and numerous they form effective protection regardless of their taste or smell. It has even been suggested (with no supportive evidence) that self-anointing may be some form of residual behaviour aimed at keeping the hedgehog cool, dating from millions of years ago when hedgehogs first evolved in tropical climates.

FIG 110. Self-anointing by a juvenile hedgehog, probably only about ten weeks old. (Graeme Thompson/BHPS)
None of these explanations seems very likely, given that only a minority of hedgehogs perform this behaviour and do so only on rare occasions. Moreover the behaviour has also been reported from at least three other species of hedgehog which live in such a variety of conditions and circumstances that it is difficult to imagine any of these explanations could apply to them all. In the absence of any scientific evidence or proof, the most likely function of self-anointing seems to be to provide an enhanced form of olfactory recognition. Perhaps, instead of scent-marking a lamp post or depositing scent-filled faeces as markers, the hedgehog is scent-marking itself. Maybe self-anointing serves to signal ‘here I am’ to other hedgehogs? The hedgehog’s spines provide an enormous surface area from which scent from evaporating saliva could be wafted on the air. Perhaps the saliva is enhanced by the addition of pheromones from secretory glands under the tongue like we find in pigs. In that connection, it may be noted that several observers have pointed out that the hedgehog licks with the upper surface of the tongue to accumulate material, then uses the underside when actually anointing the spines and fur, but that could be due to anatomical limitations of not having a longer tongue and may also not always be what happens. It may be relevant that Madagascan tenrecs (similar to hedgehogs in many other ways) spread a mixture of urine and saliva on their spines and bush babies urinate on their hands then transfer that scent-laden substance to their fur and to the surfaces where they walk. Perhaps these behaviours serve to enhance and diversify the olfactory regime within which many mammals operate, unrecognised by ourselves due to the relatively inferior sense of smell possessed by humans.

FIG 111. A juvenile hedgehog, barely old enough to have left its mother’s nest, self-anointing so vigorously that it is in danger of falling over. (Julien Crowther/BHPS)
Hedgehogs have a very well-developed sense of smell and in addition there are suggestions that they may have a functional Jacobson’s organ in the roof of the mouth. In snakes this structure plays an important part in detecting scent. Many male deer and antelope engage in a behaviour known as ‘flehmen’ in which the lips are raised and the air around potentially fertile females is sampled by sucking it into the mouth, there to be analysed in the Jacobson’s organ. Maybe hedgehogs do something similar, depositing saliva, enhanced or not, on their spines to liberate an odour that can be detected by the Jacobson’s organ of another individual. That might help them convey messages to each other about their identity and location. Hedgehogs appear not to be territorial, so they do not need to put down markers to defend where they live. Maybe instead of using a lamp post like a dog to announce its presence, the hedgehog uses its own body. Its spiny coat probably compensates for the additional risk of drawing attention to itself. But this remains just speculation, confounded by the observation that relatively few hedgehogs engage in self-anointing and those that do include both sexes as well as juveniles and adults. The behaviour is also not limited to the breeding season, when it might seem to be most useful for them to be sending out social signals.
Baffled by all the contrary observations he received, Maurice Burton toyed with the idea that self-anointing might actually have no purpose at all. He suggested that it could be comparable to the habit in some birds of frolicking among colonies of ants or in clouds of smoke. He also suggested that what cats do in the presence of catmint was also behaviour with no obvious functional benefit and wondered if self-anointing by hedgehogs could be similarly pointless (to humans at least) or just a form of play. The problem with the idea of it being ‘non-adaptive’ is that there would be no evolutionary pressure for it to develop if there was no benefit in doing so. On the other hand, we humans do an awful lot of things that appear to convey little or no biological benefit, ranging from abstract art to playing golf!
RUNNING IN CIRCLES
Another apparently pointless behaviour described by various people is where a hedgehog keeps running in circles. The most detailed account was published by the Reverend Boys Smith based on observing a hedgehog that regularly visited his garden at the Master’s Lodge of St. John’s College, Cambridge. Over a period of a month, almost every night when it was not raining, this hedgehog took to running in a large circle about 14 m in diameter. The activity went on for long periods in the same part of the garden, invariably running anticlockwise. The animal ran quite fast (he thought about 130 m per minute) across a grassy lawn and a gravel drive. The running began soon after dusk, but the hedgehog also performed its circulatory ritual at various times later in the night, apparently completely absorbed in what it was doing (Boys Smith, 1967). This story was widely publicised and discussed at the time. As a result, I received many letters on the subject and one of my correspondents, Cherry Johnstone, reported a hedgehog that regularly travelled in a circle in the same part of a garden in Canterbury. Apparently it did 31 circuits one night. Others wrote to me with similar stories, describing circuits extending to 20 m or more in diameter, and with the hedgehog running so vigorously and so frequently that a deep track would be beaten into the ground (Figs. 112 & 113). More incidents were reported in letters to newspapers (Burton, 1969). Similar behaviour was also observed in captive hedgehogs (Dimelow, 1963), with the same animal always running either clockwise or anticlockwise.
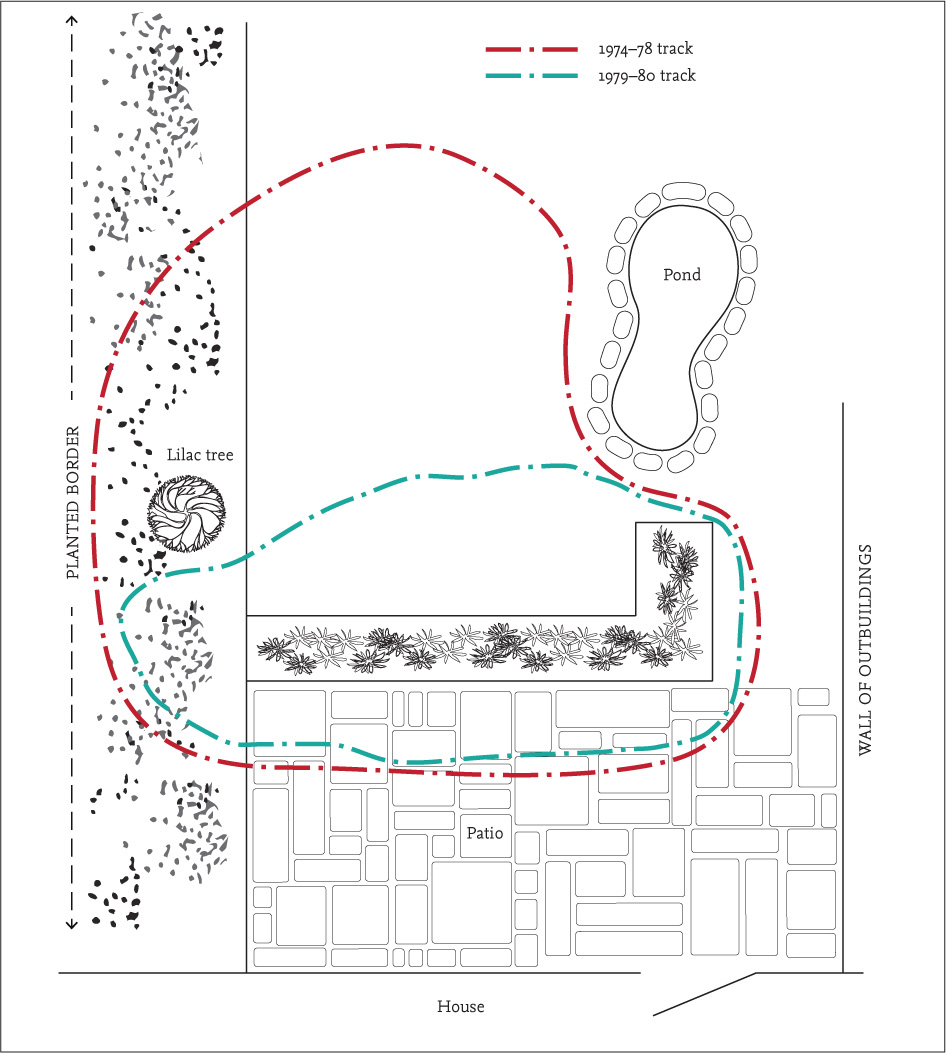
FIG 112. One of my correspondents, Alan Brack, sent me this diagram of his circling hedgehog’s activities. It would run clockwise non-stop for the 20-m circuits, sometimes 10 to 12 circuits without a pause.

