Pioneering Spanish cell biologist Santiago Ramón y Cajal made these remarkable drawings, of interconnected neurons (nerve cells) from the brain of a rabbit, in 1899, just over three decades before the invention of the electron microscope.
A Brief History of the Cell
The earliest observations of cells were made in the late seventeenth century, but their fundamental importance in the natural world only became apparent over 150 years later, in the middle of the nineteenth century. Since then, increasingly rapid strides have been taken toward understanding what goes on inside cells—and how such processes relate to growth, reproduction, inheritance, disease, and the origin of life on Earth.
Pioneering Spanish cell biologist Santiago Ramón y Cajal made these remarkable drawings, of interconnected neurons (nerve cells) from the brain of a rabbit, in 1899, just over three decades before the invention of the electron microscope.
When seventeenth-century natural philosophers and physicians gazed through microscopes at plants, animals and fungi they were treated to tantalizing glimpses of anatomy and physiology on tiny scales. Microscopes allowed these scientists and doctors to discover “microorganisms”—entire living things too small to see with the naked eye—and to stumble across the existence of cells.
A revolution in seeing
The facts surrounding the invention of the microscope are about as clear as the images that early examples of these instruments produced. It was in the 1590s, or possibly the early 1600s, and probably in Holland, but possibly in England, that someone first placed two lenses in an arrangement that produced a magnified image. What is known is that the new instrument, more powerful than the hand lenses already in use, quickly captured the imagination of natural philosophers across Europe.
The magnifying power and optical quality of microscopes improved gradually during the seventeenth century. Although minerals and everyday objects were frequent subjects of study, it was closeup views of living things that really caught people’s eyes. In 1660, the Italian physician Marcello Malpighi carried out microscopic studies of human flesh and found tiny blood vessels—the capillaries, which join arteries to veins. The discovery of capillaries confirmed a controversial theory: the circulation of blood, put forward by William Harvey in 1628. Malpighi studied many plants and animals with his microscopes, and in 1666, after studying a blood clot, he described “very small red particles” that “roll and turn helter-skelter”, the first confirmed sighting of what we now call red blood cells.
Robert Hooke’s drawing of “cells” in cork. What he actually observed were the spaces enclosed by cell walls of empty, dead cells. Note that “B” is, as Hooke described it, “split the long-ways.”
Tiny boxes
The most influential microscopist of the age was Englishman Robert Hooke. While employed as “curator of experiments” at the new Royal Society in London, Hooke made many observations through microscopes and telescopes, and produced a beautifully illustrated book of what he had seen. Micrographia was published in September 1665 and its exquisite drawings and intriguing text gave readers an insight into a world hidden from everyday eyes. The now famous diarist Samuel Pepys was among those captivated, noting: “Before I went to bed, I sat up till 2 a-clock in my chamber, reading of Mr. Hooke’s Microscopical Observations, the most ingenious book that I ever read in my life.”
It was Hooke who coined the word “cell” to describe what he saw when studying cork. He placed thin slivers of the material onto a dark plate beneath his microscope’s objective lens, illuminated them with light from an oil lamp focused through a thick lens, and gazed through the eyepiece. His description of what he saw, quoted below, is still intriguing.
Hooke estimated that there were about 10,000 cells to the inch (about 4,000 per centimeter) and that one cubic inch would contain about “twelve hundred millions” (about 70 million per cubic centimeter). It was an astonishing discovery; he wrote that this intricate structure was “almost incredible, did not our Microscope assure us of it by ocular demonstration.” Each of Hooke’s “cells” is a cube with sides measuring just over 20 microns, or 0.02 millimeters.
“I could exceeding plainly perceive it to be all perforated and porous, much like a Honey-comb, but that the pores of it were not regular … these pores, or cells, were not very deep, but consisted of a great many little Boxes, separated out of one continued long pore, by certain Diaphragms, as is visible by the Figure B, which represents a sight of those pores split the long-ways.”
Reconstruction of Robert Hooke’s compound microscope, and its system of illumination, copied from the engraving and description in Micrographia.
Although compound microscopes (with two or more lenses) were popular during the seventeenth century, many investigators also used “simple microscopes”—just single powerful lenses. Some of these could magnify as well, if not better, than their more complicated counterparts. One man who favored single lenses was Antony van Leeuwenhoek, a successful Dutch draper. Leeuwenhoek made tiny, near-spherical lenses by melting glass rods in a flame. He carefully ground them to the right shape and attached them to ingenious handheld metal frames that also held the specimen. He investigated everything from tongues to sand and became the first person to describe sperm cells (which he found in the males of several species, including humans). While most microscopes of the time achieved magnifications of between 30x and 60x, Leeuwenhoek’s could magnify more than 250x.
