2
Cell Structure
MICROSCOPES
Anything as small as a cell was unknown before sophisticated optics became available. During the seventeenth century, ground lenses that were being used for eyeglasses were first arranged at opposite ends of a tube, creating a small telescope. It was a short step from the invention of the telescope to the invention of the microscope. Objective lenses (those that are nearest what you are looking at) of a telescope have long focal lengths (focus on things far away); objective lenses of a microscope have short focal lengths (focus on things that are close). One of the first microscopes was constructed by the Dutchman Antonie van Leeuwenhoek. With this light microscope, the examination of specimens was facilitated by thinly slicing them, allowing light to pass through. By staining the specimens, it was possible to emphasize internal structures. For instance, staining cellular fluids pink and staining solid, hard structures purple provided increased contrast, enabling those who study cells, known as cytologists, to discern these structures more clearly.
While some light microscopes permit researchers to view objects at as much as 1,500 times (1,500×) their actual size, stereomicroscopes, also called dissecting microscopes, magnify objects from only 4× to 80×. With two eyepieces, the advantage of this low-powered, three-dimensional view is that researchers can investigate much larger objects, such as the venation of insect wings (see Figure 2.1).
After the invention of the light microscope, the next major technological advance for cell researchers was the development of the electron microscope (EM), which occurred in the early 1930s. It not only improved the ability to see smaller structures with greater magnification – so they appeared larger – but also enhanced the ability to see things more clearly, or with added resolution.
When it was discovered that the illumination of specimens with blue light under the light microscope lent considerably greater resolution than with any colors of longer wavelengths, researchers speculated that using shorter wavelengths might add even more resolution. However, wavelengths shorter than those of violet light are not visible to the human eye. This problem led to the invention of the transmission electron microscope (TEM), which utilizes a beam of electrons that travel in shorter wavelengths than those of photons in visible light. These electrons are passed through a thinly sliced specimen within a vacuum to prevent any electrons from being deflected and absorbed by the gas molecules in the air. Then the electrons are focused with electromagnets on a photographic plate, producing an image that is considerably better than that obtained with a light microscope.
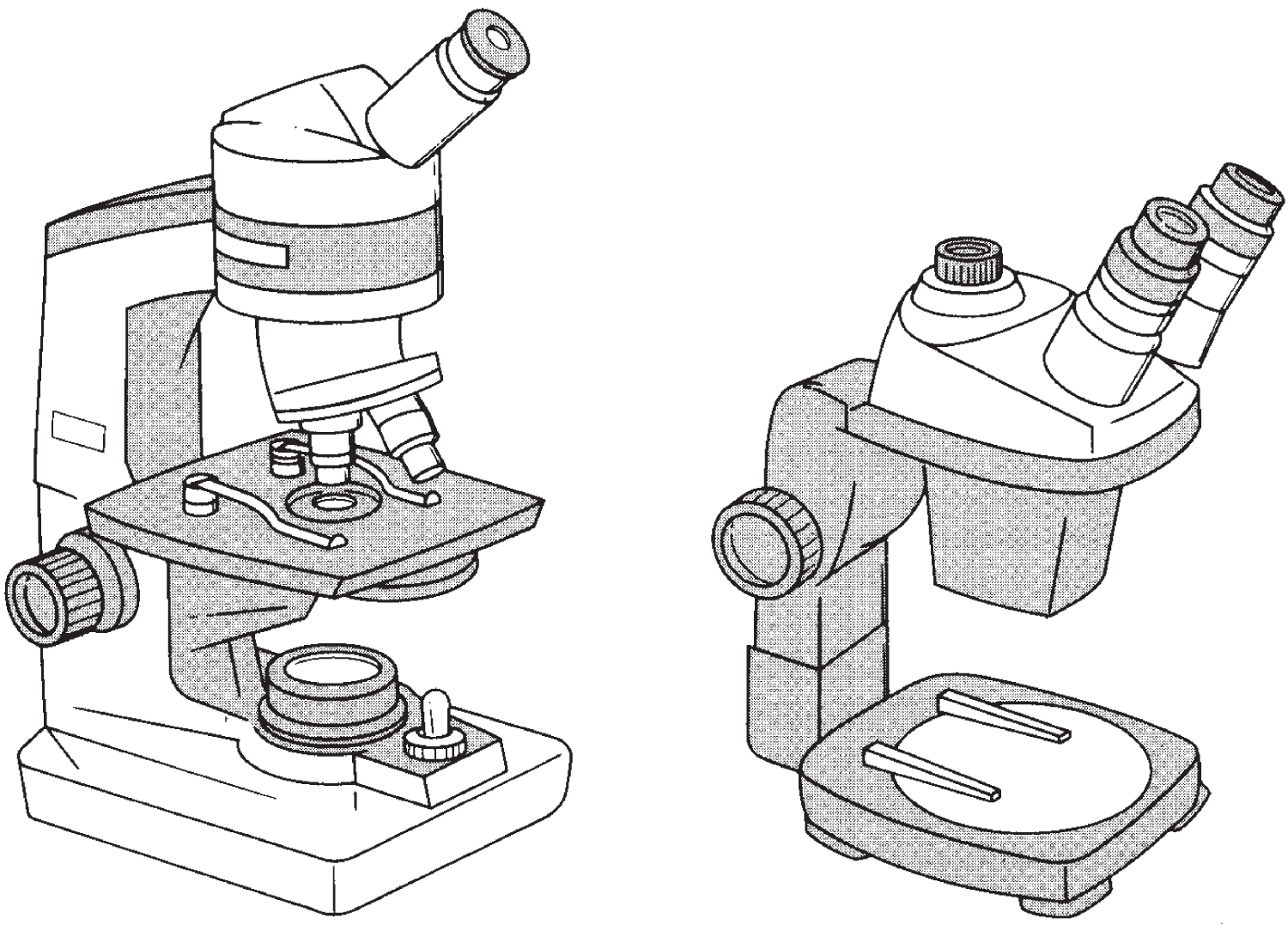
Figure 2.1 Light microscope (left) and stereomicroscope (right). The resolving power of the light microscope rarely exceeds a magnification of 1500×. The stereomicroscope, sometimes called a dissecting microscope, has two eyepieces, which render relatively large objects three-dimensional. Magnification ranges from 4× to 80×.
Then in the 1950s, the scanning electron microscope (SEM) was invented, which focuses electrons that bounce off the specimen. Since the SEM has less resolving power than the TEM, it doesn't require a vacuum and allows researchers to view some smaller organisms alive.
The most recent advances in microscope technology allow scientists to observe living cells with even greater magnification. One of the newly developed techniques is called contact X-ray microscopy. Another microscope, the scanning tunneling microscope, enables scientists to photograph molecules. An additional tool is the magnetic resonance imaging (MRI) scan machines that use radio waves and strong magnetic waves to create clear images of tissues and organs inside a body.
CELLS
With the availability of the first microscopes, researchers began to observe the microscopic structure of many substances, and in 1665 the Englishman Robert Hooke described what he called cells that he observed in a piece of cork. He used this term because the cork appeared to be composed of thousands of tiny chambers that resembled the individual sleeping rooms in monasteries at the time, which were called cells. He was not aware that he was viewing just the cell walls, which were the only structures remaining from what had once been living cells.
Hooke's initial discovery led to other advances, such as the finding that unlike plant cells, which have thick cell walls, animal cells lack such a wall and instead have only a thinner, generally more flexible plasma membrane (or cell membrane) (see Figure 2.2).
Cells were then found to exist independently or as one small part of an organism consisting of many cells, a multicellular organism. Hooke was the first to discover that some organisms consist entirely of a single cell. Such single-celled or unicellular organisms, which include the tens of thousands of species of bacteria and protozoa, carry out all necessary life-supporting functions within one cell without the help of other cells. In contrast, multicellular organisms have cells with specific functions, and together the aggregate of cells embodies a complex organism.

Figure 2.2 Linearly arranged cell walls in the annual growth rings of a pine tree. In the aggregate, it is the cellulose of each cell wall that gives the tree its rigidity.
CELL THEORY
It took about 150 years after Hooke discovered cells before several important related facts were articulated. In the first half of the nineteenth century, three German scientists, Matthias Schleiden, Theodor Schwann, and Rudolf Virchow, were the first to explain the basic tenets of what we now call cell theory:
- Cells are the fundamental units of life.
- Cells are the smallest entities that can be called living.
- All organisms are made up of one or more cells.
Viruses were not known when cell theory was first presented. Although viruses are much smaller than cells, we understand how viruses reproduce inside cells, and although it is somewhat controversial, most scientists do not consider viruses to be living because viruses do not possess cells, they have no metabolism, and they cannot reproduce on their own. They do not possess a metabolism, and they do not have cellular division. They only reproduce inside cells of other species.
CELL STRUCTURE AND CELL SIZE
The longest cells are certain nerve cells (neurons), which can reach over a meter in length. While an ostrich egg is 1,500 times the size of a human egg cell – which is 14 times the size of a human red blood cell, itself as much as 35 times the size of many small single-celled microorganisms – most cells do have one thing in common: They tend to be quite small. While the size range reflects considerable diversity, most cells are 0.5–40 microns in diameter (1,000 microns equals 1 millimeter, 1,000,000 microns equals 1 meter; micron is short for micrometer).
Small cell size is thought to be a function of the restriction placed on cells by the ratio of surface area to volume. Cells are constantly absorbing molecules from the surrounding medium and releasing molecules into the surrounding medium. These processes are more readily accomplished when a cell is small and the ratio of surface area to volume is quite large. As a cell increases in size, the amount of volume inside the cell increases much more rapidly than the amount of surface surrounding the cell, and in time the cell becomes too large to maintain a stable internal environment. Or, stated more simply, if a cell were to grow too large, it would not be able to expel enough of the wastes it would produce, and therefore it would die.
Many scientists believe that it is more difficult for the nuclear material to maintain control over the entire internal environment when a cell is over a certain size. Therefore, if a small nucleus is most often the rule, then an upper limit is placed on the size of most cells.
CYTOPLASM AND NUCLEOPLASM
Except for the nucleus (or nuclei), everything within the plasma membrane is called the cytoplasm. The nucleoplasm consists of the contents within the nuclear membrane.
The cell's interior is composed largely of a complex solution as well as a heterogeneous colloid.
A solution is a homogeneous mixture of two or more components in which the particles of the different substances are so small that they cannot be distinguished.
A colloid usually contains particles that are too small to be seen but are large enough not to form a true solution. The particles don't settle out at an appreciable rate. Different areas inside a cell may be in different colloidal states.
Some areas are in a sol state – that is, the colloidal particles, which are usually macromolecules, are randomly dispersed throughout the area.
