
THE EARLIEST KNOWN BIRD, Archaeopteryx lithographica, was essentially a feathered, flying-gliding dinosaur that lived in the late Jurassic period, some 150 million years ago. It is known from a fossil record in southern Germany, and its discovery in the early 1860s was quickly taken as confirmation of the idea that birds evolved from reptiles. The skeletal structure of this half-bird, half-reptile suggested that it was probably more a glider than a flyer, but most significantly its rounded wings and tail were feathered. An even earlier bird-like fossil, Protoavis, has been found in late Triassic deposits in Texas, but its status is controversial. Because of the hollow structure of bird bones and their propensity for being crushed, very few early fossils remain, and the evolutionary trail that leads through the Cretaceous period, from dinosaurs to the birds we know today, is unknown.
During the past 25 years, however, knowledge of the evolutionary path of birds has undergone remarkable revision with the aid of cladistics – a method of classification in which organisms are grouped together on the basis of similarities due to recent origin from a common ancestor – and molecular phylogeny. These analytical tools now place birds not as evolving during the Triassic period (254-206 million years ago) from basal archosaur reptiles but directly from a subgroup of the Dinosauria, with their closest relatives being modest-sized dinosaurs such as the dromaeosaurids and troodontids. The evidence for the relationship between the birds and these theropod dinosaurs is based on the skeletal structures in the forelimbs, pelvis, hind limbs and skull.1 Four of these meat-eating theropod dinosaurs, with bird-like bones and traces of feathers, were discovered in Liaoning Province, China, in the late 1990s. They had the forelimbs of a primitive bird and the tail of a dinosaur (Fig. 1).
FIG 1. Unnamed oviraptorosaur, one of the Chinese theropod dinosaurs that may be the ancestors of modern birds. (Image from an article by Christopher Sloan, National Geographic Magazine, November 1999. Reproduced with permission)
The true birds of the Cretaceous (145-65 million years ago) were the Neornithes (modern birds), flapping or running around the shallow tidal waters of the ancient oceans. They included the ‘fish bird’ or Ichthyornis, a small, tern-like seabird, some 20 cm in length and an efficient flying machine, and a large flightless diver called Hesperornis, up to 160 cm in length, that had a lower jaw resembling that of mosasaurs, the swimming reptiles (Fig. 2). Only the lower jaw had teeth. Its wings were vestigial but it was apparently a strong swimmer with powerful legs. Hesperornis was effectively a well-developed type of water bird. It is clear that by this point in geological history the avifauna was well diversified.
The earliest identifiable anseriform fossils, Romainvillia stehlini and Eonessa anaticula, representing two separate, distinct and extinct subfamilies, were discovered in two widely separated geographical areas – France and the United States – and date from the late Eocene period, some 40-35 million years ago. The first known fossils of wildfowl belonging to the present-day subfamilies Anserinae and Anatinae come from the Oligocene period, 34-24 million years ago. One of these fossils was of a waterfowl, Cygnopterus affinis, somewhat similar to the mute swan, discovered in Oligocene deposits in Belgium.2 But it is not until the more recent Miocene period, 24-5 million years ago, that the fossil record reveals a total of 27 anseriform species, including goose bones that could be ascribed to the genera Branta (black geese) and Anser (grey geese).
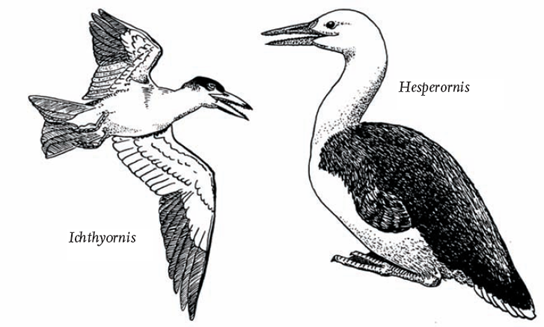
FIG 2. Ichthyornis and Hesperornis, true birds from the Cretaceous. From R. Freethy, How Birds Work (Blandford Press, 1982).