In 1675, Leeuwenhoek observed tiny living creatures in a sample of rainwater that had been standing for a few days. These microorganisms were far, far smaller than any living things anyone had ever seen. Leeuwenhoek called them “animalcules.” For the next year he studied river water, well water, and seawater, some of which he left standing for several days or weeks. Mostly, he saw protozoa and single-celled algae, which are about the same size as Hooke’s cork cells—some quite a lot larger. But in April 1676 he saw animalcules that were much smaller, and these he described as being so tiny that you would need to lay more than a hundred end to end to measure the same as a grain of sand. This was almost certainly the first observation of bacteria.
A letter that Leeuwenhoek wrote in 1683 contains the world’s first illustrations of bacteria. The letter detailed his microscopic investigations of his own dental plaque: “I have mixed it with clean rain water, in which there were no animalcules, and I saw with great wonder that there were many very little animalcules, very prettily a-moving.” Leeuwenhoek also wrote that “there are not living in our United Netherlands so many people as I carry living animals in my mouth this very day.”
Leeuwenhoek (see here) was prolific; he made more than 500 microscopes and wrote hundreds of letters informing scientific societies about his discoveries—including 190 or so to the Royal Society in London. The drawings (see here) are taken from one of his letters. Leeuwenhoek’s microscope (see here) was a handheld metal frame with screws to adjust the stage (sample holder) and to move the lens for focusing.
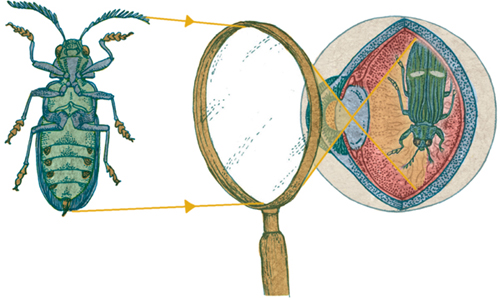
2 WITH MAGNIFYING GLASS
3 OPTICAL MICROSCOPE
HOW OPTICAL MICROSCOPES WORK
Light passing through or bouncing off any point of an object spreads out in all directions, in straight lines, or rays. Any rays that pass through the eye’s lens form an image on the retina (1). All the rays from any particular point of the object always reach the corresponding point of the image, thanks to the focusing ability of the eye’s lens. It is possible to illustrate the extent of the image produced on the retina by choosing just two points—one at the top and one at the bottom of the object—and just one ray from each point. The two rays chosen here pass through the center of the lens, without bending. The apparent size of an object is determined by how much of the retina the image takes up, which in turn is determined by the angle at which these two rays enter the eye: the visual angle. Lenses and microscopes change the visual angle, by bending light—and by doing so, they can enlarge the retinal image, making an object appear much bigger (2).
A basic compound light or “optical” microscope (3) consists of a light source, a stage on which to place the specimen and two lenses (or sets of lenses) called the objective lens and the eyepiece. The objective lens, the one closest to the specimen, produces a magnified “real image” of the specimen inside the microscope tube. This simply means that if a piece of paper were placed there, the image would be projected. The eyepiece acts as a magnifying glass, enlarging that first image to produce a very high magnification overall. The total magnification is the magnification of the objective lens multiplied by the magnification of the lens or lenses in the eyepiece.
Surprisingly, perhaps, the idea that living things are made of cells did not come from the observations of the first microscopists, such as Hooke, Leeuwenhoek, and Malpighi. Instead, it originated as a philosophical thought borrowed from physical science.
Living molecules
In his influential book Philosophiæ Naturalis Principia Mathematica, published in 1687, the English scientist Isaac Newton popularized the notion that matter and even light might be made of tiny, indestructible particles. This idea had a long pedigree, not least because the alternative is continuous matter, which is difficult to understand. Many naturalists wondered whether living things might be made of particles of a different kind; because they believed living things are fundamentally different from nonliving matter, the particles themselves would have to be alive. In 1749, French naturalist Georges-Louis Leclerc, the Comte de Buffon, called them “living molecules.”
At the same time, biologists were busy familiarizing themselves with the microscopic anatomy, or histology, of plants and animals. Some wrote about their observations of “cellular tissue” and even began to make the connection between their observations and the idea of living molecules. However, many of the “cells” were optical illusions caused by dirty lenses or out-of-focus microscopes—and, in plants at least, the word “cellular” often meant “populated by empty spaces.” By the early nineteenth century, however, scientists had begun focusing their ideas—and their microscopes—more acutely.
