what is dreaming and what does it tell us about memory?
You are terrified and running along a dark, narrow corridor. Something very evil and scary is chasing you, but you’re not sure why. Your fear is compounded by the fact that your feet won’t do what you want—it feels like they are moving through molasses. The pursuer is gaining, but when it finally catches you, the whole scene vanishes . . . and you wake up.
Almost by definition, a dream is something you are aware of at some level. It may be fragmentary, disconnected, and illogical, but if you aren’t aware of it during sleep then it isn’t a dream. Many people will protest, “I never remember my dreams!,” but that is a different matter entirely. Failing to remember a dream later on when you’re awake doesn’t mean you weren’t aware of it when it occurred. It just means the experience was never really carved into your memory, has decayed in storage, or isn’t accessible for easy call back.
We all intuitively know what a dream is, but you’ll be surprised to learn there’s no universally accepted definition of dreaming. One fairly safe catch-all is “all perceptions, thoughts, or emotions experienced during sleep.”1 Because this is very broad, there are also several different ways of rating, ranking, and scoring dreams. For example, one uses an eight-point rating system from 0 (no dream) to 7 (“an extremely long sequence of 5 or more stages”).2
Physical Bases of Dreams
But let me backtrack. One aim of neuroscience is to map the brain loci of thoughts and mental experiences. Everything we see, imagine, or think about is linked to neural responses somewhere in the brain. Dreams also have a home. We saw in chapter 6 that neural activity in the primary sensory areas of the neocortex produces the impression of sensory perception. This means that neurons firing in the primary visual cortex create the illusion of seeing things, neurons firing in the primary auditory area create the illusion of hearing things, and so forth. If that firing occurs at random, these perceptions can feel like crazy, randomly fragmented hallucinations. It is easy to imagine that the random imagery and sensations created in this way could be woven together to create a complex, multisensory hallucination which we might call a dream.
In 1977 two Harvard scientists, Allan Hobson and Bob McCarley, proposed a theory about how dreams occur called the activation-synthesis model.3 This draws on knowledge of sleep physiology to propose that dreams are generated in precisely the way described above. It turns out that chaotic firing of neurons in the brainstem is a core characteristic of REM sleep. Because these brainstem neurons communicate with the neocortex, their chaotic firing could trigger responses in the primary sensory and motor areas. The activation-synthesis model proposes that the brain may combine and synthesize these neocortical responses to create a story (Fig. 19). This explanation works well at the physiological level: We know the nightmarish dreams epileptic people sometimes have are caused by partial seizures which entail high-intensity, chaotic activity in the brain’s emotional system. In fact, electrical stimulation applied externally to the cortex can lead to dreamlike perceptions even during wakefulness.4 Both the data from epileptic patients and the electrical stimulation data show that chaotically triggered brainstem activity can lead to the subjective perception of a dream.

Fig. 19 The activation synthesis hypothesis of dreaming
Allan Hobson extended the activation-synthesis model by taking the pharmacology of sleep into account. He proposed that high levels of the neurotransmitter acetylcholine, in combination with the low levels of aminergic neurotransmitters such as norepinephrine which occur during REM sleep (see chapter 4), could cause a sense of bizarreness (e.g., unusual juxtapositions, distortions, illogical reasoning, and sudden scene changes). The absence of aminergic neurotransmitters interferes with the top-down control of the cortex by the brain areas which are involved in higher reasoning and normally force us to think logically. This idea is supported by the observation that the dorsolateral prefrontal cortex, one of the main centers of higher thought, is relatively inactive during REM sleep. In fact, Hobson drew a parallel between dreams and psychoses, since both entail hallucinations and illusions, as well as abnormal responses in the dorsolateral prefrontal cortex.