FIG 113. He also sent a photograph showing how the hedgehog had beaten a trail into the grass during its rapid running. It would sometimes break off to feed elsewhere, then return to its regular track, night after night.
Manic running in circles is unlikely to be normal behaviour and is probably the result of some form of sickness in the animal. In the 1960s, it was quite common to find wild animals in a disoriented state, apparently unable to walk or fly properly. This was often the result of pesticide poisoning. In those days, various organochlorine compounds were widely used for timber preservation and for killing insect pests on agricultural land. These man-made chemicals do not readily decompose in natural systems and small amounts accumulated in animals that then became concentrated in the bodies of predators higher up the food chain. Instead of killing these larger animals outright, the toxic residues caused sickness and sterility, with impaired locomotory abilities and a lingering death. The reported cases of hedgehogs running in circles seem to have come mainly from the period when such chemicals were in widespread use. There appear to have been few, if any, reports from earlier than the 1960s, despite this being such a conspicuous form of behaviour that it would surely have been noticed and described in earlier books about mammals if it were commonplace. Also, few such stories are coming to light now that the most harmful cumulative poisons have been withdrawn from use. So it is possible that some hedgehogs accumulated residues of DDT, Dieldrin, Lindane or similar compounds through ingesting minute quantities in the individual prey items that they ate. Over time, the toxic residues could build up in the body and disrupt normal behaviour.
However, it is not clear why that kind of poisoning should be manifest in the form of circulatory ambulations. An association with agricultural chemicals was pure speculation, so when I was presented with a hedgehog that ran in circles, I decided to have it euthanased and autopsied in order to resolve the question. A bacterial infection in the middle ear was found. This offers a very plausible explanation for running in circles because the middle ear is the part of the sensory system that enables vertebrates to orientate themselves and maintain their balance when they move about. If one ear became infected and no longer operated in synchrony with the other, an animal would find it hard to travel in a straight line. Running in circles in an agitated manner would be a likely consequence. The pain associated with a severe infection might well result in a hedgehog adopting a distracted state, uncaring of what was going on in its surroundings and behaving exactly as described by many observers. One test of this hypothesis would be to treat a ‘circle-running’ hedgehog with antibiotics and see if it became cured of the affliction, but I have never been given another circling hedgehog upon which to experiment (perhaps because of what I had done with the first one).
SUMMER NESTING
At the end of a night’s activity, the hedgehog withdraws to a sheltered place to rest for the ensuing day. Unlike the large hibernation nests which are relatively easy to see in the sparse vegetation of winter (see Chapter 6), summer nests are very hard to make out. We must rely on radio-tracking to discover where hedgehogs normally lie up each day and even then it may not be possible to find exactly where the animal is sleeping without excessive disturbance and destruction of the surrounding vegetation. Most of the early accounts of hedgehog ecology made casual references to lying up in shrubbery or thick vegetation during summer days, based upon occasionally and accidentally coming across them. The first systematic observations are from Nigel Reeve’s study on Ashford Manor golf course in 1978 and 1979, with various subsequent studies elsewhere serving to confirm that his observations represent normal hedgehog behaviour (Reeve, 1982; Reeve & Morris, 1985). Nigel’s radio-tracked animals usually spent the day in a specially constructed nest, although they sometimes merely used a sheltered place in thick vegetation (which he referred to as a ‘lair’). Other authors, including myself, have also sometimes found hedgehogs merely sheltering in long grass or other dense vegetation, with no evidence of nesting material. This may be quite common in fine weather conditions, as Elizabeth Dimelow’s captive hedgehogs often did not bother to make a nest when the temperature exceeded 16°C (Dimelow, 1963).
Nigel Reeve located 67 daytime resting places in one summer, but only 58 of them were sufficiently accessible to permit close inspection without undue disturbance. Two-thirds were well supported externally, such as those that were sited under bramble bushes or tucked up against solid objects. One nest was at the end of a passage into a tree stump and four were in burrows. These were tunnels a metre or so in length with a single entrance. They could have been dug by the hedgehogs themselves, but were more likely to have been excavated by rabbits. The use of rabbit burrows for nesting has been widely reported before and they are probably an important form of shelter in open and treeless landscapes.
Only 41 of Nigel’s nests could be inspected closely enough to discover their precise composition. They were mostly (83 per cent) formed from naturally fallen leaves, creating a small mound, the rest being made of grass and leaves or grass alone. The nests were usually well constructed, implying that feeding time had been sacrificed in favour of nest construction. This suggests that nests are important to the animals in summer as well as in winter, and the availability of suitable sites and materials may be an ecologically limiting factor in some habitats (see Chapter 6). Summer nests tend to be smaller and less substantial than hibernation nests, although they are usually constructed using similar materials. They are used periodically throughout the active season, often abandoned and then reoccupied later or possibly used by a different hedgehog. Occasionally the same nest may serve as a hibernaculum during the ensuing winter and in a few cases Nigel found the same sites being used in consecutive summers, although probably not exactly the same nest.

FIG 114. Comparative use of summer nests by male and female hedgehogs. Each horizontal row in these diagrams represents a different nest. Females tend to return to the same nest night after night for long periods (top, in green), whereas males (above, in red) use a variety of nests, enabling them to range over a wider area on consecutive nights.
During the active season, a hedgehog will use several different nests. One of Nigel’s males occupied 10 different nests in 118 days, widely distributed within its summer home range of 39 ha. Another male used 14 nests in 68 days (with one day in a lair), within a home range of similar size (Fig. 114). A female used only 5 nests in 88 days, clustered more centrally in her smaller 14 ha of summer home range. There was a clear tendency for both sexes to use a nest only once before moving to another: of the 136 periods of occupancy recorded, two thirds were for one day only among the males and about half among females. There is much individual variation in the frequency with which a hedgehog changes its nest, but several studies have shown that males move from one nest to another significantly more often than females. In Nigel’s study, males changed their nesting place about every three days. Three males used at least 10 nests each in the same summer, one of them making 42 changes among 11 nests in 4 months. By contrast, non-breeding females spent an average of 9.6 days in the same nest before moving on and several used the same nest every day for 2 weeks or more. In another study, hedgehogs were tracked in woodland and in 3 weeks 1 female used at least 3 different daytime resting places and changed her nest 4 times, while the male made 6 changes among 4 different nests in 19 days (Morris, 1986). Mothers will normally remain at the same nest throughout the time that they are rearing young, often more than a month, although they will sometimes move their family to another place. In a longer study (in Ireland), Amy Haigh found a total of 117 different day nests, with individual hedgehogs using up to 11 nests during the summer and returning to a particular nest up to seven times. Her hedgehogs built both day nests and hibernacula in farmland hedgerows (Haigh et al., 2012b).
TABLE 7. Number of summer nests used and frequency of changes; data from two seasons combined. * indicates juvenile. (Based on Reeve & Morris, 1985).

In most studies, hedgehogs have built a nest, used it for a while and then gone away. Some would return to it again later, suggesting that they ‘know their way around’ and may perhaps have a selection of nests that could be used as needed. This would make sense, given that valuable time will have been spent in nest construction and it would be more efficient if an existing nest could be found and used again instead of making a new one. Exactly this behaviour was observed among rehabilitated hedgehogs released at unfamiliar sites after a period of veterinary care. They soon learned their way about in their new home and, having built nests, went directly back to them at the end of a night’s foraging. Nest construction appears to be instinctive, as juveniles with no previous experience of life in the wild behave the same way as adults (Morris et al., 1992; Morris & Warwick, 1994).
Hedgehogs will also use rabbit burrows, in summer and in winter. Even with radio-tracking they are hard to locate underground, so the importance of burrows may have been overlooked. At sites in New Zealand where rabbits were numerous, more than half of resting hedgehogs spent the day in a rabbit burrow.
Captive hedgehogs will often share the same nest during the summer, but there is no published information suggesting that this is a frequent occurrence among hedgehogs in the wild, a potentially vital activity in sustaining the life cycle of their parasites (see Chapter 11). Nigel found no instances of hedgehogs living together, but observed seven cases of non-simultaneous nest sharing where one hedgehog used a nest that had been previously occupied by another. In six of these, the ‘visitor’ was a male; all the nests being visited had been occupied by a female. No female ever occupied a nest known to have been used previously by a male. This type of ‘home visiting’ will have been under-recorded because it can only be evident among the few radio-tagged animals being studied at a particular time. Visits by non-radio-tagged animals would pass unnoticed. We also do not know whose nest it was initially unless a radio-tagged animal was actually observed to build it. There must be plenty of existing nests available to freely moving hedgehogs if only they can find them. However, there seems to be no organised pattern of use comparable with the satellite setts of a badger main sett, for example. Several nests are potentially available to a hedgehog at any one time and their use is spasmodic and unpredictable.
The tendency for males to change their nests more often than females may be connected with the larger home range of the former. It is expedient to have several widely dispersed nests instead of having to return repeatedly over long distances to the same one. This would allow a male hedgehog to range more widely in search of females and also enable him to exploit potential feeding areas that were a long way from the centre of his normal activities. Meanwhile female summer nests will include those that they use to raise their young. These must be bigger and more robust structures than the temporary nests that are only occupied for a day or two. Nursery nests are often large and resemble hibernation nests, frequently with the inclusion of paper and soft fabrics torn to pieces with the teeth while being held down by the front feet. Once the young are born, the nest will be used every day for about a month. Although a mother will sometimes move her young to another nest, she will cease her normal behaviour of frequent nest-changing until her offspring are weaned. In one of my studies, a female had a nest with five young, estimated to be five to seven days old as they were still blind and dependent upon their mother for milk. She had been out of the nest foraging every night for the preceding five nights, which almost certainly spanned the births, but she did not appear to have returned to feed her litter during the hours of darkness (Morris, 1986).
NOCTURNAL WANDERINGS
Hedgehogs can clearly learn the geography of their surroundings, finding their way back to a previously used nest for example. Experiments show that they will also learn good places to feed, perhaps using visual cues, and will return there repeatedly (Cassini & Krebs, 1994). They will go back to a regularly placed food bowl in the garden even after it has been removed. Displaced animals can also find their way back to a garden from a distance of more than 1 km, and a reader’s letter in Woman magazine in 1956 reported a hedgehog that had returned from 3 km away. Several studies of marked hedgehogs suggest that some remain faithful to their normal home area year after year, while others may disperse over long distances, especially in unfavourable habitat. Patrick Doncaster found some of his animals travelled up to 3.8 km away (Doncaster et al., 2001), even dispersing through an existing population instead of joining it. At least three animals ‘homed’ by returning to their release point. A hedgehog that I marked in Jersey travelled over 5 km from where it had been first found, apparently prevented from going further by its arrival at the coast. A marked hedgehog released in Devon was reported to me having apparently gone 15 km, crossing roads and other challenging obstacles on the way, and it probably never went back. It is possible that the disturbance associated with capture or marking (and sometimes translocation) of these animals caused them to disperse further than they might otherwise have done. In New Zealand, several dispersed more than 5 km, one managed 10 km in 6 months and another was found 12 km away after 26 months. Dispersal ensures that the hedgehog population is well spread out, reducing competition for food and ensuring genetic diversity is maintained. After hedgehogs were released on South Uist, the population spread at about 1 km per year on average for six years and 2 km per year thereafter. Dispersal and long-distance movements appear to be a natural part of hedgehog ecology, at least for some individuals, which is all the more reason to be concerned about the creation of artificial barriers to free movement as discussed later.
TABLE 8. Evidence of dispersal movements. Returns from a small project carried out with the help of the BHPS where numbered tags were attached to 123 hedgehogs due for release by hedgehog carers. The tags bore a telephone number to report sightings and 11 were seen again after release (a 9 per cent rate of returns). The distances covered in relatively short periods suggest much more activity than is normally thought likely for hedgehogs, although the reported dispersal of 15 km is so remarkable as to invite speculation that the tag number was misread, especially as it was a female and these do not usually travel so far as males. No. 00784 crossed a major road and a river.