Illustrations by Matthias Schleiden, detailing his observations of plant cells and their nuclei (cytoblasts), from an 1847 book by Theodor Schwann.
Coming together
In the 1820s French botanist Henri Dutrochet boiled plant tissue in nitric acid to dissolve away the material that held the cells together. He watched as the cells separated into countless individual, self-contained “globules,” concluding that cells make up “the fruits, the stems, roots, leaves and flowers on all the plants on the surface of the planet.” The German botanist Franz Meyen reached a similar conclusion in 1830, observing: “Plant cells occur either singly, so that each forms an individual, as in the case of some algae … or they are united together to form greater or smaller masses, to constitute a more highly organized plant.”
Some botanists observed a dark spot inside certain plant cells. In 1831 British botanist Robert Brown gave it a name: the “nucleus.” In the 1830s, another German botanist, Matthias Schleiden, suggested that nuclei were the source of new cells after he supposedly saw new cells forming around nuclei and then emerging from inside the cell. In 1837, Schleiden described what he had seen to his colleague, the zoologist Theodor Schwann, who recognized the description of the dark spots as something he had seen in the cells of tadpoles. Schwann was convinced that in animals, too, nuclei seemed to give rise to new cells.
Two out of three
Schwann thought he also saw nuclei in the spaces between cells. He suggested that all nuclei crystallize from a hypothetical substance which he called “cytoblastema,” believing it to be present inside and outside cells. In 1839, he established the first coherent cell theory, based on three principles: first, that every part of every living thing is either made of cells or made by cells; second, that the life of a living organism as a whole is due to the fact that its cells are alive; third, that cells come into existence in or near other cells, from cytoblastema. The last idea was quickly gunned down by biologists who had observed cells dividing in two (binary fission) and realized that they were reproducing.
French biologist Barthélemy Dumortier had watched binary fission as early as 1832, even writing that it provided a “perfectly clear explanation of the origin and development of cells,” but his observation had remained controversial. Robert Remak, a Polish-German embryologist, carried out painstaking studies of developing embryos in the 1840s and noticed that every new cell arose from the division of preexisting cells.
In 1855, German biologist Rudolf Virchow made Remak’s observations his own and established and publicized modern cell theory. He dropped Schwann’s idea about cytoblastema, replacing it with a Latin phrase: omnis cellula e cellula (“every cell comes from a cell”). This simple idea is crucial to understanding how living things grow and reproduce and, ultimately, for making sense of the process of evolution by which species rise and fall over time.
Cell theory, combined with improvements in microscopy, made sense of the living world—in particular growth, reproduction and inheritance. Asking ever deeper questions, cell biologists turned to the chemistry of living things: biochemistry.
Seeing more clearly
Theodor Schwann’s idea about cell nuclei crystallizing from cytoblastema is not as absurd as it might seem. The nucleus becomes far less prominent when a cell is about to divide, so it really can seem to appear and disappear. It took two technological advances to show that the nucleus is present all the time and to begin working out its role.
The first of these advances was better microscopes. In the 1830s, British physicist Joseph Jackson Lister introduced microscopes with lenses that corrected for spherical aberration (distortion of the image) and chromatic aberration (annoying colored fringes around the image). Ernst Abbe, a German physicist, pushed optical microscope design to its limits in the 1870s, immersing lenses in oil to maximize magnification, resolution, brightness, and contrast.
The second important advance was histological staining—the use of dyes that are absorbed only by certain structures in the cell to make those structures stand out. Working with brain cells in 1858 German anatomist Joseph von Gerlach noticed how carmine (cochineal) was taken up by the nucleus and its contents but not by the rest of the cell. The introduction of a range of synthetic dyes in the 1860s opened up the technique, leading to the discovery of several other organelles (see here).
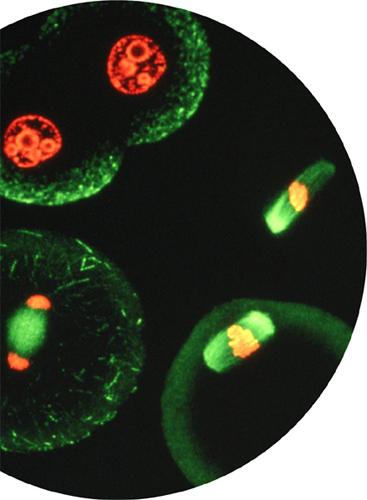
Micrograph showing the dramatic changes that take place during cell division. This modern image uses fluorescent stains to highlight the nuclear material (red) and the protein components of the cell (green).
