Before Europe, before the euro, before the three Reichs—the third and shortest by the Nazis, the second of the German Empire, and the first the 844-year rule of the Holy Roman Empire; before the rats carried plague pathogen hitchhikers who ravaged people wherever they met; before that crookback villain Richard III; before Magna Carta; before the last time invaders conquered Britain in 1066 with an arrow in the king’s eye; before the first Holy Roman Emperor, the great European conciliator Charle-magne; before Vikings and northern Scots set foot on the volcanic wilderness of Iceland, the very first to do so; before the Council of Nicaea and the foundations of modern Christianity; before the fall and rise of the Roman Empire, and the great expansive Alexander and his masterful science tutor Aristotle; before the city states of the Greeks; before the Minoans and the Mycenaeans, and to the north the Saxons, Picts, and Goths; before all of these peoples, tribes, customs, cultures, wars, invasions, technology, writing, before all of history, Homo sapiens were already here. Europe has been our land, from the eastern Steppes, to the Atlantic tip of Trafalgar, for millennia before the whole of history. Of all the versions of people that emerged in the world, Homo sapiens have been the sole human occupants of the continent and the islands of what we now call Europe for more than 30,000 years.
Earlier people had been loitering around the continent for up to 2 million years. These included Homo erectus, a species of humans that had great success leaving the motherland of Africa and making it all around the world, as far east as Java, and all over western Europe. One 1.4-million-year-old erectus tooth has been found in a cave in Bulgaria, other remains in Georgia, in France, and dotted all around the mainland of Europe. Evidence of humans in Britain dates back almost a million years, for most of this period connected to what is now the Netherlands by dry land, and they left tools and useful bits of mammoth in the sand and clay of what is now the beaches of north Norfolk. These days, the rapidly eroding coast is exposing these remnants to us. But for all these people and the bits and bobs they left behind around Europe, their bones are too old, maybe too wet, to have preserved their inbuilt genetic historical textbook.
The Neanderthals made their homes all around Europe (see Chapter 1)—in Germany (where the name we gave them comes from), all over France, and scattered in the east of Europe, Wales, Israel, and further toward the east. They were the first true born and bred Europeans: Neanderthals diverged from our best-guess common ancestor, Homo heidelbergensis 500,000 to 600,000 years ago, possibly somewhere in middle Europe, and those Germans didn’t last later than 200,000 years ago. We now know that anatomically modern humans—that is, us—didn’t arrive in their lands until some 60,000 years ago, having slowly moved north and west from the Middle East and before that Africa. We know that we overlapped in time and space with the previous Europeans—the Neanderthals—and we carry their ghosts in our DNA. Coexistence in Europe probably lasted around 5,000 years, which is in evolutionary measure a click of the fingers, but is a time long enough for maybe 200 generations, epic migration, cultural development, plenty of sex and death, and the general business of living. Even though they endure in us, we outlived those people, maybe hunted them, some have suggested even butchered and ate them, and no Neanderthal bones or tools have been found that are more modern than around 30,000 years old.
So, for the time being, the Europe of the last few tens of thousands of years has been our land. In the last decade, and very significantly in 2015, a glut of new research has transformed our understanding of the origins of the Europeans mostly using DNA from ancients and the living, but also encompassing archaeology and language. Between the end of the Neanderthals and the beginning of history, the settlers in Europe changed physically and culturally, and over the next few pages, we’ll see how genes and culture, primarily farming, shaped modern Europe. Just as with our very ancient ancestors in Chapter 1, we’re reconstructing the past with old bones and DNA combined, but the bodies are few and far between. “Far between” is less of an issue, as distance between bodies can give us a good idea of distribution over a continent. “Few” is more of a problem, because genetics is a comparative business, and the more data we have, the better resolution the picture is. But that’s the reality of people who’ve been corpses for thousands of years.
The oldest genome of a European comes from a 37,000-year-old square-jawed man who washed up on the banks of the mighty River Don in southern Russia. He’s called Kostenki today, and his DNA showed similarities with more recent European hunter-gatherers, as far afield as in Spain 30,000 years later, but few with East Asians. We know that key physical characteristics synonymous with eastern Asians arose some 30,000 years ago. Thick straight hair, a density of sweat glands, and a particular tooth shape largely specific to the people of the east appeared in China at that time, though these visible changes all stem from one single gene, and the overall genetic similarities and differences between peoples are measured at a far greater depth than skin deep.
The record for the oldest complete modern human genome comes earlier and from the banks of another mighty Russian river further east. The Irtysh is the chief tributary of the Ob, which together cut thousands of miles across the middle of Russia from China to the Arctic Kara Sea. A Russian artist named Nikolai Persitov carves jewelry and sculptures out of mammoth bones, and while on the hunt for these raw materials in 2008, stumbled over a femur—a thighbone—jutting out of the eroding banks of the Irtysh in a southern district. It was of a 45,000-year dead man, now known as Ust’-Ishim after the place of his resurrection. He gave up his genome to Svante Pääbo and David Reich, and their teams in Leipzig and Harvard, who together assembled his complete DNA as we would do with a living person. Ust’-Ishim’s DNA shares similarities with both eastern Asians and western Europeans, unlike Kostenki, who had little genetic resemblance to the east. Ust’-Ishim is therefore likely to be ancestral to both Europeans and Asians, and so represents—at least genetically—the people who had migrated up from Africa before one group, over thousands of years, turned toward the dawn, while the other one went into the sunset.
The majority of the latest round of genetic analysis of Europe has focused on the last 10,000 years, a time from which the evidence of farming around the world is strong. This includes domestication of various animals, swine, goats, sheep, llamas (dogs had come earlier, maybe 15,000 years ago, as companions for protection and hunting, fed with the scraps of the spoils), and the cultivation of plants, and processing them with Neolithic tools. Why we began farming is disputed. The climate was more stable during this time in Europe, possibly wetter, and some of the big animals that flourished during the cold—woolly rhinos and mammoths, and other megafauna—were all set for extinction. Domestication might have been easier and needed as Europe became more stable and temperate. There may have been health benefits for a regular food chain, but this has been disputed as some archaeological remains suggest that people got shorter after domestication, which can be associated with a less nutritious diet. Please ignore this sentence, as it will only be of relevance on page 77. The “Paleo Diet” is a popular fad that eschews processed foods and carbohydrates in favor of the only foods imagined to be available to the hunter-gatherers of the Paleolithic: no dairy or processed grains, no lentils, beans, peas, or other human-designed veg. Nuts are OK, but no peanuts, as they’re a farmed product. It is almost certainly built on bunkum foundations, as indeed most fad diets are.
By the time of the agricultural revolution, we see multiplication and expansion of genes that encode salivary amylase, an enzyme in your spittle that initiates the digestion of complex molecules. Some people have eighteen copies of it, but chimpanzees only have two. Amylase digests starchy, carbohydrate-rich foods, and helps generate glucose from them, which would provide much needed energy for the evolving and highly energetic brain. It works many times better on carbohydrates that have been cooked. We’re not sure when we began cooking; the range of evidence is wide, but food cooked on fires was definitely part of our menu by 300,000 years ago, so before we were anatomically modern. The expansions of these amylase genes, the presence of cooked food, and the massive expansion of our brains that is observed over our Paleolithic evolution point to a positive selection for eating nutrient-rich tubers. This is hypothetical at the moment; it fits the picture quite neatly, and fits into our overall picture of how culture affects genes and vice versa. Diet is not easy to validate in the deep past, but the clues from genetics suggest that the basis for the Paleo Diet is nuts, even if you’re not allowed to eat peanuts.
As with all fad diets it probably works a bit, but not because of the content of the diet itself, but because the act of dieting prompts people to eat less and think more about their food, and not to shovel huge portions of pasta or chips on their supper plates. So go ahead and diet, but don’t pretend that it’s based on some evolutionary precedent. And remember that whatever we did in the deep past, we live longer and better now than at any point in the history of humankind.
Regardless, the agricultural revolution that occurred at the beginning of the current epoch, the Holocene, coincided with the first evidence for farming, even though the reasons for this revolution remain unclear. But it did irreversibly change everything. This transition to a domestic life fundamentally changed us in our bones and our genes, as we’ll see soon. The land changed more obviously, as you would expect, when it became worked in this most unnatural way. I like to travel on trains in Britain, and marvel at the green rolling hills. I pine for them when I’m abroad. I like to ponder how entirely unnatural they are, how they’ve been designed and built over thousands of years, how the hedgerows—so critical to the biodiversity of the land—were put there by people to separate crops and animals and predators and property. Even the coarse wild brush in the highlands of Scotland and much of northern England has been grazed and unnaturally tended and grazed again continually for millennia.
Estimates are that the hunter-gatherers who were all but wiped out by the agricultural revolution numbered around 2 million 12,000 years ago. Agriculture spread like a virus over the continents from its birth somewhere in the Middle East (and dotted in other spots in Africa and China), and would be the dominant business of humans for most of the rest of history. Farming today is industrial, and dominated by monolithic corporations who control almost all the food we eat. But this book is not about that. In its inception, farms were subsistence smallholdings, territories carved out of cleared forests and walled fields, and permanent residences. Crops, by their nature, are seasonal, and with farming came a need to plan, to store foods in pots and jars and silos for lean years. With these plans came surplus, some years, and that draw would bring others into the bountiful communities, which would grow and flourish. We humans are a technological species, science is in our souls, and farming is a technology quite the opposite of natural. From humble beginnings, innovation in tilling the land radically brings greater harvest efficiency. With that comes economic disparity: Some have surplus, others have less, which means some families get bigger, and have more children who live, more culture, more technology; and the cycle continues. By the time of the Roman Empire, there were a quarter of a billion farmers on Earth, and foraging was a living now limited to a couple of million people at most, flung far into the corners a long way from Europe, in Australia, South America, and in pockets in Africa.
Argument about the impact, locations, and breadth of the agricultural revolution has preoccupied anthropologists for decades, and is set to continue. We can now add DNA to our arsenal of evidence, alongside—not in competition with—the archaeology of ancient farming, the lie of the land, the broken pots and pans of kitchens and larders, and of the lovely bones, both ours and of the beasts we kept. Arguing about agriculture is a European preoccupation. For forty years we’ve been bickering about farming in the European Union, and the Common Agricultural Policy.
This tale is also all about farming policies, and it starts not far from Brussels. Eight thousand years ago there were probably about 5 million people on Earth, the current population of Norway. They were spread far and wide. By this time, people had reached the tip of South America, Australia, and most places in between (though not into the South Pacific islands or New Zealand), and we have hundreds of corpses and tons of archaeology from these times. We’ve only just begun harvesting their genomes, yet already the picture of the modern evolution of us is being rewritten. Much of the work has been done in large collaborative groups of geneticists, archaeologists, and historians, and the powerhouse of the genetics work is headquartered in David Reich’s lab in Harvard. Reich’s international team plucked the DNA from nine people in 2015. One was a man from a murky cave dwelling in a village outside Loschbour in Luxembourg. One was a woman from Stuttgart. The rest were from a cave in the small Swedish town of Motala. They’ve all been dead for more than seventy-five centuries, but, though they were almost certainly unaware of it, they were standing on the edge of a revolution that had already begun, and would shape Europe and the world for evermore.
Loschbour was a hunter. In his final resting place, he was surrounded by the paraphernalia of a life stalking and killing animals, flint blades knapped to mount on spears and be used to butcher the meat and tailor the hides. In the flatlands of northern Europe 8,000 years ago, these beasts would be wild boar and deer. He was one of the last, though, as immigrants were coming from the east, with new customs, new knowledge, and a penchant for herding rather than hunting.