TABLE 9. Dispersal movements by hedgehogs released after a period of confinement in captivity. All were released into unfamiliar locations and might be expected to scatter widely as a result. Some remained close to the original release point, and others may have gone further than indicated after contact was lost or after two months when the study ended. Clearly, even hedgehogs with no previous experience of independent life in the wild can and do undertake surprisingly long-distance movements.
MOVEMENTS: HOME RANGE
Occasional dispersal movements or returning from far away are one thing, but what about normal activities? The hedgehog is a species of woodland margins, gardens, hedgerows and farmland, all habitats that are characterised by great physical and biological diversity. Prime hedgehog areas include mixtures of long grass, short pasture, bushes and trees offering a wide selection of potential food items and also suitable nesting sites. This much has been known for centuries, but exactly how a hedgehog behaves in these places, how far it travels and how much space it needs have all been key questions, frequently asked, but with the answers obscured by the cloak of darkness under which hedgehogs operate.
My own ineffective attempts at radio-tracking in 1966 were intended to throw light on the subject, but actually revealed little. Nevertheless, it became clear that hedgehogs (at least 24 of them in the space of a few weeks) would congregate in profitable feeding areas where the short-grazed turf made foraging more fruitful. Few remained in the hibernation areas that they had used the previous winter, as though a seasonal change in range area had occurred. One animal was followed for 300 m from its daytime nest to a favoured feeding area and it returned later in the night to the same nest, finding its way in the dark across ground traversed by the scent trails of countless people, dogs and deer (Fig. 115). I also found that some animals would use a different nest at the end of each night. All this was new information, with the promise of much more to be discovered once better equipment became available. But it would be another ten years before that happened, when new types of transmitters and receivers began to offer the prospect of studying movements and home range in the sort of detail needed.
The concept of ‘home range’ was devised by an American mammalogist keen to make sense of patterns of recaptures in studies of small mammals caught in a grid pattern of regularly spaced ‘live catch’ traps and then released and captured again somewhere else. Traps could be set for a few nights and each time an animal was caught this could be marked on a map of the trap grid. After a few nights, one could draw a line linking all the capture points for a particular animal and show where it had been, its ‘home range’, and compare this with other individuals or different species. But using traps like this means only one capture could normally be marked on the map for each trapping period. Even checking the traps every couple of hours (and disturbing the animals in the process) still gives only a snapshot of the general areas visited. With the development of radio-tracking in the 1960s, the movements of animals could be explored further and in more explicit detail. Home range became widely accepted as a description of an animal’s ‘normally used area’. As a radio-tagged animal walked about, its location could be determined very frequently, giving a much more realistic reflection of its spatial activity. With improvements in equipment and techniques of analysis, the home range concept was extended and elaborated upon to paint a very detailed picture of an animal’s movements.

FIG 115. A combination of radio-tracking and mark-recapture showed six hedgehogs that wintered in the plantations at the edge of Bushy Park in 1967 travelled to an open area of grassland to feed in the following summer. Female 113 had a nest with young in a patch of bracken in October of 1967, commuting between it and the feeding area during the night.

FIG 116. Nigel Reeve with his home-made radio-tracking equipment following hedgehogs at night on Ashford Manor golf course.
My own attempts at using radio-tracking involved crude home-made equipment which was not only heavy and impractical, but my hedgehogs quickly carried their transmitters out of range of the receiver. I was never going to learn much about how hedgehogs spent their time. Nigel Reeve took up the challenge ten years later, benefitting from a better system that could detect tagged hedgehogs more than 200 m away (Reeve, 1979). This made it possible to follow several animals at once and from far enough away as not to influence their behaviour (Fig. 116). Several position fixes could be obtained for each animal every night and plotted on a map. The usual way of describing the results of such studies was to join up all the outermost points visited, forming a convex polygon whose area could then be calculated to represent the animal’s home range.
Nigel’s study focused on the hedgehogs that inhabited Ashford Manor golf course, an area of approximately 40 ha, and its adjacent gardens. For the first time anywhere, not just in Britain, he was able to gather information about how much space hedgehogs used and how far they travelled in a night. The main study area comprised mown grass with small patches of woodland and shrubbery separating the fairways. The hedgehogs’ calculated range areas were based on all the sightings and position fixes obtained during one active season – i.e. a yearly range. On average, it was about 32 ha in males and 10 ha for females. This can be visualised as males using the whole of the 18-hole golf course and females about half of it or less. Although there was considerable variation in range area between individuals, males generally appeared to use about two to three times as much space as females, a statistically significant difference. Similar results have been obtained in many subsequent studies.

FIG 117. Over time, radio-tracking equipment improved, with longer range, longer battery life and smaller transmitters. This one has a luminous tag attached to help locate the animal more precisely in the dark without approaching too closely and disturbing it.
The basic problem with any estimates of home range size is that the figure obtained is hugely dependent upon the number of observations used to calculate it. If an animal is located three times, its apparent home range is a triangle of a certain size. If it is seen a fourth time, the range area is increased, perhaps substantially so if it is found way off to one side, creating a rectangular range area. None of Nigel’s home range estimates were based on fewer than 30 location fixes and most used more than that (up to 532), but they were spread over an entire season. It is tempting to list all the subsequent published estimates of home range size for comparison, but are they really comparable when they have all persisted for varying lengths of time and used different methods for analysing their results?
TABLE 10. A selection of ‘average home ranges’, based on observations carried out over a week or more. A much smaller area would be used on a single night.

