Social networks can be difficult to understand in part because they are difficult to manipulate. We cannot give you a friend the way we might give you a placebo. But if we could somehow strand a group of strangers on a desert island and watch how they become connected, and for what purpose, we might be able to observe social networks as if we were conducting an experiment. This does not sound like something that could be done. Except that it has been done, and not by curious social scientists but by television producers.
In the summer of 2000, CBS piloted Survivor, a show that would become a sensation and spawn the reality-television craze. The premise was simple: take sixteen average Americans, chosen from all walks of life, and abandon them in the middle of nowhere to fend for themselves. Every three days, the “Survivors” would gather in a tribal council to vote someone out of the tribe. At the end, a sole Survivor would win $1 million.
The highest-rated season of Survivor took place in the Australian outback in 2001, drawing nearly thirty million regular viewers who tuned in each week to watch the social dynamics, among other things, unfold. In the first episode, contestants were forced to remain silent until they arrived on site and were placed into two competing tribes, Kucha (an Aboriginal word for “kangaroo”) and Ogakor (an Aboriginal word for “crocodile”). They had five minutes to gather their gear and start a grueling five-mile trek to their camps. Debb Eaton, a forty-five-year-old female corrections officer at a men’s prison in New Hampshire, quickly took charge of the Kucha tribe, but her initiative would be her undoing. Her fellow tribe members became so annoyed with her bossiness that she was the first person voted out.1
Some Survivors quickly made friends, while others plotted to eject fellow tribe members. For example, Jerri Manthey, an actress from Los Angeles, accused Kel Gleason, an U.S. Army Intelligence Officer from Fort Hood, Texas, of smuggling in beef jerky and refusing to share it. Tribe members searched his belongings and found nothing, but because they were already suspicious of his background and trustworthiness, the damage had been done. Kel was unanimously voted out at the next tribal council.
Every three days, the Ogakor and Kucha tribes competed in “immunity challenges” that forced the losing tribe to eliminate one of its members. As a result, the strongest members of each tribe were highly valued because they could help other tribe members avoid early ejection. On the other hand, the strongest members were also targets for ejection since they would be harder to beat in head-to-head competition when the number of tribe members eventually dwindled. As players schemed to form coalitions, they lobbied their fellow castaways using arguments such as these to eliminate the strong and the weak.
During the third week, the two tribes merged. Under this entirely new social arrangement, immunity challenges pitted players against one another, and the winner could not be ejected at the next tribal council meeting. Now the goal was clear: get rid of the strongest individual player. We might think that this would automatically lead to the demise of the second strongest person (the one who did not win immunity), but another potent force was at play. Over the course of the preceding three weeks of struggling against the elements, each tribe had bonded, and these coalitions endured even after the merger.
Nowhere was the power of social connection more evident than at the first immunity challenge of the merged tribe. Contestants were forced to stand on wooden pillars above a river. The object was simple: the last person standing on a pillar would win immunity. One by one, the contestants gave up, jumped into the river, and swam ashore. Some gave up because they were exhausted. Others gave up when Jeff Probst, the show’s host, offered to give them peanut butter in exchange. After nine uninterrupted hours of standing, there were only three contestants left, and then Alicia Calaway, a personal trainer from New York City, jumped in the water. The two remaining Survivors were Keith Famie, a professional chef from Detroit, Michigan, and Tina Wesson, a nurse from Knoxville, Tennessee. As it turns out, the two of them were both from the Ogakor tribe. But Keith was in greater danger of being voted out of the competition, and so he said to Tina, “I need this one,” and she willingly dropped into the water. This gave him immunity and prevented the former members of the Kucha team from being able to coordinate their votes to cast him out. Tina later said, “It was harder to come in than to stand out there. I knew that for the good of our team, I had to let Keith win.”
The season climaxed on day forty-one, when the remaining three contestants were Tina, Keith, and twenty-six-year-old Texan Colby Donaldson. Colby had just won immunity and would decide which of the other two would be eliminated at the tribal council. Tina was popular with contestants who had been previously ejected from the tribe—the very people who would decide which of the final two contestants would win. Meanwhile, Keith had alienated several players by insisting that his skills as a chef were necessary to their survival. Most viewers thought Colby would eliminate Tina so that he could compete against Keith. But he shocked the nation and probably gave up a million dollars when he chose to eliminate Keith instead.
After the vote that night, the episode concluded with many scenes from previous days that made it clear that Colby and Tina had done well in part because they had formed a very strong alliance that had helped them through many difficult situations. Colby had chosen friendship over what seemed like certain victory. And, indeed, when the final vote was tallied before a live studio audience in Los Angeles, Tina Wesson was declared the winner.
Many people questioned Colby’s decision and claimed that he had miscalculated. But another plausible interpretation is that friendship and loyalty had trumped self-interest. This is exactly the dilemma that most of us face every day: Do we help our friends or help ourselves? And what are the consequences? Will we look dumb if we help others? Will we look mean if we do not? Is it possible to be nice and survive? And how can we possibly make these decisions when we have many friends in a dancing pattern of shifting alliances and interests?
Part of the fascination with the show was not only the characters but also their complicated interactions. Survivor presents a series of interlocking, connected biographies—a sociography, actually, something akin to a novel. And, like a Russian novel, the story follows the shifting connections between people in the group and all the social complexity, as well as the group’s fluid morality. Shows like Survivor are alluring precisely because they mirror the ancient struggles within our minds and among our peers.
Like ants, bees, penguins, wolves, dolphins, and chimpanzees, human beings are social animals, living in close proximity to one another in groups. In fact, the most important feature of the human environment is the presence of other members of our species. Because we lack any real predators, the only major threat to humans is other humans. If we did not need other humans so much, it would make a lot of sense to avoid them.
Our particular relations with other humans are therefore crucial. We deliberately choose to form social connections with specific individuals, with whom we share greater or lesser intimacy and affection, for brief or lengthy periods of time. And unlike other social species, we have a special capacity to imagine what others are thinking and feeling, including what they are thinking and feeling about us. Our embeddedness in social networks means that we must cooperate with others, judge their intentions, and influence or be influenced by them.
In short, humans don’t just live in groups, we live in networks. In previous chapters, we have shown how social networks affect everything from emotions to health to politics. But the question remains: why do humans form these networks in the first place? Through several strands of new research, we are finding a surprising answer: our desire to form connections depends partly on our genes.
Evolution likely plays a role in the way we connect to one another because the very act of connection is itself subject to natural selection. Recall the burning house we discussed in chapter 1. You want to get water from the river as quickly as possible to douse the flames, and a group of people has to get organized to help you. Some network formations will work better than others. We might say that the one that works best is the most “fit” for the task.
Now suppose that we had a tournament where we let different groups (like Kucha and Ogakor) compete to put out fires. In each round, we’d set two fires, then let each team try to put one out as fast as it can. After each game, we would eliminate the team with the slowest response time and invite a new team to play. After many rounds of this, teams that play quite well would emerge, suggesting that they had figured out efficient ways to organize themselves and work together. The teams that ran randomly back and forth to the river would be eliminated, but the teams that coalesced in a bucket brigade would not. The envious and selfish teams would be eliminated, but the helpful and collaborative teams would not. In this way, the teams with the most fit social networks would become the winners. The structure of the social network would adapt to meet the functional requirements it needed to fulfill. We form a line because water needs to flow. And in fact, some species exhibit bucket-brigade behavior because it is an efficient adaptation to their environment; consider ants that pass food items from one worker to another, for example.2
In the modern world, we see tournaments that influence network structures all the time. In American football, the offense has to put seven of its eleven players on the line of scrimmage (just behind the ball) at the beginning of a play. However, the other four players can stand wherever they want. This freedom has led to an enormous variety of formations, including the T, I, ace, pro-set, single-wing, double-wing, shotgun, pistol, eastern shotgun, wishbone, flexbone, wing-T, and A-11. Some formations have specialized purposes, like the self-explanatory goal-line formation. Each has advantages, depending on which players need to interact and what the objective is. For example, the shotgun formation puts the quarterback farther away from the ball so that he has more time to pass to a receiver while avoiding rushers from the defense.