Identical twin daughters
The nucleus does not disappear during cell division. Instead, it splits into pieces, which are shared out between the two resulting “daughter” cells. In 1882 German biologist Walther Flemming named the material from which the fragments are made chromatin (because it readily absorbed the colored stain he was using). Working with cells from salamanders, Flemming also noticed that during cell division, the chromatin becomes arranged into distinct strands, later named chromosomes; these are pulled apart by tiny, barely visible ropes to form the nuclei of the two daughter cells. The perfectly coordinated dance of tiny colored strands inside a cell nucleus was (and still is) a wonder to behold. He called it mitosis, a term still used today. An easily overlooked point—but one that reveals a profound unity throughout nature—is that soon after Flemming observed mitosis in salamander cells, others soon saw the same dance happening in other animals, plant cells, and fungi (bacterial cells do not have nuclei and reproduce differently, as explored in chapter four).
Biologists studying mitosis during cell division noticed that each of the daughter cells ended up not with just a random half of the chromatin material but identical sets of chromosomes. Quite how the cell manages to do that would remain a mystery for decades to come. However, it suggested that the chromosomes carry information essential to the proper functioning of the cell, or perhaps even the whole organism—something like a set of instructions.
Modern views of two organelles discovered in the 19th century. Far left: A colored transmission electron micrograph of a Golgi apparatus. Left: A phase contrast light micrograph of chloroplasts (the green blobs) in plant cells.
After it became clear that there are a number of structures inside cells akin to the organs in the human body, German zoologist Karl Möbius suggested the name “organulas” (little organs). The term for them later became “organelles.” The first organelle to be discovered was the nucleus.
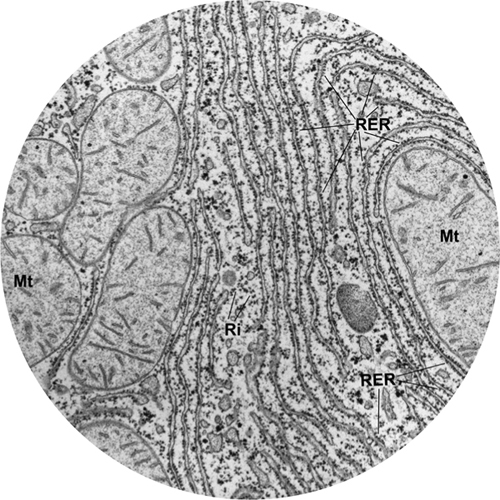
Determined to find out how plant cells manufacture starch, German botanist Julius von Sachs discovered the chloroplast (a structure essential in photosynthesis) in the 1860s—although it wasn’t named until the 1880s. Several biologists had reported seeing granules in muscle cells from the 1840s onward; they were identified in other types of cell in the 1890s, and called mitochondria. The “Golgi apparatus” was discovered by Italian physician Camillo Golgi during an investigation of the nervous system in 1897, although no one understood its function for decades to come. Another large, if inconspicuous, organelle is the endoplasmic reticulum. It was discovered in 1902 by Italian pathologist Emilio Veratti.
Early observations of meiosis during the production of sperm cells in the worm. Drawings in The Text-book of Embryology by Fredrick Bailey and Adam Miller, first published in 1909.
Gregor Mendel, the Austrian monk who founded the science of genetics with his plant-breeding experiments in the 1860s. The importance of his findings went unrecognized for over three decades.
Sex cells
The nature of the information carried by chromosomes became a little clearer in the last few years of the nineteenth century. Biologists noticed that the nuclei of egg and sperm cells (sex cells, or gametes) have half as many chromosomes as cells in ordinary tissue (somatic cells). The process by which the set of chromosomes in a nucleus is reduced by half is another remarkable and intricate chemical dance that came to be called meiosis.
During fertilization, the two nuclei fuse together to form a complete, and new, set of chromosomes, different from the chromosome set of either the male or female parent. In that way, both parents contribute equally to the chromosome set of the new individual created by fertilization. It was clear that the chromosomes have something to do with heredity—passing characteristics from generation to generation. They provide instructions for cell housekeeping and a set of instructions on how to build a unique organism, both contained in one tiny part of each tiny cell.
