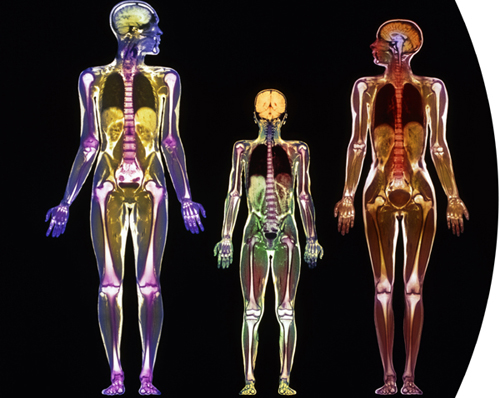
Colored magnetic resonance imaging (MRI) scans of a man (left), woman (right), and a nine-year-old boy (center). It is possible to pick out various organs and tissues, such as the lungs, bones, and brains.
Taking in the Cytes
Humans are complex multicellular organisms, so there is remarkable diversity and specialization among our cells. Our bodies manufacture around 200 different cell types, including adipocytes, erythrocytes, and neurocytes. This last chapter, drawing on what we know about cell biology, takes a tour of the scientific marvel that is the human body—while enjoying the cytes.
Colored magnetic resonance imaging (MRI) scans of a man (left), woman (right), and a nine-year-old boy (center). It is possible to pick out various organs and tissues, such as the lungs, bones, and brains.
Most people learned something about cells at school but perhaps never realized exactly how important, interesting, and stunningly beautiful they can be. All the cells featured in this chapter fall into these categories. Knowing what goes on inside them only makes them more fascinating and even more beautiful.
Erythrocytes (red blood cells)
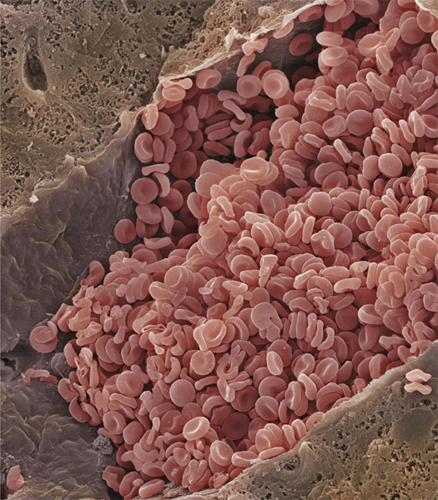
We begin with a classic and instantly recognizable cell: the disk-shape, biconcave (indented on both sides) red blood cell. In one sense, the red blood cell is not actually a cell at all. It has no nucleus and, therefore, no chromosomes—and is technically called a cell fragment. All blood cells, red and white, begin as multipotent blood stem cells called hemocytoblasts. Several divisions and differentiations result in the formation of immature red blood cells called normoblasts, and it is at this stage that the nucleus is expelled. Two more divisions later, and the tiny, enucleated red blood cell is circulating and maturing, ready to do its vital job of carrying oxygen around the body.
Most of its mass (95 percent of the dry weight) is hemoglobin protein, each molecule of which has four iron atoms at its center. Each iron atom can hold one oxygen molecule. It is to these iron atoms that the oxygen binds after it has diffused across cells in the lining of the lungs. Each hemoglobin molecule can hold four molecules of oxygen, and a typical red blood cell contains about one-quarter of a million molecules of hemoglobin, so each red blood cell can hold about a million oxygen molecules.
A teaspoon of human blood contains about 25 billion red blood cells. Over two million are created every second in a healthy adult human body. And every second the same number are destroyed, ready for recycling in the spleen, each one having lived for about four months. One of the breakdown products of all that hemoglobin is a yellow compound called bilirubin. It is responsible for the yellow color of nearly healed bruises and the brown color of feces as well as the background yellow color of urine.
Red blood cells account for about one in four of all the human body’s cells, but only about one-tenth of the body’s mass. This is because they are also among the smallest of our cells—about 200 of them lined up next to each other would cover about one twenty-fifth of an inch (1 millimeter).
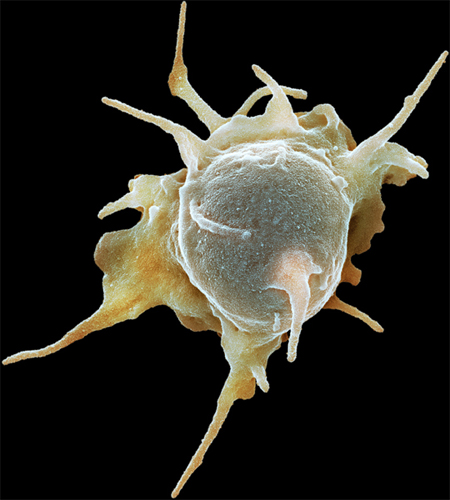
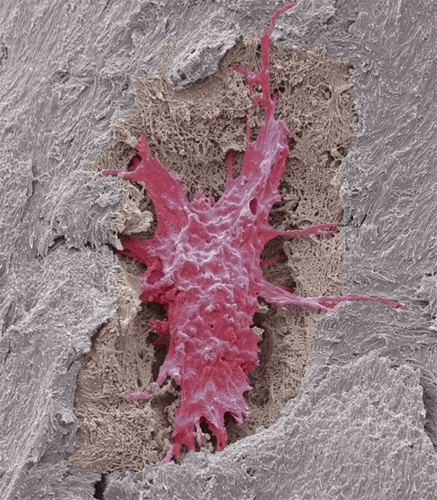
Colored scanning electron micrograph of an activated platelet (thrombocyte). Activated platelets secrete chemicals that cause the formation of a fibrin mesh, which traps platelets and red and white blood cells, forming a clot.
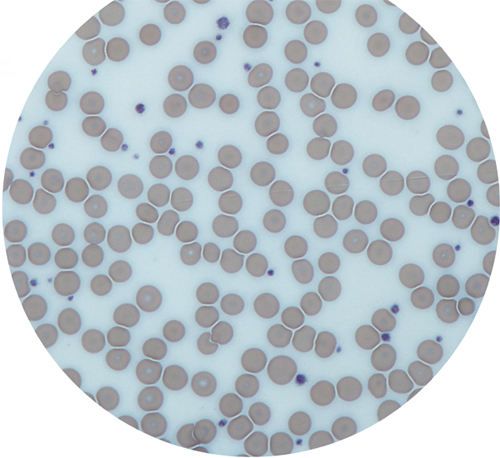
Light micrograph, its field of view dominated by red blood cells with several platelets (smaller, dark stained).