One reason for the enormous variety of offensive formations is that coaches constantly retool their strategies in order to beat commonly used defensive plays. Occasionally such innovation yields a completely new way of playing the game that the defense must then adapt to. Emory Bellard, an offensive line coach for the Texas Longhorns, invented the wishbone formation in the summer of 1968. Because the team had a quarterback, halfback, and fullback who could all run well with the ball, Bellard wanted to give the quarterback three choices (the “triple option”) that could be executed after a play began. The quarterback would first decide whether to give the ball to the fullback who was close behind him. If he kept the ball, he would break in one direction. As the quarterback ran, he could cut inside or flick the ball to a halfback running with him in the same direction. With two halfbacks behind and flanking the fullback, the four men in the backfield made a pattern that looked like a wishbone, which led Mickey Herskowitz of the Houston Chronicle (and everyone else thereafter) to call it the wishbone formation. The extra halfback served as a blocker for the play, and the symmetry of the formation meant that the play could break left or right, so that the defense had to prepare for four different backs who might carry the ball.
Texas tied the first game it played with the formation and lost the next. However, Texas went on to win its next thirty games, including two national championships. Bellard took the successful strategy with him when he moved to Texas A&M University and then to Mississippi State University, and other teams later imitated the formation, including Oklahoma, Army, and Air Force. However, defensive coaches soon developed strategies to beat the wishbone, such as the backbone defense. These new counterstrategies eliminated the original advantage of the wishbone and forced offensive coaches like Bellard to continue searching for the perfect offensive line.
Naturally occurring social networks are not like football teams or reality shows. No coach tells us how to pick our friends, and no one eliminates us if we lose a game or fail to put out a fire. However, human beings are and always have been subject to an analogous set of constraints that determine what kinds of social-network structures work and endure. Among early hominids, individuals who lived in social networks that enabled a group to acquire more food or to fend off attackers were more likely to survive and reproduce. As a result, over a long period of time, the individuals who naturally formed networks or who had specific traits conducive to forming particular kinds of networks would have had a selective advantage and might eventually have made up the largest part of the population. The networks we form today exploit different tools (like cell phones and the Internet) and operate in a different environment, but the urges we have to connect and to organize ourselves into groups of friends evolved at a time when genetic evolution favored some patterns over others.
Natural selection is brutal. Only the “fittest” individuals survive and get to reproduce and pass their genes on to the next generation. This creates a puzzle when it comes to social interactions. Suppose you tend to share food with friends who are not related to you. If food is scarce, then your generosity will help your friends become more fit. And it will make you less fit (less able to survive). Therefore, the genes that contribute to your desire to share food are less likely to be passed on than are the genes that would make you keep all the food for yourself. So, where does sharing come from?
This is the puzzle of cooperation and altruism: people who are willing to help others should be, it might seem, less likely to survive than people who care only about themselves. More formally, cooperators who are willing to pay a personal cost to help a group of people are less likely to survive than free riders who do not pay a personal cost but benefit from the group’s activities. For example, when humans learned to hunt large game hundreds of thousands of years ago, this gave a fitness advantage to the groups that knew how to do it. But if it is risky to take down a mastodon, why not let someone else do it? If you are the most selfish person in your group, then presumably you would be more likely to survive.
In one of the most celebrated openings to a novel in modern fiction, Ian McEwan, in Enduring Love, provides a vivid illustration of the problem of cooperation. A helium balloon is hovering near the ground in a green English field in strong winds. Curled up inside the basket is a frightened boy, and outside, hanging to a rope, is his grandfather, desperately trying to control the balloon before it is blown away. He calls for help, and five men come running. Each of the six men now has hold of a rope attached to the balloon, and no one is heeding the suggestions of any of the others; there is no leader to guide them. A new gust of wind comes, lifting the balloon ten feet off the ground, dragging the men into the air, dangling. If they all hold on, they will return to the ground swiftly and safely. But one lets go, and the balloon, freed of this ballast, jolts higher. Instantly, the others face a difficult decision. Another one lets go, then another, increasing the pressure on the others to follow suit, until just one remains. That man, a really good man, one in whom “the flame of altruism… burned a little stronger,” holds on. He is carried away trailing the balloon like the tail of a kite until, to the horror and humiliation of the other men watching, he has no choice but to let go, three hundred feet above the ground, and fall to his death. As McEwan noted, these six men, hanging a few feet above the ground, “enacted morality’s ancient, irresolvable dilemma: us, or me.”3
The good news is that people very often ignore their selfish tendencies when interacting with people to whom they are connected. In Survivor, Tina could have forced Keith to continue standing on the pillar, but instead she cooperated and jumped in the water when he asked. And across a wide variety of laboratory experiments that study dilemmas in altruism and cooperation, people choose to help other people about half the time, even people they will never interact with again. So it seems that a naive application of evolutionary theory to whether it makes sense to help other people would be incorrect. Selfishness does not always pay. If it did, we would all be selfish.
Now, in the real world, outside the laboratory, there are many other considerations, because we live in a web of interactions with one another and because we have repeated or sustained interactions across time. Tina might have given up because she knew she would be competing with Keith in future challenges and would need his help. Evolutionary theorists call this direct reciprocity. If you have several opportunities to cooperate with the same person, one way to get that person to help you is to promise future cooperation.
In an ingenious and famous study designed to examine reciprocity, political scientist Robert Axelrod showed that a cooperative strategy called “tit for tat” often is more effective than always cooperating or always being selfish.4 In tit for tat, you cooperate the first time you meet someone, and thereafter simply copy what that person did the last time you interacted with him. This behavior is basically an inverse Golden Rule: do unto others as they have done unto you. If someone cooperates, then reciprocate that cooperation the next time around. If someone does not cooperate, punish him next time by withholding cooperation. Simple, but effective.
In a world full of people using the tit-for-tat strategy, cooperation will happen all the time. But in a world of purely selfish people, a person engaging in tit for tat does not fare so well. If you are using the tit-for-tat rule, the first time you meet a selfish person you will cooperate with him but he won’t cooperate with you. You learned your lesson, and you will copy him in any future interaction, but that first meeting means he is a little more likely to do better since he got something from you on the very first interaction. If you don’t meet some other people who are willing to take a chance on cooperation pretty soon, you and your genes are doomed.
It was this situation that led mathematician Chris Hauert and his colleagues to consider another possibility in an important evolutionary model published in Science in 2002.5 In Axelrod’s study and in most previous theoretical models, individuals were forced to interact with each other. But what if they could choose not to interact? Rather than attempting to cooperate and risking being taken advantage of, a person could fend for herself. In other words, she could sever her connections to others in the network. Hauert called the people who adopt this strategy “loners.”
Using some beautiful mathematics, Hauert and his colleagues showed that in a world full of loners it is easy for cooperation to evolve because there are no people to take advantage of the cooperators that appear. The loners fend for themselves, and the cooperators form networks with other cooperators. Soon, the cooperators take over the population because they always do better together than the loners. But once the world is full of cooperators, it is very easy for free riders to evolve and enjoy the fruits of cooperation without contributing (like parasites). As the free riders become the dominant type in the population, there is no one left for them to take advantage of; then, the loners once again take over—because they want nothing to do, as it were, with those bastards. In short, cooperation can emerge because we can do more together than we can apart. But because of the free-rider problem, cooperation is not guaranteed to succeed.