Loschbour was pulled from his grave in 1935, but DNA was extracted from an upper molar in 2014. Stuttgart emerged from the tomb in 1982, and had her DNA taken from a molar too. She was a farmer from around 5,000 years ago, and was found with the paraphernalia typical of the Linear Pottery Culture—decorated pots and gourds, stone tools, and evidence of animal husbandry. The Motala clan were disturbed from their mausoleum only in the last decade, and samples were taken from the teeth of nine skulls and the tibia and femur—the shin and the thigh bone—of an individual. Like Loschbour, they were hunters too.
All three were genetically different. Stuttgart and Loschbour had genes for dark hair, Loschbour and Motala had genes for blue eyes. Stuttgart had sixteen copies of the amylase gene indicating a carb-rich diet; Loschbour and Motala had fewer. These indicators paint a crude picture of what they looked like, and a more detailed image of some of their cultural practices. But by interrogating the subtleties of their DNA, David Reich and his colleagues were able to unearth the foundations for 10,000 years of European occupation. Our understanding of the genetic basis of Europeans had come from studying the genes of modern, living Europeans. This has plenty of merit, but we now know that the DNA of a population today does not necessarily reflect the DNA of the population in the same spot from thousands of years ago. By comparing the genomes of Loschbour, Motala, and Stuttgart with 2,345 modern Europeans, Reich could pick out the various contributions that these early Europeans have made to the overall structure of Europeans today. What they showed was that we Europeans are drawn from three different groups of people. They’re not the ones represented by Loschbour, Motala, and Stuttgart, but the differences between them highlight different proportions of DNA that we see today.
The first European Homo sapiens were the hunter-gatherers who had moved up from Africa via central Eurasia 40,000 years before, and overlapped and mated with the indigenous Neanderthals. Between 9,000 and 7,000 years ago, we see the genetic fingers of eastern farmers reaching into this population. They didn’t usurp them or wipe them out, though. We see the two populations living not exactly side by side, but at the same time, some hunters, some farmers, and we see the slow integration of genes from the hunters enter into the genomes of the farmers.
And then around 5,000 years ago, another major wave of easterners arrive. The Yamnaya came from the Russian Steppes, driving sheep, riding wagons, making bronze jewelry, and covering their dead in ochre as part of ritual burials. They came and rapidly their way of life spread into middle Europe, bringing their culture and genes, and their fair skin. Farming came to dominate and eventually entirely replace hunting and gathering. Pale skin came to replace dark skin, and we will see more of that later in this chapter.
If you are pale skinned, you are almost certainly a product of these three waves of European immigration. We are in a process of reevaluating human history through these new models of gene flow. There was a time, not that long ago, when we assumed that the history of humankind was simply a series of what evolutionary biologists call “founder events”—small tribes moving further from the motherland of Africa, setting up camp, growing, and then budding off to form new small tribes who would repeat the process in new uncharted territory. Early genetic studies (meaning “in the last decade,” such is the ridiculous pace of this science) indicated that this model may well have been accurate, as small samples of chunks of repetitive DNA seemed to cluster together. You would also expect to see a decline in genetic variation the further we moved from Africa, which is broadly what we see. The latest analyses incorporate the fact that current residents of a geographical area are not necessarily very good representatives of the residents of the deep past. This is obvious if we look at areas that have been subject to migration by modern Europeans: The majority of the peoples of Australia or North America today are from Europe in the last 500 years, and so their genomes are not representative of the indigenous people who were there first. But this impermanence is also not necessarily true for much older populations of humans. The assumption, for example, that the Siberian farmers are going to be most similar to the first settlers of the American continent because of their proximity to the Bering land bridge 15,000 years ago (now the Bering Straits) is not correct. In fact, digging up old bones and wheedling out their DNA shows that today’s Siberians are more like East Asians, but the ancient Siberians were more like Native Americans, mixed in with some northern Eurasian. All this means is that we made assumptions about patterns of migration that were much more linear and spread like ripples, rather than the picture that has emerged in the last couple of years, which says that we moved in all directions all the time, and laid our hats and flowed our genes in a matted crisscross, instead of a nice clean radiation. Oceans and mountains are good barriers to gene flow, but on big open continents the horizon is the limit. Genes flowed out of Asia and into the Americas.
These ancient genomes don’t just tell us about movement and migration. DNA is one of a suite of tools we use now for reconstructing the past, and we build up these pictures with the agglomerated knowledge from archaeology and geology. DNA also reveals behavior. Culture can become embedded in our cells just as it gets buried in the floors of caves, bogs, and dwellings.
It is a strange thing that that some adult humans drink milk. Mammals are, of course, defined by lactation, by mammary glands and the suckling of our infants. It is specifically for neonates, and when mammals grow up, they put away this childish thing and stop drinking milk.
But not everybody does. In the West it’s fairly normal to drink milk in various forms into adulthood. Europeans and their American descendants do, and some African and Middle Eastern pastoralists do it too. But it was not always thus. Most adult humans today, and almost every human in history, do not even have the capability to digest milk. We all have an enzyme called lactase, encoded by a gene called LCT, and its sole job is to digest milk. The sweetness in milk comes from a sugar called lactose, and lactase seeps out of your stomach lining and slices lactose in half to produce the sugars glucose and galactose. Elegant names are not always a preoccupation of biologists.
For most of human history lactase has been active only in babies. After weaning, the gene’s activity is radically reduced, and as a result, for most adults, for most of human history, milk has been off the menu. Most people, for most of human history, will have experienced a full deck of problems that come with drinking milk past weaning. Symptoms include bloating and cramps, vomiting, diarrhea, flatulence, and borborygmus, which is a technical word for a rumbly tummy. The absence of lactase, or its reduced activity, means that the lactose doesn’t get digested in the small intestine, so it passes into the colon, where it encounters bacteria that can break it down and it ferments, causing gas buildup. That’s the direct cause of the bloating and fartiness, but also the increased pressure triggers diarrhea, and so on. This is called lactose intolerance, and admittedly though not particularly pleasant, it’s not the worst condition someone can have, and is pretty normal for most people if they drink milk into adulthood. Which is why most people don’t.
Except if you’re of European descent. Your lactase continues to work throughout your life. This unusual phenomenon is called lactase persistence, and although a splash of milk in tea is the English way, and even a mug of hot chocolate might seem very normal to us, we are the weird ones. There are a handful of African populations, some in Southeast Asia, and a few Middle Eastern peoples whose lactase persists, but for the majority of modern humans, milk equals tummy troubles.
The genetics of lactase persistence are well understood. A very small number of individual changes to the DNA in and around the lactase gene account for its persistence. The vast majority of Brits, and northern and western Europeans (including places colonized by them) have a single change, a C becomes a T, around 13,000 letters of DNA before the start of the lactase gene. This type of mutation is not uncommon, but it’s really where the language metaphor to explain genetics falls down. Although one hopes that the coherence of the chapters and paragraphs and sentences in this book has an overarching theme, in general each idea follows on from the last. In genetics, though, one gene can have a direct effect on another, yet can be miles away on the same chromosome, or even further away on another chromosome. So the random sentence 13,000 characters ago on page 69 was bizarrely irrelevant and totally ectopic then, and I instructed you to ignore it. But it is directly related to this one, and that is precisely what happens frequently in the genome.
Thirteen thousand nucleotides upstream of the beginning of the lactase gene is a region that controls its activity, and a mutation in that distant control center accounts for the vast majority of milk drinkers. By looking in this region for this and other minor variations that have no real effect, and comparing these accumulated changes with other genomes around the world, we can make estimates of how old the lactase mutation is, which comes out in the region of 5,000 to 10,000 years BCE. And the presence of these particular genotypes altogether in a cluster that includes the lactase gene and other bits and genetic bobs, indicates that it was all seriously favored by natural selection.
Now, unless it conferred some other advantage (for which there isn’t any real evidence), it’s hard to see what the evolutionary advantage might be for lactase persistence in the absence of a regular supply of fresh milk. And so we think of this as a classic example of how we have invoked shifts in our genome with our own practices—a gene-culture coevolution—experienced only in communities that were practicing dairy farming with domesticated milky beasts. What advantage having both access to milk and the ability to process it might seem obvious: In fact, it’s really the realm of intelligent but speculative guesswork. A regular supply of nutritionally rich food is one; avoiding the boom and bust cycles of seasonal crops is another possibility.
By 6,000 years ago, milk had become a part of Neolithic life. Shards of crockery recovered from digs in Romania, Turkey, and Hungary hold traces of caked gunk, and these got squirted into a gas chromatograph by Richard Evershed’s team at the University of Bristol. In that long, extremely thin tube, carried by an inert gas such as helium, the ingredients of these residues are separated into their constituent molecules, which move at different speeds as they trickle away from the carrier, and with a reference guide, can be precisely identified. In among the detritus of being buried for several thousand years, there was the signature dairy fats. There’s not many ways they could’ve got there unless they were used to store milk. By 5500 BCE, we were making cheese. Sieves and pottery colanders resembling modern cheese strainers had been found in Poland, and in 2012, again, telltale residues were scraped off these ancient dishes. The suboptimal washing-up skills of the people who owned this crockery again revealed fat from milk. Cheese, of course, is a strange thing in itself, and odd that we should eat it. It’s milk that has gone bad, probably the first processed food, but it may have been a useful way of storing the nutrient-rich milk in solid form, possibly more like a glob of mozzarella than a wheel of Stilton. We know that, much later, the Romans ate goat’s and sheep’s cheese, but used cattle primarily as beasts of burden. They noted that people of Germany and Britain liked to drink milk.
These examples of dirty dishes left for millennia are few and far between, and we need other ways to peer into the past. Lactase persistence is now effectively universal in Europeans (those African and Middle Eastern peoples who also are lactase persistent have different mutations and are unlikely to have a common origin). At some point, maybe 8,000 years ago, it was absent. So the question is to pin down when it emerged and how it spread. In 2007, Anglo-German geneticists dug up bodies from eight locations in eastern Germany, Hungary, Lithuania, and Poland, all from between 5,800 and 5,000 years ago. The Germans’ graves are from cultures of people who were potters, and are known to have had farming practices, probably the oldest in northern Europe. From their teeth and ribs and other bits of bone, the scientists extracted ancient DNA. None of these guys had the mutation that endows the owner with the ability to process milk.
These points of data, genomic and archaeological, set up the rough time frame. They are, admittedly, somewhat imprecise, but such is the nature of the past, truly a foreign country. Where dairy farming began would be more difficult to ascertain.
But not impossible. Mark Thomas at University College London has been in pursuit of the origins of dairy for years (alongside his many other ancient genetic detective works). In 2009, he put together the pieces of genetic and archaeological evidence into a computer model, a kind of statistical jigsaw puzzle, from which the clearest picture of the roots of milk farming would emerge. The distribution of lactase persistence genes in and around northwest Europe today, including Scandinavia and Ireland, had suggested that its origin might have been around there. There had also been suggestions that its evolutionary advantage might have been to do with lower levels of vitamin D in these northern tribes; milk drinking can compensate for that obscuring of the sun.