Unfortunately, Hobson and McCarley’s activation-synthesis model is less convincing when we think about the actual characteristics of dreams. For a start, dreams don’t only occur in REM sleep; they occur throughout the night in all stages of sleep. Perhaps more telling is the content of dreams. Studies have shown that many dreams are sensible, logical, and thematic, though potentially somewhat disconnected. Could chaotic neural activity really produce this pattern? And what about those recurrent nightmares that haunt us night after night? How could these be caused by semi-random brainstem responses? These combined characteristics of dreams (occurrence throughout the night, recurrence, and in some cases thematic logic) imply that they are generated in a structured way.
Not only are dreams not generated by chaotic brain activity, there is also strong evidence against the idea that primary sensory areas orchestrate dreams. Scientists often learn about how the body and the mind work by studying patients who are damaged or deficient in some way. When people suffer damage to the primary visual or sensory cortices, this causes no apparent deficit to vision or touch in dreams, even though it may cause either complete or partial blindness and inability to feel the things they actually do touch. A good example of this comes from people who have massive lesions to their primary sensory and motor cortices which can lead to paralysis of the arm, leg, and trunk on one side of the body (this is called hemiplegia). Amazingly enough, people with hemiplegia still enjoy normal dream imagery about bilateral movement in all of these body parts. Similarly, aphasic patients, who have great trouble talking due to extensive damage to the areas of the brain that control speech, have completely normal speech-related dreams. The experiences of these brain-damaged patients show us that the primary sensory and motor cortices are not responsible for generating imagery in dreams. Instead, dream imagery appears to be created in a more cognitively mediated fashion which involves the higher-level sensory and motor association cortices, which process sensory signals in a more refined way. For example, damage to the visual association cortex (which processes more complex visual information such as movement, orientation, color, size, and shape, and which normally only engages when you are paying attention to what your eyes are seeing) can lead to a complete loss of visual imagery in dreams.
With the great advantage of hindsight, some modern critics of the activation-synthesis model have suggested that, in its enthusiasm for the role of random activity in the brainstem in triggering the neocortical activity which underpins dreams, this model may have overinterpreted the reduced activity in the dorsolateral prefrontal cortex during REM sleep and not paid enough attention to other areas of the prefrontal cortex which are highly activated in REM sleep. Examples of such areas are the anterior cingulate, which is known to play an important control function in the brain (for instance, keeping the wayward amygdala in check in frightening situations); and the ventromedial prefrontal cortex, which is thought to be responsible for the sense of self. With these structures online in REM, it seems odd to assume that the dreams which occur during this sleep stage are generated in an entirely uncontrolled fashion.
A newer model of dreams was proposed by Mark Solms, then at the Royal London School of Medicine, in 2000. This model argues against Hobson and McCarley’s activation-synthesis ideas by suggesting that, far from being caused by chaotic activity in the brainstem and cortex, dreams are actually generated by the thinking part of the brain. Solms reviewed data from brain-damaged patients and discovered that people with lesions to the ventromedial prefrontal cortex seem to lose their ability to dream. His work suggests that the brain’s reward system, which originates in the midbrain and connects through the ventromedial prefrontal cortex as it projects upward to the rest of the brain, provides the basis for dreams. Lesions to the ventromedial prefrontal region dramatically disrupt this system. Chemical stimulation of the reward system (for instance, through the administration of L-dopa, a drug which can be converted to dopamine, the brain’s primary neurotransmitter of reward) causes both psychotic symptoms and excessive, unusually vivid dreaming. Blocking the action of dopamine using drugs—such as haloperidol, which clogs up receptors for this neurotransmitter—inhibits excessively vivid dreaming. Incidentally, haloperidol is often used to treat schizophrenia precisely because of its ability to inhibit psychoses.
Interestingly, the ventromedial prefrontal cortex, which Solms found to be so important for dreaming, is the same part of the prefrontal cortex that was often intentionally destroyed in order to restore the sanity of the patient in the heyday of invasive psychiatric procedures. It turns out that 70 to 90 percent of such prefrontal lobotomies also resulted in a complete loss of dreaming, which once more supports the parallel between dreaming and psychosis, as well as the importance of the reward system for dreaming.