To deal with free riders, another type of person is needed: punishers. People everywhere feel the desire to enforce social norms they see being violated. Some people honk when a car cuts them off in traffic, even though the honk does not change the outcome. Others risk confrontation by asking people smoking in a no-smoking area to stop. And on many occasions, innocent bystanders are willing to testify in court to crimes they have witnessed, even though this potentially exposes them to retribution. These people all pay a small cost themselves to impose a cost on someone who does not cooperate. And this is a different kind of connection. Cooperators connect to others in order to create more; free riders connect in order to leach off those who create; and punishers connect in order to drive away free riders.
Theories of punishment have been around for a while, but they have always gotten stuck on how such a behavior could have evolved in the first place.6 In a world of free riders, a punisher must constantly exert energy to correct each and every transgression (which sounds exhausting). Punishers could easily be overwhelmed. But in a world of disconnected loners, punishers would not have to punish anyone. Using this reasoning, we created our own model and showed that small groups of interconnected, interacting cooperators and punishers could coevolve in a world of people who otherwise keep to themselves, and this pushes the whole population toward higher overall levels of cooperation and connection.7
Hauert and his colleagues later extended our model, showing that under general conditions it would create a mixture of people who cooperate and free-ride and people who connect and disconnect.8 Moreover, they demonstrated that the population was frequently in transition, meaning that we might expect to find different proportions of individuals of different types at any given moment. Unlike the models that predict too much cooperation or too little, the extended model showed that cooperation would happen frequently in a world where it was possible to monitor and punish free riders and where there was variation in the tendency of people to join groups. In short, the model predicted two things: some people will cooperate, and others will not; and some people will be well connected to the social network, and others—the loners—will not.
This kind of variation in human behavior is very hard for traditional economists to explain. The standard way of thinking about human beings in economics is that every person makes a decision without considering the interests of others (except insofar as the interests of others impinge on one’s own). From this perspective, the reason any cooperation arises is that the choices of the individuals involved are incentive compatible: I scratch your back because I think you are going to scratch mine. If we happen to be in a situation where it is possible for you to walk away, then I will refuse to help you. And you and everyone you know would also refuse if you were in my shoes. In other words, there is no inherent variation in how much people are willing to help others. And social connections are meaningless.
Indeed, Homo economicus inhabits a brutal, dog-eat-dog world in which concern for the well-being of others does not exist. The expression Homo economicus, a slightly tongue-in-cheek construction, was first used at least one hundred years ago to describe a vision of our species as one that relies on self-interest to obtain the maximum personal good at the lowest possible cost. But even earlier, in 1836, philosopher John Stuart Mill was already propounding a model of “economic man” who “inevitably does that by which he may obtain the greatest amount of necessaries, conveniences, and luxuries, with the smallest quantity of labour and physical self-denial with which they can be obtained.”9 Implicit in this vision is that people are lazy and greedy but also rational and self-interested and self-directed. Such a model leaves no room for altruism. Further, it leaves utterly unexamined how people come to want what they want to begin with.
We propose an alternative. Homo dictyous (from the Latin homo for “human” and the Greek dicty for “net”), or “network man,” is a vision of human nature that addresses the origins of altruism and punishment, and also of desires and repulsions. This perspective allows our motivations to depart from pure self-interest. Because we are connected to others, and because we have evolved to care about others, we take the well-being of others into account when we make choices about what to do. Moreover, by stressing our embeddedness, this perspective allows us to formally include in our understanding of people’s desires a critical source: the desires of those around them. And, as we have seen, this applies to everything from our health behaviors to our musical tastes to our voting practices. We want what others to whom we are connected want.
Indeed, social networks can involve an endless flow of desires, propagate idiosyncratic disturbances, and give rise to tastes. We have basic desires (such as an interest in sex) that do not depend so much on the specific desires of those around us. But we also have many arbitrary desires—in music or clothes, for example—that are strongly influenced by others. In other words, some of our tastes may be for things that are made more desirable when others desire them. Once such beliefs arise, for whatever reason, they can spread and amplify within the network. Whether these beliefs, desires, or tastes arise as random irregularities or whether they have a more consistent and fundamental origin (for example, political ideology and religiosity have partly genetic bases), they nevertheless get magnified in the network, and they flow through it.
Beginning in the 1970s, nontraditional economists began to test some of the most basic assumptions of their discipline, asking questions about cooperation and the origins of tastes. And many were shocked by what they found. In 1982, a group of economists developed a simple but clever experiment called the “ultimatum game” in which two players bargain over $10 given to them by the experimenter.10 The first player is instructed to make an “offer” to the second player for how to divide the $10. He could offer to give it all away, keep it all, split it fifty-fifty, or divide it any way that it is possible to divide $10. The second player then gets to decide whether to accept or reject the offer. If she accepts, then they divide the money as agreed and both get to keep it. If she rejects the offer, then neither gets anything. Game over.
Now, traditional economists who assume that all actors in their models are self-interested Homo economicus reasoned as follows. The second player would rather get something than nothing. Even a penny is better than nothing. And the first player knows this, so he knows that the second player will accept any offer greater than zero. Economists therefore predicted that the first player would offer the second player a penny, keeping $9.99 for himself, and the second player would accept the offer.
But that is not what happened at all. In the initial experiments with American college students, economists found that subjects frequently rejected low offers. Offers of $2 were rejected about half the time, and lower offers were rejected even more frequently. Moreover, the people in the first player’s shoes seemed to understand that this would happen, since unfair offers were rare. The most common offer was an exact fifty-fifty split, and, on average, the first player earned a little bit more than the second player, but not much more because rejected offers caused both players to lose everything. Given that the first player seemed to know what kind of offers would be accepted and rejected without even discussing this with the second player, his behavior was consistent with that of someone who was purely self-interested and trying to make the most possible money for himself. However, the second player’s behavior was completely inexplicable. Why would she reject an extra one or two dollars from an anonymous person whom she had never met and would never cross paths with again?
The ultimatum-game experiments led to the development of another set of experiments involving a new so-called dictator game to study the effect of the second player’s power to accept or reject an offer. In this game, the first player is given $10 and allowed to divide it between herself and the second player any way she likes. But now the second player can do nothing. The first player’s offer is automatically accepted no matter what. Because the second player has no power, economists expected that the first player would keep everything, and many did. But many more did not. The average first player gave away about $2 to the second player. The results of this extremely simple experiment were difficult to explain if we thought of behavior as being driven purely by self-interest. People were literally taking money out of their own pockets and giving it to anonymous strangers.
The experiment further showed that there was inherent variation from person to person that might have an impact on how people behave. In our own work, we have shown that people who give more in the dictator game are more likely to vote, donate to a campaign, run for office, attend a protest, and help Hurricane Katrina victims.11 They also score higher on psychological tests that assess how “humanitarian” a person is. And just as Hauert’s evolutionary models predict, we find variation in the willingness of individuals to bear personal costs to help others. Some people care only about themselves. But the majority of us take other peoples’ well-being and interests into account.
Situations like these are not entirely fanciful. In fact, real-life ultimatum games occur all the time, sometimes quite colorfully. For example, in 2006, a contractor named Bob Kitts was tearing down the walls of an eighty-three-year-old house near Lake Erie, Ohio, when he found two green metal boxes carefully suspended by wires inside a wall. In the boxes were white envelopes that bore the name “P. Dunne News Agency” and contained $182,000 in Depression-era bills. Saying that he was not raised to act any other way, Kitts notified the owner of the house, Amanda Reece, a high-school classmate who had hired him for the remodeling project. Pictures they took show them happy and grinning, sitting on the floor next to a coffee table covered with carefully laid-out piles of bills.