But that’s not what Mark Thomas’ team found. They computer simulated a scenario of how lactase persistence might have become so widespread by plugging in archaeological data—such as the carbon 14–dated presence of the artifacts of dairy farming, with a careful comparison of the specific genetic differences between three groups of ancient people—foragers, non-dairy farmers, and dairy farmers. We know that the lactase persistence gene was selected by evolution, and didn’t just drift into ubiquity, so the simulation trots out a setup where that version of the lactase gene supersedes the versions in the hunters and milk-less farmers. It maps not where it arose, but where it began to fill the population at the expense of earlier types. When collected, the simulations plot a map showing the highest chance of the evolution of the lactase persistence allele, and it locates it in an area engulfing Slovakia, with Poland to the north and Hungary to the south. This fits with the archaeology, and the residues found in those Hungarian and Polish farmyard digs. At 7,500 years ago, these people were farmers with structured garden farms, where they grew wheat, peas, lentils, and millet. They husbanded cattle, swine, and goats, and occasionally hunted boar and deer on top of their agrarian lifestyle. They used flint and wooden tools, but not metal, and used earthenware vases, jugs, and pots with lined designs, from which we derive their name: Linear Pottery people.
The same Bristol team that scraped the gunk from the dairy pots also inspected ancient crockery for other clues to the diets of our ancestors, and in 2015 found the taste of honey. They analyzed thousands of fragments of pots and pulled off traces of beeswax from just four, dotted as far afield as Turkey, Denmark, and Algeria. In other studies they found the remnants of meat. This collected diet data shows a picture of a people cooking and farming, collecting honey, herding beasts—boar and cattle—and milking them, storing that milk in pouches and pots, sieving the curd from the whey. And as those skills developed and spread across the continent, so did the gene that renders that dairy produce so valuable to us.
There are two important lessons in this story. The first is the answer to a question that people often ask: Are we still evolving? It’s hard to determine in the present, because we can’t very easily see the changing flow of different versions of genes in populations around the world. Evolution is a fourth-dimensional process, it occurs over time, as well as in space, and it happens slowly in most cases. So we have to be patient in assessing its impact, and at the moment we simply do not have enough data. But we can definitively say that we have evolved in our recent past, during a time when we were who we are today. The pressures of natural selection have undoubtedly changed as a result of the evolution of our behavior and culture, but milk shows that, just as in every organism that has ever lived, over time our genes continue to change.
The second point is that the world in which we live is shaped by us, by our practices and culture, by our very existence, and our DNA responds to that in turn. Genes change culture, culture changes genes. Farming has been arguably the single greatest force in changing human culture and biology. So, just think tomorrow morning when you’re pouring milk and dribbling honey into your porridge, that it’s your ancient European ancestors (if indeed you are European) who began a revolution in culture and in your DNA that allows you to tuck into your breakfast. Bon appétit!
One of the more obvious differences between modern humans is skin color. We make crude visual distinctions and effectively meaningless categorizations based on average skin tones, such as black or white. The question of race is explored in depth in Chapter 5, and I will explain why geneticists ascribe no scientific value to these broad racial attempts at definitions of peoples. In parts of Africa and Australia and the islands of the Indian Ocean, people have the darkest of dark skin, almost black in its true sense. In Scotland, or the north of Sweden, some are so pale as to be almost translucent. All skin tones in between exist in abundance. This is not some liberal fantasy, it’s simply a truth that a Pantone swatch of human skin is a continuum.
Nevertheless the physical characteristics of the Europeans are unusual, just as their love of milk is. We can broadly say that many Europeans are blond and some have blue eyes with pale skin. Some are even redheaded. These are mutations in the genetic sense; as mentioned before, this simply means the DNA—the genotype—is altered from what came earlier, and the physical result of that change—the phenotype—is different.
At least eleven genes appear to have a direct role in determining skin and hair color. Overall, it’s all down to the density and type of melanin that you produce. There’s two types of melanin: eumelanin (pronounced you-melanin), which comes in either black or brown, and phaeomelanin, which is reddish and is the root of ginger hair, which we’ll come to shortly. Specialist cells called melanocytes sit at the base of the skin, and produce melanin following exposure to ultraviolet light from the sun. The melanin collects in little specialized pockets called melanosomes, which shuffle between cells, which is why people tan in the sun. They move into position atop the cell nucleus as a means of protecting the DNA within from UV damage, which can slice through the double helix with abandon. In hair follicles, the melanocytes transfer the melanosomes into the base of the growing hair, and that is how your hair is colored.
The melanin is the end point of this sequence of metabolism. The range of hair and skin colors is partly dependent on the types and density of melanin you have, but also a whole battery of other genes that seem to influence how your melanin plays out in your hair and skin. The protection that eumelanin offers to the cell nucleus is an adaptation to sunny weather. Pale skins tan to protect themselves, in a somewhat post-horse-bolting way (so my advice is wear sunscreen), but those in the blazing sun near the waist of the earth are preemptively protected by being full of melanin.
With the genes identified, we can, with relative ease, say when these changes happened and where. Of course, it’s not at all straightforward, and they appear at different times and in different forms. But we can say with confidence that the Africans who populated the Middle East and southern Europe 50,000 years ago were dark skinned. We also know from the DNA of burials in Hungary and Spain and the man from Loschbour in Luxembourg that these hunter-gatherers had dark skin around 8,000 years ago. There’s no trace of genetic fair skin between those two dates. In 2015, Iain Mathieson (and a host of others) identified two important genes with tedious names, SLC24A5 and SLC45A2, and showed they also did not have variants that are associated with depigmentation, that is, pale skin. Seven thousand seven hundred years ago, the Swedish clan in the Motala cave had both pale skin versions, as well as a version of the gene HERC2/OCA2 that also plays a role in light skin and blond hair, but also clear blue eyes. The Swedes have looked like that for a long time.
The new genetics created by genomics allows us to move from speculation about characteristics that might be under the pressure of evolutionary selection to certainty. The most comprehensive study to date arrived in winter 2015, and brought with it the largest European resurrection on record. Iain Mathieson’s landmark paper contained the genomes of 230 people who had died between 6500 BCE and 300 BCE. Some of them we knew about already: Loschbour, Motala, and some of the Hungarian and Polish bodies were in there, but dozens of newly discovered, long-time dead were invited to the party, at least the bits of DNA the scientists could finagle out of the densest bone in their skulls. Reach behind the fleshy lobe of your ear, and you’ll feel a hard bone rising from your skull. This is called the mastoid process. About an inch directly inside your skull from there is a hard ridge of bone called the petrous. Its name comes from the Latin word petrus, meaning rock, as in petrified and Peter, the rock on which the Christian church is built. This is because it’s rock hard. It is therefore a treat for preserving DNA.
So, in this mix were Anatolians, Iberians, Yamnayans, Poltavkans, and Srubnayans—all peoples from the coast of northern Spain to the Altai Mountains in Siberia, some 2,300 years old, some 8,500. With a gang this big, the signs of natural selection become easier to detect, and that’s what they fished for. Again indicative of how far this field has come, Mathieson et al. hunted for patterns of natural selection among more than a million individual changes in the letters of DNA that we call SNPs across all these genomes. Lactase persistence came out top, and a few other diet-related genotypes, almost certainly adaptations to changing diets brought on by agriculture. A couple of them are variants most closely associated with vitamin D, again suggesting a change in diet, and in our changing ability to acquire essential dietary supplements.
They saw selection for DNA associated with celiac disease, a form of irritable bowel sensitivity to the wheat protein gluten, but it’s likely that this was carried along by natural selection with other dietary adaptations; one variant found helps process an amino acid called ergothioneine, which is not abundant in wheat. Early farmers of cereal crops may have found advantage in this allele, and an irritable bowel came along for the ride. They saw the selection of genes for lighter skin and blue eyes, though we know that the people of Motala were already blue-eyed blonds. Light skin from the east spread quickly through Europe, and so the population structure over time was upheaved with a change in people, how they looked, and in agricultural practices. They looked at height too, which we know is a trait heavily influenced by genes, and a little but significant amount by the environment (see Chapter 6). The Yamnaya were nomads and pastoralists, probably shepherds from the western steppes of Russia, who we know were tall from their bones. We can also spot tall genes in their DNA, which they brought with them from the east 4,800 years ago, and these spread alongside all the farming and dietary changes that were sweeping through the continent. That was mostly in central Europe and to the north though. In Italy and Spain, selection favored the short, possibly because of colder weather and poor diets.
The picture of 10,000 years of European union slowly comes into focus. More than anything specific, what these studies show is evolution by natural selection in action in the last 10,000 years. It shows the interplay between movement of people, introgression of cultures, particularly farming cultures, and their profound, measurable effect on the most comprehensive record we can muster. In the genomes of the dead we can see natural selection at work.
We can’t necessarily see why particular genes are being selected though. These are not the types of adaptation that we might glean from natural history programs on television—big visible changes like tusks for showing off to females, or a variety of beaks for highly specialized nut-cracking as you might see in Darwin’s famous finches. But they are Darwinian evolution nevertheless—subtle changes in DNA, and subtle changes in how common that DNA is in breeding populations. The spreading of alleles through a population is the measure of natural selection. There will be many more signals of selection as we unearth more bodies and get more DNA from our European ancestors. These techniques will be and are being applied across Asia and in the Americas, and into the last places on Earth that humans migrated to, the South Pacific and New Zealand. The mix of history, archaeology, and now DNA is building a new picture not just of migration, but of the evolution of us—how we came to be what we are.
“People who haven’t red hair don’t know what trouble is.”
L. M. Montgomery, Anne of Green Gables, 1908
Extinction should be considered troublesome, so Anne was right, if the newspapers are to be believed. Pale skin and blonds may be an adaptation to the north, and compared to the hair of Africans, eastern Asians, southern Asians, and the indigenous people of the Americas, flaxen hair is a rarity. But there’s an even more unusual hair, and that’s red. Red hair is caused by changes in a single gene, and exists in the overall global population at about 4 to 5 percent, making it beautifully unusual. Its increased prevalence in Scots (and the Welsh and English, and other northern European populations) is probably due to a degree of isolation in an ancestral group at some point in our ancient history, but we don’t really know. Around 40 percent of Scots carry at least one copy of this allele, and one in ten are redheads, but worldwide it is the most unusual hair color.
There are interesting stories within this gene. The protein it encodes is called melanocortin 1 receptor (MC1R), and belongs to a broad class with the equally unwieldy name G protein-coupled receptor. These are long, bendy molecules that straddle the cell membrane, and upon receiving the appropriate molecular signal from outside the cell, trigger a metabolic pathway. In the case of MC1R, a molecule sent from the pituitary gland to melanocytes prompts these cells to produce melanin in skin melanosomes. Though most people on Earth produce eumelanin, which is brown or black, in people for whom their MC1R contains a redhead mutation, phaeomelanin is produced. The melanosomes feed into the base of a hair follicle and this is what makes redheads redheads.
Of course, as is always the case in human genetics, it’s not quite as simple as this, and much more interesting. The protein is 317 amino acids long, and there are several different mutations, all of which switch eumelanin to phaeomelanin. All human proteins are made up from different combinations of 20 amino acids, each of which is encoded in three letters of DNA in a gene. In MC1R, if at position 151 you have the amino acid cysteine instead of the more common arginine, you have red hair. If at position 294 you have a histidine instead of an aspartic acid, you have red hair. There are several other mutations that I won’t list here that have the same effect, but this goes some way to explain why not all red hair is the same.
In June 1997, J. K. Rowling introduced the world to a boy named Harry Potter and his best male friend Ron Weasley. Ron has many ginger-haired siblings, including identical twin brothers Fred and George. Three weeks after that first Harry Potter book was published, and presumably a magical coincidence, the first major study of ginger-haired twins was published, including twenty-five pairs of Weasley-ish twins. Three major variants were identified that associated very strongly with red hair, but something interesting emerged from the control group. These included dizygotic twins who were discordant for red—that is, nonidentical twins, one a ginger, the other not. Five out of thirteen of these tested had identical MC1R genes. As was beginning to become abundantly clear at that time, just the presence of a redhead allele was not enough to guarantee a redhead. Genes never work in isolation, and almost never have just one role. The discordance of these twins showed that there must be other genetic modifiers that powerfully influence the expression of the MC1R gene to the extent that the phenotype can be either red or not.