The other parts of a cell may be in a gel state, in which the colloidal particles interact and form a spongy network. A colloid in the gel state forms a semisolid.
Changes from the sol to the gel state may be stimulated by changes in pressure, agitation, temperature, pH, salt concentration, or the concentration of other substances.
Some mixtures containing proteins and lipids have properties of true solutions as well as some sol-gel capabilities of a colloid, and this often typifies the internal fluid environment of a cell.
CELL MEMBRANE
The cell membrane (also called plasma membrane) is the membrane surrounding the cytoplasm at the cell's surface. These membranes contain varying amounts of proteins and lipids. The specific types of lipids, which are fat-soluble substances located in the cell membrane, are primarily phospholipids and steroids. The total thickness of the cell membrane, which is sometimes also called the plasma membrane, is only about 80 angstroms (there are 10 million angstroms per millimeter; there are 10 billion angstroms per meter).
MOVEMENT THROUGH THE CELL MEMBRANE
In addition to providing protection, shape, and strength to the cell, the membrane helps regulate the flow of materials in and out of the cell. Many processes work in concert to maintain the cell's constant internal environment. The condition of constant internal environment is referred to as chemical homeostasis, and the methods a cell employs to maintain this constant environment are described below. The two basic methods require different energy input. When the movement of molecules occurs on its own, without any organized energy input from the cell, the movement is termed passive transport. When the cell expends energy to move molecules from one location to another, the process is known as active transport.
Diffusion
One type of passive transport, called diffusion, may occur in a dynamic system such as a gas or liquid, in which particles may be moving in a manner that appears random (see Figure 2.3). In a system where diffusion occurs, a net movement of particles can be observed over time from regions of higher concentration of that substance to regions of lower concentration of that substance.
The particles moving from the area of greater concentration will produce a concentration gradient ranging from the point of highest concentration to the point farthest away, where the lowest concentration occurs. The particles don't move in a straight line, but the concentration gradient is instead a result of many small movements. The tendency is for molecules to move down a concentration gradient – that is, they go from an area of highest concentration of that molecule to where the concentration is less. This movement leads to the point when the molecules are eventually dispersed evenly throughout the environment, where they are all able to move equally freely.
The movement of the particles can proceed at different rates, depending on such factors as the concentration and temperature of the medium. Some membranes are impermeable to the movement of anything through them. Other membranes may exclude some substances, but they are permeable to others. A partially or differentially permeable membrane is called a semipermeable membrane.
Osmosis
Another example of passive transport is osmosis, which involves the movement of water through a semipermeable membrane (see Figure 2.3). The pressure exerted by dissolved particles in a solution or a colloid, known as osmotic pressure, moves water across a semipermeable membrane. When a semipermeable membrane separates two solutions of pure water, the two sides are at equilibrium – that is, the osmotic pressure is equal on both sides. When the osmotic pressure is the same on both sides of a semipermeable membrane, each solution is said to be isosmotic or isotonic. This means that each medium has the same concentration of osmotically active particles, and each has the same osmotic pressure, so neither tends to lose or gain appreciable quantities of water via osmosis.

Figure 2.3 The top three boxes illustrate diffusion. Water is represented by clear space with no dots. The dots represent the solute, which is the component that is dissolved in a solution (the solution is the solvent). The dissolved solute molecules in the solution enter the enclosed area and move about within the medium (with diffusion the medium doesn't have to be a liquid, it can also be a gas). Eventually the molecules become randomly distributed throughout the enclosure. The bottom three boxes illustrate osmosis. The percentage of dissolved materials inside the enclosure is greater than the percentage outside the enclosure. To counteract the difference, water molecules diffuse through the membrane. The osmotic process continues until the percentage of water molecules inside the membrane equals the percentage outside, unless the membrane becomes stretched to capacity and can no longer accept additional molecules. At such a point it either remains firm (turgid), or it may burst, or in some cases a mechanism such as active transport pumps water out of the cell.
Another alternative exists when a cell is in a medium with a higher concentration of osmotically active dissolved or colloidal particles, rendering a higher osmotic pressure. In such a situation, water moves across the cell membrane into the medium that is hyperosmotic (also called hypertonic) relative to the solution inside the cell.
When a cell is in a medium with a lower concentration of osmotically active dissolved or colloidal particles, and the solution thus has a lower osmotic pressure than that inside the cell, the cell is said to be in a medium that is hypoosmotic (or hypotonic) relative to it, so water moves into the cell.
To avoid confusion, one should be careful to note the point of reference of these terms. For instance, when the internal environment of a cell has a higher concentration of osmotically active particles than the external medium, this means the medium inside the cell is hyperosmotic relative to the external environment. Either way one chooses to state it, the water still moves from the hypoosmotic environment into the hyperosmotic environment. Water moves from where the osmotic pressure is low to where the osmotic pressure is high (to where there are more osmotically active particles). If this process is allowed to continue, all things being equal, the solutions on either side of the semipermeable membrane should eventually become isosmotic to each other.
If particulate matter is poured into one of the two previously isosmotic solutions, and the particles dissolve into solution, then the equilibrium will be upset and the osmotic pressure that now exists between the two sides will be unequal. Water will move from the area of lower concentration of dissolved molecules.
The permeability of different types of plasma membranes varies significantly. An amoeba's plasma membrane has less than 1% of the permeability to water of the membrane of a human red blood cell. Most plasma membranes are quite permeable to water and other substances such as many simple sugars, amino acids, and lipid-soluble (fat-soluble) substances. Plasma membranes are relatively impermeable to larger complex molecules such as polysaccharides and proteins.
Permeability to small molecules indicates there are pores through which these particles can pass. It may be that these pores open and close according to the cell's needs at a given time. The membrane's permeability to lipid substances is related to the lipids that make up part of the membrane. Lipids are able to dissolve right through the membrane. In the cell membrane are carrier molecules that help move certain substances in and out of the cell. They are described in the following discussions on facilitated diffusion and active transport.
Facilitated Diffusion
One of the two main types of transport that involves active transport, as opposed to passive transport, is known as facilitated diffusion. Here, carrier molecules move substances either in or out of a cell in a manner that increases the rate of diffusion across the membrane. The mechanism involved combines molecules that are to be moved across the membrane with the carrier molecules in the membrane. In such a manner, molecules are picked up on one side of the membrane and released on the other side.
A common example of facilitated diffusion is the rate at which cells take up glucose, an important simple sugar in many biological systems. Most cells absorb glucose much more rapidly than the rates at which such absorption would occur without any added help. In concert with the carrier molecules, as the cell starts to use the glucose rapidly, the concentration on the inside of the cell decreases, and because of the increased concentration gradient, glucose is moved into the cell more rapidly than usual.
Facilitated diffusion works in both directions. Sometimes it is equally important to move substances into the cell as it is to move other substances out of the cell. Certain hormones can affect the rates at which some substances move across cell membranes. For instance, insulin increases the rate of glucose uptake by some tissues. While glucose normally moves along a concentration gradient, facilitated diffusion helps increase that rate.
Active Transport
To move substances such as glucose across a cell membrane against a concentration gradient, energy has to be expended. The process by which substances move against their concentration gradients is termed active. There are many types of active transport systems. Some take up amino acids; others move peptides, nucleosides, or potassium. These substances are moved with an appropriate carrier system. Both of the most extensively studied active transport systems use the high-energy molecule adenosine triphosphate (ATP).
In some cases, it appears that specific globular proteins within the cell membrane temporarily change shape under certain conditions. The modification may temporarily open a pore wide enough for the transported molecule to pass through. When active transport is involved, energy from ATP may help change the conformation of the carrier molecule so it will pick up the molecule that is to be moved, or ATP may help detach the transported molecule from its carrier once it has been moved to the other side of the membrane (see Figure 2.4).
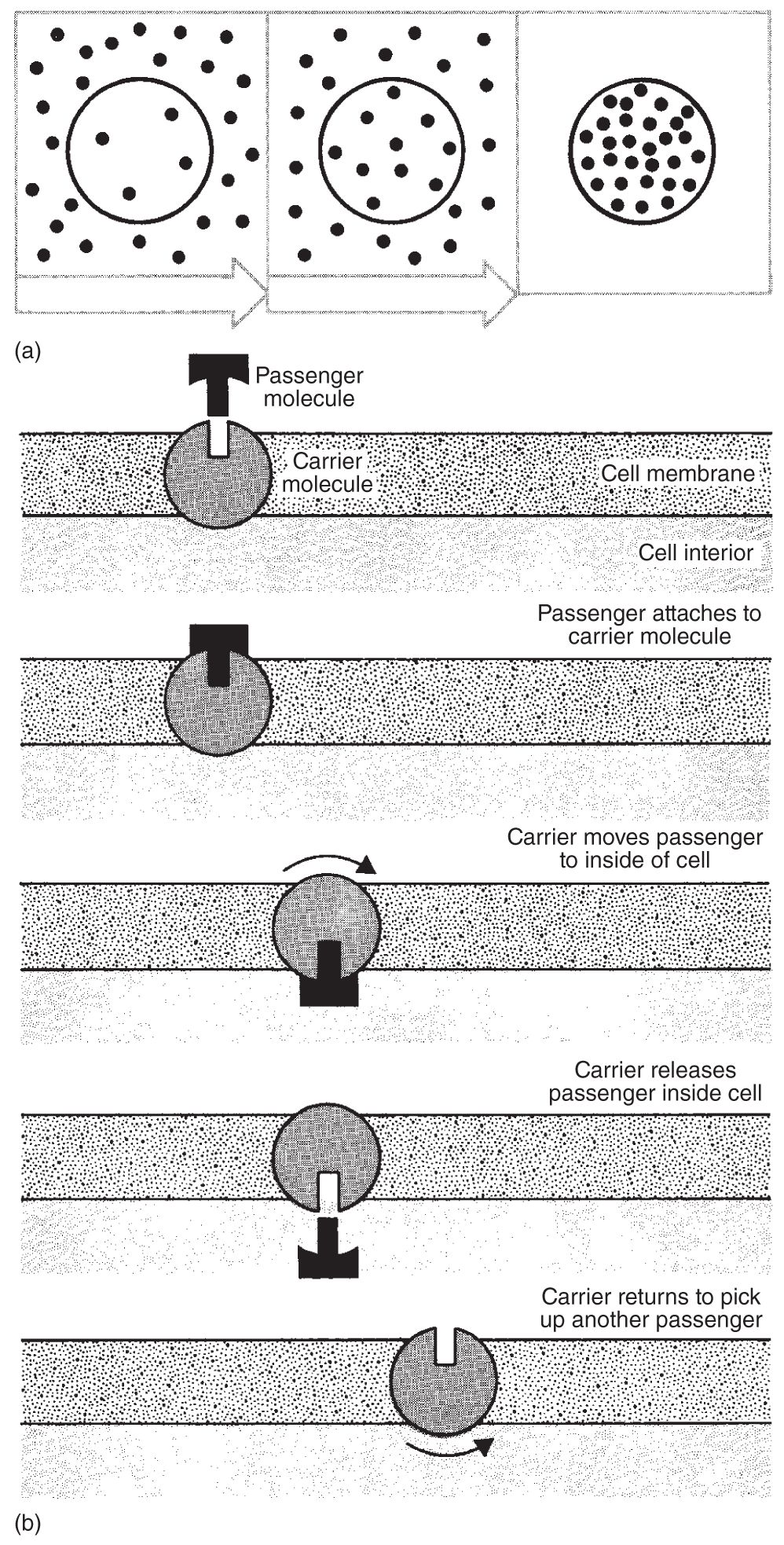
Figure 2.4 Active transport: (a) A cell concentrates certain kinds of molecules inside, or may move them outside the membrane, utilizing a process that expends energy; (b) the process works with carrier molecules as illustrated.