The trouble began not long afterward when they discussed how to share the money. Reece offered Kitts 10 percent. He wanted 40 percent. Perhaps if Reece had been familiar with the percentages the ultimatum-game experiments had revealed, she would have made a better first offer, and they would have been able to avoid a whole lot of trouble. Because they were unable to agree on a fair split, the information leaked, and the local newspaper, the Cleveland Plain Dealer, published a story about the find in December 2007, whereupon Dunne’s descendants got involved. All twenty-one of them. Naturally, they wanted the money. In the meantime, Reece spent some of it on a trip to Hawaii and claimed that $60,000 was stolen from a shoe box in her closet. After that, not much was left to be shared among Reece, Kitts, and Dunne’s descendants. A lawyer for the Dunnes put the situation starkly: “If these two individuals had sat down and resolved their disputes and divided the money, the heirs would have had no knowledge of it. Because they were not able to sit down and divide it in a rational way, they both lost.”12
The economic experiments we have discussed were carried out almost exclusively on university campuses in the United States. Research such as this typically attracts hungry undergraduates in need of cash, which has led some to remark that we now know more about American college sophomores than any other animal. But when researchers in other countries replicate the ultimatum-game and dictator-game experiments with local college students, they find largely the same results. The main notable exception is among undergraduates studying economics: they have been taught that the self-interested choice is the most rational one, and so they are significantly less likely to cooperate.13
Anthropologist Joseph Henrich wondered if the results would generalize to people in the nonindustrialized world, so he tried the games with the Machiguenga, an indigenous group he was studying in the Peruvian Amazon.14 Oddly, these people behaved much more “rationally” than their counterparts in the developed world. Those acting as the first player in the ultimatum game made lower offers, and those acting as the second player tended to accept them, a result much closer to what traditional economists had previously predicted.
The “Machiguenga outlier” quickly became a curiosity to a variety of researchers, and several decided to join Henrich for a three-day conference on the subject at UCLA in 1997. These researchers designed a study in which they would scatter to all corners of the globe to administer the ultimatum and dictator games to people in a variety of small-scale societies. The list of countries and peoples studied included Bolivia (Tsimané), Chile (Mapuche), Ecuador (Achuar, Quichua), Kenya (Orma), Tanzania (Hadza, Sangu), Indonesia (Lamalera), Mongolia (Torguud, Khazax), Papua New Guinea (Au, Gnau), Paraguay (Aché), Peru (Machiguenga), and Zimbabwe (Shona). The group also conducted experiments in the exotic locales of Ann Arbor, Michigan, and Brentwood, California, as controls. Subjects were offered high stakes in most cases: they bargained over a day’s wages or more to ensure that the decision would be taken seriously.
It turned out that the Machiguenga were not alone. From one society to another, there was a wide range of variation both in the initial offer and the tendency to reject unfair offers. The Ann Arborites of Michigan performed as previously observed among university students, with average offers by the first player of about 44 percent in the ultimatum game. The Hadza of Tanzania and the Quichua of Ecuador offered much less than university students, about 27 percent on average. Meanwhile, the Lamalera of Indonesia and the Aché of Paraguay offered somewhat more (58 percent and 51 percent, respectively).
The anthropologists involved in the study were curious why some groups would offer more than others, so they studied several aspects of each society, including the nature and origin of the local language and the peoples’ relationship to the physical environment. Some groups lived in forests, others on the plains, and others in deserts. Some groups were hunter-gatherers, others were shepherds; still others were small-scale farmers. Some groups lived sedentary lives, staying in one place, while others were nomadic, roaming widely. Their societies varied in complexity too, with some based on families, and others based on groups of families, tribes, or villages.
But the variables that seemed to be most closely related to differences in group behavior were explicitly social. One of these variables was anonymity, a measure of how frequently each group interacted with strangers. For example, the Achuar of Ecuador hardly ever saw strangers, but the Shona of Zimbabwe encountered them all the time. The groups also varied in how frequently they engaged in market transactions. The Hadza foragers in Tanzania were almost completely self-reliant, and they therefore had little interaction with markets; whereas groups like the Orma of Kenya frequently bought and sold livestock and also worked for wages from time to time.
The researchers found that the groups who were in greater contact with strangers were also more likely to engage in what they call prosocial behavior. That means they cooperated with others in the ultimatum game by making higher offers, but they were also willing to reject low offers. In other words, as groups expand their networks beyond the family unit, they appear to behave less and less like Homo economicus and more like Homo dictyous. The economists’ simplification of a person who would offer little or nothing to his counterparts would thus seem to apply only, if ever, to isolated individuals bereft of social interaction, a state that cannot easily be found even in the most remote parts of the globe and that has never really characterized the human condition.
Over the course of human history, we have gradually come together to live in clans and villages and then towns and cities. We have become progressively more connected, even to strangers. And, as we will see in chapter 8, this development has moved from the real world into cyberspace. But we are not all interchangeable cogs in a machine. Human beings clearly exhibit strong differences in both their tendencies to care about others and their abilities to connect. While our life experiences may influence whether we help strangers or make friends, it is also clear that we carry these differences deep within us from a time long ago when our ancestors were first learning to live together in small groups.
Anthropologists think we began hunting large game about half a million years ago. This was quite an innovation because big animals were plentiful at the time, and a single animal could sustain a group for several days. To be successful, however, our human ancestors had to cooperate. And we have already seen that participating in group activities yields a diversity of coexisting strategies. Some people go it alone, while others contribute to the group. Some people free-ride on the efforts of others, while others willingly punish free riders. If this has been the human condition for hundreds of thousands of years, then maybe our network behavior is not simply the product of markets or of a growing population density. It could be that we have evolved genetically to adapt to the risks and opportunities of cooperating in groups. If so, then we should expect to find evidence that cooperative behavior and our tendency to connect are written in our DNA.
To test this theory, we started in an unlikely place. In the summer of 2006, we traveled with our colleague Chris Dawes to a sleepy town called Twinsburg in the middle of rural Ohio. An annual festival is held there, complete with carnival games, hay rides, and funnel cake; in many ways it seems like any other country fair. But this festival is different. It started in 1976 when some of the citizenry thought it would be nice to honor the town’s name by setting aside one day during their bicentennial festivities to celebrate twins. Thirty-seven pairs showed up that first year, and the idea quickly caught on. By 1985, more than one thousand pairs of twins were visiting the Twins Days festival, and they continue attending at this rate today, making it the largest annual gathering of twins in the world.
Each year, things start off with a wiener roast for the twins and their families, followed by events that are open to the public like the “Double-Take Parade,” where the twins march by twos from the center of town to the fairgrounds, and a series of twin contests, including several “most alike” and “least alike” competitions. And tucked away to one side is another spectacle, albeit more subdued. Dozens of researchers from around the world travel to Twinsburg each year to study the health and behavior of twins. In tents, researchers ask twins questions about their childhood, take saliva and blood samples, administer vision and hearing tests, and even do dental exams. Each of the studies pays a little money that the twins can use during their time at the fair. In fact, there is usually a rush right before lunch when hungry volunteers descend on the tents to earn funds for their funnel cake.
Researchers like to attend the festival because twins provide a unique opportunity to study genes. Some pairs of twins are identical: they share exactly the same variants of every single gene in their DNA. Other pairs are fraternal, sharing only half of their genes on average. Differences in genetic similarity turn out to be a powerful natural experiment, allowing us to estimate how much genes influence a given trait. For example, identical twins almost always have the same eye color, but fraternal twins often do not. This suggests that genes play a role in eye color, and in fact geneticists have identified several specific genes that are involved. In the same way, scientists can estimate the role genes play in any other trait by comparing the similarity of identical twins to the similarity of fraternal twins. If there is no difference, then genes play no role. If there is a difference, then the magnitude of the difference gives a clue as to how much genes are involved.