In addition to the ventromedial prefrontal cortex, Solms found that lesions to the three-way junction between temporal cortex (which mediates sound processing and also general knowledge), parietal cortex (which mediates attention), and occipital cortex (which mediates vision) also abolished dreams. This region is important for mental imagery, so it isn’t surprising that lesions here disrupt dreaming.
Dreams Are Limited by
Cognitive Ability and State
Even among the healthy, not everyone is capable of normal dreams. The complexity, length, and thematic coherence of dreams all seem to be partially determined by the general cognitive capacity of the dreamer. For instance, there is evidence that autistic people have shorter dreams with fewer elements in them, and they recall less detail about the dreams they do have. This suggests a possible link between the ability to dream and a person’s general capacity for creative, imaginative, emotional thinking. In a similar vein, children younger than five years old tend to have unstructured dreams with poor narrative development; in the dreams of five- to eight-year-olds, these properties are predicted by measures of visuospatial intelligence.5 Interestingly, people with schizophrenia have brief dreams with limited hallucinatory content. These dreams contain a lot of aggression, and this is usually directed toward the dreamer. Particularly violent dreams can precede a psychotic scene in wake, and patients with very severe symptoms can have trouble distinguishing between wake and dreaming.6 The dreams of people with depression vary a lot. Some studies have shown that depressed people have short, bland dreams. Other observations have suggested that they have more negative and masochistic dreams. These differences are probably due to the fact that depression is a hugely diverse condition, and different studies probably included people with different subtypes of depression.
Do Dreams Serve a Purpose?
In contrast to the activation-synthesis model, which views dreams as epiphenomena—a simple by-product of neural processes in sleep—other scientists have suggested that dreams serve an important function. As usual in psychology, there are lots of different ideas about what this function could be. Sigmund Freud’s suggestion that dreams express forbidden desires is of course the most famous of these, but there are lots of other theories about what dreams might do, many with more empirical support than the Freudian view. For example, the threat simulation hypothesis suggests that dreams may provide a sort of virtual reality simulation in which we can rehearse threatening situations, even if we don’t remember the dreams. Presumably, this rehearsal would lead to better real-life responses, so the rehearsal is adaptive.7 Evidence supporting this comes from the large proportion of dreams which include a threatening situation (more than 70 percent in some studies) and the fact that this percentage is much higher than the incidence of threats in the dreamer’s actual daytime life. Furthermore, studies of children in two different areas of Palestine show that those who live in a more threatening environment also have a much higher incidence of threat in their dreams. Reactions to these threats are almost always relevant and sensible, so the rehearsal (if that’s what it is) clearly involves plausible solutions, again suggesting that they provide a kind of valid simulation of potential real-life scenarios.
Another suggestion is that dreams influence the way you feel the next day, either in terms of mood or more basic bodily states. Forcing people to remember the nastier dreams from their REM sleep definitely puts them in a foul mood, and nightmares (defined as very negative dreams which can wake you up) may even lead to ongoing mood problems. On the other hand, there is also evidence that dreams could help to regulate long-term mood. For instance, a study of dreams in divorced women showed that those who dreamed about their ex-husbands more often were better adapted to the divorce.8 Amazingly enough, dreams also seem able to influence physiological state: One study showed that people who were deprived of water before they slept, but then drank in their dreams, felt less thirsty when they woke up.9
The content of dreams can be influenced in lots of different ways. For instance, recent work has shown that sleepers tend to initiate pleasant dreams if nice smells are wafted at them in REM sleep, and they have negative or unhappy dreams if stinky, unpleasant smells are sent their way.10 Some people can achieve lucid dreaming, in which they control the sequence of events in their dream, and evidence suggests that these techniques can be learned by intensive practice and training. All of this is highly tantalizing, of course, because (though it tells us nothing at all about the original evolved purpose of dreams) it suggests we might not only be able to set ourselves up for pleasant experiences while we sleep, but we might also eventually be able to use these techniques to treat mood disorders, phobias, and other psychological problems. We already know that hypnotic suggestion can cause people to incorporate snakes, spiders, or other things about which they have phobias into their dreams, and—when combined with more benign forms of these menacing objects—such incorporation helps to remove the phobia. Hypnotic suggestion can also make dreams more pleasant, and mental imagery practiced during the day can be used to modify (and often nullify) persistent nightmares.