During the Pliocene period, some 5-2 million years ago, the number of species representing each genus of wildfowl increases rapidly in the fossil record, so that by the start of the Pleistocene period, about 1.7 million years ago, most of today’s wildfowl species had evolved as we know them today.
Wildfowl (ducks, geese and swans) all belong to a single large family, the Anatidae, which makes up the bulk of the order Anseriformes. According to the classification put forward by Jean Delacour and Ernst Mayr in 1945 and later modified by Paul Johnsgard in 1978, the Anatidae embrace some 151 species.3 This is the classification adopted by the British Ornithologists’ Union (BOU), and followed in Chapter 3. More recently Livezey proposed several changes to the classification of the order Anseriformes, based on cladistic analysis of morphological characters.4 This classification recognises two small families – the screamers (Anhimidae) and the magpie goose (Anseranatidae) – alongside the Anatidae; the number of subfamilies within the Anatidae is increased from three to five; and a number of fossil species are included, making a total of 173 species in 55 genera.

FIG 3. Greenland white-fronted geese in flight. These geese fly approximately 5,800 km on a round trip from their wintering grounds in Ireland and Scotland to their breeding grounds in west Greenland and back again. The spring journey from Ireland/Scotland to their staging grounds in Iceland takes some 14-17 hours. After resting and refuelling there for about a month they fly another 14-17 hours, including crossing the Greenland icecap, to their breeding grounds. Neck collars – each with a unique three-digit alphanumeric code – have been fitted to some 2,800 birds for scientific research. Much of our detailed knowledge of wildfowl comes from intensive studies using such devices, and more recently satellite tracking. (Alyn Walsh)
Despite a great diversity of size, body shape, plumage coloration, ecology and behaviour, the Anatidae are united by certain common features in physical structure, habitat and behaviour. They are all adapted for an aquatic or semi-aquatic life. The front three toes are connected by webbing – only the Hawaiian and magpie geese have partially webbed feet – and the smaller hind toe, sometimes lobed, is positioned higher. The legs of wildfowl are set far apart, and located not in the centre of the body but towards the tail end of the bird to facilitate more effective swimming and diving. When walking on dry land wildfowl appear to waddle, a necessary movement to prevent the bird from toppling over, as its centre of gravity must be shifted backwards over each leg, thus producing a mild rotating effect as the bird moves forward. In flight wildfowl have characteristically long necks and short tails, creating the impression that the wings, often sharply pointed, are set well back on the body. Most wildfowl are fast and strong flyers (Fig. 3), with a well-developed keel protruding from the sternum to accommodate the bulky flight muscles.

FIG 4. Black scoter: upper and lower mandibles open, revealing the lamellae that are used to sieve or strain solid food items from water by many duck and swan species. (Joe Blossom/NHPA)
The diagnostic feature that separates wildfowl from all other birds is the bill, generally flattened, broad and blunt, which is covered by a thin skin and tipped by a horny plate in the shape of a fingernail known as the ‘nail’, which in some species is bent over in the form of a hook. The edges of the mandibles are set with rows of horny lamellae (Fig. 4), and the tongue is thick and fleshy, covered with horny spines and papillae. The range of size, shape and nature of the bill of each species is witness to adaptive variation, a response to the great diversity of food eaten by wildfowl, ranging from grass to molluscs. Geese are essentially pluckers of vegetation, and have conical and often short stubby bills with serrated cutting edges to assist in the cutting of grasses; the shoveler, a dabbling duck, has a relatively enormous spatulate bill with well-developed lamellae along its edges for sifting, straining and extracting food from mud and ooze; the mergansers sport long and narrow bills with saw-tooth-like lamellae, which help to grip small fish underwater.