The twin-study method is not without its critics. Some people argue that if twins self-identify as being identical, then they will strive to become more similar, their parents will treat them the same, their friends will treat them the same, and so on. Hence, they would resemble each other for social rather than genetic reasons. If identical twins dress alike and eat the same foods and enjoy the same movies, it might be because their social environment encourages them to be similar. But this claim has been tested in ingenious ways. Occasionally, a pair of twins will mistakenly think they are identical and so will all their friends and family members, but a simple genetic screen shows they are not. If the social environment is really what makes identical twins more similar, then these misidentified identical twins should be just as similar as real identical twins. Yet, when scientists tested a variety of characteristics (intelligence, personality, attitudes, and so on), they found that the misidentified twins are only as similar as fraternal twins. That means that it is the genetic state of being identical and not the self-perception of being identical that drives similarity.15
At Twinsburg, we used the twin-study method to measure how much genes play a role in a simple test of cooperation called the “trust game.” In this experiment, we paired each twin with a person she did not know and assigned them the roles of player 1 and player 2. We gave player 1 $10 and asked her to choose how much money to give to player 2. We also told both players that each dollar sent to player 2 would be tripled. For example, if player 1 gave away all $10, then player 2 would receive $30. Player 2 was then asked to choose how much money to send back to player 1 (but this time it was not tripled). So if player 2 received $30 and wanted to split it fifty-fifty, then he could send back $15 to player 1 and keep $15 for himself. Player 1 would earn an extra $5 as a result.
This game is called the trust game because the first player’s decision indicates how much she trusts the second player to return some of the money she is giving away. The more she gives, the more she trusts the other person. By the same token, the second player’s decision indicates how “trustworthy” he is. The more he returns, the more he is reciprocating the first player’s initial generosity. Higher values of both trust and trustworthiness indicate more cooperative, prosocial behavior.
Over the course of two summers at Twinsburg, we had about eight hundred twins play this game, each of them with a person they did not know. We then compared the identical twins to the fraternal twins. The results indicated that genes significantly influenced both trust and trustworthy behavior. And in one of those happy accidents that occur so often in science, by chance we traded e-mails with David Cesarini, an economist from MIT who was doing exactly the same thing with several hundred sets of twins in Sweden. Amazingly, he had observed nearly identical results in his own study. So we joined forces, and both studies were published side by side.16
Since then, David Cesarini and Chris Dawes have found that genes influence behavior in the dictator game and the ultimatum game that we described earlier. What this means is that cooperation, altruism, punishment, and free-riding are written into our DNA. There can be no doubt that our life experiences have a big impact on all of these characteristics, but for the first time we had found evidence that diversity in these social preferences may be at least partly a result of our genetic evolution.
Human sociability and social networks have ancient genetic roots: apes form social ties, hunt in groups, maintain enduring social bonds, and derive advantages in terms of how long they live and how well they reproduce from these ties. But humans take these traits to a whole new level. The tendency to form social unions beyond reproductive ones is biologically encoded in humans: we seek out friends, not just mates. And through our research, we have discovered that genes also play a role in more complicated aspects of social-network structure. In fact, genes have a big impact not just on whether we are friendly but also on where exactly we land in the vast social network that surrounds us.
To explore the role of genes in social networks, we analyzed 1,110 twins drawn from a national sample of 90,115 adolescents in 142 schools (this is the same Add Health data set used to study adolescent sexual behavior discussed in chapter 3).17 These students had been queried about their friendship networks, and the entire friendship network at each school was known, as well as the student’s precise position in it. We started by studying the most fundamental building block of a human social network: the number of times a person is named as a friend. We found that genetic factors were very important, accounting for about 46 percent of the variation in how popular the kids were. On average, a person with, say, five friends has a different genetic makeup than a person with one friend.
This result by itself was not too surprising. We already knew, for example, that facial symmetry is heritable and is associated with beauty, and that this could help to explain why some people naturally attract more friends. But what was surprising was that even higher-order structural aspects of a person’s network position appeared to be under genetic influence. Your genes affect not just how many friends you have but also whether you are located in the center or at the periphery of the network. On average, people located in central parts of the network have a different genetic makeup than those located at the periphery.
We also studied the effect of genes on how interconnected your friends are. Recall that transitivity denotes the probability that any two of your friends are also friends with each other. People with high transitivity live in densely clustered cliques where everyone knows everyone else. People with low transitivity, in contrast, tend to have friends in several different groups. Such people often act as bridges between completely different groups of people. In our study, we found that transitivity is significantly heritable, with 47 percent of the variation explained by differences in genes. Therefore, on average, a person with five friends who know one another has a different genetic makeup than a person with five friends who do not know one another.
One bizarre implication of this finding is that if we knew which genes were involved in transitivity, we could use that information from one person to predict whether two other people were likely to become friends. If Tom, Dick, and Harry are in a group, this means that Tom’s genes affect whether Dick and Harry are friends!
How can this be? Recall that in chapter 3 we discussed how people commonly meet their partners through others. Introductions are a key feature of human life. But not everyone goes around getting their friends together. In fact, some people go to great lengths to prevent connections. In a memorable episode of the television show Seinfeld, the character George Costanza worried so much about his two sets of friends meeting each other that he claimed it would “kill independent George.” That episode hit the zeitgeist, spawning a “Worlds Collide Theory,” which, according to the Urban Dictionary, “states that a man must keep his personal side (i.e., friends) separate from his relationship side (i.e., girlfriend). Should the two worlds come into contact with each other (by means of his girlfriend becoming friends with his friends), both worlds blow up.”18
Based on our findings about the heritability of social-network attributes, we developed a mathematical model for how social networks form. This “attract and introduce” model is built on two simple assumptions. First, some people are inherently more attractive than others, whether physically or otherwise, so they are nominated more often as friends. Second, some people are inherently more inclined to introduce new friends to existing friends or to attempt marriage matches among their friends (and so these people indirectly enhance their transitivity).
As a result of these two behaviors, we also affect how central we are to the network. Diversity in these choices (whether conscious or not) yields an astonishing variety of locations we can inhabit within our social network, and this in turn has important implications for our lives. If genes influence whether we are in the middle or at the periphery of a social network, they can also affect how rapidly we get a piece of gossip (the center is better) or how likely we are to become infected with an epidemic disease (the center is worse).
The variation in whether certain positions are indeed better or worse, depending on circumstances (acquiring information, avoiding a germ), helps explain why not every position in human social networks is the same. If it were always beneficial to have a certain number of friends and a certain number of connections between our friends, then our social world would look like a boring, predictable lattice or the atoms locked in a salt crystal. We would all have the same kind of network.
Traits that are always adaptive tend to reach what geneticists call fixation in the population: in the long run, everyone becomes the same. But when there are conflicting pressures—under some circumstances, a trait is beneficial, but under others, it is not—then it is possible to maintain diversity in the population in the face of natural selection. If it were to our advantage to all be the same height, then we would be, because long ago evolution would have culled the very tall and the very short. Likewise, if only one kind of social network and location within it were optimal, we would all have the same kind of social networks and be located in identical positions within them.
There are probably many reasons for genetic variation in the ability to attract or in the desire to introduce friends. More friends may mean greater social support in some settings or greater conflict in others. Having denser social connections (greater transitivity) may improve group solidarity, but it might also insulate a group from beneficial influence or information from people outside the group. But most important, social networks may serve the adaptive function of transmitting emotional states, material resources, and information between individuals.
Some traits that we have shown to spread in social networks also appear to be heritable (such as obesity, smoking behavior, happiness, and political behavior). That means that a full understanding of these traits will likely require a better understanding of the factors that mediate the influence of genes on social-network structure as well as how the patterns that we see in our networks today might have evolved.