But these are home ranges based upon studies that lasted weeks or even months. What do hedgehogs do in a normal single activity period? It seems unlikely that a hedgehog could patrol 30 ha in one night (almost the entire area of Nigel’s 18-hole golf course). In a small study of hedgehogs in woodland, two females averaged 1–2 ha per night and a male 2.4 ha (Morris, 1986). This might suggest that hedgehogs in woodland could have smaller home ranges than those on the golf course, perhaps because the animals move about more easily in the open habitat than in woodland. But activity on a single night should surely not be compared directly with a whole year’s observations. And which of those figures best represents a hedgehog’s home range – the ‘normally used area’? On different nights the animals visited different places, not always the same localities. Adding position fixes from successive nights and measuring the progressively enlarging polygon enclosed by the outermost points provided an estimate of the minimum total area familiar to each animal in that woodland study. The largest ‘cumulative home range’ thus obtained in the woodland was 11 ha, the ‘normally used area’ beyond which the animal seemed not to go. Again it suggests smaller home ranges than on the golf course. But this was based on only a three-week study; the golf-course figure was for the whole summer season. Field studies tend to last for different lengths of time, driven by the needs and convenience of the investigator. When seeking comparative data in the literature, it also becomes evident that investigators may not have stayed up all night and their calculated home ranges will be incomplete. Clearly the time dimension needs to be considered and information given about hedgehog movements and home range needs to be taken with a pinch of salt.
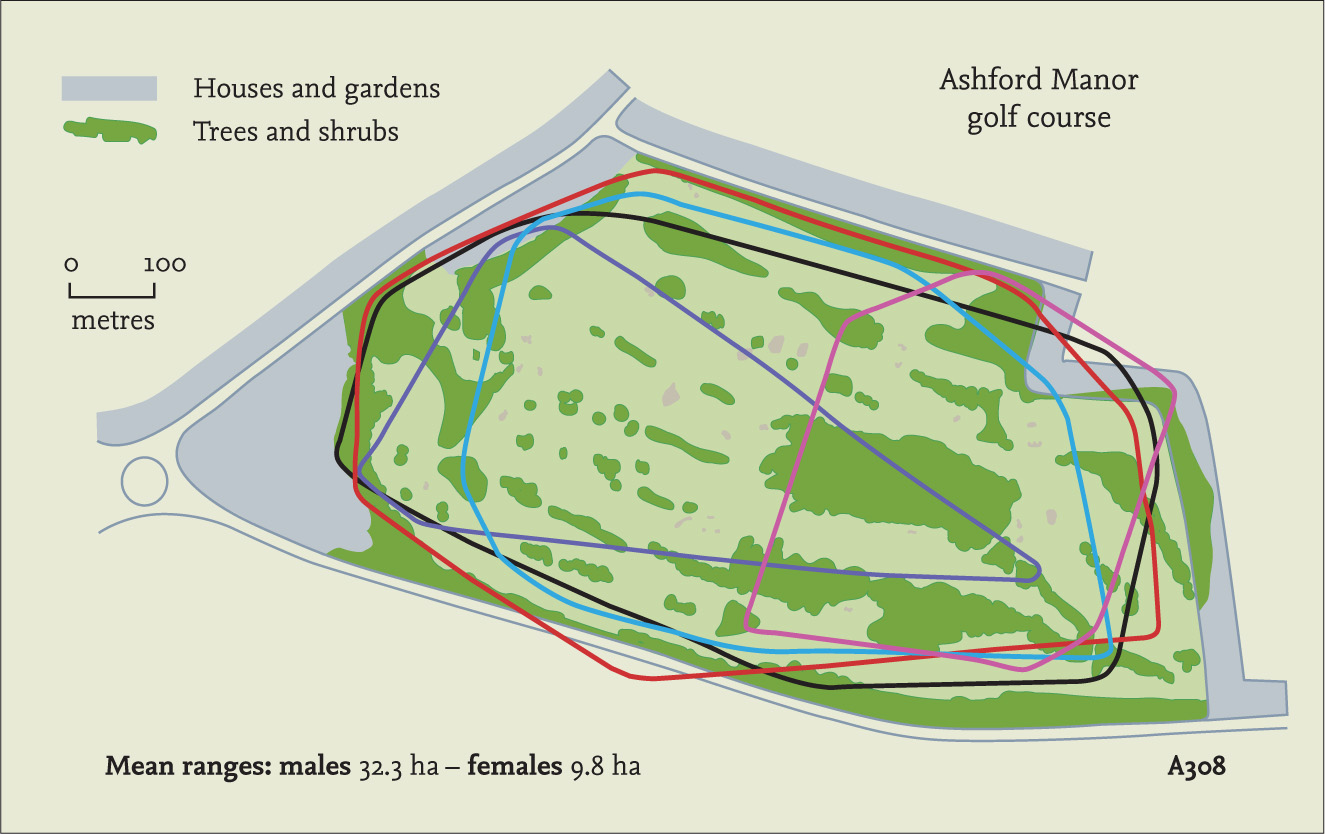
FIG 118. Areas used by five adult males in the course of one summer, showing they are not territorial. Males ranged over about three times more of the golf course than females.

FIG 119. The village of Newtown on the Isle of Wight, surrounded by farmland and isolated by tidal creeks. Here, we tracked hedgehogs that used a typical assortment of fields and gardens.

FIG 120. Penny, one of my students, helping to track hedgehogs at Newtown. By 1987, radio-tracking equipment had vastly improved and the receiver was small enough to be carried about quite comfortably.
In an attempt to obtain data in a structured manner, a group of my students radio-tracked five hedgehogs fairly continuously for three weeks on the Isle of Wight (Morris, 1988). The aim was to find each animal at least six times per night, spread across six hours. In practice, it sometimes proved difficult to achieve even this as the animals scattered widely. Nevertheless the study showed that there was considerable variation in range area used per night, twice as large on some nights as on others. It became clear the two females that were being closely monitored were generally using about 2–5 ha per night, but sometimes more than 20 ha for a very active male (Fig. 121). But the animals were not always visiting exactly the same places each night. So their cumulative home range was also estimated by adding new locations to those already obtained. After the sixth night, the animals rarely ventured far outside the area within which they had already been found. Plotted on a graph, the asymptote occurred at about 10 ha for the females, and this could reasonably be regarded as the ‘normally used area’, i.e. home range in conventional terms. The very active adult male was also closely tracked and his cumulative asymptotic home range area was approximately 37 ha, reached after the seventh night (Fig. 122). (In another study by one of my students, asymptotes showing cumulative range sizes were also all established within ten days or less.) Since the protocol followed was the same as for the study in woodland, the results can be directly compared, although sample sizes were small. It appears that about 2 ha is a normal amount of space used in a night, for females at least. Males are likely to range over a much larger area and are far more variable in the extent of their activity. Winter nesting may occur elsewhere, owing to different habitat requirements, so the total area used in a year may be larger still.

FIG 121. Range areas used each night by ‘B’, our most active male hedgehog. They varied considerably in both size and location. By contrast, the female hedgehog ‘K’ ranged more consistently each night over a smaller area and mostly in the same places each night. These differences between males and females are typical of all the studies undertaken so far.

FIG 122. After six to seven nights, hedgehog ‘B’ seemed not to enlarge his area of activity by very much, so his ‘cumulative home range’ was about 40 ha.
Most of the published information about hedgehog movements is based on radio-tracking carried out in the middle of summer. Since activity varies seasonally (Rautio et al., 2013a), figures for average home range size can be no more than a rough guide. When home range areas are determined by drawing polygons on a map, they will offer only a crude reflection of reality, not least because they often include areas that the hedgehog will never visit. They do, however, give a rough idea of how much space an animal needs and what is familiar to it, and these are factors that we need to know if we are to understand how to help hedgehogs survive in our increasingly human-dominated landscapes. All of the studies so far published, involving various habitats, indicate that hedgehogs travel more widely than previously realised and males travel further than females, although this differences persist only until the end of the breeding season.
TERRITORIALITY
Territory is defended space. Animals may defend it actively by interacting aggressively with intruders or passively by setting out scent markers to signal possession (with an implied threat of aggression towards unwanted visitors). Whilst it can often be belligerent, a hedgehog does none of these things in defence of a defined space. They do not establish and hold a territory the way that robins do in a garden. Why should they bother? Their food is plentiful and widely distributed and they are not restricted to one particular type of food or place where it might be found. They have no need to keep other hedgehogs away from their home range or nests, so what would be the benefit of spending time and energy in territorial defence? All the studies of hedgehog movements so far reported, here and abroad, show that individual hedgehog home ranges overlap considerably, even in New Zealand where hedgehog population densities are much greater than in Britain and hedgehogs might therefore have a reason for defending resources within their home range. In Finland too, hedgehog home ranges overlap despite living in very different habitat and climate (Rautio et al., 2013a).
Male home ranges often encompass those of two or three females, for understandable reasons, but there is no attempt made to guard mates or repel alternative suitors. Male home ranges also overlap the ranges of other males, sometimes completely. Instead of defending a personal territory, hedgehogs appear to have a different form of social organisation in which they may sometimes defend the space that immediately surrounds them at any one time, but not an extensive area of habitat. Adult males change their daytime resting place frequently, allowing them to range over the largest area possible in search of females, and simply avoid contacting each other. Mutual avoidance allows non-simultaneous use of the same areas by several individual hedgehogs as they forage and behave in a sexually promiscuous manner (see Chapter 7). Despite not defending a territory, some hedgehogs do show a marked tendency to remain in the same general locality year to year, and the same recognisable individual may be a faithful visitor to the same garden for many weeks or even years. Conversely, every attempt to mark hedgehogs and keep a record of which ones are found again has identified a significant proportion, perhaps 20 per cent that are seen once and never again. They are often young males. It is as though the population includes nomads of no fixed abode or maybe they are the ones that engage in long-distance dispersal movements. We still have much to learn.
MOVEMENTS: DISTANCES TRAVELLED
Home range is a concept not a definitive entity and a quest for accuracy is beguiling but perhaps futile. Moreover comparisons between habitats and different studies will not be valid unless time is taken into account and a robust protocol adopted governing the minimum number of location fixes used in subsequent analyses. But in our Isle of Wight study, estimated sizes of home ranges were strongly correlated with the total distance travelled per night, offering the prospect of using these calculations as another measure of activity. Over three weeks, the females generally and consistently travelled about 1 km per night (Fig. 123). The very active adult male managed more than 3.14 km on one occasion, taking him nearly seven hours, almost the entire period of darkness (Fig. 124). The following night he did almost the same distance again, but over a slightly different area (Fig. 125). Again a comparison may be made with the woodland hedgehogs, where females averaged about 630 m per night and the male 868 m (and a maximum of 1,138 m). Comparable figures for hedgehogs in Regent’s Park (London) in 2014 and 2015 were 892 m per night in males and 821 m per night for females (Nigel Reeve, pers comm.).