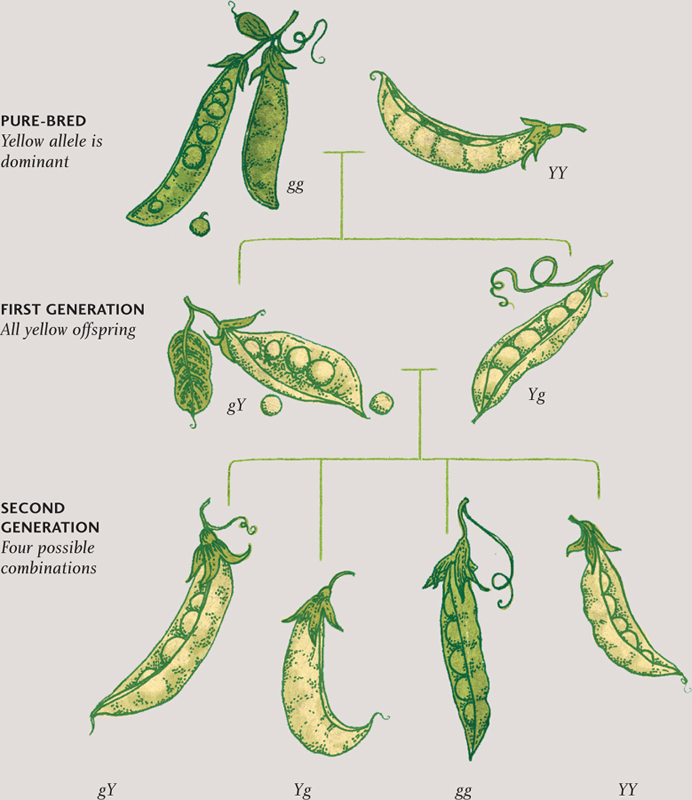
In the first few years of the twentieth century, biologists studying heredity rediscovered the work of an Austrian monk, Gregor Mendel. In the 1860s, Mendel had conducted experiments on pea plants, trying to work out the rules of inheritance. He established that each physical trait is carried by factors that come in pairs, one from each parent. Where an individual inherits two different forms of a particular factor, one form is usually dominant. So, for example, if a purebred pea plant with yellow seeds is crossed with a purebred plant with green seeds, the offspring will all have yellow seeds. But cross two plants from that generation, and some will have green seeds, because they have retained the nondominant (or recessive) factor and passed it on to their offspring. Only if an individual plant inherits two copies of the recessive green-seed factor will it have green seeds.
German biologist Theodor Boveri and American biologist Walter Sutton realized that Mendel’s factors are somehow carried by the chromosomes—one from each parent—and that Mendel’s laws of inheritance are applied inside the cell nucleus. In 1909, Danish botanist Wilhelm Johannsen came up with a name for this bold and exciting new science: genetics. Mendel’s “factors,” carried on chromosomes, became genes. In a laboratory in New York, a team of scientists led by geneticist Thomas Hunt Morgan began the process of mapping certain physical characteristics to certain chromosomes, and even to certain regions of particular chromosomes.
MENDELIAN INHERITANCE
Science is a never-ending journey toward the truth; any answer to one question opens up a deeper layer of questions. In this case, a new line of investigation was: “What is chromatin, and how does it carry information?” The answer would begin with chemistry—biochemistry—and would end up spawning a whole new discipline: molecular biology.
Breaking it down
Because a cell is, in essence, nothing more than a bag of chemicals, it is vital for anyone trying to understand the cell to determine its contents. In the nineteenth century, biochemists had worked out that chloroplasts in plant cells manufacture glucose and starch. These compounds are carbohydrates that are used as a source of energy; they are also built up into larger molecules of cellulose (the main structural material of plants). The scientists also knew that cells contain fats—in the cell’s outer envelope, or membrane, and inside the cell, too. And they were aware that cells manufacture proteins, which are the main component of most biological materials. Hair and nails are made almost exclusively of proteins, for example, and skin owes its toughness and elasticity to two proteins, collagen and elastin.
In the 1870s biologists also began discovering enzymes—catalysts that encourage chemical reactions to take place. The fermentation of sugar, for example, does not happen spontaneously, but it does proceed in the presence of an enzyme called zymase. The nature of enzymes was a mystery until the painstaking experiments of American chemist James Sumner in the 1920s proved that enzymes are, in fact, proteins. So ubiquitous are proteins in the cell that many biologists believed that proteins had to be the carriers of heredity. But they were wrong.
During the twentieth century the mysteries of reproduction and inheritance finally succumbed to science’s relentless quest for understanding. Biochemistry and molecular biology have laid bare the machinery of life at the smallest scale. Meanwhile, electron microscopes have given cell biologists access to ever clearer views of cells’ components, and a range of new techniques for looking inside living cells has provided intimate knowledge of those components at work.