Samples taken from a couple of Neanderthal genomes (one from El Sidrón in northwest Spain, another from Riparo Mezzena cave in Italy) indicate that their MC1R had an alteration at position 307 (a glycine where we have an arginine). As mentioned in Chapter 1, this variation is not found in modern humans with red hair, and there is no Neanderthal hair that has survived the ravages of epochs. But there are some cunning tests we can do to try to work out the color scheme. By inserting the version of the protein that these Neanderthal people had in Spain into cells in a petri dish, we can see not the color itself, but the activity of the cells and then speculate about the color that might result from the cell’s behavior. The different mutations we see in living redheads can reduce the function of melanocytes in different ways, and indeed these cellular tests show a reduced function of the melanocytes. But does that mean ginger? Possibly. The physical distance between the source of the two Neanderthal genomes sampled suggests that they weren’t a couple of freaks, and that we just happened to sample unusual DNA by chance. While blond and pale skin is almost certainly an adaptation to northern exposure, the variant we see in these two chaps is unlikely to be. Think of the tar-black hair of most Italians and Spaniards today. We piece together the past with the clues we can find, and build up a hypothesis we can test and puzzle over. In this case, the truth is that we don’t really know—for now.
In July 2014, the world woke to the shocking news that the perfect storms of climate change and genetics had conspired to mark ginger hair for extinction. The first headline I saw was from the Scottish paper the Daily Record, with understandable concern given the prevalence of ginger in Caledonia. A scan revealed that every mainstream British newspaper carried the story, the Daily Mail, the Times, Guardian, Telegraph, Independent, Mirror, and the Sun, with various pictures of sexy redheaded celebrities, frequently the actors Christina Hendricks, Julianne Moore, and Damian Lewis. Or Prince Harry. Social media and news websites were ablaze with horror. Around the world, National Geographic, the Week and a host of other apparently sensible magazines and news outlets reprinted the story. The headline in the Independent was typical:
GINGERS FACE EXTINCTION DUE TO CLIMATE CHANGE, SCIENTISTS WARN
Broadly, the newspapers were reporting that, according to researchers, climate change is going to make Scotland less cloudy and more sunny. Therefore, the selective pressure that nurtured the allele for red hair is eliminated, and red hair will no longer be of any use to bearers, and will drift off into the great evolutionary dustbin of once-useful traits. This is an excerpt from the article in the Independent, followed by a gallery of famous redheads:
Dr Alistair Moffat, managing director of Galashiels-based ScotlandsDNA, said: “We think red hair in Scotland, Ireland and in the North of England is an adaption to the climate.
“I think the reason for light skin and red hair is that we do not get enough sun and we have to get all the Vitamin D we can.
“If the climate is changing and it is to become more cloudy or less cloudy then this will affect the gene. If it was to get less cloudy and there was more sun, then yes, there would be fewer people carrying the gene.”
Well, no.
Alarm bells started to trip for many familiar with the themes of genes and natural selection. As mentioned earlier, there is good evidence for pale skin being an adaptation to the cold of the north, but none that red hair is too. Furthermore, I’m unaware of any evidence that says that Scotland will become less cloudy as a result of climate change.
Who is Alistair Moffat, and what is ScotlandsDNA? In fact, it is a genetic ancestry testing business, a partner company to Britains-DNA (see Chapter 4), and Alistair Moffat is their founder and chief executive. The story was drawn from a press release from ScotlandsDNA, which coincided with the promotion of a new additional service that tests for the presence of red-hair alleles in a customer’s genome.
Alas, a fiction can fly around the world before the truth has managed to pick the sleep from its eyes in the morning. Many condemned the errors inherent in the content quickly, and focused on the discomfit of PR dressed up as research that journalists sometimes fall for.
In this particular case, it is hard to think of a way that the ginger allele might be extinguished. The selective pressure asserted in this tale is that redheads exist as an adaptation to cloudy weather in Scotland. There is no evidence for that. In order for these pressures, if we imagine they are real, to push the ginger allele toward extinction, ginger hair would have to be a powerfully maladaptive condition, meaning that it causes harm to the individual in the environment in which it exists.
I need not point out that being redheaded is not a maladaptive condition. It’s a very lovely condition. It is an absurdity, offensive to both redheads and geneticists—a group that contains both family and friends—to suggest that red hair might be subject to a force of natural selection so powerful that oblivion awaits. Even actually maladaptive genetic traits, actual diseases with well-understood modes of inheritance, such as cystic fibrosis or Duchenne muscular dystrophy are not likely to go extinct, because carriers of a single copy live healthily and pass the faulty gene on to their children. Via genetic screening and expert advice, these conditions will diminish, we hope, but the prospect of full eradication is as yet unlikely.
After careful consideration lasting no longer than forty-six seconds, I thought of three plausible ways that redheads will vanish from existence:
1. Humankind goes extinct.
2. Not only would every redhead have to permanently stop having sex for some reason, but every carrier of the gene would too* (which means that everyone on Earth would need to be tested).
3. Redheads, and all carriers of a red allele are exterminated (which also means that everyone on Earth would need to be tested).
To be frank, option 1 is the most likely. The absolute truth is that, like so many human characteristics, we don’t genuinely know if ginger-hairedness is a mutation that has a physiological advantage. It is unknown if it is an adaptation to northern climates, to the beloved gray weather of Scotland or Scandinavia. It might be sexually selected, as despite the lazy mockery that red hair sometimes invokes, some of us find it enormously attractive. It might be that the mutation, random as they all are, was neutral, and had no noticeable effect, but drifted in populations largely isolated somewhere in the northern realms of Europe, and became fixed in these populations at the low, special frequencies we see today.
Such is my confidence that red hair will not go extinct as predicted by ScotlandsDNA, I publicly offered, in an article in the Guardian, to support and assist in the repopulation of the ginger people of the earth, should this auburn apocalypse transpire, by maintaining the various alleles in subsequent generations through vigorous procreation. Naturally, I sought my wife’s approval (after I had attempted to explain the extreme unlikeliness of this happening).
In fact, my services would not be further required at all, because I had already done it. When I had my genome sequenced, by BritainsDNA—the very same company behind this entirely specious extinction nonsense—there it was: MC1R Val60Leu. I carry a mutation for ginger hair. At position sixty in the MC1R protein, instead of the amino acid valine, I have a leucine. In the gene, a single letter of DNA is changed; sometime in my ancestry, a G became a C. This was partially surprising, as I’ve got black hair on my head (though it’s flecked peppercorn these days; weary melanocytes give up the ghost as one ages),* no doubt from my Indian heritage. My father had dark brown hair (nowadays he’s a silver fox), and his family, mostly from the northeast of England and Scotland, are to our knowledge largely ginger-free too. It’s also not very surprising as Rutherfords hail from the northeast of England and Scotland, where redheads are unusually common. A few years ago, in the throes of capricious youth, I stopped shaving for a summer and sported a beard. In among the black was a crop of hairs that were the perkiest auburn, like poppies in a scorched field. I plucked a couple and under microscope compared them to a friend’s very obviously red hairs and they were indistinguishable. So it’s there in my genome. My wife is blonde, and of my three children, two are blond, and two have brown eyes; my flaxen son has eyes so blue he could be Swedish. One of them may well carry a MC1R red allele. My wife’s sister is a strawberry blonde, and has two children by a very ginger-haired man, and they have the most glorious bright red hair you could imagine. So, what are the chances of this oh-so-northern-European trait going extinct? Roughly, somewhere between none and zero.
Red hair appearing exclusively in beards is not uncommon, though we don’t really know why. Forgive us; it’s not really been a research priority over the last few decades. The ginger twin study showed that my mutation, Val60Leu, occurred most often in “fair/blonde and light brown” hair, three things I emphatically am not. That again is typical for genetics—presence or absence of gene variants are rarely fully absent or fully present in populations. Such is the nature of human variation: We’re very variable. One version of the allele exists at a higher frequency in Irish redheads. One version has a slight association with a lower pain threshold. One version seems to subtly affect how an individual responds to an anesthetic that dentists frequently use. This is just one gene, with many versions, and it has a variable phenotype, though we can mostly see the outcome with fiery clarity. Even when we know the genome intimately, and the patterns of inheritance, and the history of the DNA, and the migration patterns of the people who carried it, and evolutionary pressures that led to the perpetuation of the genes and the phenotype—even when we know all that, how it manifests can still be mysterious and surprising. Anyone who says differently is selling something.
While we’re in the northwest of Europe, let me indulge in some national pride to scrutinize this sceptred isle, and the finest genetic analyses of a people yet undertaken: Just who are the British?
Archaeologists sometimes use technological cultures as being definitional of people or eras. These are often broad and scattered characteristics incorporating a multitude of skills. The Beaker people, for example, made distinctive drinking cups and vessels over a couple of millennia beginning almost 5,000 years ago, and also had bronze and other metalworking skills, shot arrows, and were spread all over Europe, with many subdivisions across the land. Or before them there were the Linear Pottery people, who were similarly widespread and made pots (though not in a line: the linear name refers to bands of lines on these pots incised into the clay. It’s not the most descriptive definition, but so it goes; that’s what they’re called). These are broad, sometimes woolly definitions that are helpful for us to see vast transitions in culture across large areas and huge populations. Within those potters were the farmers who edged out the hunters of the northwest in the Neolithic.
The tide of people in Europe has been incessant from the first humans onward. Although the sea is not a barrier to gene flow, continuous landmasses make for easier journeys, and the band of sea between us and the French, Dutch, and Scandinavians has slowed down a free flow across our borders over history. We’ve not been aggressively invaded since 1066. We’ve not always been an island nation, though. My homeland, East Anglia, was not the gentle coast of my youth: It was simply continuous land to what is now the Netherlands. This terra firma, called Doggerland, played host to the estuaries where both the Thames and the Rhine flowed out into what is now the English Channel. Today, its shadow is referenced in the Shipping Forecast on BBC Radio 4 as Dogger Bank, a 7,000-square-mile submarine sand mound that was once dry land. Twenty-one thousand years ago, during the period when northern glaciers covered these lands at their maximum girth, the sea was more than a hundred meters lower than today, and the land surrounding Dogger was solid too.
Across this icy bridge trudged mammoths and hairy rhinos, during and before the last Ice Age—and people. We don’t really know who these folk were. Neanderthal teeth have been found as far west as north Wales. Other than that, the human remains are scant, though the detritus of their lives is easier to come across. There’s Boxgrove Man, a 500,000-year-old forty-year-old Homo heidelbergensis, identified only by his shin bone, which has teeth marks on it, from either cannibals or animal scavengers. Around his last resting place on the South Downs are hundreds of Acheulian axe heads that span from 800,000 until his time. And there’s Pakefield in Suffolk, which 700,000 years ago was in the middle of Doggerland, and was the home of an unknown people who also knapped blades out of the flint still common on the Suffolk coast today.