FIG 123. Distances travelled by male ‘B’ varied each night between about 600 m and 3,000 m. Typically, for female hedgehogs, ‘K’ travelled between 600 m and about 1,200 m each night.
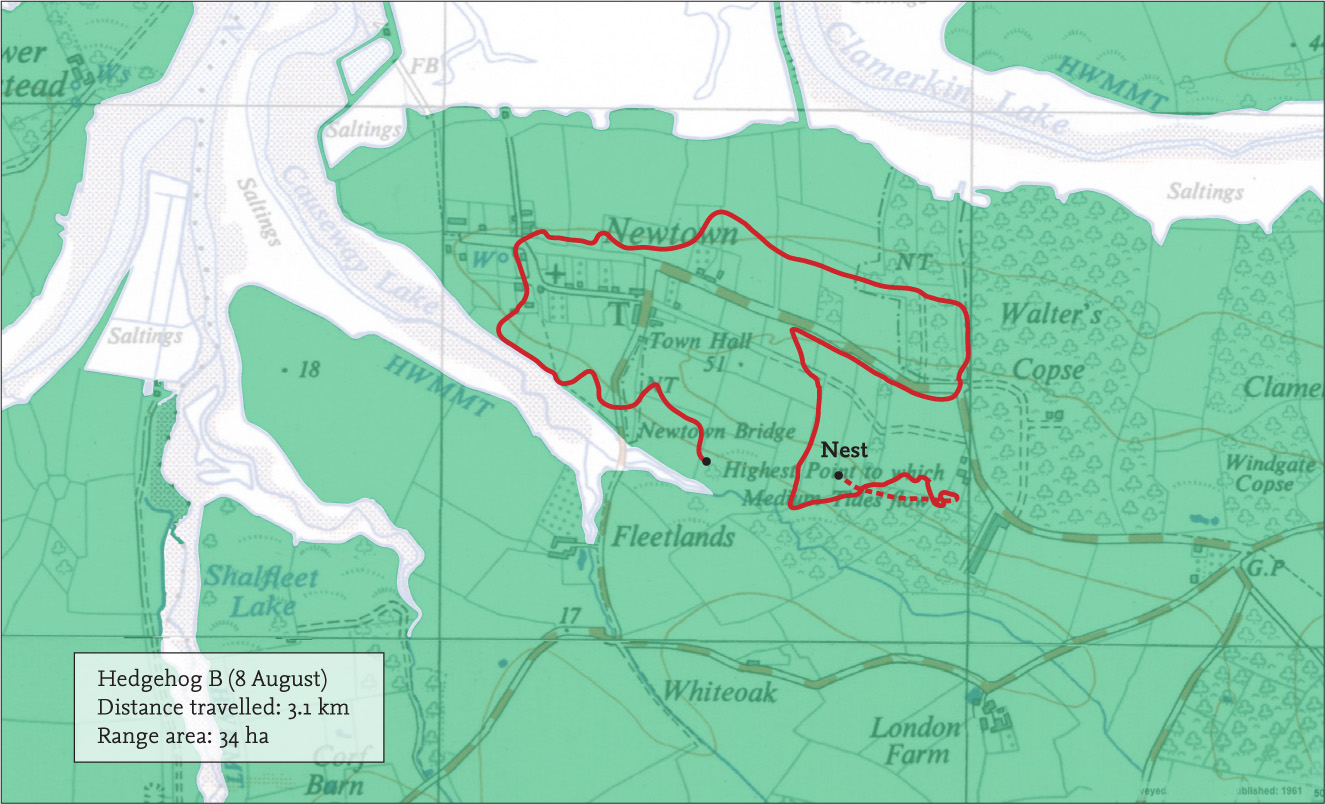
FIG 124. A ‘trackogram’ for ‘B’ on his most energetic night out, travelling a distance of over 3 km and ending at a different nest to where he started out. (the dotted line represents his track between the known site of his daytime nest and the first point where we managed to catch up with him after he set out from it before it was fully dark).

FIG 125. The following night, he set off early from his nest and travelled almost as far again, ending at yet another nest on the outskirts of Newtown village.

FIG 126. Locations of eight nests used by male ‘B’ in three weeks, showing that he was able to roam more widely in search of females than if he had returned to the same nest at the end of every night. It is also clear that he was able to find again nests he had used previously.
These figures and the consistent difference between males and females are in line with what Nigel had found on the golf course, suggesting that there is nothing particularly misleading about his data despite the artificial nature of the habitat (Fig. 127). Many subsequent studies have also confirmed that males move around more widely than females, especially adult males during the height of the breeding season. The distances travelled reported in the literature are generally similar, but always with a lot of individual variation among the hedgehogs. At Hawkes Bay in New Zealand, distances travelled averaged 908 m per night (ranging between 477 and 2,264 m) and in England Hans Kruuk found hedgehog footprints on sand exposed at low tide, indicating travel of over 3 km during a tide cycle lasting probably less than eight hours. So hedgehogs do move about a lot. We could even compare their efforts with human activity. If a hedgehog 20 cm long covers 3 km in a night, it has travelled 15,000 times its own length, equivalent to a human journey of 22 km or 14 miles.
However, calculations of how far hedgehogs travel in a night are based on adding the straight line distances between consecutive position fixes. But hedgehogs don’t walk in straight lines for very long. They wander and dither about in their search for food. So if an animal is only located six times in a night, its total distance travelled will appear to be one figure, but if it is located 20 times, that figure will be greatly increased owing to all the extra data points that are included as they zig zag about. More position fixes mean more accuracy in defining what the animal is doing. But following one animal closely to achieve this accuracy means a sample size of only one, as it becomes difficult or impossible to get comparable data from following additional animals at the same time. One of my students attempted to clarify this by using a spool of very fine thread attached to a hedgehog’s back. As it wandered about, the line snagged in grass tussocks and twigs so the actual track followed could be measured in daylight next morning, a technique known as ‘spooling’. This showed that the actual distance travelled by the animal was 40 per cent greater than computed as a straight line between position fixes, even when these were taken at only five-minute intervals. But a reasonable-sized spool will not contain enough thread to leave a trail for the whole night, only for a short sample of the activity period.
So, even figures for distance travelled per night have to be taken as indications rather than definitive statements. These limitations apply to all similar studies of animal movements of course, although they are rarely spelt out when numerical data are published. Nevertheless, computed distances travelled are useful as they do indicate the minimum journey length and they are closely correlated with spooling distances, providing a useful index of distance travelled for comparative purposes. Distance travelled will closely reflect energy expenditure, a parameter that is grounded in biological reality. The calculated size of a notional home range will be affected by the geometry of the position fixes used and may be distorted by inclusion of habitat patches that are never visited (a lake, for example). Nevertheless, estimations of home range are useful, especially when comparing behaviour in different habitats, and vital when we start to discuss how large an area is needed to sustain a secure hedgehog population.
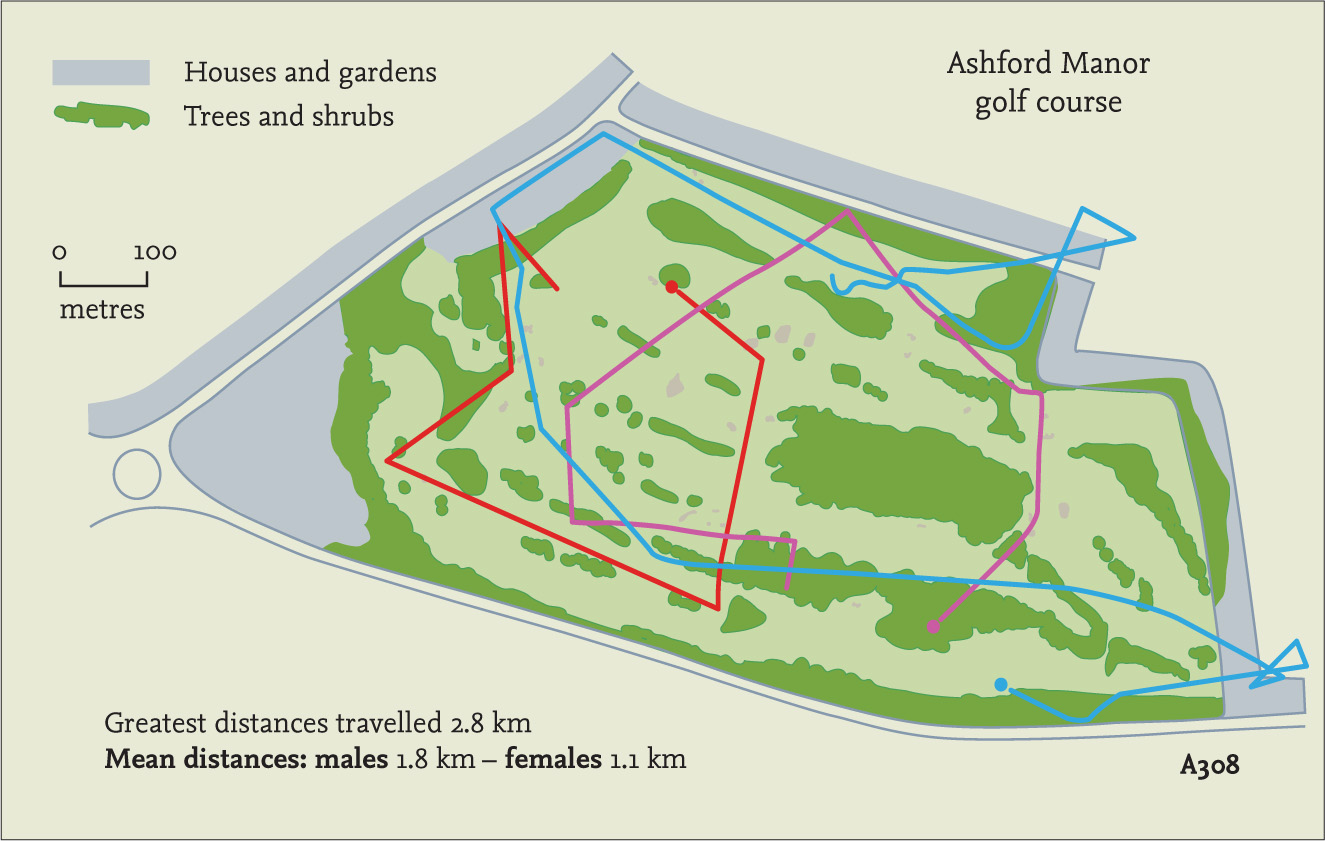
FIG 127. Nigel Reeve’s study showed that male hedgehogs (three examples shown here) travel further each night than females, and further than anyone had previously imagined likely.
With so much variation between different habitats and among individual animals, perhaps ‘precision’ has little meaning. What we need to know is approximately how big an area does a hedgehog need and how far does it normally travel in a night. It turns out that hedgehogs require much larger areas, and travel far further, than anyone had previously imagined. We also need to know how the hedgehog uses that habitat in order to improve its suitability, as suggested in Chapter 13, or at least to avoid compromising the way that the animals can move about to feed and mix with each other (Baaker et al., 2014). Without these studies, imperfect though they may be, we can have no idea how to set about helping hedgehogs to survive in an increasingly challenging world.
GPS TECHNOLOGY
All these studies of hedgehog movements and activity relied on conventional radio-tracking, with people following the animals in the field at night. During the past 30 years, huge improvements in computers and software have enabled far more sophisticated forms of data analysis which help to identify key areas within a home range, but there have been few attempts to apply new analytical techniques to the study of hedgehogs. This is partly because the old problems still remain, crucially including the difficulties and cost of people doing fieldwork at night (Fig. 128). A solution seems to lie in the employment of of modern Global Positioning Satellite (GPS) tags that store geographical positions computed utilising signals from the satellites used by satnav machines in cars. Periodically, data from the tag can be downloaded (or transmitted) containing a summary of locations together with the times those position fixes were obtained. The animals can go on about their business day and night and the investigator can work more normal hours in a nice, comfortable office. These devices have been used very successfully on birds to map migratory flights, so maybe this is a way forward for the study of hedgehogs?