Thrombocytes (platelets)
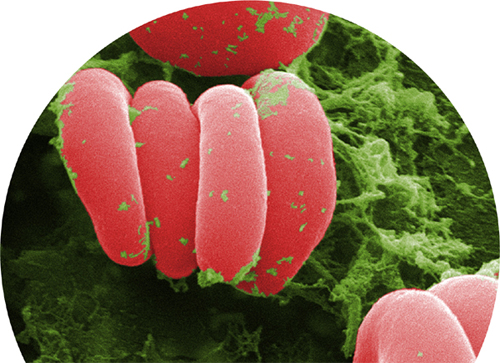
Another enucleated cell fragment found in the blood is the platelet, or thrombocyte. Platelets have a key role in the formation of clots. Without them, even a small injury would not stop bleeding and could become life threatening. They are attracted to any tear or damage in a blood vessel. When the platelets arrive, they are in their inactive, lens-shape form. As they become activated, they quickly change shape, as growing microtubules in their cytoskeleton push out waggly arms. The platelets adhere to the extracellular matrix around the damaged part of the blood vessel and to each other, forming the beginning of a clot, or thrombus.
The activated platelets now release the contents of granules they have stored within their cytoplasm. The contents include calcium, which is an essential ingredient in a cascade of reactions between soluble proteins in the blood, with the result that strands of the insoluble protein fibrin form. The fibrin strands hold the platelets, red blood cells, and the edges of the tear together as a clot. Some of the platelets’ granules contain growth factors that encourage the cell division required to heal the broken blood vessel. Activated platelets even release a bactericidal compound that will destroy certain types of bacteria if they enter the wound site.
Melanocytes produce the pigment melanin. They are found in hair follicles, and melanin is responsible for hair color. These cells are active throughout life, adding color to hair as it grows. Their activity slows with age, which explains why hair normally grows gray in later life. Melanin evolved long before humans did and is found in many other animal species. For example, it is one of the main pigments in bird feathers, many fish scales contain it, and it also forms part of the immune systems of insects.
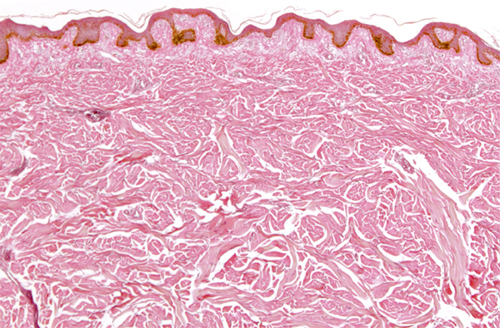
However, apart from determining the color of our hair, melanocytes are most important in our skin. They are found at the bottom of the epidermis, the top layer of the skin, and melanin is the body’s natural sunscreen; it is very good at absorbing ultraviolet-B radiation (UVB), a particular part of the ultraviolet spectrum that causes damage to DNA.
The human species evolved in Africa, near the equator, where UVB levels are high, and all early humans had dark (melanin-rich) skins to protect them. Some of the early people who migrated out of Africa tens of thousands of years ago ended up far from the equator, where UVB levels are much lower—so low, in fact, that dark skin put them at a disadvantage. A certain amount of UVB is needed for the body to manufacture vitamin D, which is essential for the uptake of calcium. So in higher latitudes, natural selection favored those individuals whose melanocytes made a little less melanin, giving rise to the wonderful range of skin tones present throughout the world today. Surprisingly, perhaps, there is very little difference in the number of melanocytes between any two people; different skin tones are caused by different amounts of melanin per melanocyte.
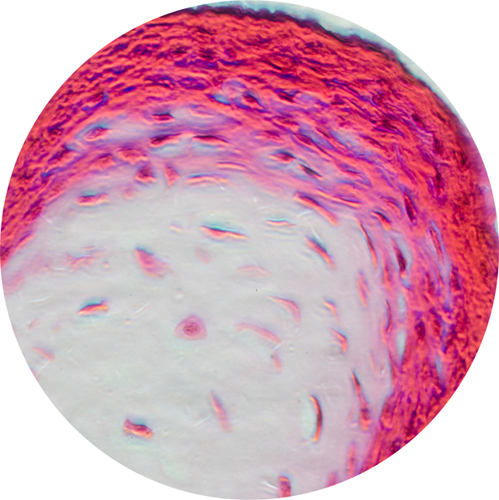
Light micrograph of a section through the dermis and epidermis of a person with fairly dark skin. The pigment-producing melanocytes are visible in a brown layer at the bottom of the epidermis.
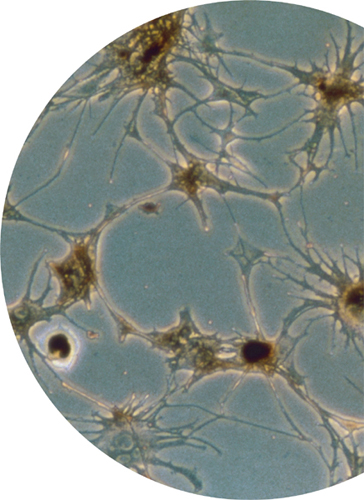
Inside melanocytes, melanin production takes place in membrane-bound organelles called melanosomes. These organelles are mobile; they can be transported to the ends of the cell’s extended branches, which are known as dendrites. In the epidermis of the skin, melanocytes even donate melanosomes to neighboring keratinocytes, the most common epidermal cell. (The keratinocyte gathers the melanosomes by engulfing the ends of the melanosome-packed dendrites.)
Light micrograph of an artificially grown culture of melanocytes, showing the cells’ dendrites (branches), along which pigment-containing vesicles can be transported.
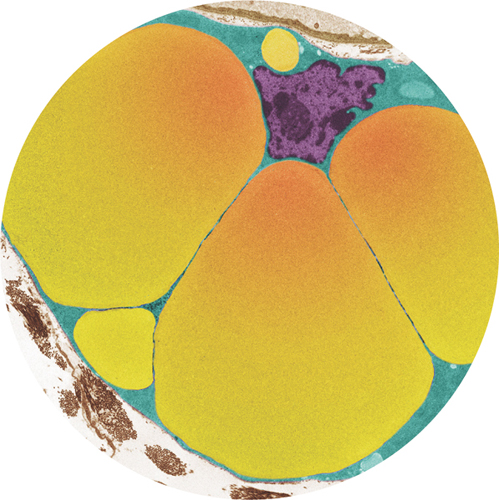
Fats are essential for building cell membranes, for high-density energy storage, and, under the skin (subcutaneous), for providing insulation against cold and protection against knocks. Fat is stored in fat cells, also called adipocytes or lipocytes. When we eat more calories than we burn the excess ends up here, as fat, even if the excess is sugar, because the liver easily converts sugars to fats. Inside an adipocyte, the fat is stored as one or more large, semiliquid globules.