Movement of Large Particles Across Membranes
The plasma membrane has the capacity to engulf small particles on either side of the cell and then fuse with these particles; or sometimes the portion of the plasma membrane that surrounded the small particles simply separates, pinching itself off and drifting away. When liquids or macromolecules are engulfed by the membrane and are then moved to the other side, the process is known as pinocytosis. The same process, but on a larger scale, that takes on larger particles is phagocytosis. These engulfed particles pinch off with part of the membrane and move in or out of the cell. The process that moves particles into the cell is endocytosis, and the process that moves particles out of the cell is exocytosis.
PLANT CELL WALL
All cells have a plasma membrane, but some also produce a nonliving outer cell wall. The cell wall is a product of the cytoplasm that supports and provides shape for the cell. Most bacterial cells have a rigid or semirigid cell wall outside the plasma membrane that is usually composed of a polymer of glucose derivatives attached to amino acids. Sometimes certain bacteria have an additional layer outside this known as lipopolysaccharide (a polymer composed of lipid and sugar monomers).
The complex polysaccharide that composes most plant cell walls is cellulose (and related compounds). The micro wall allows a wide range of compounds to pass through easily. It is the cell membrane that determines which materials may enter or leave the cell.
In complex, multicellular plants, the primary cell wall is the outer layer that is laid down first by the young cells. And in some of these plants, where two cell walls come in contact, another layer is laid down between the primary cell walls. This is known as the central or middle lamella, which is one continuous layer between the cells, and it is usually composed of pectin, a complex polysaccharide, generally in the form of calcium pectate. It is the pectin that binds the cells together.
Plant cells that become harder, or woodier, usually do so by adding layers to the cell wall, known as the secondary wall. This wall is also deposited by the cytoplasm and is inside the primary wall. Rather than being laid down in a loose network, as in the primary cell wall, the long thread-like structures that make up much of the cell wall, known as cellulose microfibrils, are laid down parallel to one another in the secondary cell wall in a series of layers, or lamellae. Each layer has fibrils oriented at angles of about 60° to the fibrils of the next lamellae. The parallel fibrils, the number of layers, their orientation, the total thickness, as well as added substances such as lignin, help strengthen the cell wall.
PLASMODESMATA AND GAP JUNCTIONS
Plasma membrane connections, known as desmosomes, are thin cytoplasmic connections that often run from one plant cell to the next through pores in the cell walls. These pores, called plasmodesmata, facilitate intercellular exchange of materials such as sugars and amino acids (see Figure 2.5).
Gap junctions, sometimes called nexus junctions, are areas of low electrical resistance between animal cells across which electrical impulses, small ions, and molecules may pass. They seem to function similarly to the plasmodesmata that connect plant cells.
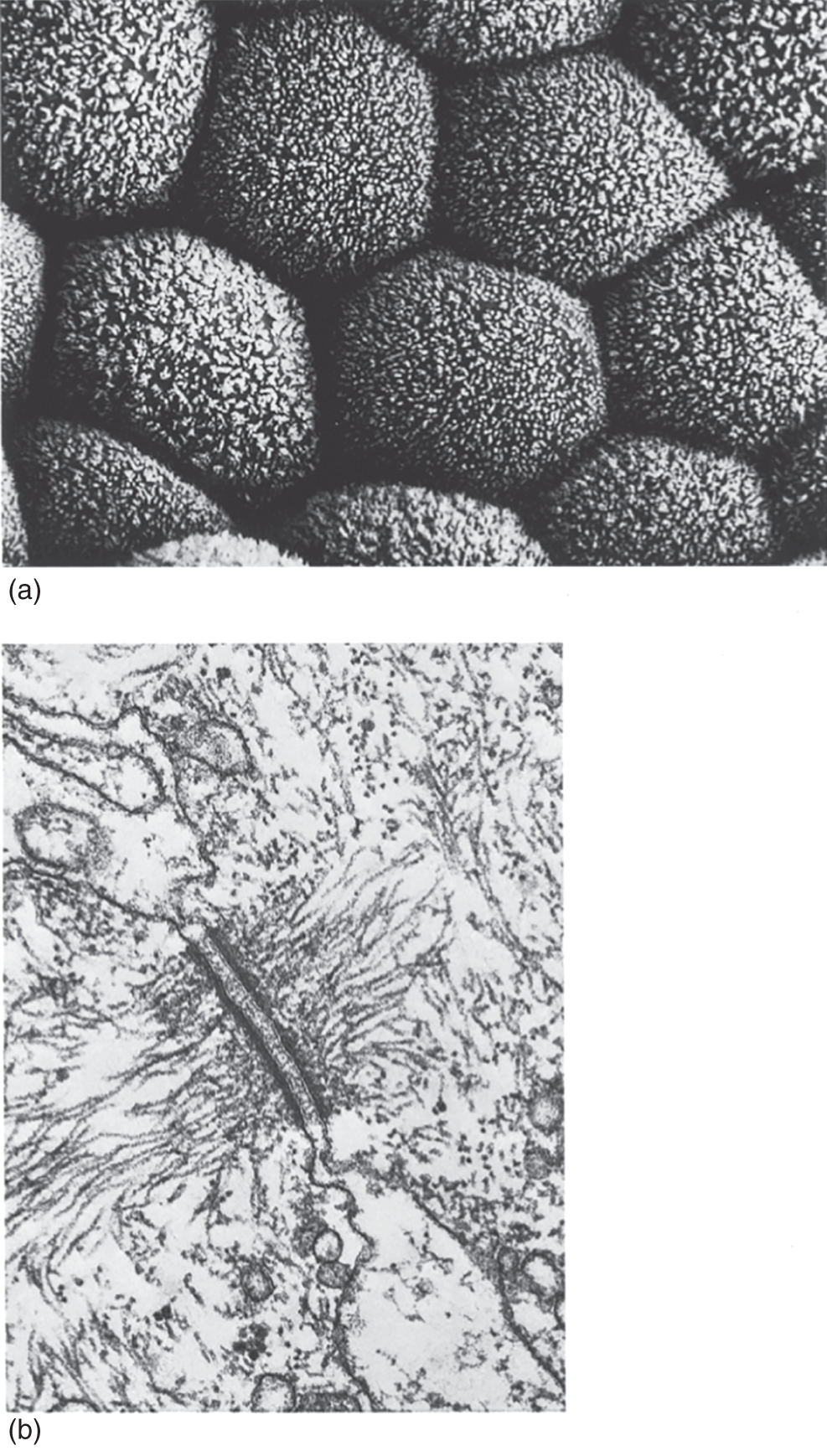
Figure 2.5 Cells have specialized junctions that facilitate the passage of materials across plasma membranes. Shown in (a) are the surfaces of sweat gland cells, and (b) desmosomes, which form below these surfaces in plasma membranes of adjacent cells.
When animal cells are in water with a low relative osmotic pressure, water will move from the area of low osmotic pressure into the cells with higher osmotic pressure, and they will ultimately burst due to the cells' distention, or turgor. Plant cells can usually withstand the swelling without bursting because their sturdy cell walls counterbalance the low external pressure.
CELLS' INTERNAL STRUCTURES
Nucleus
Relatively large, distinct, membrane-enclosed structures of several kinds are found in most kinds of cells, though not all. The notable exceptions are bacteria and cyanobacteria (often referred to interchangeably as blue-green bacteria or blue-green algae). The largest, most well-defined subcellular structure, or organelle, is the nucleus, unless even larger vacuoles are present. Vacuoles are large, fluid-filled spaces found in many cells, particularly plant cells. The nucleus contains the long, thin structures composed of deoxyribonucleic acid(DNA) and protein – the chromosomes. These structures contain genes, which are the individual units of information that inform the cell what to do and how and when to do it. All the instructions concerning the life processes of the cell emanate from these chromosomes (see Figure 2.6).
Nucleic acids are a major group of organic compounds that are composed of subunits called nucleotides, which contain carbon, hydrogen, oxygen, phosphorus, and nitrogen. It is the sequence of nucleotides that gives the specific nucleic acid its distinctive properties. DNA and ribonucleic acid(RNA) are two important nucleic acids found in cells. They are fundamental in the storage and transmission of genetic information that controls the cell's functions and interactions.
A dark area within the nucleus of most cells, seen when the cells are not dividing, is known as the nucleolus (see Figure 2.6). This area contains a high concentration of RNA and protein. Sometimes a nucleus contains several nucleoli, which are formed by a specific region on one chromosome called the nucleolar organizer.
All the nuclear structures are embedded in a viscous colloidal material, the nucleoplasm. The nucleus is bounded by a membrane, the nuclear membrane, which consists of two complete membranes similar in structure to the cell membrane. The nuclear membrane is very selective, restricting certain substances that readily pass through the cell membrane into the cytoplasm but are unable to pass through the nuclear membrane.