These old bones are almost always named something “Man”—Neanderthal Man, Cro-Magnon Man—which can be a bit tedious given that half of all humans have not been that, but the skull found in a pit near a golf course in Piltdown, Sussex, in 1912, was only half man. For a short while he was a celebrity, but turned out to be a shameless fraud, an orangutan’s jaw jammed into an old but modern human skull, the work, probably, of Charles Dawson, who was so desperate to become a proper grown-up scientist, join the Royal Society, and gain international repute that he engineered this and other elaborate deceits over many years.*
On the beautiful Norfolk coast, there’s Happisburgh (pronounced Hays-bruh), a small village with a lighthouse, a church, a superb pub, and not much else, apart from almost a million years of human occupation. The North Sea grinds away at the sandy coast every year, and houses inch closer to toppling into the brown seawater below. As a result, the past just falls out of the cliffs, and pokes out of the beach. A Paleolithic axe head was picked up by a man walking his dog in 2000, and now the annual digs there recover more tools, and the remnants of hunting and butchery of bison, and the fragmented remains of rhinos, elephants, horses, elk, and a colossal beaver. And then in 2013, human ichnofossils were found. These are trace remains, the evidence of living activity in the absence of the bones themselves. In the clay bed only revealed at low tide, there are footprints. They look like they were made by a size 9 Doc Marten boot, but the dating of these beds says that those feet in ancient time walked upon England’s mountains around 800,000 years ago. Toes and details are not present, as they’ve been weathered by oceans and time, so we don’t know what species these feet were.* Nevertheless, Britain has been host to humans for almost a million years.
The ice retreated, and the seas rose. Some believe a mega-tsunami crashed into Doggerland and slid the land into the sea forever, and since 6500 BCE, just when the farmers were arriving in western Europe, we have been an island. That makes us interesting for both conquerors and geneticists. We put names to people with the cultures from archaeology that appeared to share cultures and geographical origins: Celts, Picts, Angles, Saxons. Some of them are better defined than others. The Romans at least came from Rome, though for the 400 years that the Romans were in Britain most of them were conscripts from Gaul. For all the attempts to put discrete labels on the people of the past, genetics has a habit of defying them.
We haven’t been significantly invaded since 1066, though immigration has trickled continuously since. Before that, though, there were plenty of invasions en masse, rulers who came and stayed, or came and went, and left their cultures behind. The question for the twenty-first century is whether they left their genes. This was the subject of a mammoth project called People of the British Isles in 2015, run by Peter Donnelly and Sir Walter Bodmer, two geneticists not primarily known for their interest in history, but in the genetics of disease, in particular cancer. But genomes are DNA, and DNA is data, so it’s a short jump from the pernicious variation in a disease gene to the benign variation that tracks evolution and history. Of course, overall we are incredibly similar in terms of our genomes. But the levels of detail we can now extract from DNA, using delicate and highly perceptive statistical techniques, are unprecedented, and reveal secrets from the past undetectable by any other means—wispy fingerprints only noticeable with forensic analysis.
Two thousand and thirty-nine people were carefully selected across Great Britain, who spat in the tube and gave up their history. They had been chosen wisely: All had four grandparents who were born within fifty miles of their own birthplace, in order to exclude the waves of immigration from the colonies and elsewhere that came in the nineteenth and twentieth centuries. Many were of retirement age, so their grandparents were decidedly Victorian, which lengthens the odds of isolating DNA that has been as static as possible across the generations that span thousands of years. Six thousand Europeans from ten countries were the template from which they checked, looking for similarities and differences between the people of Britain and the people on the mainland.
The zoom you use on DNA determines the resolution of picture it paints. “Human” becomes “non-African” becomes “broadly northern European.” Magnifying to the minutely fine-scale shadows of British DNA reveals a history of the people, not just kings and queens, but proper hoi polloi. This powerful lens is a great leveler, a historical source less tainted by victory or politics. The genome is a neutral record of sex, sometimes on a grand scale, and admittedly sometimes reflecting the politics of the time. The delicacy with which the traces of ancestry can be extracted can change, or reinforce history. These are patterns invisible but for the subtlest of statistics, long lost webs of family trees that are just threads of silk. From the spit in the tube, half a million individual spots on the genomes of the Brits were identified and fed into a computer, all individual SNPs, and neutral changes, meaning that they don’t convey any significant advantage or otherwise to the owner. Each one was compared to each other one, and compared to each of the 6,000 European controls. This is history enabled by computer power.
The software shuffles people into groups as similar as it can, based on the 500,000 variables on each genome, but by design doesn’t take into account geography. It was only asked to sort people who were genetically more similar to each other than to the rest of the sample, seventeen groups in all, all color coded with a symbol. Then, when the sorting was done, Walter Bodmer’s team placed each grouping on a map of Britain according to the location of their grandparents.
Only at this resolution are they revealed, but the differences are stark. If we were all completely homogenized, then the map would look like pebbles on a beach, and with fewer people in the samples, that is what the map would’ve come out like. But it didn’t. The ancient genetic map of Britain shows clear clusters of people who were more wispily similar to each other than to the rest of the country. The differences are slight but striking, and only visible en masse.
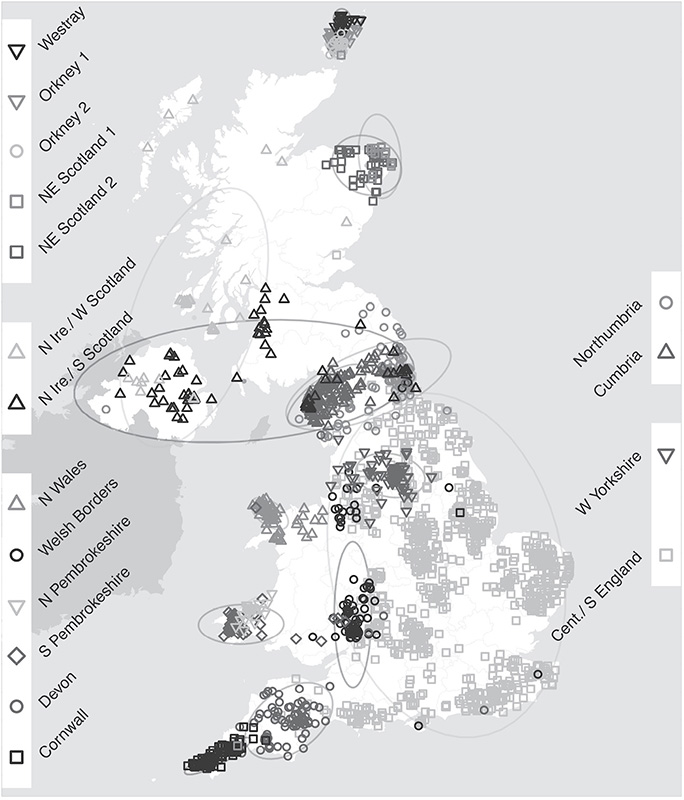
The fine-scale genetic map of the people of Great Britain.
The People of the British Isles project sorted the DNA from 2,039 Britons into genetically similar clusters, and placed them on a map according to where they were born. The map reveals local constellations that reflect the history of the British Isles. (© Stephen Leslie, University of Melbourne, Australia. Based on data published in Nature 2015. Contains OS data © Crown copyright and database right 2012. © EuroGeographics for some administrative boundaries.)
The most genetically distinct are the people of Orkney—a cluster at this level of detail that cuts them off from the rest of us. Maybe that is not so surprising as Orcadians are an island people on their own, and remote even by Scottish standards. Around a quarter of their genome appears to come from Norwegian Vikings, as precisely as 1,100 years ago (assuming twenty-nine generations based on the number of genetic changes, and a generation being twenty-eight years). That places an invasion from Vikings in 916, who stuck around and got on handsomely with the locals.
At this resolution, the Cornish are distinct from the people of Devon, and the borders of this genetic difference are very close to the county lines, reaching from Bodmin Moor to the Tamar Estuary. The Welsh, they look distinct from the rest of Britain too, and within Wales, the north and south settle apart as well. Even within southern Wales, the divisions of culture are borne out in the genome. An invisible line slices through the southernmost tip of the Gower Peninsular, at the bottom of Wales. It bisects Pembrokeshire to the point where that county meets its neighbor to the east, at the River Taf north in Carmarthenshire. Since the sixteenth century this domain has been known as Anglia Transwalliana—Little England Beyond Wales—and the boundary called the Landsker Line. It’s a language barrier more than anything else, and has no formal or legal status, though it has lasted a thousand years. To the south is a pocket of English speakers, whereas the rest of Wales spoke Welsh (at least historically; English is spoken everywhere today). It’s drifted over the centuries, a few miles north or south depending on the local movement of people and traditions, but largely it’s preserved as a vague border a few miles deep. There’s a neat-ish line of forts that map roughly along the line too, Norman castles from Roch on the west to Laugharne on the east, but it’s a stretch to say these were frontiers protecting this informal border.
The People of the British Isles study showed that the Landsker Line is there in the genome too. Residents of Anglia Transwalliana leapfrog over 150 miles of south Wales to share more genetically with the English. Earlier genetic maps of the Welsh had not spotted this and had shown no significant difference between the Welsh as a whole and the English, which is no fault of those genetic cartographers; it’s simply that the latest techniques can pick out these differential ghosts from a genomic mass of similarity.
These are the finest of differences, but they show the remnants of peoples who have, on average, stayed put rather than nomadically roamed. It won’t surprise the people of West Yorkshire that they pop out on this map as being distinct from the surrounding people, a tribal gathering called Elmet. Nor people of Cumbria, which overlies a Celtic settlement called Rheged. The map says profound things about history too. As well as the shadows of regional populations in the UK, the comparisons with European DNA show where bits of the modern British genome originated, or didn’t. There’s very little ancient Italian. The acres and tons of archaeological evidence for 400 years of the Roman Empire in Britain are rich and wondrous. But the biological traces are comparatively absent.
The Celts are a people whom we romantically think of as being tough Welsh, Scottish, Breton, or Irish, with a style of art and culture instantly recognizable with often abstract twisted shield, bolts, and crosses. They slide up the west coast of Britain, from Cornwall, through Wales, and into Scotland, Picts to the north, Saxons to east.
But they’re not a cohesive group of people at all. According to the British genome, Scottish Celts are more different from Welsh Celts than either are from the English. The same goes for the Cornish, who resemble the Breton Celts 250 miles to the south in France. Fiefdoms that we refer to as Celtic stick out, such as Elmet in the West Riding of Yorkshire, Rheged in Cumbria, and Dumnonia in Devon and Cornwall, but are dissimilar to each other at this genetic resolution. Similarities seen in cultures, such as farming techniques, or the patterns on decorative trinkets may be common, but the DNA shows that they are not necessarily rooted in shared ancestry. They may have traded at some point, or copied, or it might be coincidence. But, according to the genetics, there wasn’t a point where a group of genetically similar people spread into the extremities of the British Isles and settled into a culture that we now call Celtic. That word is a modern invention of a presumed people that isn’t reflected in Britain’s DNA.
Wave after wave of invasion until the Normans came shaped the cultures and histories over centuries. Comparing the genetics of the British now with Europeans today can tell us who came and, via close relations with the indigenous residents, left the biological signatures of their visit.
The south is the largest solid block of color on the genetic map, though; a bent rectangle roughly cornered by Newcastle, East Anglia, Kent, and Dorset. We call them the Anglo-Saxons, but they didn’t. They were more like a scattering of peoples from northeast mainland Europe comprising tribes such as the Mierce, Gewisse, or West Seaxe. They followed, or maybe contributed to, the decline of Roman Britain, coming from the countries that are now southern Norway and Denmark and Belgium in the fifth century, maybe migrating away from the influx of people from the east, such as Huns and Bulgars and the unexcitingly named Alans. Following on from the Romans’ exeunt, many theories have been proposed over the years of what happened when we plunged into what is daftly called the Dark Ages—as if Rome was the light, and a shroud fell over Britain as sophisticated culture retreated from our shores. It is true we didn’t write down as much after they skedaddled, but culture abounded, as did agriculture, and historians continually debate how good or bad things were. Some theories suggest that warmongering Saxons turned up and wiped out the abandoned natives, or freed them from imperial tyranny. The genomic map showed a solid block of red that corresponded to the areas that Rome ruled and that the Saxons inherited. It shows admixture, a virtually even spread of genes from older Britons and from European Saxons. They came, they saw, and they just integrated into the lives of the existing population—migration, intermarriage, and assimilation. Unlike the Romans, they stayed, and their genes have endured ever since.