These globules dominate the cell’s volume, and its nucleus is pushed against the cell membrane. If we gain or lose weight as fat, the number of adipocytes remains the same—each cell simply grows or shrinks to take on or lose fat. In situations of extreme weight gain, however, extra adipocytes are created. Fat cells are mitotic: they are routinely replaced by differentiating precursor cells called lipoblasts. Each fat cell lasts about ten years—so about one in ten is replaced each year. When they are replaced, they are destroyed and dismantled via apoptosis, and the fat globules transferred to the replacement cells.
Fatty tissue is simply made of fat cells bound together by connective tissue, mostly collagen. There are two types of fatty tissue: white fat and brown fat. The cells of brown fat are different from those in white fat. Brown fat cells contain a lot of mitochondria (they are what give the brown color) that burn the fat to produce heat. Newborns and young children have a lot of brown fat around their trunk to protect them against hypothermia. Adults have far less.
Fat cells produce a hormone called leptin, which has a powerful effect on appetite; the more leptin circulating in the blood, the more satisfied a person feels. The innate desire for high-energy food, left over from a time when food was hard to come by, is a stronger influence on appetite; the easy availability of sugary and fatty foods means that more and more of the world’s adipocytes are cramming in fat.
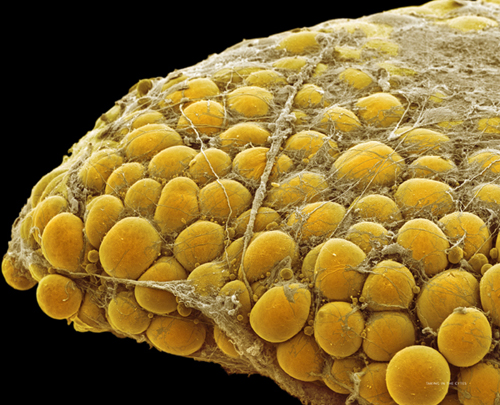
Colored transmission electron micrograph of an adipocyte, showing the large vesicles containing fat (yellow). The nucleus (purple) is pushed to the edge of the cell.
Colored scanning electron micrograph of fat (adipose) tissue. Individual adipocytes are held together with connective tissue, mostly collagen.
Colored scanning electron micrograph of an osteocyte (red) in a lacuna (depression) in mineralized osteoid (gray).
Bone is a dynamic material that is constantly being remodeled: cells called osteoblasts build it up, and cells called osteoclasts reabsorb it. Both cell types move around inside the bone in an amoeboid fashion. To make new bone, osteoblasts lay down tightly woven collagen fibers with a few other proteins mixed in—a composite material called osteoid. Crystals of carbonated hydroxyapatite (a form of calcium phosphate) mineralize the osteoid. Osteocytes are osteoblasts that become trapped within the matrix of mineralized osteoid; they are the most common type of cell in mature bone.
Osteocytes can live for decades. Once they are in position, they are responsible for sensing stresses on the bone. They feed this information to each other via long dendrites that interconnect them with their neighbors. The signals they share help to make sure the bone remodeling is appropriate to the level of stresses on the bone. They also produce growth factors if a bone sustains damage, encouraging the rapid remodeling necessary to make the bone strong again.
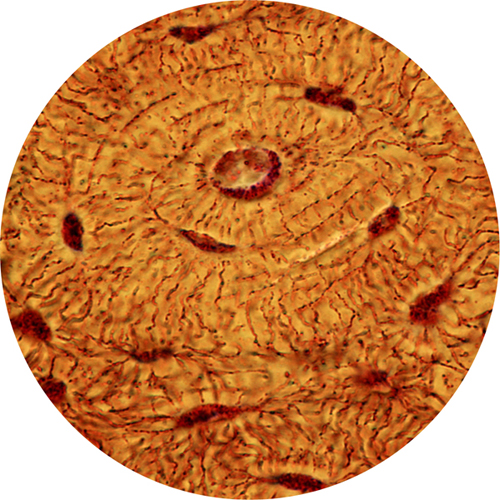
Light micrograph of an osteon—the basic unit of compact bone. At the center is the Haversian canal, which carries nerve and blood supplies. The osteocytes are the dark shapes and the thin lines that connect the osteocytes are microscopic canals, or canaliculi.
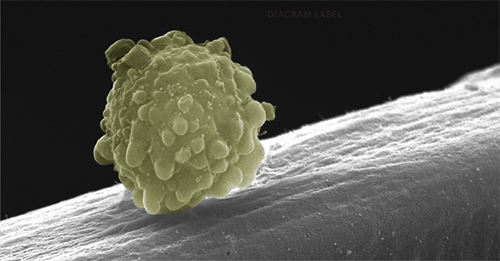
Colored scanning electron micrograph of a multipotent muscle stem cell migrating along the polysaccharide surface of a muscle fiber.
Myocytes (muscle cells)
There are two main types of muscle cell: smooth and striated (meaning it is patterned with fine parallel lines). Both work in the same way, with protein fibers contracting and relaxing to change the cell’s shape, although the inner structure of the two types is slightly different. Smooth muscle, which is involuntary (not under conscious control), is responsible for important housekeeping tasks, such as squeezing food through the digestive system and opening and closing our iris as light levels change. Striated muscle comes in two varieties. Involuntary cardiac striated muscle keeps our hearts beating without us having to think about it, while voluntary skeletal striated muscle makes our bones move relative to each other and keeps our lungs breathing.
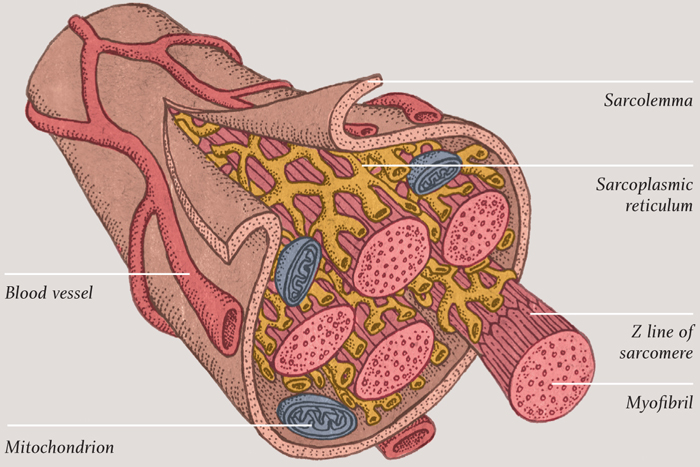
Striated muscle cells are more interesting and more beautiful to look at than smooth muscle cells—and most interesting among them are the skeletal cells. These are long, tubelike structures that contain many nuclei, because they are formed from several cells that have merged together. The cell membrane is a phospholipid bilayer, as with any other cells, but it also has a thin layer made of the sugary polymer polysaccharide tightly bound to it. The resulting cell coating is called the sarcolemma. Beneath that outer coating, the inside of the cell is dominated by thick bundles of fibrous protein called myofibrils. Clustered around these structures, perhaps not surprisingly, are many mitochondria, which can provide a constant supply of ATP to enable the myofibrils to do their job.