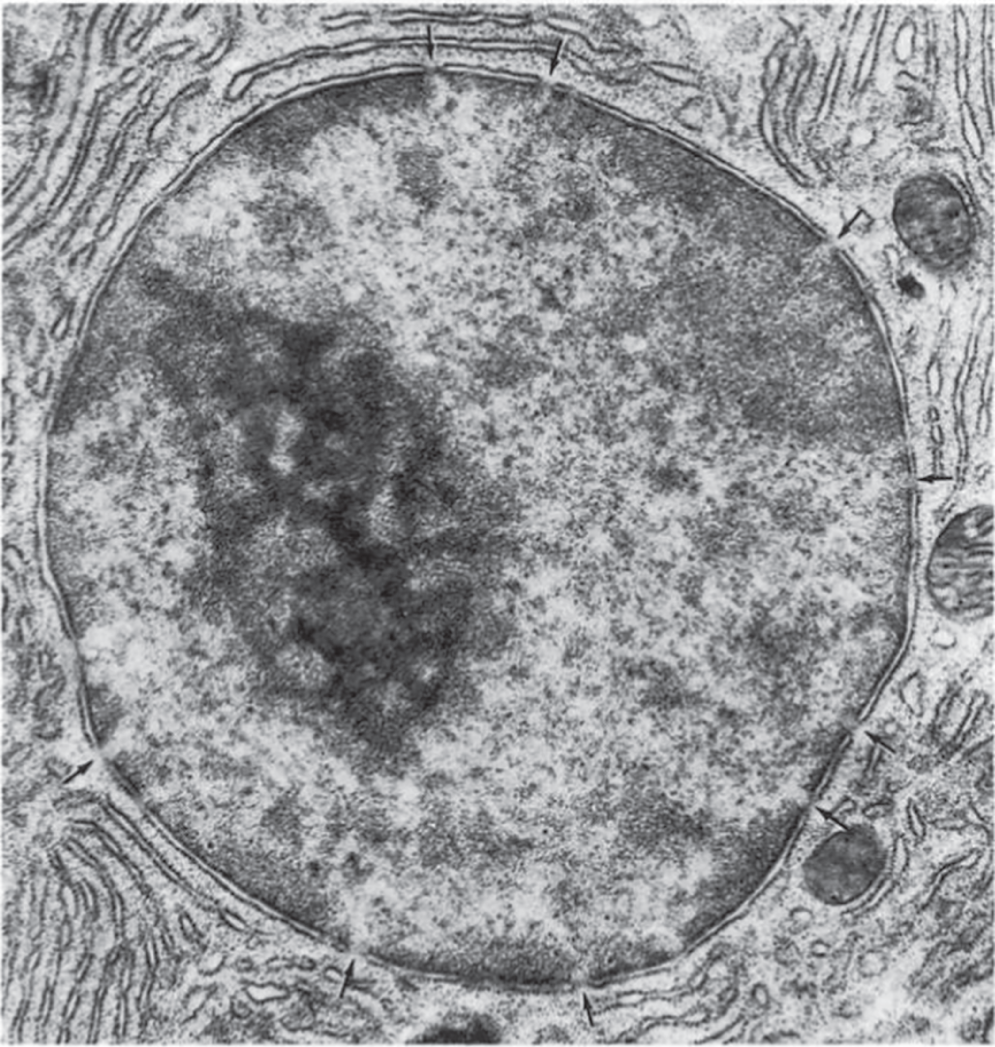
Figure 2.6 An electron micrograph of a nucleus. The arrows point to pores in the nuclear envelope's double membrane. The large dark area in the left portion of the nucleus is its nucleolus. Most of the other dark material inside the nucleus is chromatin.
The nuclear membrane is perforated by many large pores that facilitate the passage of substances in and out of the nucleus. Attached to the outside of the nuclear membrane are many thin membranes of endoplasmic reticulum(ER) that appear to be important in assisting the two-way flow of materials to and from the nucleus.
Endoplasmic Reticulum
The endoplasmic reticulum is a series of double-layered membranes found throughout the cytoplasm that provide surface area for enzymes to catalyze chemical reactions, particularly those leading to protein synthesis. The ER forms a multibranched channel between the outside of the cell and the nucleus, connecting the cell membrane and the nuclear membrane, thus expediting the transport of cellular materials.
Endoplasmic reticulum may be present with or without the outer surface lined with small particles called ribosomes, which are the actual sites of protein synthesis. Endoplasmic reticulum without ribosomes is known as smooth endoplasmic reticulum, and this is where some steroid hormones and other lipids are synthesized. The ER that is lined with ribosomes is referred to as rough endoplasmic reticulum. These terms, smooth and rough endoplasmic reticulum, relate to the structures' appearance in electron micrographs (see Figure 2.7).
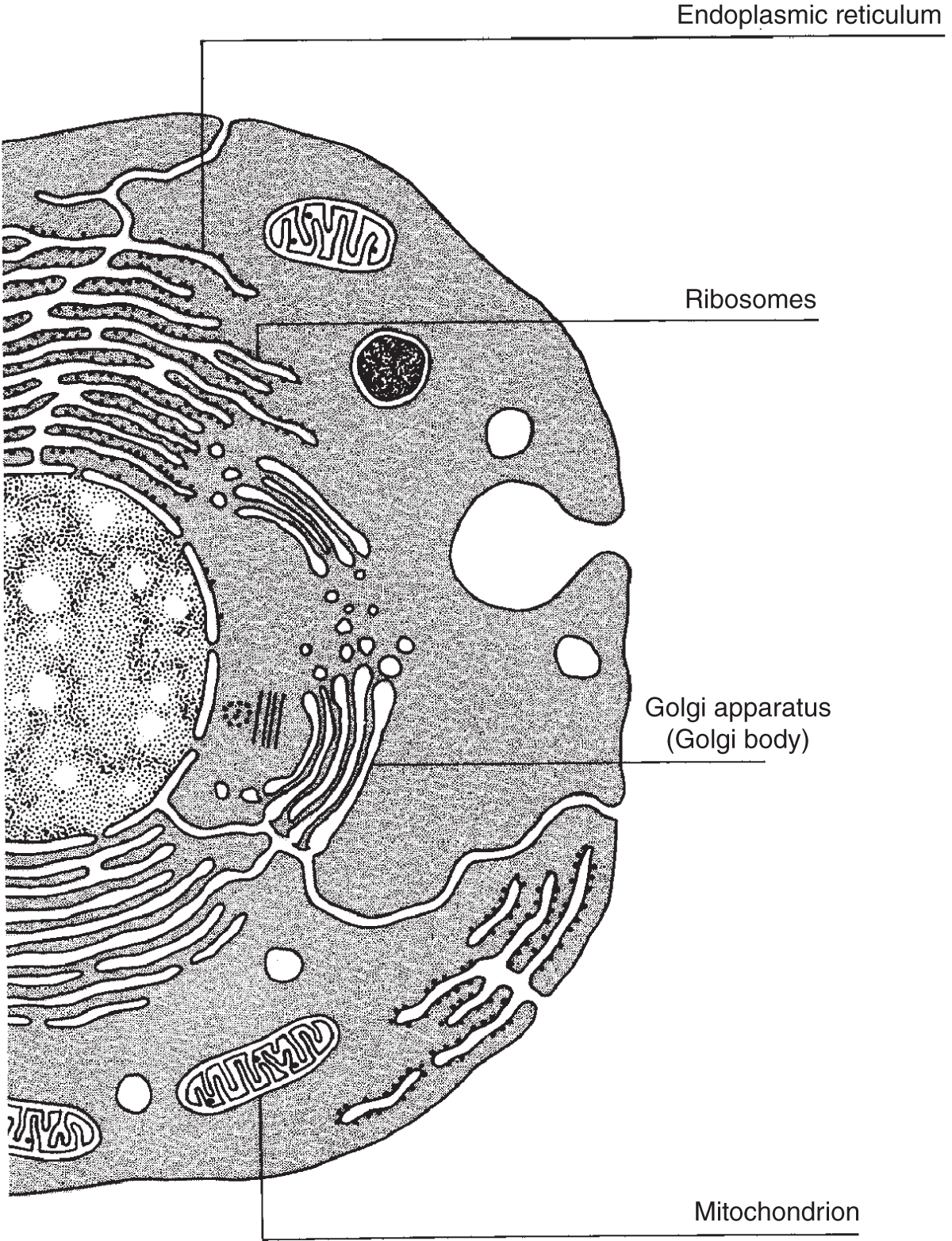
Figure 2.7 Here are four of the major organelles located in the animal cell as they would appear under an electron microscope. Note that both smooth and rough endoplasmic reticulum are shown.
Ribosomes
Ribosomes are tiny granules that can exist either in the cytoplasm as free-floating organelles or along the outer surface of the ER. Cells contain three main types of RNA, each synthesized from DNA. The RNA found in ribosomes that functions in protein synthesis is known as rRNA, which stands for ribosomal RNA.
Ribosomes contain high concentrations of rRNA and protein. In addition to existing singly, and attached to ER, some ribosomes occur in clumps or clusters, which are collectively known as polyribosomes or polysomes.
Golgi Apparatus
The Golgi apparatus (or Golgi body) is an organelle that looks like a stack of flattened, smooth, oval bowl-shaped membranes. It is thought that certain chemicals that are synthesized in the ribosomes move along the ER to the Golgi apparatus, where they are concentrated and stored until released by secretory vesicles produced by the outer portion of the Golgi apparatus.
In addition to functioning as an area where substances such as proteins pass through and are stored before being secreted, the Golgi apparatus is also involved in processing some proteins and carbohydrates, synthesizing them into more complex molecules by combining them into glycoproteins and glycosaminoglycan (previously called mucopolysaccharides), some of which are secreted in the form of mucus (see Figure 2.8).
Lysosome
A lysosome is a membrane-enclosed storage vesicle. The membrane is composed of a single layer, unlike the double-layered membrane of the nucleus. Lysosomes appear to arise from Golgi bodies. They contain strong digestive (hydrolytic) enzymes that act as the cell's digestive system, making it possible for the cell to process within its cytoplasm materials such as the particles taken in by pinocytosis or phagocytosis (defined earlier in this chapter under the heading “Movement of Large Particles Across Membranes”).
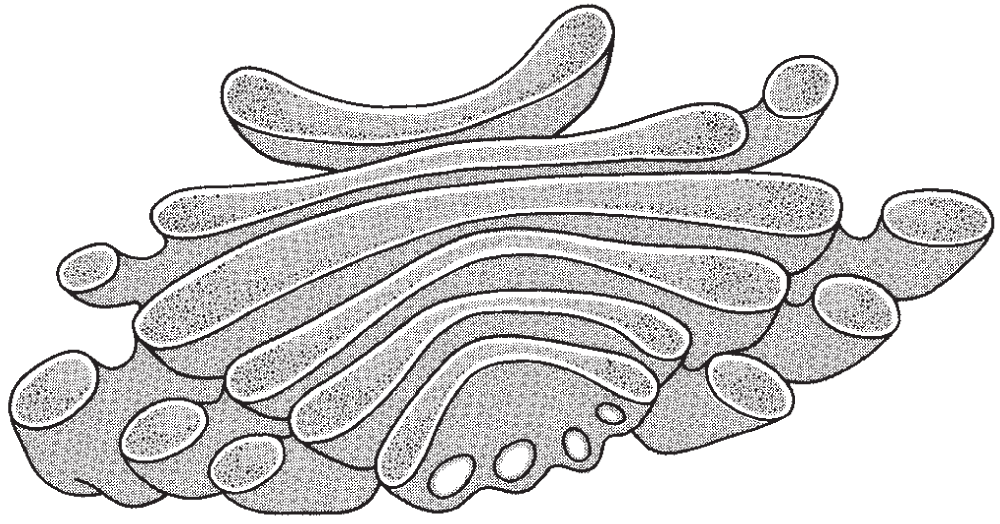
Figure 2.8 This cross-section diagram illustrates the Golgi apparatus, an organelle that provides capsules in which hormones and digestive enzymes are stored.