FIG 128. The radio-tracking methods we used required long, lonely and uncomfortable hours of stumbling about in the dark, matching hedgehog time with our own. This is difficult to reconcile with the need for daytime activities too. The alternative of automated tracking using GPS technology has considerable appeal!
GPS tags on hedgehogs were first tried in New Zealand in a study of potential predators of rare ground-nesting birds (Recio et al., 2011). They eliminated the need for observers to follow animals closely in the dark or triangulate them to establish their locations. Position fixes could be gathered every five minutes, not just ‘once an hour with a bit of luck’, allowing a much more precise analysis of an animal’s movements. The study used 27 hedgehogs, but each was followed for only four to eight days because of limited battery life in the GPS tag. Despite their rather large backpacks, the hedgehogs seemed to behave normally and many of them put on weight during the study, suggesting that they were not impeded by the additional burden of their tag.
The main problem with GPS tracking, especially with hedgehogs, is that if the animal goes behind a building, down into a ditch or even under wet vegetation, they can lose the signal from one or more of the vital GPS satellites, resulting in a false position fix. It may then appear that the animal makes an occasional long detour and then quickly seems to come back again as normal contact is resumed, having apparently made a lengthy excursion at high speed. Without ‘ground truthing’ to check exactly where the animal really is in comparison with what the GPS satellites suggest, one cannot be fully confident of the results obtained. This New Zealand study, for example, suggested that some hedgehogs had surprisingly large home ranges, up to 95 ha, but had they really been to every point that the system said? One way to avoid this problem is to use an algorithm in the analysis software that rejects any position fix that is an improbable distance from the previous one, based on normal speeds of travel.
For birds and mammals that live in open habitats, this technique undoubtedly works well and yields massive amounts of valuable information, but hedgehogs are a different matter. A brief test of GPS tracking tags on hedgehogs in Norfolk generated unsatisfactory results, partly as a result of the animals losing contact with the satellites. A more thorough test was carried out by Lee Glasby and Richard Yarnell in Nottinghamshire. Despite some technical difficulties, most of their position fixes were successful, the failures being mainly around dawn and dusk when the hedgehogs were probably hidden away in their nests, screened from the satellites. After five days or so, the cumulative home range for each of nine hedgehogs was not significantly different from the results obtained using conventional radio-tracking (See Table 11).
TABLE 11. Comparison of average cumulative home range after a few days revealed by GPS tracking and conventional radio-tracking (from Glasby & Yarnell, 2013).

None of this comes cheap, but the GPS technology offers more detail at lower cost, without disturbing the animal’s activity and without the problems of working at night. However, the duration of such studies is limited by battery life, less than eight days in the Nottinghamshire study. GPS tags can be made to operate for longer by gathering less information or having larger batteries, but battery life may improve with advances in technology. Habitat type has a significant influence too. Performance and accuracy of the GPS tags was reduced in vegetated and built-up habitats and also when the tags were placed within hedgerows to see the effect of screening vegetation. Further use and field-testing of GPS tags should enable a protocol to be developed that identifies the likely ‘rogue position fixes’ to eliminate most of them. It appears that this technique would still have limited value for a study of nesting behaviour or activity in urban gardens, for example, because of all the obstructions that might screen or reflect signals from satellites. Maybe other forms of automatic tracking will be developed for use in difficult habitats.
A WIDER RELEVANCE: THE REGENT’S PARK STUDY
Investigating hedgehog movements is not just a matter of academic curiosity. Regent’s Park supports the last surviving hedgehog population in central London and GPS tags have been used there by Nigel Reeve, John Gurnell and colleagues to find out how hedgehogs move about within the park (see www.royalparks.org.uk/managing-the-parks/conservation-and-improvement-projects/hedgehogs). They discovered that shrubberies and flower beds are favourite places for nesting and foraging, but the area most popular with the hedgehogs is the car park for London Zoo. More precisely, it is the surrounding rough grassy areas, which are used by a quarter of the hedgehog population for foraging and nesting (Fig. 129). It is also where engineers planning to build HS2 (the high-speed rail link between London and the north) want to create a depot for large equipment, serviced by heavy lorries. This could sound the death knell for the Regent’s Park hedgehog population because so many of them use that area. The issue received national attention when a Parliamentary committee debated the future of the HS2 project. To the visible irritation of the HS2 lawyer, hedgehogs seemed to be getting in the way of a key British infrastructure project costing billions of pounds! The use of this area is opposed by the Zoological Society of London (ZSL), whose car park for visitors is likely to be taken over. ZSL also raises funds and campaigns vigorously to help endangered species around the world. It would be embarrassing if hedgehogs became extinct just outside their front door. There ought to be ways to create some similar habitat elsewhere at the edges of the park, but time is short and free space is very limited. Meanwhile, the issue has usefully drawn public attention to the fate of the hedgehog nationally and within Regent’s Park. It also shows the value of tracking studies that can tell us what goes on in one of the last hedgehog populations left in central London and, by implication, in many similar urban sites elsewhere.