There is little evidence that people actually learn during their dreams. The fact that they can learn during sleep is a different matter, but dreams themselves don’t appear to be a good forum for imprinting new information into the hippocampus (after all, we don’t even remember our dreams most of the time). Studies of language learning illustrate this well. Although learning efficiency is predicted by an increase in the percentage of the night that is spent in REM, the dreams which are experienced during this extra REM don’t have much to do with language. If they relate to it at all they are most often about the frustration of not being able to understand something and not about the mechanics of how to construct or decode a sentence.
Memories in Dreams
What’s the most recent dream you can remember? Was anyone you know in it? Did it happen in a place you know well? Were you doing something familiar? Most dreams incorporate fragments of experiences from our waking lives. It’s common to dream about disconnected snippets like a particular person, place, or activity. But do dreams ever replay complete memories—for instance, the last time you saw your mother, including the place, activities, and people? Memories like this are called episodic because they represent whole episodes instead of just fragments; studies of dreaming show that these types of memories are sometimes replayed in sleep, but it is quite rare (around 2 percent of dreams contain such memories, according to one study).11 Most of our dreams just recombine fragments of waking life. These fragments are relatively familiar and reflect the interests and concerns of the dreamer. This means cyclists dream about cycling, teachers dream about teaching, and bankers dream about money.
Some researchers have capitalized upon dream reports to gain insight into the process by which memories are immediately incorporated (i.e., in the first night after they were initially experienced). Freud famously referred to this as “day-residues.” One study showed day residues appear in 65 to 70 percent of single dream reports.12 On the other hand, a more recently described phenomenon called the dream-lag effect refers to the extraordinary observation that, after its initial appearance as a day residue, the likelihood that a specific memory will be incorporated into dreams decreases steadily across the next few nights after the memory was formed, then increases again across the following few nights (Fig. 20).
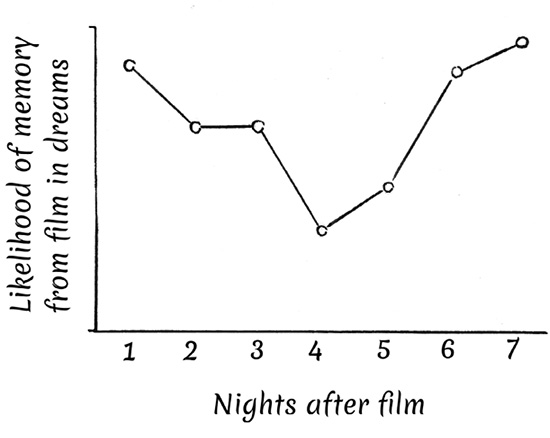
Fig. 20 The dream-lag effect in memory for a film
Thus, it is very common for memories to be incorporated into dreams on the first night after they were initially experienced (if I have a car crash today, I’m likely to dream about it tonight). The likelihood of such incorporation decreases gradually across the next few nights, with few memories incorporated into dreams three to five days after they occurred. Extraordinarily, however, the probability that a memory will be incorporated into a dream increases again on nights six and seven after it was initially experienced. What is going on here? Why are memories less likely to be incorporated into dreams three to five days after they originally occurred than six to seven days afterward? One possibility relates to the need for consolidation. Memories may be inaccessible at this stage because they are being processed in some way which takes them temporarily “offline.” Notably, this effect is only true for people who report vivid dreams, and it also appears to only be true of REM dreams. As with most research, the dream-lag effect raises more questions than it answers.
Why Do We Have Different Kinds of Dreams at Different Stages of the Night?