Lysosomes also break down old parts of the cell into the organic molecules from which they were constructed. These molecules are then reused by the cell. The importance of lysosomes has been established in studies concerning the metamorphosis of frog larvae. When tadpole tails shrink in size, their constituents are being broken down by lysosomes, and these organic molecules are then reabsorbed to fuel the growth of new structures such as the frog's budding arms and legs.
Mitochondria
Often referred to as the powerhouse of the cell, mitochondria supply the cell with chemical energy in the form of the high-energy molecule ATP. Slightly larger than the lysosomes, these double-membraned organelles (see Figure 2.9) are often the largest of the organelles in the cytoplasm, although plastids, vacuoles (described later), and the nucleus are also quite large in many cases.
Because they supply cells with energy, mitochondria are most abundant in cells with high-energy requirements, as in the growing root tip of a plant or in an animal's muscle cell. Each liver cell contains as many as 2,500 mitochondria.
The outer membrane separates the mitochondria from the cytoplasm, and the inner membrane has many inwardly directed folds, or cristae, that extend into a semifluid matrix. It is along the cristae and in the matrix where the energy-transfer enzyme systems that manufacture ATP are found. ATP is used in cell division, in muscle cell contraction, and as an energy source for many other cellular activities throughout living organisms.
When more sugar is available in a cell than it requires, fat droplets found next to many mitochondria will produce fat. Then when an energy deficit arises, the mitochondria will break down the fat droplets to yield energy. Excess fat droplets are transferred to specialized fat storage cells.
Unlike most other organelles, mitochondria contain DNA, RNA, and ribosomes, and they replicate themselves. It is now thought that mitochondria did not arise as a part of the cell but were originally independently living cells that were taken into larger cells and were able to survive in a jointly beneficial relationship (or symbiosis).
Mitochondria are often found in close proximity with a smaller organelle, the peroxisome, which may produce substrates for mitochondrial activity.
Plastids
Like mitochondria, plastids contain DNA, RNA, and ribosomes and are able to replicate themselves. Plastids are rather large cytoplasmic organelles that can be seen with a light microscope. They occur in most photosynthetic plant cells and also in photosynthetic protists (single-celled organisms such as some algae). Plastids are composed of two membrane layers. The outer membrane lacks folds and encloses the organelle, regulating the movement of materials between the cytoplasm and the organelle's interior. The inner membrane folds throughout the internal proteinaceous matrix known as the stroma. These internal membranes occur in sheets called lamellae, which often form stacks or grana; these stacks are connected by single lamellae.

Figure 2.9 (a) An artist's rendering of a mitochondrion; and (b) a labeled photomicrograph of a mitochondrion in cross section.
Different types of plastids have been recognized, and each appears to develop from a common structure or sometimes from one another. Those that are white and colorless and that store oils and protein granules, as well as energy-rich starch formed from simple sugars, are the leucoplasts. Plastids filled with starch are called amyloplasts. Plastids containing yellow, orange, or red pigments are called chromoplasts. These are found in flowers and fruits. Chloroplasts are the chromoplasts that – in addition to yellow, orange, or red pigments – also contain green pigment, or chlorophyll, which uses energy from sunlight to manufacture organic molecules such as sugar from inorganic molecules. In addition to chlorophyll, some of the other pigments, including carotenoids and xanthophylls, remain in temperate-zone leaves in the autumn after the green pigments have broken down. It is these pigments that account for the autumn colors (see Figure 2.10).
Vacuoles
Vacuoles are membrane-bound, fluid-filled spaces in both plant and animal cells, although they are most prominent in some plant cells (see, for instance, Figures 2.11 and 2.12). Much of the total volume of most plant cells is composed of a single vacuole surrounded by its membrane, the tonoplast. This membrane is also called the vacuolar membrane. This separates vacuoles inside plant cells. The liquid inside the plant vacuole is called cell sap, which is mostly water with some dissolved substances. Cell sap is usually hyperosmotic relative to the external medium surrounding it, so in times of need, when available, water will move in through the vacuole's membrane via osmosis, keeping the cell turgid. Vacuoles often contain a high concentration of soluble organic compounds such as sugars, amino acids, some proteins, and several pigments. Many plant species store toxic chemicals in the vacuoles that do not harm the cell. If an animal eats the plant, however, the contents of the ruptured vacuoles can have an adverse effect on the animal.
Different kinds of vacuoles have different functions. Some single-celled organisms have contractile vacuoles, which are important in expelling excess water and waste from the cell. Many cells have vacuoles or vesicles that are formed when the cell membrane takes in small particles by phagocytosis or pinocytosis, which may then be digested by enzymes from the lysosomes.
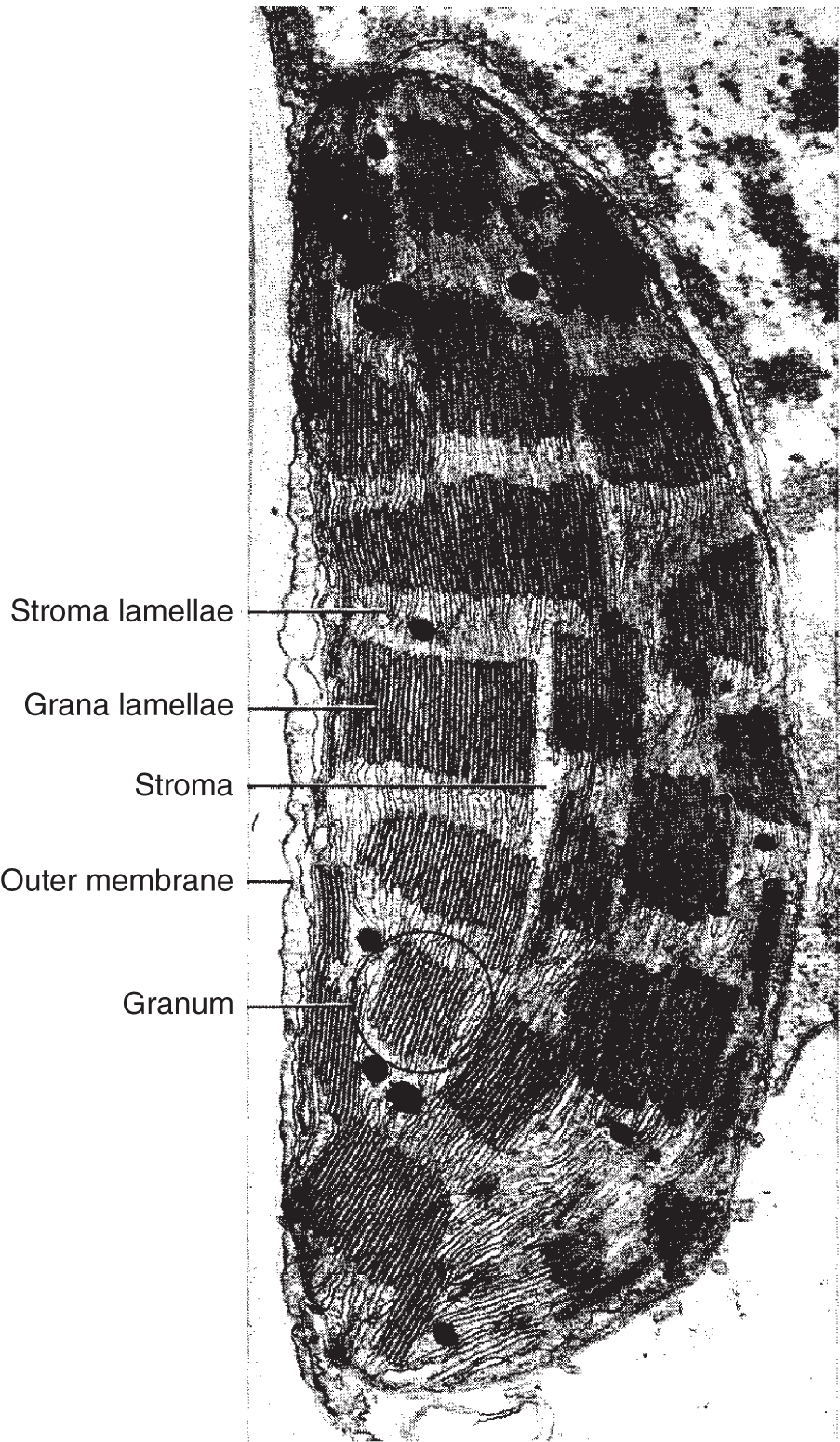
Figure 2.10 A chloroplast with structures labeled.
Microfilaments
Within a cell's cytoplasm are many long protein fibers arranged into microfilaments that are only about 6 nanometers in diameter. Microfilaments function in the maintenance of cell structure and movement. Depending on the local conditions in the cell, microfilaments can exist either as individual subunits or together in strings of subunits. Microfilaments are believed to be important components of the contractile filaments that exist in some cells. They may be associated with cytoplasmic streaming, a constant movement of cytoplasm in the cell that is evident in many plant cells under the light microscope. Microfilaments may also be involved in cell movement, and when in a more complex form, they may be associated with the contraction of some muscle cells.