Nuclear chemistry
While most biologists supposed that proteins must be the carriers of genetic information, another candidate was waiting in the wings. In 1869, Friedrich Miescher, a Swiss biochemist, isolated a mysterious compound from inside the nuclei of white blood cells. Because he discovered it in the nucleus, he called it “nuclein.” In the 1870s, a German biochemist, Albrecht Kossel, showed that nuclein is made of proteins plus another compound. That other component soon gained the name “nucleic acid” (because it was found to contain phosphate, which is acidic when dissolved in water). In addition to phosphates, nucleic acids were also found to contain sugars. By the turn of the century, Kossel had successfully established that nucleic acids also harbor five compounds: adenine (A), cytosine (C), guanine (G), thymine (T), and uracil (U). These compounds are bases (they form alkaline solutions in water) and so became known as nucleobases.
Biochemists had identified two different types of nucleic acid. They initially called these yeast nucleic acid and pancreas nucleic acid, because of where they were discovered. During the 1910s and 1920s, the Lithuanian-American biochemist Phoebus Levene identified the nature of the sugar molecules present in nucleic acids. Yeast nucleic acid contains a type of sugar called ribose, and so gained the name ribonucleic acid (RNA). Pancreas nucleic acid contains a very similar sugar, called deoxyribose; it gained the name deoxyribonucleic acid (DNA). Levene also worked out that RNA contains only the nucleobases C, G, A, and U, whereas DNA contains only C, G, A, and T, and he came up with a largely unconvincing way in which nucleic acids might be the carriers of hereditary information.
Transforming opinion
Levene’s idea was wrong, but he was looking in the right place. Working out the structure of nucleic acids—how the phosphates, sugars, and nucleobases fit together—would ultimately unravel the secrets of how cells make proteins and how chromosomes carry genetic information. Despite the interest in DNA and RNA, most molecular biologists of the 1920s and 1930s clung to the idea that inheritance was carried by proteins.
That view changed after scientists studying bacteria succeeded in transforming one bacterial species into another by mixing dead bacteria of one pathogenic (disease-causing) species with live bacteria of a benign (meaning harmless) one. Something had transferred from the virulent cells to the harmless ones—something species-defining. A British bacteriologist, Frederick Griffiths, was first to do this, in 1928. But it took until 1944 for molecular biologists to establish that what Griffiths had called the “transforming principle” was in fact the nucleic acid DNA.
In what proved to be one of the most important and well-known breakthroughs in the history of science, in 953 scientists in Cambridge and London, England, worked out the structure of the DNA molecule, and thus gave rise to modern molecular biology.
Portrait of Friedrich Miescher, who discovered nuclein, the compound containing DNA, in pus obtained from a hospital adjacent to his laboratory.
Rosalind Franklin, whose work in X-ray crystallography helped determine the structure of DNA. She discovered the double helix independently of Watson and Crick.
“Photo 51” (taken by Franklin’s Ph.D student Raymond Gosling). This image was crucial in determining the positions of the bases, sugars, and phosphates in DNA.
James Watson (seated) and Francis Crick with their model of the double-helix structure of DNA. Metal plates and rods represent nucleobases, sugars, and phosphate groups, and are held in the right place by laboratory clamps and a retort stand.
Doubling up
In London, two molecular biologists, Rosalind Franklin and Maurice Wilkins, used a technique called X-ray crystallography to work out the relative positions of the atoms present in DNA. In Cambridge, James Watson and Francis Crick used the X-ray crystallography data to build a model of the DNA molecule made of hundreds of metal plates that represented the sugars, phosphates, and nucleobases. Watson and Crick’s model revealed the fact that DNA must have a double-helix structure—something like a twisted rope ladder.
The nucleobases form the ladder’s “rungs”; they are strung along two sugar-phosphate backbones that form the ropes, and they join together in the center, preventing the two ropes from coming apart. It is the sequence of nucleobases along the backbones that carries genetic information—forming the letters of the code of life. The letters spell out recipes for creating proteins. In general, each recipe, encoded in a finite sequence of nucleobases, represents a single gene.
By the mid-1960s, molecular biologists had worked out the code by which sequences of As, Cs, Ts, and Gs of the nucleobases represent the instructions to make proteins – and the role of RNA in the whole process. When a protein is to be made inside a cell, the two ‘ropes’ of the DNA double-helix ladder unzip, and a strip of RNA is made as a copy of the code for the relevant gene. The piece of RNA is used as a template for building the protein for which the gene codes. Yes, it’s another intricate, well-choreographed molecular dance—but, unlike mitosis and meiosis, it’s going on every second of every day, not just when a cell divides. The actual protein-building part is carried out by molecular machines called ribosomes, which were discovered in 1954 by Romanian-American cell biologist George Palade. We will explore clearly and in more detail how this remarkable scheme works in chapter two.
A researcher removes DNA (in solution) from a plastic micro test tube using a precise micropipette, ready for sequencing.