Dreams aren’t all the same. Everyone is aware of the difference between good and bad dreams, but we don’t tend to notice that some dreams are more logical and structured while others are more bizarre. Some dreams are so highly realistic that it is difficult to convince ourselves they aren’t real, while others are fuzzy and indistinct. Some dreams are fragmented, jumping rapidly from one topic to another, while others move forward in a more coherent story. Recent analyses have suggested that these differences are far from random; instead they may be driven by the physiology of various brain states and the extent to which structures like the hippocampus and neocortex are in communication during different sleep stages.
Dreams occur in all stages of sleep, but they seem to become increasingly fragmented as the night progresses. In general, they appear to be constructed out of a mishmash of prior experience. As mentioned above, dreams contain disconnected memory fragments: places we’ve been, faces we’ve seen, situations that are partly familiar. These fragments can either be pasted together in a semi-random mess or organized in a structured and realistic way. The dreams that occur in non-REM sleep tend to be shorter but more cohesive than REM dreams, and often they relate to things that just happened the day before. REM dreams that occur early in the night often also reflect recent waking experiences, but they are more fragmented than their non-REM counterparts. Conversely, REM dreams that occur late in the night are typically much more bizarre and disjointed.
Simply thinking about where these memory fragments are coming from and how they are connected together may provide an explanation for the difference between early and late-night dreams. The various elements of an episode are thought to be stored in the neocortex, but they are not necessarily linked together to form a complete representation. For example, if your memory of having dinner last night involves memories about a specific place, specific sounds, specific actions, and maybe even memories about other people who were there, each of these bits of information is represented by a different area of the neocortex. Even though they combine together to make up a complete memory, these various neocortical areas may not be directly interlinked. Instead, the hippocampus keeps track of such connections and forms the appropriate linkages, at least while the memory is relatively fresh. However, communication between the neocortex and hippocampus is disrupted during sleep, so this process is also disrupted. During REM sleep, both the hippocampus and those parts of the neocortex which are involved in a current dream are strongly active—but they don’t appear to be in communication. Instead, responses in the neocortex occur independently, without hippocampal input, so they must relate to memory fragments rather than linked multisensory representations. Essentially, when memories which have been stored in the neocortex are accessed or activated during REM, they remain fragmentary instead of drawing in other aspects of the same memory to form a complete episodic replay. These fragments aren’t linked together in the way they might be if you thought of the same place while you were awake (or indeed in non-REM sleep). For instance, cortical representations of both someone who was present for your dinner last night and of the place where it was held may be triggered, but these will not necessarily be linked together, and they may not be linked to the idea of dinner or eating at all. Instead, seemingly unrelated characters and events may be activated in conjunction with the memory of this place. One possible driver for this is the stress hormone cortisol, which increases steadily across the night. High cortisol concentrations can block communication between the hippocampus and neocortex, and since concentrations are much higher early in the morning, this could provide a physiological reason for the disjointed properties of late-night (early morning) dreams.
Irrespective of how it happens, it is clear that dreams not only replay memory fragments but also create brand-new, highly creative mixtures of memories and knowledge. This process has led to the creation of many works of literature, art, and science, such as Mary Shelley’s Frankenstein, the molecular formula of benzene, and the invention of the light bulb. An especially good demonstration of this somnolent creativity comes from a study of 35 professional musicians who not only heard more music in their dreams than your normal man-on-the-street but also reported that much of this (28 percent) was music they had never heard in waking life. They had created new music in their dreams!
Although we don’t quite understand how dreams achieve this type of innovative recombination of material, it seems clear that the sleeping brain is somehow freed of constraints and can thus create whole sequences of free associations. This is not only useful for creativity, it is also thought to facilitate insight and problem solving. It may even be critical for the integration of newly acquired memories with more remote ones (see chapter 8). In fact, this facilitated lateral thinking could, in itself, be the true purpose of dreams. It is certainly valuable enough to have evolved through natural selection.