Wildfowl plumage is thick, with the feathers underlain by a dense coat of downy feathers that are loose, soft and fluffy. The mallard has approximately 12,000 feathers, including downy feathers, while the tundra swan has some 25,000, representing about 10 per cent of the swan’s body weight.5 Underneath is an additional insulation layer of subcutaneous fat, a feature generally lacking in other birds. Anyone who has roasted a goose will have been rapidly brought to an appreciation of the extent of the oil stored in this fatty coating. Wildfowl, unlike most other birds, shed their flight feathers simultaneously at the end of the summer and are generally flightless for up to four weeks. During this relatively short period they take refuge on lakes, generally safe from predators and close to their food. Many of the brightly coloured male ducks moult their body feathers twice: in late summer they moult into an ‘eclipse’ plumage that renders them less conspicuous while they regrow their flight feathers, and then later in the autumn a second body-feather moult restores their splendid plumage, readying them for the business of courtship. Most other non-wildfowl species undergo a progressive moult of their feathers, allowing them to retain their power of flight. A large proportion of the time budget of wildfowl is spent on preening and feather maintenance. To assist with the waterproofing of the feathers a large and well-developed sebaceous gland, located just above the tail, is a continual source of oil for smearing, by both bill and head, on the feathers of the preening bird.
Geese and swans share a high level of social and family organisation, generally remaining paired for life over a relatively long life. The young remain with their parents after the breeding season, migrating together to the wintering grounds, where the family generally remains intact, and returning together to the spring staging grounds and often the summer breeding grounds. If the adults fail to breed during that summer, more often than not the family will stay together for the second winter, and possibly further winters. While much bird behaviour is instinctive, geese and swans are unusual in that their young must ‘learn’ the migration routes by travelling with their parents southwards from the Arctic during the autumn and returning in the spring.
Wildfowl, ratites – the ostrich, rhea, emu, cassowary and kiwi – and a few other birds are unusual in possessing an erectable intromittent phallus, an adaptation to facilitate fertilisation in the aquatic environment. It is considered a primitive feature amongst birds.
According to Livezey’s classification, the family Anatidae consists of five subfamilies, of which only three are relevant to Britain and Ireland. The swans and geese comprise the subfamily Anserinae, characterised by large size, long necks, relatively long legs and a body adapted for walking. They have a single annual moult rendering them both flightless (for up to four weeks) and vulnerable to predators, display little plumage, structural and voice differences between the sexes, and exhibit relatively simple courtship displays. The three swans of Britain and Ireland belong to the genus Cygnus. They are all-white in adult plumage with longer necks (either as long as or longer than body length), shorter legs and larger feet than geese. There are eleven species of geese on the British and Irish lists, several of which are further divided into subspecies or races. The ‘grey geese’ belong to the genus Anser and display generally uniform plumage coloration, while the ‘black geese’ belong to the genus Branta and exhibit a more elaborate plumage with contrasting whites, blacks, greys and, in the case of the red-breasted goose, an additional colour, red.
The subfamily Tadorninae includes the shelducks and sheldgeese, with three species recorded in Britain and Ireland. The true ducks belong to the other subfamily, Anatinae. They are generally smaller than geese and swans, have shorter necks, and are better adapted for swimming and diving. Unlike the geese and swans, they have a double body-feather moult as well as the single flight-feather moult, the sexes have different plumages, and their courtship behaviour is complicated, with males indulging in elaborate displays in contrast with the low-key behaviour of the drabber-plumaged females. The ducks are further subdivided into five tribes, of which four have been recorded in Britain or Ireland. The two largest tribes are the sea ducks or Mergini, with 16 species recorded from these islands, and the Anatini or dabbling ducks, of which there are 13 species on the British and Irish lists (all in the genus Anas apart from the muscovy and mandarin ducks).
A complete classified list of all wildfowl species that have occurred in Britain and Ireland – either in a wild state or in self-sustaining populations following deliberate introduction or escapes – is contained in Appendix 1. The Livezey classification is followed in Appendix 1, although Chapter 3 retains the more familiar BOU/Johnsgard sequence, placing the swans before the geese. Appendix 2 contains a list of all other wildfowl that have occurred in Britain and Ireland as recorded in recent years by the Wetland Bird Survey (WeBS), either as escapes from wildfowl collections or as non-self-sustaining introductions.