The result of DNA sequencing—a small section of a genome with each of the four nucleobases represented by a different color.
The entire sequence of nucleobases—across all the DNA in all the chromosomes inside the nucleus of an organism’s cells—is known as that organism’s genome. From the 1970s onward molecular biologists began developing techniques for sequencing DNA—reading the sequence of nucleobases. Soon they were sequencing entire chromosomes and eventually entire genomes. The first genome to be sequenced, in 1995, belongs to a bacterium. A draft version of the three billion-nucleobase-long human genome was completed in 2000, and the Human Genome Project was declared complete in 2003. Of course, everyone’s genome is unique, so the project will never really be complete.
Molecular biology, together with genetics, have made remarkable strides in advancing our understanding of what makes living things alive—and how cells manufacture proteins and how information is passed from parents to offspring. Because every cell comes from another cell, and DNA may be slightly altered as it passes from generation to generation, molecular biology is important not only in making sense of living things today but also in answering questions about the history of life on Earth—evolution—and even its origins more than three billion years ago.
Getting a reaction
There is more to biochemistry than the nucleic acids that carry genetic information – and there is also more to biochemistry than knowing what chemical compounds are present in a cell. It is also important to work out how fats, carbohydrates, and proteins interact with each other and how they are broken down and built up. The set of chemical reactions that keep living things alive is called metabolism.
Two of the most important metabolic reactions are photosynthesis and cellular respiration—although they are actually pathways consisting of many separate, successive reactions. These two reactions are essentially the reverse of one another. It is through photosynthesis that the cells of plants and certain other organisms capture energy from sunlight, storing it in the form of carbohydrates. Cellular respiration releases that energy.
THE ORIGIN OF LIFE
No one knows exactly when or how the first cells came to be, but some of the key molecules of life occur naturally in a number of lifeless locations. For example, nucleobases of extraterrestrial origin have been found in meteorites, and amino acids, the building blocks of proteins, have been detected in comets. Another possibility is that the first self-replicating organic molecules were manufactured in the tumultuous conditions of hydrothermal vents—cracks in Earth’s crust at the bottom of the ocean. Either way, the world’s first cells would probably have been simple oily blobs that contained tiny chemistry laboratories, protecting early self-replicating RNA and DNA from an otherwise harsh environment.
The building blocks of proteins—amino acids—have been detected in outer space, and these crucial ingredients for life may have been delivered to Earth by comets crashing into our planet during its formative years.
Finding the energy
Working out the many complex compounds and reactions involved in photosynthesis and cellular respiration were mammoth tasks, achieved in the 1930s and 1940s. Both reactions involve a remarkable compound called adenosine triphosphate (ATP), which acts as the cell’s energy currency. In photosynthesis, ATP is one of the end products; in cellular respiration, it is both one of the initial reactants and one of the end products. German biochemist Karl Lohmann discovered ATP in 1929, but the manner in which it stores and releases energy was not uncovered until the 1940s.
ATP is built up and broken down by an enzyme called ATP synthase (enzymes always end with the suffix “-ase”). This extraordinary molecule is the world’s smallest motor: one part of the molecule literally rotates continuously inside the other part. It took decades for biochemists to work out the structure and behavior of this molecule. Every one of the 50 trillion or so of your living cells contains thousands of ATP synthase molecules embedded into the membranes of mitochondria. Each one rotates about twenty times every second, building three molecules of ATP per rotation, and keeping you alive.
Despite the improvements in microscope technology in the nineteenth century, some features of the cell remained unseen. This is because of theoretical limits to the magnification and resolution possible with optical microscopes. Light waves change direction, or diffract, as they pass around edges or small objects—in the way that waves in water change direction as they pass around the entrance to a harbor. Diffracted light waves from two edges or two objects close together interfere (just as the spreading waves from two adjacent openings in a harbor wall would create choppy water). This interference compromises the quality of the image you can achieve, to the point where you can discern no detail whatsoever. However good the microscope, it is thought to be impossible to overcome this interference. Ernst Abbe, the German physicist who improved microscope design in the 1870s, worked through the math and found that optical microscopes would never be able to resolve two objects separated by less than half the wavelength of light – about 200 nanometers (0.0002 millimeters). Several significant organelles and other features inside the cell are a lot smaller than that.
In 1931, two Germans, the physicist Ernst Ruska and the engineer Max Knoll, invented a new kind of microscope, which uses electrons instead of light to produce images. In 1933, an electron microscope exceeded the theoretical magnification of optical microscopes. By the late 1930s, electron microscopes could resolve objects 10 nanometers apart—by the mid-1940s, 2 nanometers. One of the first triumphs of the electron microscope in cell biology was the discovery of ribosomes—the molecular machines that build proteins.