Even if dreams serve a vital function, and even if they are important for memory consolidation, most still seem to fade from memory very soon after waking up. In fact, there is probably a physiological reason that we forget our dreams. When we are awake, the hippocampus monitors activity in the neocortex, keeping track of all the major neural events that happen there, binding together their elements and (most critically) remembering them. When we sleep, however, this pattern changes because the hippocampus becomes much less responsive to inputs from the neocortex. This means that the cortical activity isn’t being recorded in the same way as it would be during wake, so the episodes, themes, and stories created in our dreams are not captured. Thus, the same physiological disconnect which makes us construct bizarre combinations and sequences of events when we dream also tends to keep us from remembering them later on when we are awake. If you think about it, this disconnect is probably adaptive since it could be confusing if we remembered our dreams so well that we mixed them up with reality.
Of course, this intentional amnesia for dreams doesn’t always work. If it did you wouldn’t be reading a chapter about dreams because we wouldn’t know they existed. It is unclear why dream amnesia is only partial, but this incomplete pattern suggests that communication between the hippocampus and neocortex is not entirely cut off during sleep—it is just slowed or downgraded (think about a slow Internet connection), so some of the stronger stimuli occurring in the neocortex will still eventually make it into the hippocampus and be remembered. Reduced connectivity of this type could also explain why the dreams we remember are often emotional: Emotional stimuli are stronger, and so perhaps this helps them to penetrate even when there is limited connectivity.
Dreams as Memory Replay?
Another possible mechanism for the preferential remembering of emotional dreams relates to memory replay. The idea that dreams may be a conscious manifestation of the same spontaneous replaying of memories which underpins consolidation during sleep is just too tantalizing to neglect. Although it is unlikely that every single memory replay could surface in this manner, it is possible that a small percentage of replays (the tip of the iceberg) do. Researchers have tested this idea by checking for a relationship between the dreams people report having and the memories which improve during that same epoch of sleep. Specifically, they have looked to see whether people who report dreaming about a newly learned task (like exploring a maze, for instance) improve more across sleep than people who make no such reports.13 Although it is still relatively early days for this idea, the existing data provide fairly consistent support. One problem with these studies is that because people forget their dreams, researchers have to test vast numbers in order to get a decent sample of “rememberers.” The latest wave of experiments have tried to get around this by making the to-be-remembered tasks as emotive as possible. For example, instead of running around fairly mundane 3D mazes on a computer as in older studies, people participating in these experiments are immersed in interactive video game scenarios in which they are racing a clock for survival, and bogeymen may jump out at any corner of the maze while the participants’ potential monetary reward visibly dwindles in one corner of the computer screen. This has led to a greater degree of dream incorporation (and memory), but the jury is still out on how these relate to subsequent memory.
Summing Up
This chapter has introduced dreams as physical responses which are linked to specific patterns of brain activity. We mentioned the idea that dreams are epiphenomena that result from the random neural firing in the cortex triggered by disordered activity in the brainstem, as well as the idea that dreams are generated by more thinking parts of the brain such as the anterior cingulate. We saw that lesions to the ventromedial prefrontal cortex abolish dreaming, possibly because the dopaminergic reward system which projects through this region is essential for generating dreams. We also explored some potential functions of dreaming, touching on threat simulation and mood regulation, as well as memory replay. Focusing more specifically on memory, we saw that there is a period several days after learning when memories are less likely to be incorporated in dreams, and this is thought to reflect a gradual consolidation process which renders memories temporarily unavailable. Finally, we reviewed the possibility that dreams are important for memory consolidation because they reflect memory replay, and we saw that while memories are sometimes replayed in a more complete form early in the night, only fragments are replayed later in the night, perhaps reflecting the limited connectivity between the hippocampus and neocortex at this time.
The next chapter will take a different angle on consolidation by considering how sleep helps us to abstract overall principles or “gist” from a corpus of information, thus facilitating the construction of general knowledge, as well as creative connections and inferences.