One thing is certain in human history: Nothing is forever.
In the ninth century, the Vikings came from the north. The Danish started bothering our shores in the 800s, with frequent pirate raiding parties to the east coast. They came again with intent in 865: An army led by Ivar the Boneless, Sigurd Snake-in-the-Eye, and Halfdan Ragnarsson arrived in Suffolk to conquer the Anglo-Saxon territories. They marched north and took York and Cumbria, and over the next few years marched back down to London and conquered East Anglia too. Danelaw was established, a ruling body and land that blocked out most of England, with Wessex under Alfred to the west. This territory included England and Scandinavia, with undulating borders and politics, and an influx of Norwegians, until 1066, when Harald Hardrada got an arrow in his throat at the Battle of Stamford Bridge. Harold Godwinson took the English reins for a short spell, until he got an arrow in his eye from a Norman conqueror down south in Hastings later that year. And thus ended the permanent residence of the Vikings in Britain.
Yet there is virtually no trace of the Danes in the British genome. Compared to the Angles’ and Saxons’ and even the Norwegians’ genetic legacy in the north of Scotland, there’s an absence of Danish DNA despite a long adventure here. This says something about their 200-year rule. They didn’t integrate. They may have shaped the lands and defined and defended borders, and given us the days of the week (and hundreds of other words: “a berserk freckly husband is a blundering guest in hell”), but they don’t appear to have left any distinctive DNA. The Vikings might not have been as bloodthirsty as popular depictions; they farmed and wrote and created great art. Their first interactions with the Brits were piratical parties, with assumed rape and pillage, yet the former left no genetic trace. Like the Romans before them, it seems that those Vikings wielded their power from above, absolute Cnuts and Haralds ruling from the top down, with no enduring relations with their subjects.
In earlier and other chapters, I’ve gone to lengths to explain how we are all descended from a very small group of people, and that all lines of ancestry cross in the surprisingly recent past. You are descended from Vikings, because everyone is. If it now seems like I’m separating all these pockets of Brits out as distinct, it’s a facet of the zoom, and the People of the British Isles project doesn’t negate that. Because every person has so many lines of ancestry above them, a branch from everyone will cross at some point. But you still may well have more branches from Vikings than from Angles, and indeed, more lines from Norwegian Vikings than from Danish, who didn’t seem to leave many British descendants at all, as they ruled from afar, and like the Romans before them, didn’t get stuck into the locals much. And although this sifting puts the Welsh as distinct from the rest of the British Isles, it also mocks any modern nationalistic pride that you might derive from that. The north Welsh are as different genetically from their southern compatriots as southern Englanders are from the Scottish, or the people of Devon from the Cornish. Even if you’re more Welsh than your neighbors, it’s only in the details, and you’re still part Viking, Saracen, Angle, Saxon, and, as we shall soon see, part Holy Roman Emperor.
This map of the British Isles is unprecedented. It ties history and people—the people—the normal folk who arrived and traded, settled and integrated in farms or homesteads with people already there. It shows a picture of simultaneous homogeneity and diversity across our lands. What a beautiful map it is too; to see those symbols cluster according to ancient lands is invigorating and reassuring. We’re all the same people, we’re all slightly different, and the differences show up in funny pockets, reflecting a land that has such a profound and rugged history, but that has been pretty stable now for centuries. I think of my children, mutts with their genomes from south Wales and Ireland from their mother, and from the northeast and Scotland from me, plus a dollop of south Asian to spice things up. They are our twenty-first-century national dish: a very British curry. The same techniques can now be applied to us in the postcolonial era with the rich flow of the peoples of the world that now make up our countries, and it can be rolled out across every nation on Earth.
There are other strange tales further to the north. Sixty years after people first set foot on Iceland, the baron of Thingvellir murdered a local. Soon after, he stood trial before a court of his peers above a wide volcanic plain. As punishment, Thingvellir’s land was seized by the local families, and they decided to exile him, not abroad, but to the interior of their island. There, in its barren, lunar terrain he would surely live out his days quickly. The new common land became the site of the world’s first national democracy, grounds set for assembly and lawmaking. The Icelandic parliament has now moved into the capital, Reykjavík, but it is still called the Althing. Thingvellir’s bleak, brutal but green land (when not blanketed in snow), perches atop a jagged cliff overlooking the mile-wide valley that separates the North American tectonic plate from the Eurasian one. Iceland straddles a rip in the earth’s mantle, and the two plates riven through the land are slowly pulling it apart, at the speed of fingernail growth. There is the source of Iceland’s living rock, still angry to this day—Eyjafjallajökull and Bárðarbunga, and a smattering of other minor volcanoes blowing their tops in the last five years.
Iceland is a weird place. The living landscape is virtually extraterrestrial—glacial and barren, volcanic and lunar, and its culture and language rich and bewitching. Its latitude means that the winters are black all day, and the summer sun barely dips below the horizon. Even the neighboring Scandinavian countries think the Icelanders are a bit odd. The country has an extremely uncommon history, which makes it paradise for a geneticist. It is my second favorite country on Earth, other than the land of my birth.* The tales of the first families are logged in the Icelandic Sagas, collectively one of the most important European texts. Written over a sixty-year period, they contain events of world-changing significance, such as the first steps of European feet on North American lands, and plenty of ghosts and trolls, sex, violence, drinking, and subsequent puking, as surely befits the Viking reputation.
The genealogy of virtually every Icelandic settler is recorded. Gardðar Svavarsson was probably the first,* lost in the north Atlantic in the early 860s, but he only stayed a winter. Flóki Vilgerðarson was next according to the Landnámabók—The Book of Settlements—which documents Iceland from first residents to its time of writing in the twelfth century. Ingólfur Arnarson and his family arrived in 874 CE, and took their place as the first permanent residents in the southwest fjords, a place he called Reykjavík—the Bay of Smokes. Other settlers followed that year, and the Landnámabók lists more than 400 families. By the formation of the Althing in 930, according to the Landnámabók, there were 1,500 farms and place names and more than 3,500 people.
Island populations often breed strange and interesting genomes. Because Iceland is so small, with a precisely known chronology of inhabitation to the present day, we have the most comprehensive log of everyone who has ever lived there since the ninth century. There have only been thirty-five generations of Icelanders. Since the end of the era of settlement, there’s been very little immigration into Iceland of note. The population has never topped 400,000. These are all matters that make genealogy and genetics significantly easier. The Icelanders knew this, and decided with great foresight to add DNA to their database of identity. Theirs is now the most comprehensively studied genome of any people, both ancient and modern.
Since the beginning of the twenty-first century, the genetic genealogy of Iceland has been explored. As often happens in this business, the mitochondrial DNA and the Y chromosome were first, as they leave the most obvious genealogical tracks. And those tracks are again unusual. Two thirds of the mitochondrial types are most closely associated with Scotland and Ireland. The rest and the vast majority of the Y came from Scandinavia. With little immigration, and a perpetually small population, these numbers give a much better handle on the origin of these DNA types than in admixed countries like the UK, or continuous landmasses such as Europe.
Consider what that means. Male Scandinavians bred with Scottish and Irish women. In the People of the British Isles project, the Orcadians came out as the most different.
Norwegian Vikings came to those beautiful islands repeatedly, but unlike the Danish in Britain, enjoyed local hospitality and left their mark very clearly. Scotland and Ireland were known destinations of the Norway and Denmark Vikings’ seasonal raiding expeditions. They clearly ventured further west and north too—the men, that is—and acquired wives en route. Whether those acquisitions were forced or voluntary is unknown.
That’s the story based on Iceland’s contemporary DNA. But with such a short history, the past was not far behind. Over the last couple of centuries, Icelandic archaeologists have dug around in the volcanic soil looking for the burials of the first wave of immigrants. In 2009, the teeth of ninety-five Icelandic settlers from the tenth century were wrenched from their jaws, and gave up their DNA to the small army of Icelandic geneticists led by Kári Stefánsson. He had traced his own ancestry back to 910 CE, to a warrior poet described in the Sagas as famously unattractive.
The graves are scattered around the island, many in and around Reykjavík, some to the north, but mostly on the periphery, as the interior is an unforgiving dominion. These were people who are thought to have died before 1000 CE, including probable jurors at Baron Thingvellir’s murder trial. The National Museum’s skeleton collection hosts 780 former Icelanders, but most of them haven’t been carbon dated, so their ages are not well documented. There’s a neat historical cheat code here though, which is that Iceland officially converted to Christianity around 1,000 years ago, and burial practices changed as a result. The Norse buried their dead in north-south orientated graves around the homestead, mostly as singles, sometimes in small family plots, and included weapons, jewelry, animals, or boats that might serve them on their way to afterlives in Helgafjell, Hel, or Valhalla. Christians like cemeteries and east-west graves with none of the fun stuff.
The DNA extracted from these settlers confirmed to a degree the earlier studies of contemporary mtDNA, but was different in a very interesting way. The regions of the ancient mitochondrial genomes they looked at were more similar to contemporary Scots, Irish, and Scandinavian mtDNA than they were to contemporary Icelandic. This is a facet of the phenomenon of genetic drift. This occurs when the sampling of a larger population is not representative of the overall population. Imagine you have a bag of a hundred marbles, fifty blue and fifty green. If you sampled eighty, you’d probably get close to forty of each, and the ratio would be maintained if you were to seed a new bag based on those eighty. If you only took one, though, you’d have a 50:50 chance of getting a blue or a green, and if that was the basis of a new bag of a hundred marbles, suddenly, you only have blues. This effect is seen frequently in evolution when a founding population is small compared to the people from whom they came. This is called a founder effect, when their misrepresentative genomes become the basis of an entire population. In small groups it happens faster, and that is what we’ve seen in the Icelanders. Their founding genomes have remained similar in the countries whence they came—the north of the British Isles and Scandinavia. But there on the island, with no significant input from other sources for more than thirty generations, the genomes of the Icelanders have quickly drifted away from their founding mothers.
The Icelandic interest in genetics has moved on apace as the genomic landscape has evolved. Kári Stefánsson and his company deCODE are the driving force behind all things Icelandic and genetic, and they’ve set the bar high for the rest of the world. In 2015, they published the most complete set of the genomes of a people yet, the complete genetic readout of 2,636 Icelanders,* and sampled more than 100,000 others. This scale reveals patterns unseen in individuals, and so many rare or unspotted disease-causing or disease-related genes were found. Patients were found with a version of the gene MYL4, which causes an irregular heartbeat called early onset atrial fibrillation. They found versions of genes that trigger gallstones, others that strongly associate with Alzheimer’s disease. A very odd thyroid condition showed its hand in a mutation that boosted a hormone if it came from the mother, but lowered it if from the father. That is difficult to explain, but it highlights the complexities of human genetics that we are only just beginning to understand.