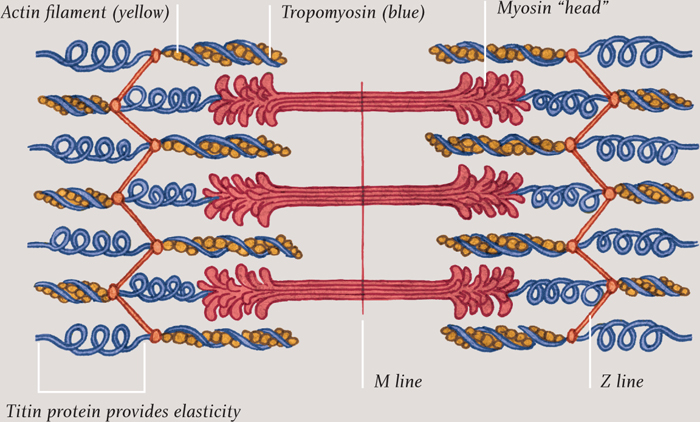
SARCOMERE (BASIC UNIT OF MUSCLE FIBER)
Illustration of a sarcomere, the basic unit of striated muscles. The Z line boundary is what gives this kind of muscle its striped appearance under a microscope.
Illustration of a skeletal muscle cell, showing the bundles of myofibrils, each one composed of a long line of successive sarcomeres.
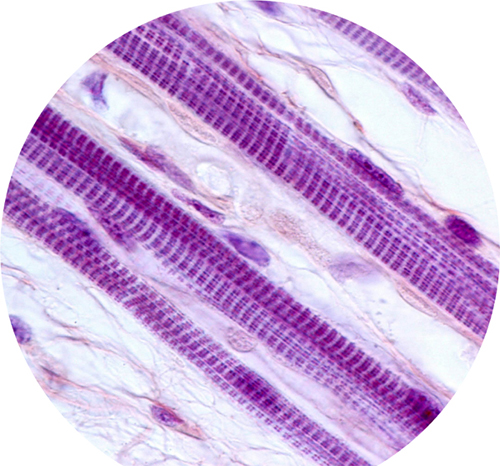
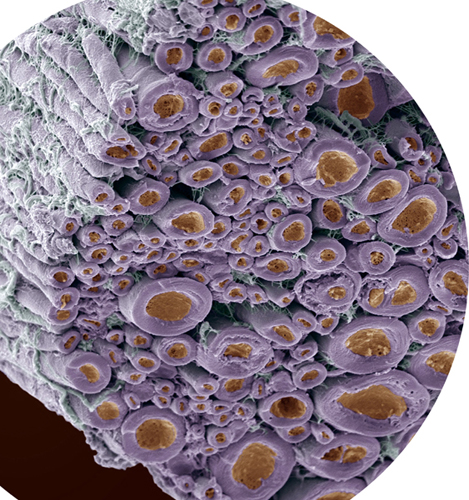
Light micrograph of striated skeletal muscle fibers. Every muscle contraction you have ever made was the result of motor proteins “walking” up and down inside tiny subunits in these fibers.
The job of a myofibril is to contract and relax. When it is relaxed, it can be pulled back to its original length by other muscles (muscles are arranged in opposing pairs—think biceps and triceps). Each myofibril is divided along its length into sections called sarcomeres. The contraction of the myofibril is the result of the sarcomeres shrinking, as two fibrous proteins, actin and myosin, slide past each other. The actin forms thin strands that are connected to both ends of the sarcomere—half of the strands to one end and half to the other. The myosin strands are thicker and are interspersed among the actin strands. When ATP molecules, produced by the mitochondria bind to the myosin the ends of the myosin molecules attach to specific sites on the actin strands. They pull the actin, drawing the ends of the sarcomere together by a tiny amount. Then the myosin lets go of the actin, ready to attach and pull again a tiny fraction of a second later. Each power stroke requires another ATP molecule.
When the muscle is relaxed, a protein called tropomyosin covers over the parts of the actin molecules where the myosin attaches, so the muscle remains inactive. When a nerve impulse arrives at the muscle cell, it causes calcium ions to be released into the cytoplasm. These attach to proteins along the length of the tropomyosin, which then changes shape to reveal the sites where myosin can attach, enabling the sarcomere to contract. It is truly remarkable to think that every move we make is the result of tiny straggly ends of myosin molecules grabbing hold of and pulling actin fibers, activated by a burst of calcium ions.