Genes may influence our tendency to form social ties through their regulation of our emotional states. When core needs for intimacy, love, and social connection are not met, people often experience a feeling of loneliness. Feeling lonely is not the same thing as being alone, however, and there is often a disjunction between a person’s psychological sense of disconnection and their more objective, sociological placement in a social network. Feelings of loneliness can arise from the discrepancy between our desire for social connection and our actual social connections.
Because there is a survival benefit for those who find social relationships helpful, this would promote genetic selection of individuals with such emotional responses. Yet, we have already seen in the evolutionary models we described earlier that it is possible for loners (people who opt out of group activities) to survive. Psychologist John Cacioppo and his colleagues make this mathematical prediction more concrete, arguing that, many thousands of years ago, hunter-gatherers struggling to survive in times of undernourishment may have considered not sharing their food with their family—that is, they may have embraced the loner strategy.19 Individuals who did not feel any loneliness in the absence of family or friends may have been more likely to survive, but their offspring would have been less likely to survive the period of undernourishment given the lack of food.
Conversely, individuals inclined to share food with others may have reduced their own chances of survival but increased those of their offspring, suggesting that no single strategy is always best. As a consequence, a diversity of feelings about being connected and sharing with others were able to evolve, leading to heritable differences in loneliness in adults. And this is exactly what behavioral genetics has revealed. Recent studies of 8,387 adult twins from the Netherlands Twin Register confirm that about half the variation in whether a person feels lonely or not depends on his genes.20
We do not know whether the same genes that affect loneliness also affect a person’s social networks, but the results are suggestive. In chapter 2 we saw that emotions can spread from person to person to person and that people who feel lonely are more likely than others to disconnect from the network. As a consequence, it is possible that genes regulate the structure of our social networks via their effect on our moods. If it pays to be a loner when everyone else in the population is cooperating, free-riding, or punishing, then natural selection might favor those genes that promote the feeling and spread of loneliness. But the advantages of being connected mean that there is an upper limit on the number of people who could benefit from going it alone.
Social connection is a complex phenomenon that likely involves hundreds of genes and is probably influenced by countless gene-environment interactions. But some specific genes appear to have a truly remarkable effect. Scientists have recently shown that a single gene variant distinguishes mating and parenting behavior in small mouselike mammals called voles.21 Male prairie voles are paragons of monogamy, attaching to their first mate for life and taking care of their kids. Male meadow voles, in contrast, are much more promiscuous and less likely to care for their young. This stark difference in mating behavior suggests that evolution does not always yield behavior that we might consider moral in humans—sometimes it favors lust and the deadbeat dad. But more important, it shows what a big difference even a single gene can potentially make in the way animals connect to others. And this has led to similar research in humans, which has shown that people with a certain variant of a related gene give significantly more of their money to anonymous recipients in the dictator game—that is, they are more prosocial.22
Given the role of genes in social networks and cooperative behavior, and the fact that other animals seem to be similarly affected, it might seem that human social networks are nothing special. Humans do share much in common with other social species. For example, primate social networks (in chimpanzees, gorillas, orangutans, and so on) are built on grooming—the activity most of us have seen in nature documentaries where apes appear to be foraging through the hair of their confederates. This close-proximity activity allows one individual to get to know the other, his behavior, health, tendency to violence, willingness to reciprocate, and so on. Primates also use grooming to form alliances, and animals’ willingness to come to mutual aid has been observed to be directly proportional to the amount of time they spend grooming each other.
Primatologist Jessica Flack and her colleagues recently used a “knockout” study to show that removing important individuals from a group of pigtailed macaques significantly changed the structure of interactions in the network of grooming and play.23 This change in network structure, in turn, yielded decreased cooperation and greater instability in the group’s behavior. For an analogous example in humans, consider what happens when you take a teacher out of a middle-school classroom or when a referee ejects one of the eleven players on a soccer team. Both the person and all their ties are lost, and things do not work as smoothly.
The link between cooperation and networks does not appear to be uniquely human. Sociologists Katherine Faust and John Skvoretz tackled the issue of human uniqueness head-on in their own study of forty-two social networks sampled from fifteen different species, including chimpanzees, three kinds of macaques, patas monkeys, vervet monkeys, cows, hyenas, highland ponies, red deer, silvereyes, sparrows, tits, and human beings. Among humans, they looked at networks of managers, monks, and even U.S. senators. Within this enormous variety of network structures, there were some important similarities. They found that the type of relationship in the network was much more important than the identity of the species for making good predictions. For example, grooming relationships appeared to be very similar across species. In fact, the model that best predicted the network structure of U.S. senators was that of social licking among cows.24
However, primates have cognitive abilities that are especially well suited to making sense of social information. They can recognize individuals; distinguish kin from nonkin; assess and compare the value of resources and services offered by others; remember past interactions with particular group members; discriminate between cooperators and free riders; and assess the relative desirability of prospective rivals, mates, and allies. They also have “third-party knowledge” about the relationships between other members of their groups. For example, in one set of experiments with vervet monkeys, the investigators played tape recordings of the screams of baby monkeys, and the adults in the group looked not toward the screams but preferentially toward the female they knew to be the mother of the baby whose voice was being heard, the same way human adults hearing a baby crying on an airplane would. Crucially, primates are also able to manipulate their social network in response to events such as loss of a partner. For example, females in one species whose close kin member died responded by spending more time grooming other social contacts and expanded the number of individuals with whom they interacted. Humans behave similarly: a recently bereaved woman may join a variety of new groups in order to make new friends or to find a new spouse, and her friends may band together to help speed her recovery from grief.
Given the similar (if simpler) behavior in primates, it should not be surprising to find that many human social-network behaviors are hardwired. Humans, of course, manipulate and interact with social networks in ways more complex than other animals, and in the process our social networks can change. Paradoxically, as we will see in chapter 9, while networks depend on their members, they also are inherently stable, and new members can come and go as part of the normal process by which networks evolve and survive.
So far, we have drawn on evidence from remote human societies, our primate cousins, other social animals, and even our genes to support the idea that social networks are an ancient part of our genetic heritage. But unlike other animals, human beings cooperate with unrelated individuals in enormous and complex societies. And to navigate this complexity requires some special abilities that only humans have. In particular, our brain seems to have been built for social networks.
Compared to other species, humans have unusually large brains and unique cognitive capacities, ranging from language to abstract mathematics. Evolutionary biologists and physical anthropologists have developed a variety of explanations for the origin and function of the human brain. The general-intelligence hypothesis posits that larger brains have allowed humans to perform all kinds of cognitive operations (ranging from superior memory to faster learning) better than other species. The adapted-intelligence hypothesis posits that particular mental faculties evolved in response to particular environmental challenges. For example, caching birds that store food in a variety of locations have terrific memories, and social insects have complex communication skills.
An alternative theory about our big brains that has recently gained momentum is the social-intelligence hypothesis, which stresses the special challenges posed by living in close proximity with others and confronting a complex social environment that demands constant cooperation or competition. In other words, this is essentially a theory about networks. It suggests that humans are “ultrasocial,” with skills ranging from language to abstract reasoning to empathy and insight that are adapted to a highly social environment. It also suggests that skills have evolved in humans to create and form social groups, to manipulate the social world, and to shape the architecture of the social networks in which we are embedded. These groups can be discerned both at a large scale (where they constitute whole cultures, with their own languages and artifacts) and at a small scale (involving, say, just kin-group interactions). Once humans form particular social groups with particular social-network ties, they can then transmit their knowledge to others near and far.