They saw something else too: a very high level of genes that just don’t work at all. Many mutations in genes just subtly—or in some cases profoundly—change the shape of the protein they encode, and therefore the way that protein behaves. Those small changes account for much of what we see as human variation—red hair, blue eyes. Some mutations introduce a termination message before the end of the protein, kind of like a period before the end of the sentence. Some insert extra letters in such a way that the whole protein is out of frame; I compare this to a 35mm film running out of sync with the projector gate, so you see half the picture you’re meant to—they’re called frameshift mutations. If I only use three-letter words, it demonstrates the problem:
the big red fox ate cat pie
Inserting a random letter (x) near the beginning, but maintaining the three-letter structure really messes things up:
txh ebi gre dfo xat eca tpi e
Versions of each of these can also simply make a protein functionless, out of step with its unmutated code, and these are called “loss of function mutations” or LOFs. deCODE’s big 2015 papers showed that the Icelanders have way more of these than most people. One in twelve of their population has inherited a LOF. Across the population sampled, they found 1,171 of these totally nonfunctional genes. These were found in living people, so they are clearly not mutations that are not conducive to life, nor fatal during development. We have 20,000 genes or thereabouts, so you might think that missing one or two might not be such a bad thing. But it really depends on which one you are short of. The 1,000-plus identified in Iceland are the starting point for working out not just what each of these genes does, but what redundancy is built into our biology such that losing whole genes does not prompt loss of life.
And perhaps unsurprisingly given the island, and the small founding population, they found a high degree of homozygosity—inheriting the same gene from both parents, and thus increasing the likelihood of a recessive condition appearing. This is a facet of inbreeding, of what geneticists call a “small effective population size.” With such a persistently restricted population, incest is a constant specter, and we know how that can affect families (see Chapter 4, part 3). They know it too, and caution abounds. Whereas in a small town with typical patrilineal naming, you might raise an eyebrow if being chatted up by someone with the same surname, the Icelandic naming convention does not volunteer a meaningful alert to possible genetic proximity. The father’s first name becomes the root of the child’s surname, with daughters adding -dóttir, and sons adding -son. So for example, Iceland’s most famous child is surely the singer Björk,* whose surname is Guðmundsdóttir, the daughter of Guðmundur Gunnarsson, son of Gunnar Guðmundssón. In 2013, software writers Sad Engineers Studios created Íslendinga-App. It’s effectively a database of most of the people who have ever been recorded as having lived in Iceland, drawing its data from the Íslendingabók genealogical library built from records dating back more than 1,000 years. Two users can link their apps on their smartphones together to see how they are related, and genealogical proximity sets off the Sifjaspellsspillir alarm—the incest spoiler.
What a strange land, and a wonderful people. If there is a nation on Earth better studied, from their origins right down to their DNA, I am unaware of it. Compared to the rest of the world, their roots are not deep, and as a result their genetic diversity isn’t either. But their records certainly are. Sometimes a great lack of diversity can be fruitful, but only if you’ve got the foresight and imagination to make it flourish.
Let’s leave the human genome for a moment to dig into one that has shaped empires, culture, and devastation across Europe and beyond. It’s not just our own DNA that tells us our history. Yersinia pestis is a fairly typical rod-shaped bacteria, a single cell around two micrometers in length (ten times less than the width of a typical European head hair), and stationary with no propeller or other means of movement, as some motile microorganisms have. Yersinia is unremarkable—it was discovered by Swiss-French bacteriologist Alexandre Yersin in 1894, and acquired his name in the 1960s.
Now, imagine you are Xenopsylla cheopsis, an Oriental rat flea, tucking into a typical meal by biting an animal, say, a species of fat Russian squirrel called a marmot. You have two sawlike laciniae, which lacerate the skin, and in between them is the hollow needle we call the epipharynx. Together these form a channel through which your saliva can run into the victim, and blood is sucked out. Itchy and annoying to the marmot, but that’s how a flea gets its fill.
All animals are replete with bacteria, inside and out. Most of these organisms do nothing harmful, and many are positively useful. Animals are all different though, and a marmot is different from a flea is different from a human. Yersinia can fester in marmots with no particular effect, and does so to this day in the prairies of the Asian steppes. As a voracious flea, you suck up some Yersinia cells, which pulse down into your digestive tract along with all the rest of your bloody meal. In the warmth of your mid-gut, the bacteria start to multiply, and produce proteins that help them coalesce into lumps with clotted blood, and acquire a thin membranous skin to hold them in place. This makes it difficult for you to digest the blood you’ve already consumed, so you, the flea, still feel hungry and hop onto another animal and try to get some more food. This time, the fresh blood you suck floods into your mid-gut and dislodges some of the lumps, filled with now rampant Yersinia, which rush back down your laciniae, and into the poor cretin you’re chewing on. Yersinia might not have legs, or even the whiplash rotor that many bacteria wield. But it sure knows how to get around.
This is not a particularly unusual life cycle of a parasite. Every known organism has parasites, and the relationship between host and predator has been a major driver of many aspects of evolution. But if that second critter that the flea is trying to get some nosh out of happens to be not a marmot, but a large bipedal mammal of the species Homo sapiens, then what follows is seriously bad news. Your skin is normally the first defense against infection, but Yersinia has already breached that by persuading the flea to try to pierce the skin to feed, and then regurgitating the pathogenic cells straight inside. Once there, it starts doing a whole range of things to further its own life cycle, and in doing so, end yours. It switches on genes that make proteins that stick to your epithelial cells, that is, cells that are on any wet surface—in your gut, in your mouth, lining your blood vessels—and invade them. It makes proteins that render it impervious to phagocytosis—the process where your large hungry Pac-man immune cells called macrophages literally swallow invaders and digest them. It produces proteins that punch small holes in your cell membranes, and these allow the bacteria to seep out to infect more cells. Yersinia blossoms and proliferates in lymph nodes, where it can evade our highly evolved immune system defenses with an arsenal of nefarious tactics. The host cells rupture, and spewing their diseased guts into the intracellular milieu invokes the classic signs of an inflammatory response: tumor, rubor, calor, and dolor—swelling, redness, heat, and pain. Some of your metabolic pathways are ectopically activated by Yersinia, which prompts the macrophages to submit to the persuasive will of their invaders and commit suicide.
Cellular self-destruction is a healthy normal part of living, as cells have completed their use and need to be disposed of, such as in the webbing between an unborn child’s fingers. But this suicide is not part of the body’s plan, and absolutely unwelcome. The macrophages are wiped out, and this has the domino effect of weakening your immune system further. Pain and fever become crippling headaches and your body is plagued with swelling. Cell death in blood vessels denies the oxygen and nutrient supply to the extremities—and as the cells of your toes and fingers die, they become necrotic, blackened, and seeping with pus as gangrene sets in. Your lymph nodes swell like purple water balloons—buboes—notably in your armpits and your crotch, but by this stage you’re too feeble to do anything much. You are riddled with the bubonic plague, and a fortnight after that hungry flea puked Yersinia pestis into your bloodstream, you’re dead.
You might get it in your lungs, and cough and wheeze and spread the airborne bacteria to other people.* Or it might infect your bloodstream, with a host of other cellular apocalyptic traits, and as with bubonic and pneumonic, septicemic plague will end you, if not treated. Nowadays, we have good treatments, antibiotics that will block Yersinia’s ability to reproduce, and if taken in the first few days, plague is easily manageable. Nevertheless, outbreaks do still occur—as in Colorado in the United States in autumn 2015. These days they are restricted to individuals, and quickly identified, and subsequent quarantine and treatment with antibiotics stops the spread dead.
In the gloaming of the Roman Empire, matters were much worse. Rarely has a disease altered the course of a continent’s history so profoundly, but Yersinia pestis managed it at least twice. The first decimation occurred in the sixth century, the epicenter of the infection being Constantinople. There, Justinian ruled the Byzantine Empire, including during a plague that lasted just a year, in 541, but at its peak was an apocalypse. The historian Procopius details some of the devastation of Constantinople in his book Secret History, written at the time. He suggests that when it was at its most virulent, 10,000 people per day were dying, though modern historians estimate a more conservative but no less baffling 5,000 every day. Over this bout of infection, something like 25 million died across the empire. Numbers are difficult to verify at that time, but with the waves that followed in the next couple of centuries, this number is estimated to have doubled. This pandemic extended its necrotic fingers all over Europe, to Germany, France, Italy, Spain, and even North Africa. Historians speculate that the obliteration of so many souls over the waves that followed for the next two centuries made significant contributions to the end of Rome’s rule, and plague-weakened armies opened the door to people of the Middle East.
Procopius describes plague victims in a port near Suez in Egypt in 542 with the classic symptoms seen during the Black Death a few centuries later (which we will come to in good time):
A bubonic swelling developed; and this took place not only in the particular part of the body which is called boubon, that is, “below the abdomen,” but also inside the armpit, and in some cases also beside the ears, and at different points on the thighs.
. . . Upon opening some of the swellings, they found a strange sort of carbuncle that had grown inside them. Death came in some cases immediately, in others after many days; and with some the body broke out with black pustules about as large as a lentil and these did not survive even one day, but all succumbed immediately.
This is the first description of what has been presumed to be that most virulent and fatal form, bubonic plague. Where there is corn, there are rats, and he suggests that trading corn with Egypt may have been the source of those first infections.
Genetics says otherwise. Work has been feverish in the last few years, as ancient genetics has found its feet. The dead are abundant when it comes to the various plagues, and we’ve acquired plenty of samples of Yersinia pestis from all over the world now, and taken out their genetic code. Nineteen teeth from twelve sixth-century plague pits in Aschheim near Munich provided the source of the Code of Justinian. In among the ancient human DNA are the remnants of other species that loiter around our bodies. A 2013 study ground out DNA from those teeth and found without doubt the same Yersinia pestis we see today. This had settled a long running debate about whether that great plague was in fact bubonic. But it also allowed us to place the genes of the Bavarian graves in an evolutionary tree, just like we do with humans and any other species. The similarities were unequivocally eastern, and not African as several historical sources had suggested. It appears that the trade from China, possibly along the Silk Road, had ferried these terrible bacteria from their benign existence in the east. The reservoirs of Yersinia pestis today are in rodents in the steppes, and, though this is speculation, it is easy to imagine that these might have been the source of the fleas that were harbingers of death for so many millions.
And it came from the east a second time. Halfway between two London icons—Smithfield’s meat market and the theatres of the Barbican—is the East Smithfield Black Death cemetery. This part of town is now lush, salubrious, and fashionable, but under the surface there is little but death. A two-hectare site in one of London’s typical squares has been excavated in the years since 1986, subducting its edges under the Royal Mint. Scraping away the veneer of august London prosperity revealed a horror story. Inhumation is the word we use to describe the burial of the dead, and between 1348 and 1350 there was mass inhumation in London. The Black Death had made its way from mainland Europe to Britain, where by 1350 it had cut through a third of the population, and this site was annexed as the first plague pit.
It’s a human dumping ground, a midden more than a cemetery. Christian burials were a social necessity in those days, but people were dying at a rate where the living were barely capable of burying the dead. Some estimates suggest 200 people per day, and so East Smithfield was hastily designated a place for inhumation of thousands and last rites on an industrial scale. It’s known as a “catastrophe cemetery,” and of the 600 bodies catalogued so far, some are neatly lined up, others strewn willy-nilly, tossed to their grave in haste. The demographics of who is in the pit are way out of kilter with normal patterns of death in the fourteenth century. Mortality typically picks out the very young and the very old. In the plague pit, around a quarter are children, and most of these younger than five years old. But the rest are adults, and most of these are younger than thirty-five. These numbers show that the plague struck hard and fast, and wiped out a large proportion of an otherwise healthy population.