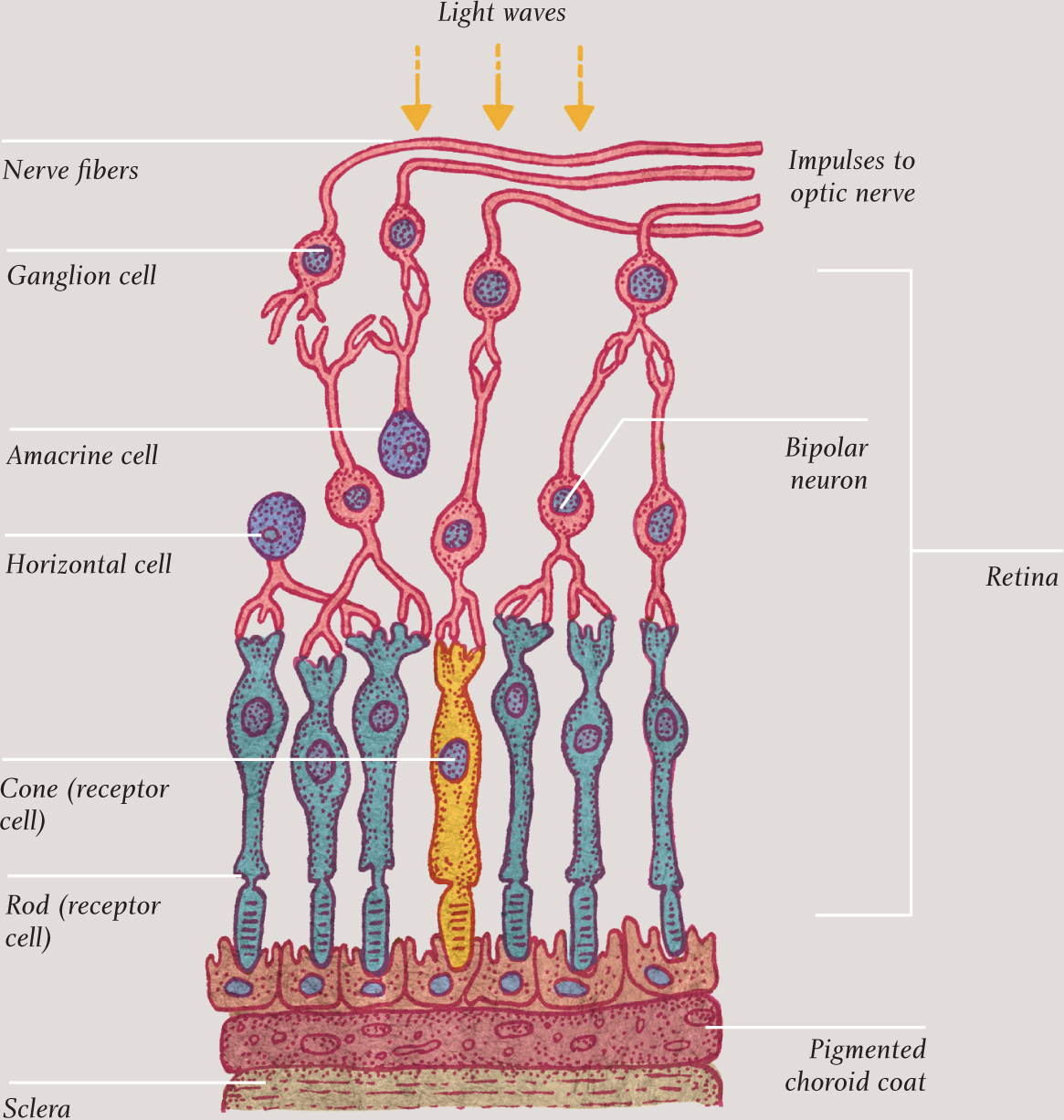
Retinocytes (rod and cone cells)
The term retinocytes, rarely used now, refers to all the cells of the retina, the layered structure at the back of the eye. There are several types of retinocytes but the most interesting and important are the ones that enable humans to see: the rod and cone cells. These have a very similar structure but slightly different purposes. The graph here shows the sensitivity of rod and cone cells to light entering the retina. The latter—of which there are three different types, each sensitive to a specific range of frequencies of light, roughly equivalent to red, green, and blue—are not particularly sensitive, so they do not work in low light, but they are responsible for color vision. Rod cells are extremely sensitive to light—far more so than cone cells—and are responsible for vision in low light. But they only detect light and dark, so they only enable black-and-white vision with a peak in the green part of the spectrum. Likewise color sensitivity peaks in that same region of the spectrum. Overall rod cells outnumber cone cells by about 20 to 1; a typical retina, with an area of 1½ square inches (10 square centimeters), has around 120 million rod cells and about six million cones. The cones far outnumber the rods, however, in the part of the retina that gives the sharpest vision, called the fovea.
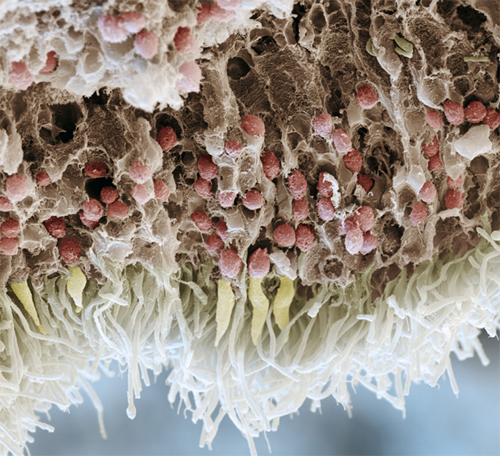
Colored scanning electron micrograph of a section through a human retina. Light enters from the top and passes through several layers of cells. The rods and cones are the white and yellow cells respectively, at the bottom of the image.
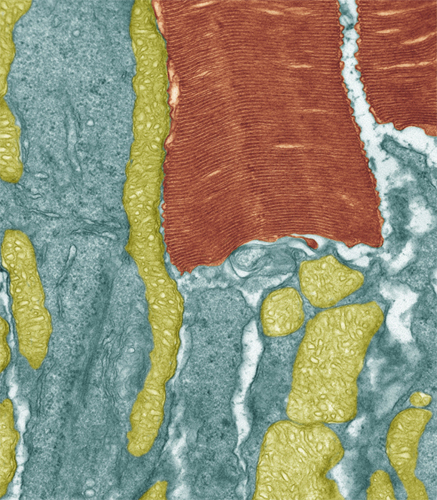
Colored transmission electron micrograph of a section through a rod cell showing the inner segment (blue) filled with mitochondria (green) and outer segment made up of stacks of folded membranes (brown).