FIG 129. The excellent hedgehog nesting and foraging habitat adjacent to the car park at London Zoo.
Another important outcome of the Regent’s Park study with wider implications was the revelation that hedgehogs rarely visited the football pitches that constitute nearly a quarter of the total area of the park. Based on previous studies of hedgehog activities, these look like ideal foraging areas with healthy short grass, but GPS fixes show that they are conspicuously ignored by the hedgehogs. Moreover, there is increasing evidence of similar problems elsewhere. Why should that be? One possibility is that the soil (heavy London clay in the case of Regent’s Park) has become so compacted that it no longer supports worms, beetle larvae and the many other foods that hedgehogs normally expect to find in short-grass habitats. After all, a footballer thrusts his whole weight into the ground on a few studs under his boot, generating pressures that may exceed 15 kg per cm2. Soil compaction is inevitable, especially if the pitch is used multiple times every week for the whole season (Fig. 130). Efforts are made to increase aeration of football pitches by driving spikes into the soil at the end of the season, but this might be making things worse. The spikes leave deep holes, but no soil is actually removed. The particles are simply thrust sideways into the adjacent soil, compacting it further. Air may be able to enter the soil down the holes, but for soil-dwelling larvae and other organisms, the soil particles are rammed so close together in the upper layers of a football pitch that movement and respiration by the soil macrofauna will be impeded. This must severely compromise the abundance and productivity of many invertebrates that hedgehogs need for food – worms and beetle larvae, for example. Treatment of ‘sports turf’ like this will also often include the use of chemicals to kill the insect larvae that damage grass roots – the very items that are high on the list of prey that hedgehogs need to eat (see Chapter 9) – although this does not appear to be a problem in Regent’s Park, where chemical treatments have not been used for many years.

FIG 130. The football pitches in Regent’s Park, London, appear typical of many across the country, but although they have not been treated with pesticides for many years, the hedgehogs still seem not to use them for feeding.
So the Regent’s Park study is not just about hedgehogs. It also helps to alert us to wider issues concerning the use and management of urban green spaces. Many studies of hedgehog ecology in the 1990s and earlier, including my own, found they were common on open fields, golf courses and parkland, leading to an assumption that grassy turf was a preferred habitat. That may no longer be the case as a consequence of heavy use and changed management practices that include the introduction of deadly efficient pesticides (see Chapter 10). Studying hedgehog movements may be generating information that has a much wider importance and there is a clear need for more detailed research into the relationship between hedgehog movements and the types and amounts of food potentially available to them, especially in amenity grasslands that are managed in different ways. Perhaps many of the parks and grassy areas of our towns and cities that we presume to be important wildlife reservoirs in built-up areas are becoming less and less viable as refuges without us realising what is happening. Maybe we need to find alternative ways of managing amenity grasslands lest our green spaces become just green deserts, although it may already be too late for some. Isolated areas of urban green space, especially those managed as sports turf, may no longer offer key foraging opportunities for local hedgehog populations and many bird species too. Under a more relaxed and supportive form of management, these habitats might one day recover. The birds could then fly back to recolonise the sites, but not the hedgehogs.
ARABLE FARMLAND
Also important are questions about how hedgehogs use the habitats available in today’s countryside because this relates directly to issues of how we might help to conserve them and assist their survival in human-dominated landscapes. Most fieldwork on hedgehogs has been carried out in grazed or mown grasslands for practical reasons and also because it is normally a good habitat for hedgehogs. But what about arable farmland? Is that a good place for hedgehogs or is it a hedgehog desert? The question matters because a quarter of Britain’s land area is now managed for arable crops. The land is ploughed and regularly doused with pesticides, both actions being aimed at eliminating the very things upon which hedgehogs feed. Can hedgehogs use this habitat or are they restricted to the field margins? If the latter, then hundreds of square miles of Britain are no use to them. An exploratory study tracking hedgehogs on the fringe of Windsor Park revealed that they used only the margins of maize fields and did not forage in the crop itself. Other attempts to study hedgehogs in arable habitats have been hampered by the difficulty of catching any of them at all, suggesting that arable farmland is not a good place for hedgehogs to be. In two cases, we had to resort to borrowing randy male hedgehogs from animal rescue centres and radio-tracking them on arable land as they sought prospective mates on our behalf, fortunately with some success.
One of those studies (Hof & Bright, 2010) investigated hedgehog use of arable fields in Norfolk and found that they kept very much to the edges, rarely more than five metres from the field boundary. This could be due to hedgehogs being unwilling to venture too far out into open areas for fear of being attacked by a predator. In this study, and others, they appeared to be very wary of using areas frequented by badgers. But this may not be the only explanation. Sampling the soil for invertebrates showed that worms and beetles were more abundant around the field margins than out in the crop area. The best fields had a strip of rough grassland between the crop and hedge offering better foraging prospects (Fig. 131). Moreover, there are no suitable nesting sites or materials out in the crop. Altogether, this suggests that large areas of Britain’s arable farmland are sub-optimal habitat at best and even perhaps no use to hedgehogs at all. When choice of habitat was measured (including woodland and amenity grassland), hedges and field margins scored best and cropped areas worst. In fact, only 6.5 per cent of position fixes in the Norfolk study were on the cultivated land. A study at two sites in Norfolk and two others in Yorkshire (Pettett et al., 2017a) found arable land and woodlands were the least-used of the available habitats by a sample of 78 hedgehogs. In the first such study in Ireland (Haigh, 2011), arable land proved rather more attractive to hedgehogs, possibly linked to an abundance of molluscs not seen elsewhere. This may reflect a difference in farming methods, differential use of molluscicides, for example, or seed drilling instead of deep ploughing (affecting slug survival rates). But in Britain, it does appear that the major historical change from pastoral farming to arable during the twentieth century is likely to have rendered much of the countryside unsuitable for hedgehogs, causing a reduction in their overall numbers. The radio-tracking studies show that agri-environment subsidies for farmers that encourage wide grassy margins and thick hedgerows can do a lot to stop the hedgehog’s decline and will benefit many other species too.

FIG 131. Radio-tracking studies show the importance for hedgehogs of ‘wild’ field margins in arable landscapes.
URBAN HEDGEHOG ECOLOGY
Suburban gardens are an important habitat for hedgehogs and may become even more so in the future, both as a refuge from badgers and as an alternative to trying to live in arable farmland, which is one of the largest available habitat types in the countryside. Householders who have marked their hedgehogs often report that ten or more visit their garden, implying a high local population density. This may in part be due to the attractive and supportive effects of bowls of food, often set out specially to attract hedgehogs. In France, the population of hedgehogs in towns is said to be up to nine times greater than in the French countryside (Hubert et al., 2011).

FIG 132. Recent studies suggest that gardens are becoming a vital refuge for future hedgehog populations, given the intensification of farming and prevalence of badgers in the countryside.
Although hedgehogs are a familiar sight in many of our town gardens, we know surprisingly little about their ecology in this type of habitat. For example, several people have written to me with details of their ‘attendance register’, recording which marked animals come each night to a food bowl in the garden. Sometimes as many as 20 individual hedgehogs will be seen during a single summer. One of my correspondents, a Mrs Margaret Larkman, recorded the visits of about 30 different hedgehogs to her garden in a single year, but none stayed as ‘regulars’ for more than about two months. Her animals showed a 1:1 sex ratio and a general indifference regarding time of day, rain and temperature. Another correspondent, Mrs Anne Ursell, experienced a similar visiting pattern, with some hedgehogs coming regularly, then being absent for long periods before returning. Three of the 18 hedgehogs visiting my friend Brian Barton’s garden in 2012 continued to do so until well into November and one was still coming in January the following year. This helps confirm the principle explained in Chapter 6 that hibernation is flexible and need not necessarily happen if food is still available.
TABLE 12. An analysis of the attendance records of 18 animals visiting a garden in 2012, at least 5 of whom did so for more than 3 months (data kindly provided by Brian Barton). Many individuals came night after night, but then were not recorded for several nights before reappearing. Regular visitors first seen in the spring seemed to disappear and were largely replaced by apparently new arrivals from July onwards. Three animals came 7 times or less, 6 others were recorded on 40 or more occasions. Records like this are of course ‘minimal’, in the sense that the observer normally goes to bed by midnight! So some animals that are apparently absent may simply have arrived later, adding to their number of nights in attendance. Nevertheless, some seem to discover the food and come on more than 75 per cent of the nights during the period of their visits; others arrive for less than 50 per cent of the feeding opportunities on offer.

A general pattern in many of these observational studies is that as the season progresses, more new animals turn up, but the ‘regulars’ stop coming. Why, having discovered a food bowl put out specially for hedgehogs, do they visit every night and then abandon the habit of coming to a good food source? Maybe they were killed, but some of them actually reappear after an absence of a few days or weeks. Where have they been meanwhile? Why go elsewhere? Why come back? A significant proportion of marked animals are seen once or twice and never again. Why? Where do they go? Are there permanent nomads in the population that just keep moving from one place to another. If so, why does this happen?
We need to know much more about urban hedgehogs, but (as usual!) studies are fraught with difficulty. Householders don’t like to have people creeping about their garden in the middle of the night, numerous fences get in the way, dogs become excited and start to bark, the police are called out. Worse still, radio transmitter signals are screened or confusingly deflected by walls and fences. Gardens harbour countless nooks and crannies into which a hedgehog can vanish, hiding both itself and its radio signals. Nevertheless, there have been a few investigations of urban hedgehogs in Britain.
One of mine (Morris, 1985) addressed the obvious question: To what extent do food bowls put out to encourage hedgehogs influence their behaviour? Food availability is likely to affect territory, home range, social behaviour and other key aspects of an animal’s life. One might expect a regular, localised supplementation of natural food to result in higher population density and reduced home range size, since the animals will no longer need to travel so far in order to get enough to eat. They will surely seek to minimise the effort of foraging and travel no further than necessary, but this may disrupt natural spacing behaviour. The hedgehogs might remain in a small garden because there is no need to go anywhere else, just as the good people who put out this extra food hope and imagine, but do they stay or commute? There are important welfare issues involved too. Maybe hedgehogs will become reliant on the extra, easy food and suffer when people go away on holiday. Perhaps the animals will forsake natural foraging in favour of easy and unnatural food, then suffer from dietary deficiencies. In other words, this well-meaning attempt to help hedgehogs may do more harm than good, hence the need for a study.