At some point in evolutionary history, primates applied the skills initially developed for finding a mate and maintaining a stable reproductive union to relationships that were not reproductive. As primatologists Robin Dunbar and Susanne Shultz argue, the everyday relationships of many primates entail a type of attachment that is found only among pairs of individuals bonded for reproduction in other species. Primates of the same or opposite sex could form long-term, stable relationships to their mutual benefit. But each of these relationships brings with it the potential for many more relationships, since every friend is also potentially connected to every other friend. There is only one relationship between two people, but there are three possible relationships between three people, six between four people, ten between five people, and so on. Since the number of possible relationships grows exponentially with group size, it probably takes a big shift in cognitive capacity to keep up with all the drama of a full social life.
More direct evidence of the social-brain hypothesis comes from the use of functional MRI to study the neural correlates of social decision making. Neuroscientists have found that we use a very large part of the brain called the default-state network to monitor social interactions, and we even seem to have expanded the use of these brain regions to think about coalitions and conflicts in modern-day politics.25 Biologists have also found that color vision, which occupies about two-thirds of a human brain’s capacity, is optimally tuned to detect differences in skin color. This might be so that individuals can discern emotional states in other members of the same species. And, intriguingly, the species that have this ability also have little facial hair (like humans, sometimes called the “naked ape”), suggesting that color vision coevolved with the need to be able to see the faces of other group members and assess their moods.26 Thus, over millions of years, our social life has affected not merely our ability to monitor others and make decisions; it also may have changed the very way we see the world.
Anthropologist Michael Tomasello and his colleagues have taken the social-intelligence hypothesis a step further and argued for a variant called the cultural-intelligence hypothesis, positing that higher cognitive functions are based on a whole complex of social skills. He writes: “There should be an age… before children have been seriously influenced by written language, symbolic mathematics, and formal education at which humans’ skills of physical cognition (concerning things such as space, quantities, and causality) are very similar to those of our nearest primate relatives but at which their skills of social-cultural cognition (specifically those most directly involved in cultural creation and learning, such as social learning, communication, and theory of mind) are distinctively human.”27
The cultural-intelligence hypothesis finds support in experiments on chimpanzees, orangutans, and two-and-a-half-year-old human children using an omnibus Primate Cognition Test Battery—an IQ test of sorts—that offered treats for a variety of tasks, ranging from locating a reward, tracking a reward after invisible displacement, discriminating quantity, understanding that things can change in appearance, understanding functional and nonfunctional tool properties, and so on. The IQ tests administered in this head-to-head comparison of human toddlers and adult primates included two in which the toddlers had a particularly strong advantage: following the direction of an actor’s gaze to a target, and reading the intentions of others. These tasks showed that even at this young age, when physical cognition in children is similar to that of apes, humans outperform them on tasks in the specifically social domain.
In short, the human brain seems to be built for social networks. Over time, evolutionary selection has favored larger brains and greater cognitive capacity to respond to the demands of a more complicated social environment. Individuals living in social networks confront a set of cognitive challenges not faced by solitary individuals or by those in disconnected groups. These challenges arise from the need to understand others and to cooperate with them, as well as to occasionally act altruistically for the benefit of the group. A bigger brain is needed to avoid self-destructive aggression, to hunt down a mastodon, and to avoid getting voted off the island.
There is increasing evidence that religion and the inclination to form social networks are both part of our biological heritage and that the two may be related. Religion is one means of integrating people into a collective. A belief in God can have relevance to social networks in a direct way: God can actually be seen as a part of the social network. This involves not just the personification of a deity but the addition of a deity into the social fabric.
One way to make social networks stable is to arrange them so that everyone is connected to a node that can never be removed. There would then be a short path from each person to every other person through this particular node. But even the most popular person in a society could not fill this role since, realistically, a single individual cannot be connected to absolutely everyone. And even if someone could be so connected, the effect on the network would not be permanent because humans are mortal.
But such considerations do not apply to God. If God were seen as a node on a network, large groups of people could be bound together not just by a common idea but also by a specific social relationship to every other believer. People could perceive a specific social tie to others, and everyone would be one degree removed from everyone else. People who felt a connection to God would have a way of feeling connected to others, because through God everyone is a “friend of a friend.”
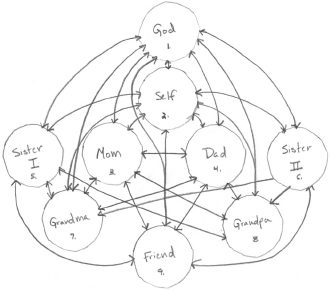
This is not merely an abstract idea. People often see their networks in just this way. For example, in the early 1980s, psychologist Catalin Mamali became interested in how people perceive their relationships to others and how they form mental maps of their relationships. She developed a method to ascertain these mental maps by asking people to identify others with whom they interact, and to draw their relationships in a kind of network graph.28 Her subjects were told to think of people they were “close to” and who were “highly significant” to them and to draw their interconnections. Specific examples of such people were provided: parents, siblings, spouses or intimate partners, children, best friends, friends, and neighbors. Very unexpectedly, however, quite a few people following these instructions chose to include God as a node on their networks and even explicitly made God connected to everyone. The figure below is one such “social autograph” from an eighteen-year-old college student.
The idea that God can be personified and seen as part of a human social network is further supported by the fact that people tend to have an increase in religious belief after the death of a loved one: it is as if a connection to God is strengthened when one loses a connection to people. People might also turn to God in order to support a belief in an afterlife; this belief can sustain the hope of being reconnected with others who have departed. And the very fact that most gods are personified is consistent with the idea of God’s inclusion in social networks and the sense that many people have, and that religions foster, that “God is among us.”
There is a tendency for socially isolated people—and not just bereaved individuals—to anthropomorphize the world around them, from mermaids in the ocean to faces on rocks. Studies by psychologist John Cacioppo and his colleagues show that people who are experimentally induced to feel lonely will alter their mental state in order to personify, and feel connected to, “gadgets, greyhounds, and gods.” Religion is the opiate of the people, Karl Marx famously noted, but it turns out that it is the opiate of disconnected people in particular. Cacioppo and his colleagues administered personality tests to ninety-nine people (fifty of whom believed in God and forty-nine of whom did not) and then randomly assigned them to receive one of two possible interpretations of their performance on the test, regardless of how they actually performed. One interpretation was: “You’re the type who has rewarding relationships throughout life” (that is, you will be connected); and the other was: “You’re the type who will end up alone later in life” (that is, you will be disconnected).
“Social autograph” of relationships identified by an eighteen-year-old college student (subject 1987SA) showing how she feels she is connected to important people in her life. (Courtesy of C. Mamali.)
The subjects then rated the extent to which they believed in ghosts, angels, spirits, God, and so on. Not surprisingly, subjects who reported believing in God before the start of the study reported a strong belief in these supernatural agents. However, regardless of their belief in God, those who were told they would end up disconnected reported an increased belief in supernatural agents.
Lest we think these findings are specific to God, these investigators also did an experiment in which subjects were encouraged to believe that they were more connected or more disconnected, and then they were asked their feelings about pets. The investigators reasoned that if disconnection made people seek out and personify agents in their environment in order to regain a sense of connection, this sensibility should also apply to pets. In fact, this is what they found.29
Making people feel disconnected did not, of course, turn atheists into deeply religious people, but it did nudge people in the direction of believing in God. To the extent that the saying “there are no atheists in foxholes” is true, we can add a wrinkle: this is especially likely to be true if you are all alone in the foxhole.
Our claim is not that God is really a part of social networks, of course, but, rather, that one way to understand religion is to study its role in the function of social networks. Religious sensibilities are partially hardwired in our brains, and they are related to our desire for social connection to others, not only a spiritual connection to God. A key function of religion, in other words, is to stabilize social connections.