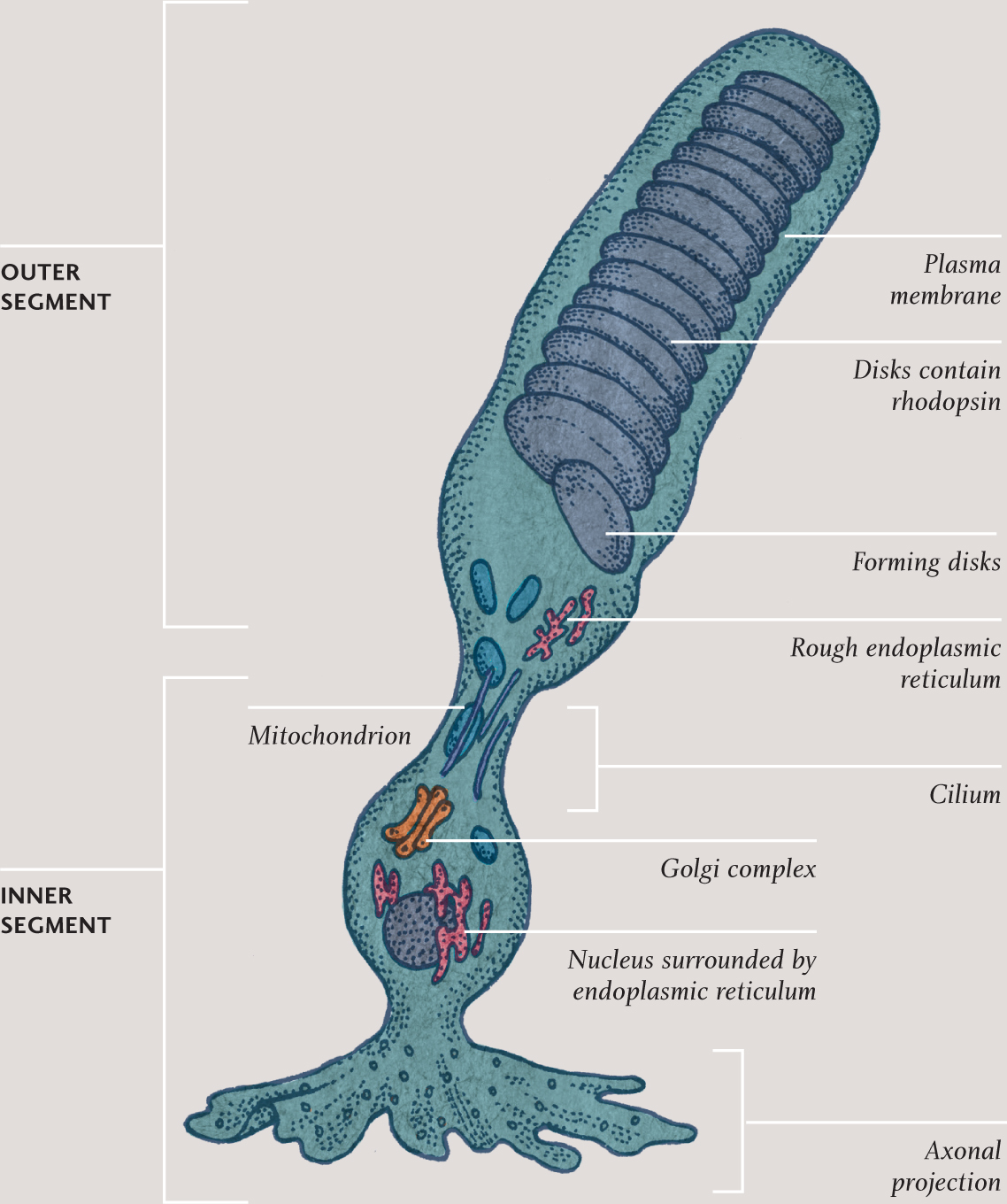
ROD CELL
Illustration of a rod cell. The part that links the two parts of the cell is a fixed cilium. The axonal projection connects to cells in other layers of the retina that pass signals on, ultimately to cells in the optic nerve.
Rod and cone cells have two very distinct ends. In both types, one end of the cell contains the nucleus and most of the organelles. The other end is dominated by stacks of membranes (actually modified cilia), formed by the cell membrane folding in on itself. In rod cells, this end is rod-shape and in cone cells, it is cone-shape. Each type of cell constantly pumps sodium ions out at one end and takes them back in at the other. Because sodium ions are electrically charged, this circulation constitutes a tiny electric current called the dark current. It is the interruption of this current, when the cell absorbs light, which creates the signal that is ultimately passed along the optic nerve to the brain.
Embedded within the membrane stacks at the business end of each rod and cone cell are pigment molecules. It is these that actually absorb incoming light. The pigment in rod cells is rhodopsin and it is sensitive to light across a wide range of frequencies, centered on blue-green. The equivalent pigments in cone cells are called photopsins. Both rhodopsin and photopsin molecules are in two detachable parts: one large opsin and a smaller retinal molecule (retinal is made from vitamin A). When a pigment molecule absorbs light, the retinal molecule changes shape and temporarily breaks away. This interferes with the circulation of sodium ions and begins the process of creating a nerve signal.
Surprisingly, perhaps, the light-sensitive end of each rod and cone cell is found in the deepest layer of the retina, farthest away from the front of the eye. Light has to pass through eight layers of cells and two membranes before it reaches the stacks containing the pigments. At the end of the cell farthest away from the light-sensitive part, and closest to the front of the retina, the cell passes on signals to nerve cells; these are bundled together to form the optic nerve that connects to the brain.
An average human brain contains around 85 billion neurocytes—nerve cells, or neurons. Each neuron in the brain receives signals from up to a thousand other neurons and, depending upon the overall effect of these signals, either fires or doesn’t fire, passing on its own impulse to many others. So the brain contains an incredibly complex network of interconnected neurons, all busily communicating with each other. Those interconnections form and break as we experience the world and learn new things. Somehow—no one exactly knows exactly how—the signals zipping here and there in the brain give us awareness, recall, and the ability to act, as well as subconsciously monitoring and regulating the vital organs of the body.
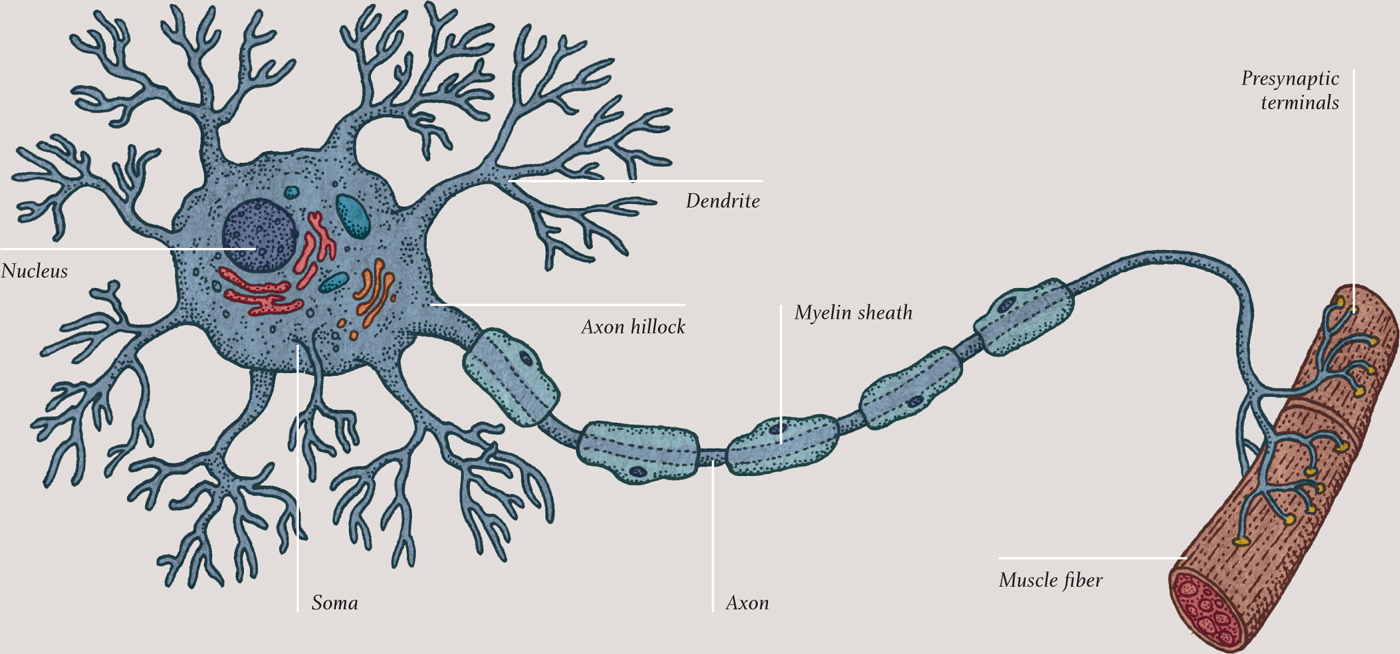
All this activity would be for nothing were it not for neurons that connect the brain to the rest of the body and to the outside world. Signals enter the brain via sensory, or afferent, neurons, whose origins are in the sensory receptors, such as rod and cone cells in the retina and pain and pressure receptors in the skin. Signals leave the brain via motor, or efferent, neurons and cause muscle cells to contract. Neurons come in several different shapes and sizes, but they all share certain features: they gather input signals at extensions called dendrites, they process the signals in the cell body, or soma, and they produce an output signal that passes along an extension called an axon. The end of a typical axon has many branches, called terminals, at which other neurons connect their dendrites—or, in the case of motor neurons, at which a muscle cell receives signals.