HOW ELECTRON MICROSCOPES WORK
The discoveries of quantum physics in the first 30 years of the twentieth century led to the inescapable and bizarre conclusion that electrons, discovered in 1898 as tiny charged particles, are, in fact, waves as much as they are particles. Similarly, light is a stream of particles, called photons, as much as it is a wave. In each case, the beams interact with objects, because they either bounce off or pass through, and if you have a lens, you can form an image of those objects (the lenses in electron microscopes are electromagnets, not glass). Because the wavelength of electrons is so very much smaller than the wavelength of light, an electron microscope can resolve objects and other details of a much smaller size than an optical microscope can.
In a transmission electron microscope (TEM), electrons pass through a slice of the specimen. In a scanning electron microscope (SEM), the specimen is given a very thin coating of metal to reflect the electrons.
New vision
Cell biologists now have at their disposal a variety of innovative techniques to use in conjunction with optical and electron microscopes. These provide scientists with unprecedented views of the inner life of the cell. Dark-field microscopy, for example, invented in 1909, is a way of making everything except the specimen appear black. Phase contrast microscopy, invented in the 1930s, provides a way of picking out details in otherwise translucent organelles without the use of staining.
Perhaps the most exciting recent innovation is the development of fluorescence microscopy. Sometimes this takes advantage of a specimen’s natural fluorescence—for example, particular proteins produce visible light when illuminated with ultraviolet radiation (UV). Alternatively, a sample can be stained with fluorescent dyes. Most exciting is when an organism’s DNA is deliberately altered to add a sequence that codes for a fluorescent protein. When that part of DNA is active, the protein is also made, and the activity can be followed in a living cell in real time. The fluorescent protein most frequently used is green fluorescent protein, discovered in jellyfish in the 1960s and incorporated into a wide range of plants and animals by genetic engineering since the 1990s.
Throughout this book are remarkable images produced by optical microscopes and electron microscopes, using phase contrast, dark-field illumination, and fluorescence. Robert Hooke and Antony van Leeuwenhoek would have been amazed.
1. Transmission electron micrograph showing mitochondria and endoplasmic reticulum inside a cell. Like all electron micrographs, there is no color.
2. Scanning electron micrograph parasitic bacteria living inside a larger cell. “False color” has been added to differentiate the different elements of the image.
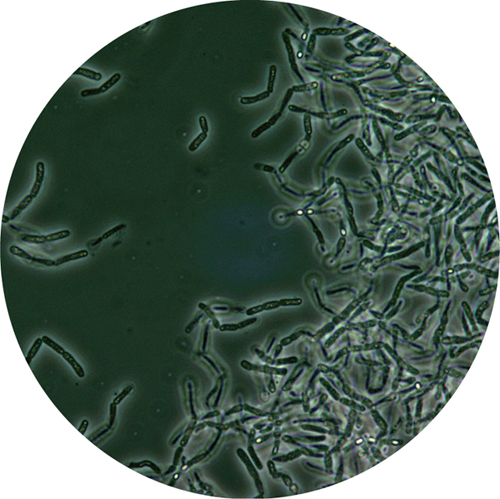
3. Phase contrast light micrograph showing the bacterium Bacillus anthracis. Dry spores inside some of the bacterial cells show up as bright dots, because they refract the light differently than the rest of the cell.
4. Dark-field micrograph of “stentor” protozoa. Due to a clever optical arrangement, all light is excluded from the image except light that has been scattered by the object. That explains why the background is dark.
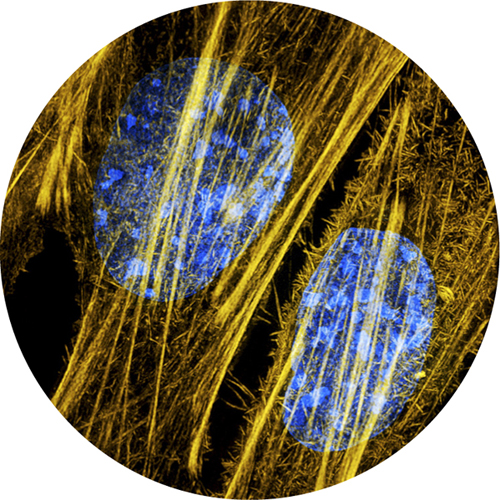
5. Very high magnification fluorescent micrograph of DNA (blue) and protein cytoskeleton filaments (yellow) inside a cell. The DNA and proteins are stained with different fluorescent dyes.