Investigations of the fundamental ways in which the mind works further support this idea. For example, functional MRI studies show that during religious feelings and altered states of consciousness, the parts of the brain that regulate the sense of self in time and space actually stop functioning. This contributes to the sensation that “all are one” and may help us overcome a built-in rigidity in the way we perceive our position with respect to others.30 In essence, the brain is fooled into believing that social boundaries do not exist or, equivalently, that everyone is connected to everyone else. Thus, people who would be willing to do something to help their friends suddenly have many more friends and may be more motivated to act on their behalf. In this way, a religious movement can bring together disparate groups of people to achieve a common goal, from caring for the poor to building great structures to, alas, launching wars on rival groups.
As we will see in chapter 8, people sometimes claim to have hundreds of friends in their online social networks. While the human brain is designed to cope with large social networks, our capacity for friends is not, in fact, unlimited. As a key exponent of the social-brain hypothesis, Robin Dunbar has argued that the evolution of larger social groups among primates required and benefited from the evolution of a larger neocortex (the outer, thinking part of our brain), and that managing social complexity required and benefited from the evolution of language. In a celebrated 1993 paper, which was published along with the comments of more than thirty other scientists, Dunbar examined the relationship between brain size and group size in a variety of primates and by extrapolation posited that the expected size of social groups in humans, based on our big brains, should be about 150. This has come to be known as Dunbar’s number.31 Animals cannot maintain the cohesion and integrity of social groups larger than a size governed by the information-processing capacity of their brains. And humans have both the largest groups and the largest brains.
But what do we mean by groups? We clearly do not see clusters of humans grooming each other on the street. In primates, it is easy to define the boundaries of a group—who is in and who is out. But in humans, there are groups as large as tribes and nation-states and as small as families and villages. Dunbar argues that a group is the maximum number of humans in which every member knows every other member, knows whether they are friendly or hostile, and knows the relationships among them. This is the number of people whom you recognize and with whom you can maintain a stable, coherent relationship, which Dunbar defines as “one that can be picked up again on meeting after an absence without any need to reestablish where you stand.” According to Dunbar, an informal definition might be “the number of people you would not feel uncomfortable about joining, uninvited, at a chance meeting in a bar.”
But how do we decide what the natural condition for humans is in order to test the prediction? Here, Dunbar creatively employed data from a variety of sources. For example, a survey of the ethnographic literature looking at all documented censuses of hunter-gatherers showed three types of social groupings: the “overnight camp,” the “band or village,” and the tribe. The mean sizes for these were 38, 148, and 1,155 people, respectively. Hence, remarkably, the size of a band or village matched Dunbar’s number. He also noted that the size of Neolithic villages in Mesopotamia in the period 6,500 BCE to 5,500 BCE fell in the range of 150 to 200 inhabitants. A more subtle point from these studies is that while tribe and overnight-camp size varied widely, the size of a band or village was much more consistent, suggesting that it is a more fundamental grouping.
Dunbar also studied the case of the Schmiedeleut Hutterites, a fundamentalist group who live and farm communally in South Dakota, North Dakota, Minnesota, and in Manitoba, Canada. The Hutterites explicitly regard a community of 150 to be the limiting size, and they make arrangements to split into two groups as they approach that number. They note that this is the maximum group size that can be maintained through the workings of peer pressure alone and that for larger groups a police force and hierarchical structure would be required.
For centuries, the size of fighting units in armies has been about 150 men. The basic unit in the Roman army (the maniple) was 120 men, and the mean size of the company, the analogous unit in modern armies, is about 180. These numbers suggest an upper limit on the size of a group in which members can work together as a coordinated team and know the strengths, weaknesses, and reliability of their brethren. One can even imagine that warfare presents a particular kind of evolutionary selection pressure, and that armies across the centuries have arrived at this working size by empirical observations of what size group is most likely to survive.
Interestingly, group size in modern armies hasn’t changed, even though modern telecommunications would seem to facilitate larger group coordination. This suggests that communication is not the crucial factor. More important is the human mind’s ability to track social relationships, to form mental rosters that identify who is who, and to form mental network maps that track who is connected to whom and how strong or weak, cooperative or aggressive, those relationships are.
Dunbar’s assessments of the relationship between brain size and group size required some further novel claims. He predicted how much time primates would spend grooming each other in order to maintain the cohesion of the larger groups made possible by bigger brains. Dunbar estimated that, for the 150-person group size predicted for modern humans, we would have to spend 42 percent of our time grooming each other! He argued that language emerged among humans, in part, to replace grooming. Basically, language is a less yucky and more efficient way to get to know our peers since we can talk to several friends at once but only groom them one at a time. In fact, in a conversation with a small group, we can assess the behavior, health, aggressiveness, and altruism of several individuals simultaneously. Plus, we can talk to someone else while engaged in another activity, like foraging for food in the refrigerator.
This is actually quite a radical idea. Until relatively recently, it had been thought that language had evolved in order to facilitate an exchange of information—about the location of predators or prey, for instance—or as a secondary consequence related to our development of tools. But the social perspective on language suggests that it evolved as a means of maintaining group cohesion. Just like emotions, language is key to the acquisition and manipulation of social information about other members of our species. A strong indicator of this fact is that most conversation is light on intellectual content, and it certainly is not focused on complex ideas about the environment we inhabit or even about culture or economics. After all, how often—unless we are ten-year-old boys—do we talk about predators or navigation, let alone particle physics or Homer?
Dunbar estimates that language would have to be 2.8 times more efficient than grooming in order to sustain the group size seen in humans. Hence, he estimates that human conversational groups should be about four people: one speaker and 2.8 listeners. But how can we quickly get a sense of the size of groups people form for conversation? One strategy was to assemble data regarding restaurant reservations. Over a ninety-eight-day period at Novak’s Restaurant in Brookline, Massachusetts, in 1968, for example, 3,070 groups made reservations. While there were peaks at two and four people, the average party size was 3.8. Everyday experience with dinner parties supports this result, of course. At large dinner parties, conversational cliques tend to form among four people. Another adventurous scholar—apparently without ulterior motive—observed the size of swimsuit-clad groups on a public beach and found a similar number.
Finally, Dunbar notes that the emergence of language had a further, unintended consequence. After language evolved to handle social interactions on a small scale, we were free to use it to do things like write poetry—just as feathers evolved to keep prehistoric reptiles warm but they wound up using the innovation to fly. In addition, and more crucially, we could use language to handle social interactions on an enormous scale, allowing us to interact in groups such as tribes and nations.
Language permits this transition to large-scale interactions in at least two ways. First, it makes it easy for us to categorize people and interact with them as types rather than as individuals; for example, instead of knowing every policeman (and grooming them), we interact with them in a stylized way (“What seems to be the problem, Officer?”). Second, it allows us to teach others how they should behave with certain types of individuals (“Stand at attention when the teacher comes into the class, students.”). Hence, a person interacting with another person for the first time knows how to do so, without having the benefit of prior knowledge of that particular individual.
The tendency to form ties and to live out our lives in social networks has had an important effect on our development as a species. Social networks encouraged rapid growth in the size of our brains, which allowed us to acquire language and to become the dominant species on the planet. At the same time, these biological changes have given us the capacity to cooperate in large groups, even with complete strangers, to create magnificent and complex large-scale societies. Our connections—partly shaped by our genes but also profoundly influenced by our culture and our environment—are made and remade every day. We choose our friends, develop cultural norms about social order, make and obey rules about whom we can date or marry, enforce ideas about reciprocity, and react to events affecting people around us, in part because we are equipped with empathy. And as we will see in the next chapter, our ability to manipulate and participate in networks is highly relevant to the new opportunities and challenges that we are facing in our hyperconnected world. As we take our real-world networks online, we carry with us the tools wrought by natural selection to make a new world that nature has never before seen.