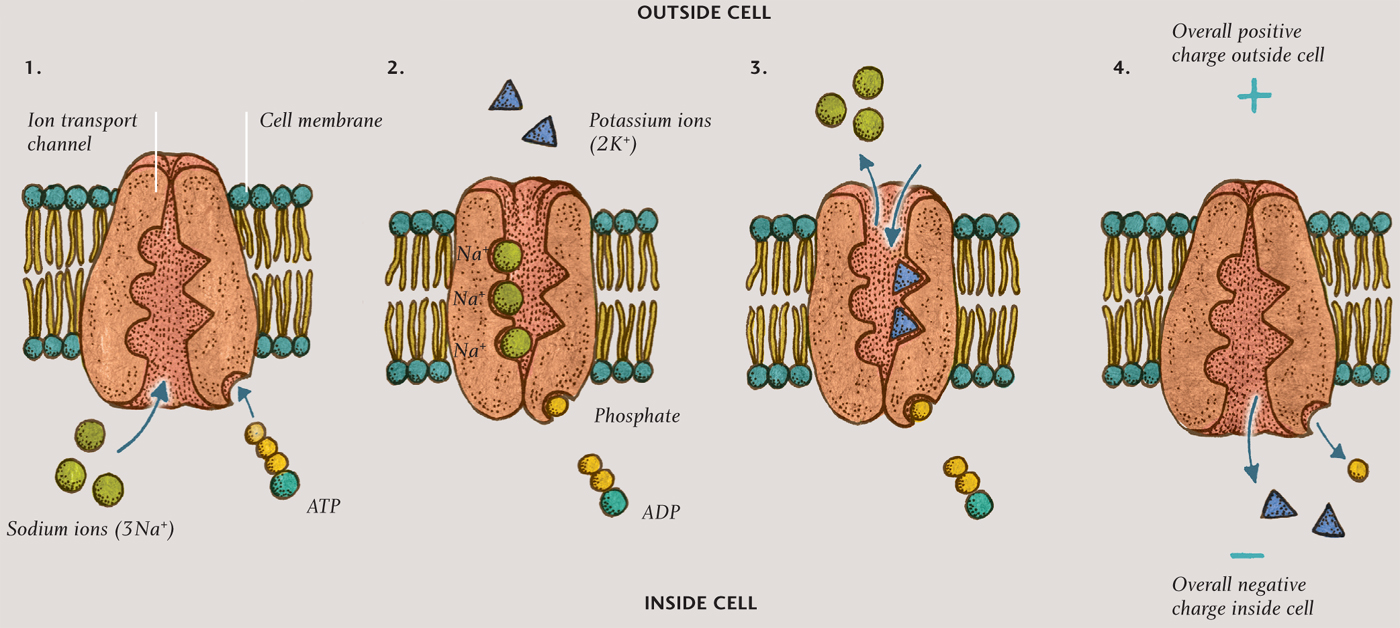
Nerve impulses are electrical in nature. When it is not firing, every neuron has more positive charge on the outside than the inside, thanks to a remarkable cluster of molecules that sits in the cell membrane. It is called the sodium-potassium pump, and it pushes three sodium ions (3Na+) out of the cell for every two potassium ions (2K+) that it pulls into the cell. The imbalance of charge across the membrane gives the inside of the cell a voltage of about minus 70 millivolts relative to the outside (the inside is more negatively charged than the outside). This voltage is called the resting potential. The cell remains in this polarized state until a dendrite receives a signal from another neuron, or the cell decides to fire. In either case, a disturbance in the ion concentration, called the action potential, passes along the axon or dendrites.
SODIUM-POTASSIUM PUMP
Illustration of the sodium-potassium pump, which creates the resting potential by pumping more sodium ions (Na+) out of the cell than it pumps potassium ions (K+) into the cell. The resulting resting potential leaves the interior of the cell more negatively charged than the outside, as shown in the final step.
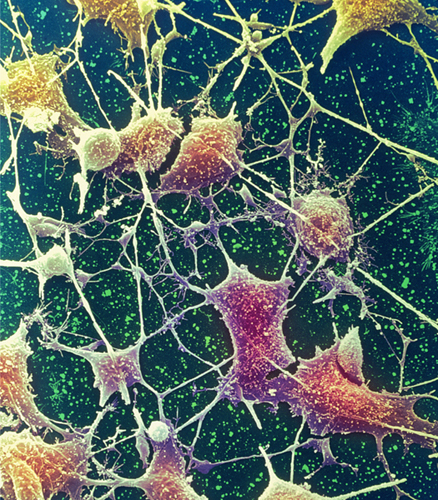
Colored scanning electron micrograph of neurones from the brain. Cell bodies are brown, and the axons and dendrites are clearly visible.
Colored scanning electron micrograph of a freeze-fractured section through a bundle of myelinated nerve fibers. Myelin sheaths (purple) can be seen surrounding the axons (brown). Connective tissue (blue) is also visible.