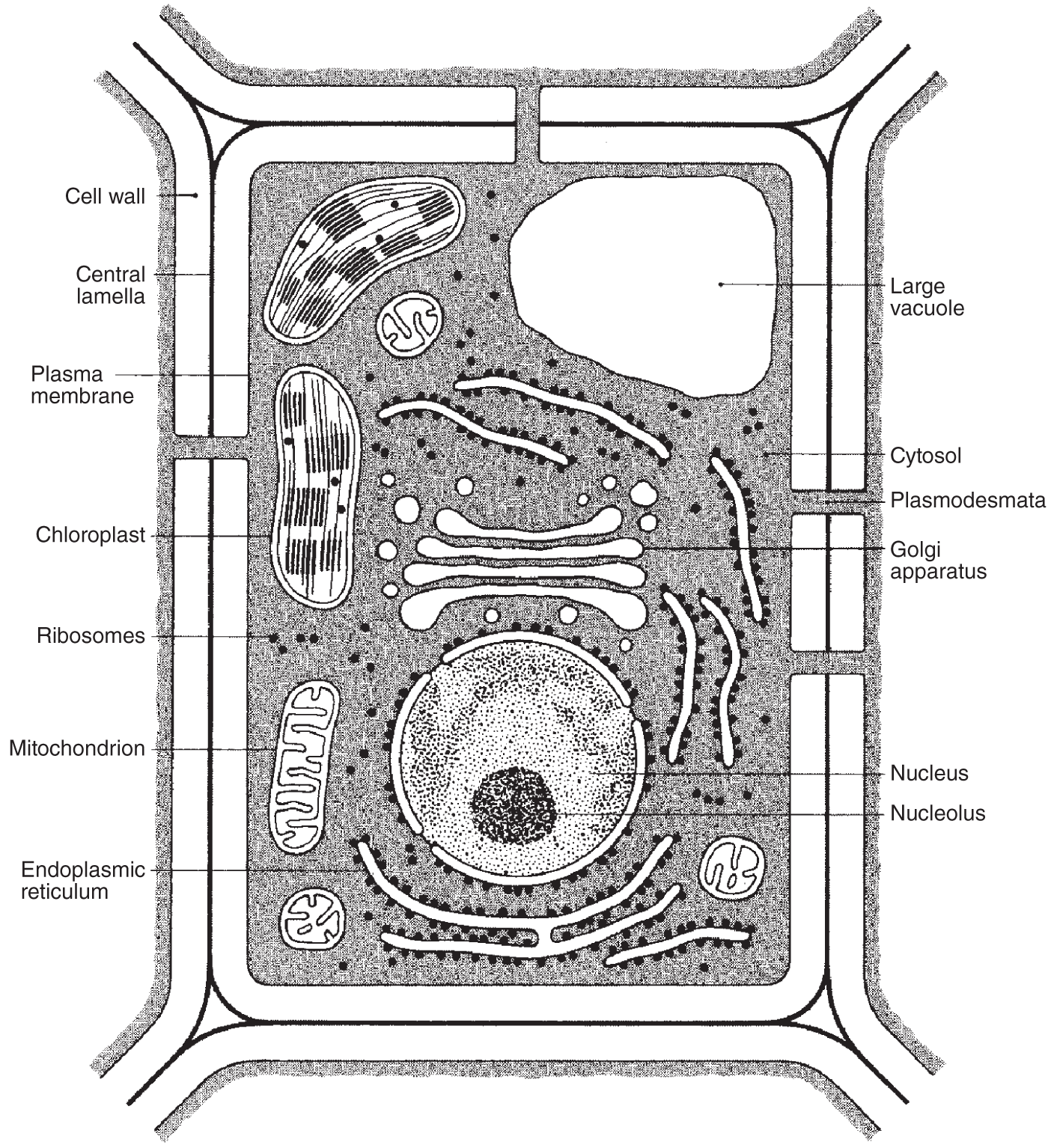
Figure 2.11 A generalized plant cell with its structures labeled.
Microtubules
Also in the cytoplasm are microtubules, which differ from microfilaments in many respects. The outside diameter of each microtubule is considerably larger than that of a microfilament, ranging from 20 to 25 nanometers (one million nanometers equals one millimeter; one billion nanometers equals one meter). Furthermore, microtubules are hollow and vary in length from less than 200 nanometers to 25,000 nanometers. Microtubules also differ from microfilaments in terms of their chemical makeup.
Sometimes microtubules act as tracts along which the cell's organelles move. Microtubules orient themselves toward each pole of a cell, and chromosomes move along them during cell division. A distinctive internal microtubular arrangement also determines the arrangement of a cell's cilia, flagella, and sperm tails, which are discussed below.
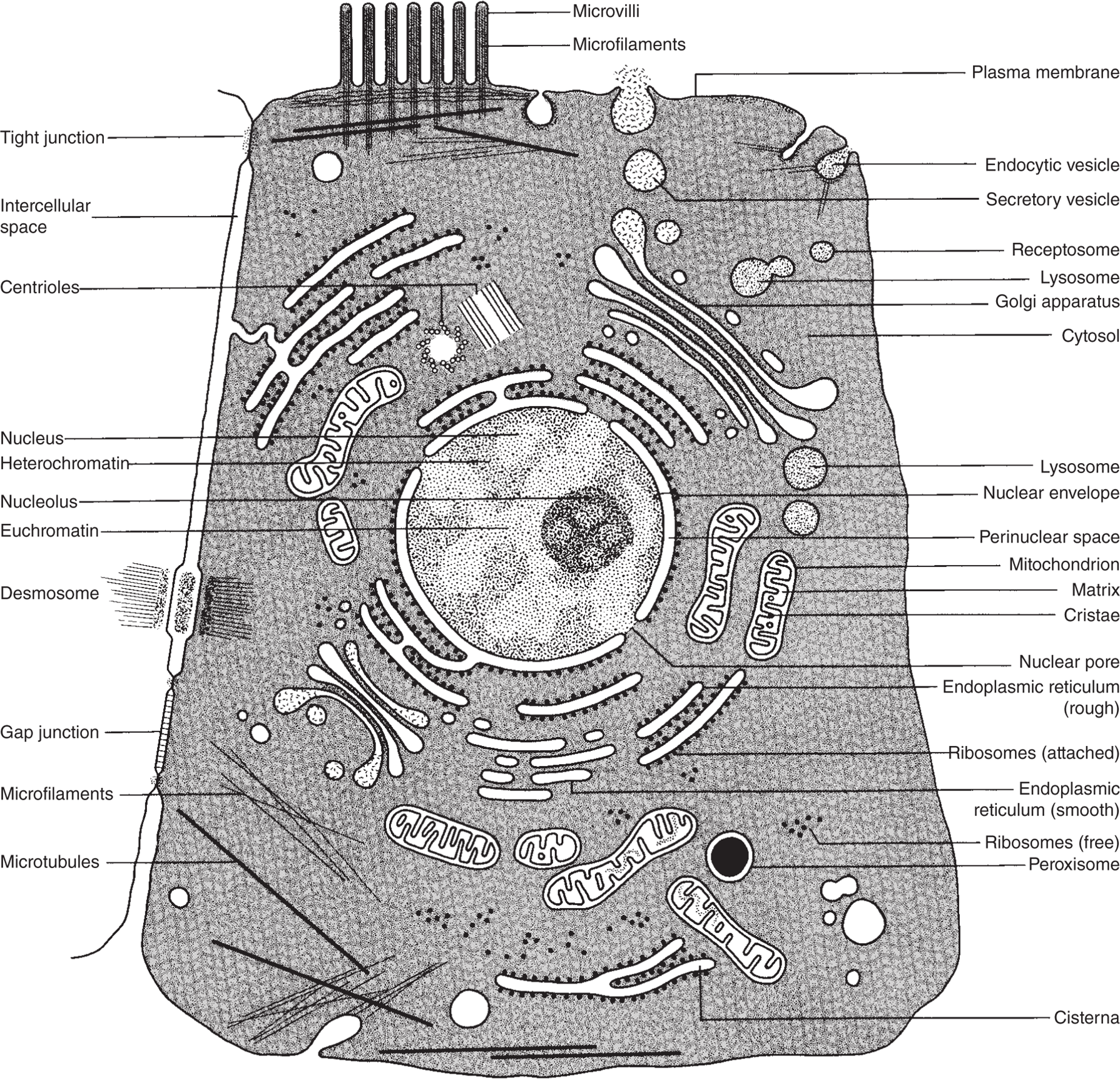
Figure 2.12 Composite animal cell.
Cilia and Flagella
Cilia and flagella (singular: cilium and flagellum) are long, thin organelles that project from the surface of some cells and have the capacity to beat back and forth or in a corkscrew fashion, moving the cell through fluids or moving fluids past a stationary cell. They appear on some protists, on certain lower plants, and on some animals. Many eukaryotic organisms have flagellated sperm cells. Eukaryotic and prokaryotic bacterial flagella differ structurally and mechanically.
The difference between cilia and flagella is mostly a matter of length (though not diameter), and sometimes they are differentiated according to number; many short ones together are called cilia, while an isolated long one is usually a flagellum.
The function of these organelles is primarily locomotory, to propel or pull a cell through liquids. Some protists also use cilia to move particles into their gullet – a part of the cell membrane that is similar to a mouth. Multicellular organisms commonly have ciliated cells lining internal passageways. Their constant motion helps move materials through tubes and ducts in the preferred direction.
In all types of cells except bacteria and blue-green algae, cilia and flagella are cytoplasmic outgrowths from the surface of the cell, surrounded by extensions of the plasma membrane. At the base of the long cilium or flagellum is the basal body, which supports and gives rise to the rest of the long stalk. It consists of nine triplets of microtubules arranged in a circle. The basal bodies may be where the protein subunits assemble to form the microtubules that extend through the rest of the long structures, which are themselves composed of a circle of nine pairs of microtubules with another pair at their center. This 9 + 2 arrangement is characteristic of all cellular cilia and flagella except those found among bacteria and blue-green algae.
Centrioles
Basal bodies and centrioles are structurally identical, although the basal bodies are found at the bases of cilia and flagella of all cells, while centrioles are found only in animal cells, generally located near the nucleus. Centrioles occur in pairs. Like basal bodies, they are composed of nine sets of triplets, all arranged lengthwise, creating a cylindrical structure.
Not all cells contain centrioles, but most animal cells have them, and they are always composed of the same typical pattern: a cylinder formed by nine sets of triplets that, when cut in a transverse section (cross section), looks like a pinwheel (see Figure 2.13).
Besides playing a role in the formation of microtubules, centrioles become active during cell reproduction. During interphase, both pairs of centrioles are located just outside the nucleus, arranged at right angles to each other. As mitosis (or meiosis) begins, they move apart and organize the mitotic apparatus known as the spindle, which is composed of microtubules extending from pole to pole through the cell's interior. It is along these microtubules that the chromosomes align at metaphase, then moving apart during the remaining stages of cell division. Toward the end of cell division (during telophase), the centrioles replicate, so each daughter cell in turn has two pairs of centrioles located just outside the nucleus. For a more comprehensive discussion of cell division, see Chapter 3.