FIG 133. Mr and Mrs Wall’s ideal hedgehog garden with lawn, flowerbeds, compost heaps and a regularly filled food bowl on the patio that we monitored in a study of hedgehog behaviour and supplementary feeding.
FIG 134. The dotted line represents the shortest straight line distance from several hedgehog nests to the food bowl. The dots indicate the actual distance covered before the animals arrived there. Clearly they knew where it was, but did not abandon natural foraging to go there immediately.
During our investigation, at least 11 different hedgehogs visited the study garden where bread and milk had been put out regularly for seven years by the kindly owners, Mr. and Mrs. Wall (Fig. 133). Six of the animals were radio- tracked closely, two of whom regularly travelled to the garden from more than 500 m away, even though they did not need to, and none actually nested in the garden itself, to the dismay of the kind people who had been putting out the food for so long. One might expect that the hedgehogs would head for the food as soon as possible in order to get there before it was all eaten, but they often took nearly twice as long as necessary to reach the garden and sometimes did not come at all. One that needed to travel 196 m from its nest to the food bowl actually travelled over 1.2 km before arriving. Another hedgehog spent four hours getting to the garden which could have been reached in fifteen minutes (Fig. 134). These animals were feeding normally on the way and again later in the night, after visiting the food bowl. They foraged widely in nearby gardens and on the adjacent golf course, seeking natural food (Fig. 135). So the food bowl was only providing a supplement to their natural diet and did not apparently deter them from feeding normally, even the blind animal ‘Number 28’. They ate a lot. The maximum amount taken by one hedgehog in one continuous feed was 94 g, and even the average meal was 54 g. Sometimes a hedgehog would return for additional feeds later in the night. If it is typical, this level of consumption must represent a huge dietary benefit for urban hedgehogs in areas where supplementary food is offered.

FIG 135. A typical ‘trackogram’ showing how a hedgehog would forage on the golf course, then detour into the garden to feed at the bowl, before returning to normal foraging back on the grassy areas of the golf course.

FIG 136. The trackogram for ‘Number 28’, a blind hedgehog that travelled across the golf course to feed at the bowl before returning to forage in the grassy areas.
Each garden did not have its own exclusive group of visitors, a disappointment to the householders concerned. The hedgehogs certainly did not defend their territory or the precious food bowl that lay within it; instead, they wandered widely and freely, criss-crossing each other’s paths. Few disputes were observed, even at the food. Hedgehogs entered and left gardens by the same shared routes and there were often three or more present simultaneously in one garden within 10–15 m of each other. A regularly filled food bowl would appear to be a valuable resource, well worth defending, yet two or more hedgehogs often fed simultaneously or in quick succession. Male hedgehogs still travelled substantially further than the females, confirming that their greater activity is not food-driven. The difference in behaviour between males and females was not disrupted by the presence of supplementary food. Nor was there evidence that the presence of extra food diminished hedgehog wanderings to any significant degree. Minimum distances covered in a night ranged up to 2.3 km, exceeding 1 km on at least 12 animal-nights. The hedgehogs sometimes went on to other gardens with food bowls available or they may have been there first. The nightly consumption of natural food may explain why only a small meal was sometimes taken, and also suggests that the hedgehogs had not become addicted to the bread and milk that was on offer, nor even heavily dependent upon it (and certainly not killed by it after seven years!). Average speeds were lower after visiting the bowl, probably as a result of more and longer resting periods. The bowl was usually empty by about midnight, allowing plenty of time for the hedgehogs to continue feeding naturally somewhere else. Leaving the bowl empty for a few nights, simulating our absence on holiday, revealed that the hedgehogs were not totally dependent upon it. They did not sit about waiting for it to be filled and they readily fed elsewhere. Even the blind one, ‘No. 28’, managed well, coming in from the far side of the golf course each evening to visit the food bowl (Fig. 136). A short study in my own garden a few weeks later confirmed similar patterns of behaviour towards the provision of extra food (Table 13).
Independently of this study, a lady who had marked her hedgehogs elsewhere reported that she had several hedgehogs coming regularly before she went on holiday. She left the food bowl empty for two holiday periods lasting a week each. Five of the animals came again afterwards and three that did not had only been rare visitors anyway. At least ten different hedgehogs came to her food bowl between May and September. Some were very regular visitors, including one who came frequently over a period of 5½ months, but was often not seen for up to ten nights before reappearing. Two animals who knew the food was available and had eaten it more than once before, were absent for 33 and 40 nights respectively before coming again. These observations provide further evidence of non-addiction and non-dependency. Altogether, it seems that natural hedgehog behaviour is not disrupted by the provision of extra food. They appear to accept it as a useful supplement to their normal diet and in periods of dry summer weather, access to moist food may be a valuable aid to survival, especially for females attempting to raise young. The extra food put out in gardens for pets and wildlife may be a contributory factor in sustaining high population densities of hedgehogs in urban areas.
TABLE 13. An ‘attendance register’ recording the occasions when six radio-tagged hedgehogs visited a garden food bowl: before, during and after a period when it was left empty. The food was watched from dusk until about midnight on 17 nights (Morris, 1985).

Roads are another issue for urban hedgehogs. One radio-tracking study (Rondinini & Doncaster, 2002) found that they avoided crossing large roads, especially motorways, and the only road casualty hedgehog I have ever seen on a motorway was actually on a slip road, not the main carriageway. This raises the prospect that major roads may be dividing up hedgehog habitats, with consequent threat to the survival of the small, fragmentary populations that remain. Information gained from radio-tracking studies has been used as the basis for generating computer models that predict likely hedgehog behaviour in respect of roads. The simulations suggested that large and small roads would be treated the same by hedgehogs, but in reality they avoid crossing large roads more often than predicted, indicating that bigger roads are treated as barriers during their nocturnal activities. Barriers that fragment habitats, especially in urban areas, may be one of the key problems for population integrity and survival (see Chapter 8). Road verges were the lowest-ranking habitat in preference analyses, while playing fields and urban gardens were preferred habitats (Rondinini & Doncaster, 2002). But what if hedgehogs are prevented from reaching those preferred sites by roads, garden fences and other barriers?

FIG 137. A marked hedgehog, one of at least seven, feeding at a food bowl on the patio while my father watches TV.
A radio-tracking study of urban hedgehog ecology in Bristol (Dowding et al., 2010a) also suggested road traffic was a threat to urban hedgehog populations. The Bristol hedgehogs frequently crossed roads, an obviously hazardous activity, but avoided actively foraging beside them or along their verges. Unlike their behaviour in quieter habitats, the hedgehogs were significantly more active after midnight, when there was a substantial reduction in road traffic and in the number of pedestrians walking about. The hedgehogs liked to use the large gardens of semi-detached and terraced houses, but then faced the threat of attack by badgers. Females seemed to stay out of the larger gardens where badgers were more likely to occur, although the males were bolder.
This and other studies of urban hedgehogs, including my own, confirm that the average garden is of inadequate size to support a hedgehog population and that they need to have access to a number of them, or to larger areas such as recreation grounds, cemeteries and allotments. In Denmark, Sophie Rasmussen found individual hedgehogs visited ten or more gardens, highlighting the need for them to be connected by accessible routes and not isolated by hedgehog-proof fences. This issue is explored further in Chapter 13. Studies of urban hedgehogs have drawn attention to the importance of green spaces in suburbia as reservoirs for many animal species, but also highlight the need to consider how those sites are used and managed if they are to continue to provide vital support for wildlife.
CONCLUSION
The hedgehog is normally active after dark and appears not to indulge in typical mammalian behaviour such as play, social activities or territoriality. Almost all of its time is spent in routine systematic foraging, which makes it all the more peculiar that occasional bizarre actions occur, including self-anointing and manic running in circles. Radio-tracking has generated much essential information about hedgehog movements, although the results may not be comparable between studies and their interpretation is open to discussion. When describing ‘home range’ for this and other species, it is important to distinguish between the area used in a typical activity period, the larger area with which the animal is familiar, and the total range area used in an entire year (including winter). This is frequently not done and figures given in print may not be directly comparable and need to be accepted with caution. Hedgehog activity differs according to local conditions and it appears that arable farmland is a challenging habitat for hedgehogs that now covers large parts of Britain. Radio-tracking studies have established that hedgehogs range more widely than expected, raising concerns about habitat continuity and also about the possible unsuitability of intensively managed sports turf, amenity grassland and other urban green space for this species and many others. Suburban habitats are becoming increasingly important as refuge areas for hedgehogs, especially where extra food is put out for wildlife in many people’s gardens. This does no harm and may be very supportive of urban hedgehog populations, but we need to know more about hedgehog ecology in built-up areas as urbanisation expands inexorably.