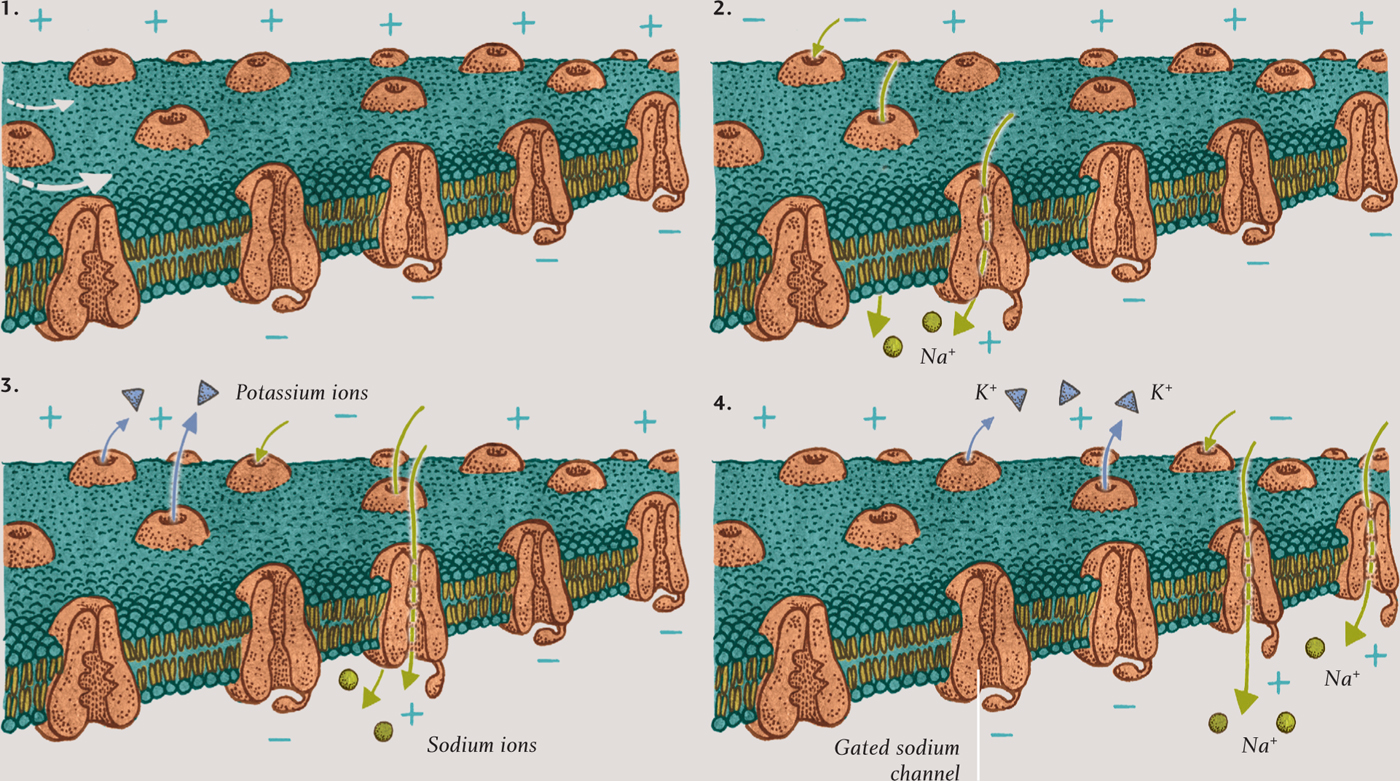
ACTION POTENTIAL (PASSING FROM LEFT TO RIGHT)
This sequence shows how a nerve signal moves along a neuron. When a neuron fires, the resting potential (1) is disturbed and sodium ions are pulled into the cell through gated channels (2). The channels close afterward (3), ensuring that the signal passes only one way (4).
The action potential moves along the neuron’s axon and dendrites because of the presence of protein clusters called gated sodium channels, which are dotted all over the membrane. These channels are normally closed, but when an action potential arises, they open and enable sodium ions to rush into the cell, depolarizing that part of the membrane. The voltage across that part of the membrane initially becomes zero and then for a moment rises above zero—and that causes adjacent sodium channels to open.
It is important that nerve impulses travel in one direction only—from input at the dendrites to the cell body, and from the cell body to the output at the axon terminals, and from brain to muscle, and from receptor to brain. The traveling action potential can go only one way, because the channels close shortly after the potential reaches its maximum and remain closed for a few milliseconds. These cannot, therefore, be opened again by the action potential they have just passed on to adjacent channels down the line. This remarkable process is how nerve signals pass within the brain and throughout the body. Some neurons are sheathed by insulated sections of a fatty material called myelin. These enable the action potential to jump quickly from gated sodium channels in one uninsulated node to the channels in the next. Neurons that are myelinated in this way transmit impulses much faster than unmyelinated neurons.
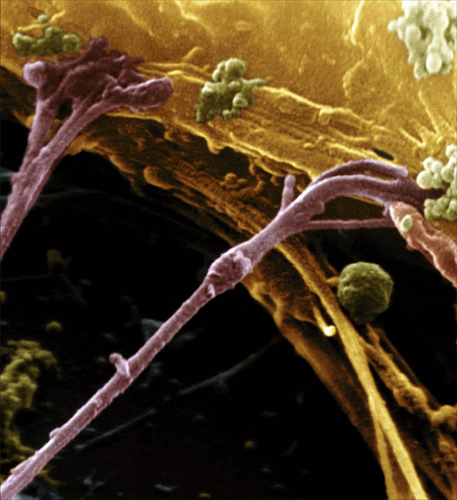
The transmission of a nerve impulse from one neuron to another is equally remarkable. The neurons are not physically connected; instead, there is a small gap called a synapse (see chapter five) between the axon terminal of one neuron and the dendrite of another. The nerve signal is transmitted across the synapse by the release of compounds called neurotransmitters from the axon terminal. Neurotransmitters are manufactured in the rough endoplasmic reticulum just outside the nucleus in the cell body. The Golgi apparatus nearby (see chapter two) squirrels away the neurotransmitter molecules into vesicles, bound for the neuron’s cell membrane. The molecules are then “walked” along the cytoskeleton by motor proteins to the axon terminals. Here, they stay, ready to be released quickly, as the vesicles burst like bubbles at the membrane in the axon terminal.
Neurotransmitters pass across the synapse and dock into receptor proteins in the membrane of the dendrite on the other neuron. Some receptors cause the membrane to remain polarized, in which case they inhibit the receiving neuron’s membrane from creating an action potential. Other receptors will depolarize the membrane, exciting the membrane to create an action potential that travels along the dendrite toward the cell body. Meanwhile, neurotransmitter molecules are released back into the synaptic gap and are reabsorbed by the first neuron, ready for the next time it fires.
A neuron has many dendrites, each receiving signals from another neuron. If enough dendrites are excited and create action potentials at the same time, reaching a threshold, they create an output signal at the other side of the cell body, at the very beginning of the axon—a part of the cell known as the axon hillock. When that is achieved, the neuron will fire.
Of all the cells in the body, neurons are perhaps the most amazing: they really do make you think.