Johannes Krause pulled DNA out of this pit, but he was after the bug, not the people. By probing the full sequence of the London plague for the very first time, Krause and his team unearthed the evolution of Yersinia pestis, and the genomic tracks of its terrible journey. An earlier study had shown that, just like the Plague of Justinian, the Black Death in the 1340s had also originated in China. With a publicly available database of the full sequence, the history and the genetics can be aligned. Over a five-year period we can track a course from Russia to Constantinople, to Messina, to Genoa, Marseille, Bordeaux, and finally London. All these ports acted as points from which radiation of the plague could crawl inland. En route, it claimed the lives of some 5 million people.
Just as in the Byzantine Empire 600 years earlier, wave after wave of outbreak crashed into Britain’s population in the centuries after the fourteenth, and it was only after the Great Fire of London in 1666 that this pandemic was crushed. Krause’s work also shows that it never really went away. The pandemic might have ended, but the strains of Yersinia that cause bubonic plague outbreaks to this day are identical.
Let’s return to human DNA for a moment. Across Europe, the Black Death left its dark mark, in the buildings, in the churches, and in the wretched huddled masses in pits below our feet. When a force of nature is this powerful and this aggressive, it can leave its signature in our DNA too. In 2014, an analysis of some of the genes in our immune system showed the hallmarks of evolutionary selective pressure in Europeans, including the Roma, Europe’s largest ethnic minority. (Britons and Americans have traditionally called the Roma “Gypsies,” but this is often considered derogatory.)
The origin of the Roma is northwest India, though populations have migrated into Europe on several occasions, and settled permanently in near eastern Europe in the eleventh century. This is an unusual and useful population structure for the archaic geneticist, as there hasn’t been much admixture between the Romanian Roma and other Romanian Europeans. And for at least a thousand years, both populations will have endured the same environment, the same climate, and the same evolutionary pressures.
Of these forces of nature, the Black Death was a big one. An Indo-European union of scientists used the Roma to scan for the imprint of plague on our DNA. Mihai Netea from the Netherlands, working with Romanian, Spanish, and Indian scientists, sifted through publicly available genome databases and looked at the myriad immunity genes that we possess, looking at 200,000 SNPs—those individual variations in spelling that make up much of the difference between you and me and everyone else. In this pool of Romanian Europeans and Romanian Roma, they saw evidence for a cluster of genes undergoing evolution by positive selection. It’s a process called a “selective sweep,” and is described in more detail in Chapter 1. This signature was absent in people of northwest India whence the Roma once came (and also absent in Africans and Chinese). The presence of positive selection and lack of mixing of genes between the Roma and the Europeans is a telltale sign that these two peoples underwent a similar evolutionary force in the last 1,000 years, and that same force was not experienced in India. The Black Death never struck the subcontinent with the same vigor with which it blew through Europe, and the burden of mortality can be a great driver of evolution. Whether that is a result of chance or some innate immunity of those people is unknown.
The results indicate a strong association, but Netea’s team back it up with some neat fine-tooth combing. Within the region of DNA that was apparently undergoing evolution in response to the plague is a family of genes with typically clunky names.* They’re called Toll-like receptors, or TLRs, and the proteins they encode sit on the surface of immune cells such as those hungry macrophages and sentinel cells. There they vigilantly await the advent of microbes with very specific markings. Upon identification of such an invader, an immune klaxon goes off, and the innate army of cells that protect us from within is activated. TLRs 1, 2, 6, and 10 are the combination that recognizes Yersinia pestis, and there they are, as a cluster, sitting right in that zone that has subtly but measurably evolved in Romas and Romanians, but not in the rest of the world. The mark of the plague is in our genes.
Procopius’ Secret History describes the Justinian bubonic plague. But as we get better at reeling in the ancient DNA from out of old bones, the forensic work in piecing together the past gets deeper and richer, and tells us a different story. The latest analysis in this cycle of death was published in autumn 2015 by the Danish geneticist Simon Rasmussen, after his team had gone on a plague fishing expedition. Unlike the previous efforts that had looked in known plague graves for the genetic shadows of the bacteria, they simply surveyed the genomes of 101 people from graves scattered around Europe by pulling out the soft core of teeth from Bronze Age tombs. This technique demonstrates quite elegantly how the game of ancient genetics has changed in the last few years, as they effectively were trawling in a pool of 89 billion fragments of raw DNA. On a fishing trip like this, scientists need to know what they’re looking for, and out of this infernal mess of DNA fragments, much of it contamination from other organisms, Rasmussen pulled out the last remnants of the genome of Yersinia pestis in Poland, Russia, Estonia, and Armenia.
Rasmussen’s team decoded these bits of Yersinia genome and compared them to other strains, and to another closely related species, Yersinia pseudotuberculosis, which is not so malevolent and lives in soil. These comparisons set a molecular clock for when all of these versions diverged from each other,* and calculates the time when a most recent common ancestor (MRCA) existed. The number comes out at 5,783 years ago, thus predating the earliest reported infections by many millennia. That doesn’t necessarily mean that we were infected in those ancient times. But among all that genetic debris, Rasmussen managed to hoick out fragments that encode some fifty-five specific known proteins that endow contemporary Yersinia with such lethal virulence. All were present except one, called Yersinia murine toxin (Ymt)—which is the one that allows the bacteria to successfully live inside the flea’s gut, by protecting it from being digested. It’s Ymt that makes the flea the carrier, but it wasn’t there 5,000 years ago. They calculate that it was in place by 1000 BCE, and probably was acquired from another bacteria species donating it. Bacteria can do that; it’s called lateral gene transfer, and is an essential part of bacterial evolution. They extend a spear, in Latin a pilus, and short bits of DNA can float across and get incorporated into another bacterial species’ genome. We can spot how this has happened in the past due to the presence of flanking bits of DNA that denote a jump from another species. So, Yersinia in those Bronze Age people was not there by dint of fleas. Without that, and with a slight shift in some of the other genes necessary for delivering full-blown plague, it is likely that infections and outbreaks were not bubonic—the most lethal form—but pneumonic and septicemic. The population of Europe has fluctuated since the introduction of farming over 8,000 years ago, in a sort of boom-and-bust cycle. One of those busts occurred at the time when the fourth millennium BCE was turning into the third, and now we know that plague pathogens were present. It is speculation, but not unreasonable, to suggest that plague had a role in the shaping of this period of Europe’s evolution, especially given the annihilations that would follow in recorded history.
Plague is a specter that has been with us for much longer than we can remember. It is the most deadly bacteria known to history, and still haunts us today. These strains are effectively identical to the ones found in the mass graves of east London, but the severity of infection is less. This isn’t because of the DNA of Yersinia, and not just because of readily available treatments of modern scientific medicine. We have evolved to cope better with this killer—genetically, socially, and culturally. Pandemics are always just around the corner, but it’s now unlikely that plague will devastate the lands as it did repeatedly in history. It is still very present on Earth, and will probably always be so, but we have taken much of the sting out of its tail.
The irony is that Yersinia pestis doesn’t care about us at all: Its life cycle is primarily enabled by hopping in and out of small mammals that it doesn’t particularly hurt, via its preferred host, the flea. We do get fleas and nits, and crabs, but not as much as our more hirsute mammalian distant cousins. The symptoms, the virulence, the catastrophic wave after wave of merciless death that it has handed out indiscriminately to us over five millennia have shaped Europe by crippling armies and toppling empires, and yet it’s just a side effect of the blind indifference of natural selection. We brought it on ourselves, by farming and trade and prosperity: Where there is grain, there are rats, and where there are rats there are fleas. Evolution is an arms race, a permanently fluctuating ping-pong between genes in the hosts that carry them and the organisms they feed on, and the spiraling cycles in the struggle for existence. Yersinia found fleas and made them its slave; the flea likes the rat, the rat skulks omnipresent in our shadows. And in the belly of fleas over 5,000 years, with nothing but a simple will to survive via that ancient and universal method of natural selection, Yersinia pestis has mutated its genes to do its bidding and, in turn, mutated our own genes in response. DNA sculpts the history of Europe, and the world.
The tales of the European adventures are all modern compared to the slow creep from the motherland in eastern Africa. Although still difficult to imagine, that journey played out over centuries or millennia. The nearer we get to the present, the more the increments decrease, and the more graspable human evolution becomes.
It’s worth taking a moment to examine the timescales. Out of Africa is a theory that is supported by reams of data and is broadly incontestable today. But what it means when we think of the movement of people over hundreds of generations and thousands of years is deceptively simplistic. Perhaps it is the word migration that is so misleading. It means something so specific in modern history, and currently is a hotly debated topic. Migration over an evolutionary timescale is an entirely different phenomenon. A thousand miles over a thousand years is barely movement at all, and the archaeological remains of those movements are scattered wide and far. But that time and those years equally provide the necessary basis for seeing the movement of genes, and how they spread through populations over generational time, of how they are selected by the subtle, largely invisible forces of evolution, and equally how they are deselected. In some senses, archaeology was waiting for genetics to come along to provide a measure that was dependent on unimaginable oceans of time in order for it to make sense.
It’s worth stressing again that although getting DNA out of ancient bones is easier now than it ever has been, it’s still incredibly difficult. The aim is to get uncontaminated genome-wide samples—not just fragments, but large chunks from all the twenty-three chromosomes. In fact, in most fossils from which DNA can be retrieved, only 1 percent is of the organism itself, the rest being contaminating bacteria that have been feeding off the remains for thousands if not tens of thousands of years. It’s in these big pieces of DNA that we can really make sophisticated comparisons and claims about the movement of people in history. There’s not exactly an abundance, and although DNA from ancient Europeans is now readily available, there’s virtually none from Africans. And the reason is largely to do with heat. DNA kept in a bone in a cold dry-ish cave is much more hardy than DNA stored in a hot humid cave. Although Africa, the nursery of humankind, has given up her bones in abundance, she will not yield the DNA of her children as easily.* But the richness of Neolithic DNA, coupled with the genes of modern Europeans, and all the bountiful archaeology that litters European caves and the homes of our ancestors is reconstructing the history of Europe in ever-increasing detail.
All these results are subject to change, or at least refinement, as a sophisticated picture of the movement of people over the world develops, fed by archaeology, language, culture, and now DNA. It is going to be much more complicated than those Dad’s Army arrows pushing over continental Europe. We can see now where the genes came from, and how culture, specifically farming culture, changed our DNA permanently. Those ghosts in our genomes were just waiting to be discovered, so they could show us how they lived their lives.
And as we progress through time, and from deep history into the last millennium, we enter a time of kings and queens whose lives and lineages are documented in an unprecedented way. The Swedish Motala clan and Loschbour in Luxembourg, and all of those we have resurrected, were just normal people, and only because DNA is the ultimate leveler is their role in the history of humans now being told. Almost everyone who ever lived will be forgotten. But as Europe—and indeed the rest of the world—evolved, tribes grew into fiefdoms, into states, and eventually into empires. People came from the east, and from Africa. Rome petered out, and upon that rock Christianity grew. And while those empires of Europe rose and fell, they were largely oblivious to the existence of two immense continents thousands of miles to the west. The Viking Leif Ericson was the first European to set foot on these lands, and he called it Vinland—possible meanings include meadow-land, or a reference to the wine-berries or grapevines that they found there. Centuries later and more than two thousand miles farther south from Ericson’s first steps into the Americas, Columbus made landfall in what we call the Caribbean. There they met locals, who became generically known to the Spaniards as indios—Indians—because Columbus thought he had reached the East Indies. These vast landmasses span just shy of half the circumference of the earth, and the first peoples had been there for more than 20,000 years. But with the arrival of Europeans, these American lands were all set to change.