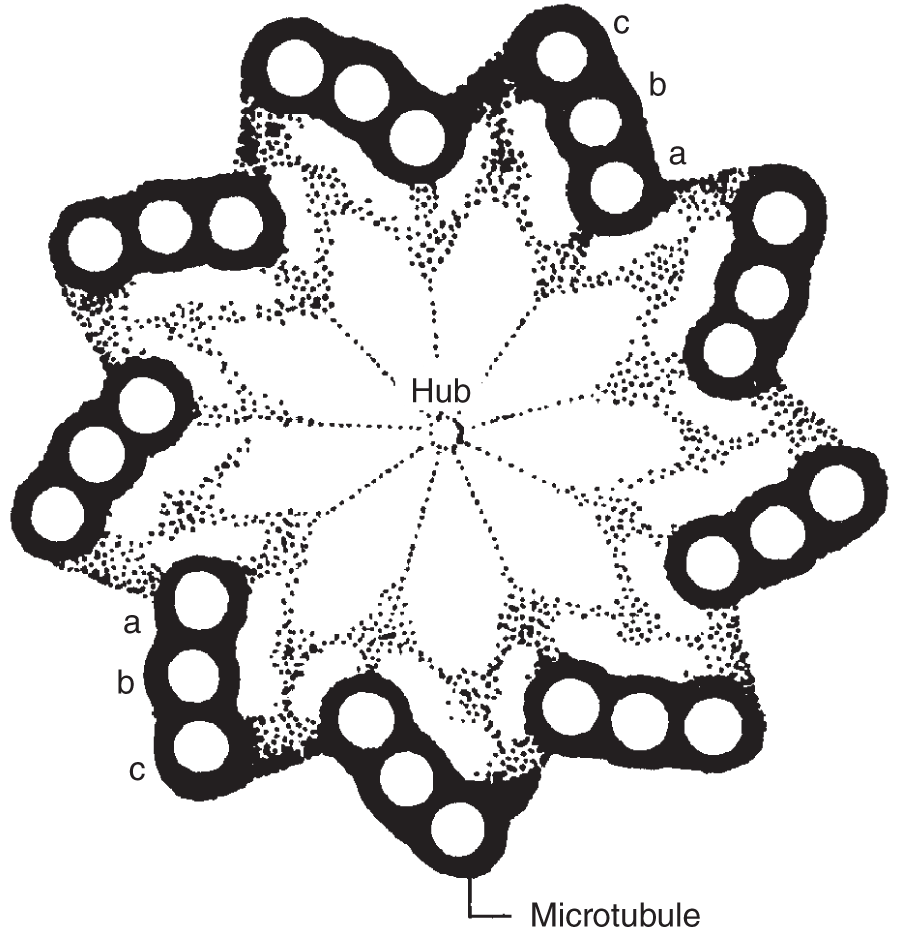
Figure 2.13 Cross section of a centriole. This drawing shows the nine triplets of microtubules, which turn in unison, creating the corkscrew or whip-like motion of cilia and flagella.
Eukaryotic and Prokaryotic Cells
The differences between cells of bacteria and blue-green algae and those of most other organisms are customarily cited to distinguish between two distinct cellular types: eukaryotic cells and prokaryotic cells.
Eukaryotic Cells
Most of the preceding part of this chapter concerns eukaryotic cells. Eukaryotes are those plants and animals with eukaryotic cells. These include all the protists, fungi, plants, and animals. Their cells have a nucleus enclosed within a nuclear membrane, inside of which are located the chromosomes – the long, filamentous structures along which occur the units of inheritance, called genes. These chromosomes are composed of nucleic acids and proteins.
Eukaryotic cells also contain mitochondria, Golgi bodies (or Golgi apparatus), ER, and most of the other organelles so far described. When present in these cells, chlorophyll is located in chloroplasts. And if these cells have cilia and/or flagella, they always possess the previously described 9 + 2 construction.
All the contents of eukaryotic cells are enclosed within a plasma membrane, and in some cases, as with many plant cells, the plasma membrane is also enclosed within a cell wall.
Prokaryotic Cells
Prokaryotic cells are thought to be more primitive than eukaryotic cells. Bacteria and blue-green algae are prokaryotes, which is to say their cells are prokaryotic. These cells do not have a nuclear membrane; instead, they have a nuclear area known as the nucleoid that in most cases contains a single circular chromosome. In prokaryotic cells, the chromosomes are composed entirely of nucleic acids (without protein). Their ribosomes are also structurally distinct from those in eukaryotic cells. When flagella-like structures are present, they don't have the 9 + 2 fibrillar structure. And prokaryotic cells contain no mitochondria, no Golgi apparatus, no lysosomes, and no vacuoles.
In addition, prokaryotic cells have cell walls containing muramic acid, a characteristic not found among eukaryotic cells. In many prokaryotic cells appears an infolding of the plasma membrane known as a mesosome, which may be involved in the production of new cell wall material following cell division, and which may be associated with the replication of DNA during cell division. The mesosome may also help break down food molecules to supply the cell with energy.
Most photosynthetic prokaryotes contain photosynthetic membranes and chlorophyll, but these materials aren't enclosed in chloroplasts.
KEY TERMS
| active transport | cell theory | chlorophyll |
| amino acids | cell walls | chloroplasts |
| amyloplasts | cells | chromoplasts |
| basal body | cellulose | chromosomes |
| carotenoids | cellulose microfibrils | cilia |
| carrier molecules | central lamella | colloid |
| cell membrane | centrioles | concentration gradient |
| cell sap | chemical homeostasis | contact X-ray microscopy |
| contractile vacuoles | inner membrane | passive transport |
| cristae | isosmotic | pectin |
| cytologists | isotonic | peroxisome |
| cytoplasm | lamellae | phagocytosis |
| cytoplasmic streaming | Leeuwenhoek, | pinocytosis |
| desmosomes | Antonie van | plasma membrane |
| deoxyribonucleic | leucoplasts | plasmodesmata |
| acid (DNA) | light microscope | plastids |
| diffusion | lignin | polyribosomes |
| dissecting microscopes | lipid soluble | polysaccharides |
| electron microscope | lysosome | polysomes |
| endocytosis | magnification | primary cell wall |
| endoplasmic reticulum (ER) | matrix | prokaryotes |
| equilibrium | mesosome | prokaryotic cells |
| eukaryotic cells | microfilaments | proteins |
| exocytosis | microscope | resolution |
| facilitated diffusion | microtubules | ribonucleic acid (RNA) |
| fat droplets | middle lamella | ribosomal RNA (rRNA) |
| fat soluble | mitochondria | ribosomes |
| flagella | mucopolysaccharides | rough endoplasmic |
| gap junctions | multicellular organism | reticulum |
| gel | nexus junctions | scanning electron |
| genes | nuclear membrane | microscope |
| glucosaminoglycan | nucleic acids | scanning tunneling |
| glucose | nucleoid | microscope |
| glycoproteins | nucleolar organizer | Schleiden, Matthias |
| Golgi apparatus | nucleolus | Schwann, Theodor |
| (Golgi body) | nucleoplasm | secondary cell wal |
| grana | nucleotides | semipermeable |
| Hooke, Robert | nucleus | membrane |
| hyperosmotic | organelle | smooth endoplasmic |
| hypertonic | osmosis | reticulum |
| hypoosmotic | osmotic pressure | simple sugars |
| hypotonic | outer membrane |
SELF-TEST
Multiple-Choice Questions
Microscope, Cells, Cell Theory
- During the seventeenth century, the first light microscopes were constructed by __________.
- Hooke
- DuPont
- Scopes
- Pierson
- Leeuwenhoek
- The study of cells is called __________.
- herpetology
- ornithology
- simology
- cytology
- tautology
- Illuminating specimens under the light microscope with __________ light was found to give considerably more resolution than when using other colors.
- red
- orange
- blue
- green
- yellow
- With a transmission electron microscope, a beam of electrons with considerably __________ than visible light is passed through a thinly sliced specimen.
- longer wavelengths
- shorter wavelengths
- brighter light
- stronger X-rays
- stronger cosmic rays
- Of the following, the most recent advance in microscope technology that enables scientists to observe the most highly detailed views of individual living cells is called __________.
- transmission electron microscopy
- light microscopy
- scanning electron microscopy
- contact X-ray microscopy
- gamma ray microscopy
- In 1665 __________ described having seen what he called cells in a piece of cork.
- Spallanzini
- Malthus
- Hooke
- Leeuwenhoek
- Marvin
- All following statements concerning cell theory except one are correct. Circle the letter preceding the incorrect answer.
- Cells are the fundamental units of life.
- All cells have cell walls.
- Cells are the smallest entities that can be called living.
- All organisms are made up of one or more cells.
- Cells arise only from the division of other cells.
Cell Structure and Cell Size, Cytoplasm and Nucleoplasm, Plasma Membrane
- A cell becomes too large to maintain a stable internal environment when
- its surface area becomes too small
- its volume becomes too small
- its surface area gets too big
- its volume gets too big
- its surface area to volume ratio becomes too great
- It has been stated that the larger the cell, the __________ to maintain control over the entire internal environment.
- easier it is for the nuclear material
- harder it is for the nuclear material
- easier it is for the chloroplast
- harder it is for the chloroplast
- easier it is for the mitochondria
- The interior of a living cell is made up of a __________.
- complex solution
- heterogeneous colloid
- solid throughout
- a and b
- b and c
- A colloid may become a gel, depending on the __________.
- temperature
- pH
- salt concentration
- pressure
- all of the above
Movement Through the Cell Membrane, Diffusion, Osmosis, Facilitated Diffusion
- The plasma membrane provides the following to the cell:
- protection
- shape
- strength
- a structure that helps regulate the flow of materials going in and out of the cell
- all of the above
- When energy is used to move molecules from one location to another, it is termed __________.
- passive transport
- colloidal collision
- gel state
- passive homeostasis
- active transport
- The net movement of particles from regions of higher concentration to regions of lower concentration of that substance is called __________.
- homeostasis
- active ambiance
- homeostatic collision
- diffusion
- gradual constance
- The particles moving away from the area of greatest concentration of that substance will produce a __________.
- gel state
- colloidal collision
- diffusionary divergence
- gradual homeostasis
- concentration gradient
- A partially or differentially permeable membrane is called a __________.
- diffusionary membrane
- semihomeostatic membrane
- active membrane
- facilitated membrane
- semipermeable membrane
- The pressure exerted by dissolved particles in a solution or a colloid that pulls water across a semipermeable membrane is known as __________.
- isosmosis
- hyperosmosis
- osmosis
- hypoosmosis
- osmotic pressure
- When there is a semipermeable membrane separating two solutions of pure water, the two sides are at __________.
- hyperequilibrium
- hypoequilibrium
- osmotic diffusion
- facilitated osmoequilibrium
- equilibrium
- When the osmotic pressure is the same on both sides of a semipermeable membrane, each solution is said to be __________.
- isosmotic
- hyperosmotic
- hypoosmotic
- osmotic
- none of the above
- Plasma membranes contain __________ that help move certain substances in and out of the cell.
- organelles
- nuclei
- nucleoli
- prions
- carrier molecules
Active Transport, Movement of Large Particles Across Membranes, Plant Cell Wall
- To move substances such as glucose across a cell membrane against a concentration gradient __________.
- no energy needs to be expended
- is called diffusion
- is called osmosis
- energy has to be expended
- none of the above
- The most extensively studied active transport systems use the high-energy molecule __________ to maintain the systems.
- ATP
- NADPH2
- NAD
- NADH2
- none of the above
- The capacity of the plasma membrane to fuse with or engulf particulate matter and then pinch off and move out of the cell is known as __________.
- endocytosis
- exocytosis
- endothelial phagocytosis
- exocrine pinocytosis
- none of the above
- When liquids or macromolecules are engulfed and moved into the cell, this kind of endocytosis is known as __________.
- exocytosis
- endothelial phagocytosis
- pinocytosis
- phagocytosis
- none of the above
- Both plant and animal cells possess a plasma membrane, but only plant cells have a __________.
- mesothelium
- endothelium
- cell wall
- nucleus
- nucleolus
- The cell wall is a product of the cellular protoplasm and is primarily composed of the complex polysaccharide __________ and other related compounds.
- starch
- glucose
- globular protein
- cellulose
- nylon
- Where two cell walls come in contact, another layer is laid down between the two cells, which is called the middle lamella. This continuous layer between the cells is usually composed of __________.
- starch
- pectin
- lignin
- globular protein
- cellulose
- The complex polysaccharide, usually in the form of calcium pectate, that binds plant cells together, is known as __________.
- cellulose
- lignin
- beta kerotene
- anthracnose
- pectin
- The long thread-like structures that make up much of the cell wall, and that are laid down in a loose network, are known as __________.
- myofibrils
- myoglobin
- fibrinogen
- cellulose microfibrils
- pectin
Plasmodesmata and Gap Junctions, Nucleus, and Endoplasmic Reticulum
- The connected network of similar membranes that connect structures within the cell is called (the) __________.
- carrier molecule
- endoplasmic reticulum
- globular protein
- adenosine triphosphate
- gamma globulin
- The thin protoplasmic connections that often run from one plant cell to the next through pores in the cell walls are known as __________.
- plasmids
- plasmodesmids
- gap junctions
- endoplasmic reticulum
- plasmodesmata
- When animal cells are in water with a low osmotic pressure relative to their interior, the water will move from the area of low osmotic pressure into the cells with the higher osmotic pressure, and ultimately the cells will burst due to their distension, or __________.
- plasma
- desmata
- turgor
- nexus
- none of the above
- Well-defined subcellular structures that are enveloped in membranes are called __________.
- bacteria
- blue-green algae
- prions
- nucleotides
- organelles
- The large fluid-filled spaces enveloped in membranes that are found in many cells, particularly in plant cells, are __________.
- chromosomes
- vacuoles
- nuclei
- chromatids
- nucleoli
- The large organelle containing chromosomes is the __________.
- nucleus
- vacuole
- chromatid
- lysosome
- endoplasmic reticulum
- It is the sequence of __________ that gives the specific nucleic acid its distinctive properties.
- genes
- nuclei
- chromosomes
- nucleotides
- nucleoli
- When the cell is not dividing, there is a conspicuous dark area within the nucleus, the _____, that contains a high concentration of ribonucleic acid and protein.
- endoplasmic reticulum
- Golgi body
- vacuole
- lysosome
- nucleolus
- All the nuclear structures are embedded in a viscous colloidal material, the __________.
- RNA
- DNA
- cytoplasm
- protoplasm
- nucleoplasm
- The nucleus is bounded by the __________, which consists of two complete membranes similar in structure to the plasma membrane.
- nuclear membrane
- nuclear wall
- nucleolar organizer
- nucleoplasm
- cell wall
- Attached to the outside of the nuclear membrane are many other membranes of __________ that appear to be important in assisting the two-way flow of materials to and from the nucleus.
- cell wall
- endoplasmic reticulum
- endothelium
- nuclear organizer
- cell membrane
- The __________ is a series of double-layered membranes found throughout the cytoplasm that greatly increases the surface area within the cytoplasm, expanding the area available for enzymes involved in chemical reactions, particularly those leading to protein synthesis.
- endoplasmic reticulum
- primary cell wall
- cell wall
- cell membrane
- plasma membrane
- Endoplasmic reticulum may occur with and without being lined with small particles, the __________, which are the actual sites of protein synthesis.
- ribosomes
- lignin
- pectin
- endothelium
- nuclei
Ribosomes, Golgi Apparatus, Lysosomes, Mitochondria, and Plastids
- Besides being found in cells singly as well as on endoplasmic reticulum, some ribosomes occur in clumps or clusters collectively known as __________.
- polysomes
- polyribosomes
- polychromosomes
- a and b
- a, b, and c
- Certain __________ synthesized in the ribosomes move along the endoplasmic reticulum to the __________, where they are concentrated.
- plastids, lysosomes
- peroxisomes, autosomes
- glycoproteins, mesosomes
- polysaccharides, mesosomes
- chemicals, Golgi apparatus
- Chemicals that are concentrated and stored in the Golgi apparatus are later released from the cell by secretory vesicles that are produced by the outer portion of the Golgi apparatus when it moves to the __________.
- cell surface
- lysosome
- mesosome
- autosome
- ribosome
- Some simple sugars may be modified in the Golgi apparatus, synthesizing more complex polysaccharides that can then be attached to proteins forming __________ and __________ that can be used for mucus secretion.
- starch, celluloproteins
- glycogen, mucoproteins
- photoglycogen, mucoglycogen
- glycopolysaccharides, glucopolysaccharides
- glycoproteins, mucopolysaccharides
- __________contain strong digestive enzymes that act as the cell's digestive system, making it possible for the cell to process within its cytoplasm, particles taken in by pinocytosis or phagocytosis.
- peroxisomes
- lysosomes
- mesosomes
- autosomes
- ribosomes
- Often referred to as the powerhouse of the cell, __________ supply the cell with the chemical energy in the form of the high-energy molecule adenosine triphosphate.
- mitochondria
- vacuoles
- endoplasmic reticulum
- Golgi apparatus
- mesosomes
- The inner membranes of the mitochondria have many inwardly directed folds, the __________, which extend into a semifluid matrix.
- cristae
- grana
- thylakoids
- lamellae
- stroma
- In plastids, stacks of lamellae are the __________.
- stroma
- amyloplasts
- chloroplasts
- leucoplasts
- grana
- The many double membranes found in plastids are called __________.
- stroma
- grana
- lamellae
- amyloplasts
- leucoplasts
- The proteinaceous matrix inside plastids is known as the __________.
- stroma
- grana
- amyloplasts
- leucoplasts
- thylakoids
- Plastids filled with starch are called __________.
- leucoplasts
- amyloplasts
- chromoplasts
- leucocytes
- granulocytes
- Most plastids that contain yellow, orange, or red pigments are called __________.
- leucoplasts
- amyloplasts
- chromoplasts
- leucocytes
- granulocytes
- The chromoplasts that contain green pigment are known as __________.
- leucoplasts
- amyloplasts
- chromoplasts
- chloroplasts
- carotenoplasts
- In addition to chlorophyll, chloroplasts also contain other pigments, particularly the yellow and orange __________.
- amylophyll
- leucophyll
- rohophyll
- chromophyll
- carotenoids
Vacuoles, Microfilaments, Microtubules, Cilia and Flagella, Centrioles, and Eukaryotic and Prokaryotic Cells
- Much of the total volume of most plant cells is comprised of a single __________.
- vacuole
- tonoplast
- ribosome
- autosome
- macrotubule
- Vacuoles are surrounded by a membrane known as the __________.
- microtubule
- macrotubule
- microfilament
- tonoplast
- amyloplast
- The liquid inside the vacuole is called __________.
- cell sap
- tonoplasm
- vacuoplasm
- a and b
- a and c
- Membrane-bound fluid-filled spaces in plant and animal cells, which in some plant cells comprise much of the total cell volume, are __________.
- tonoplasts
- vacuoles
- ribosomes
- mesosomes
- autosomes
- There are different kinds of vacuoles with various functions. Some single-celled organisms have __________ that are important in expelling excess water and waste from the cell.
- tonoplasts
- ribosomes
- autosomes
- contractile vacuoles
- mesosomes
- Within a cell's cytoplasm are many long, thin protein fibers that are only about six nanometers in diameter. Depending on the local conditions in the cell, these __________ can exist either as subunits or as filaments.
- cilia
- microfilaments
- flagella
- basal bodies
- microtubules
- The constant movement of cytoplasm in the cell is called __________.
- cytoplasmic streaming
- ciliar movement
- flagellar movement
- microtubular movement
- microfilamentary streaming
- Cilia and flagella are cytoplasmic outgrowths from the surface of the cell, surrounded by extensions of the cell membrane. The base of the long cilium or flagellum is the __________.
- centriole
- microtubule
- microfilament
- basal body
- blue-green algae
- During interphase in animal cells, but not in most plant cells, in a special region of cytoplasm just outside the nucleus are two cylindrical bodies oriented at right angles to each other that move apart and organize the mitotic apparatus of the dividing cell. These organelles are the __________.
- basal bodies
- microtubules
- microfilaments
- centrioles
- cilia
ANSWERS
- e
- d
- c
- b
- d
- c
- b
- e
- b
- d
- e
- e
- e
- d
- e
- e
- e
- e
- a
- e
- d
- a
- b
- c
- c
- d
- b
- e
- d
- b
- e
- c
- e
- b
- a
- d
- e
- e
- a
- b
- a
- a
- d
- e
- a
- e
- b
- a
- a
- e
- c
- a
- b
- c
- d
- e
- a
- d
- a
- b
- d
- b
- a
- d
- d
Questions to Think About
- Explain the events that made it possible to observe and understand cell structure and function.
- When cells were first described, what was observed during this initial discovery?
- Describe what is now known as cell theory.
- Why are most cells small?
- Define cytoplasm and nucleoplasm.
- Compare and contrast the function of a solution and a colloid in cell structure.
- What is a cell membrane?
- How do materials move through a cell membrane?
- What are the possible dynamics of cell contents when the osmotic pressure within the cell is not equal to that outside the cell?
- Compare and contrast passive transport and active transport.
- How do materials move between cells?
- Describe the nucleus: what is it?; where is it?; what is in it?; and what is its function?
- Give a brief description of each of the following structures:
- endoplasmic reticulum
- ribosome
- Golgi apparatus
- lysosome
- mitochondrion
- fat droplet
- plastid
- vacuole
- microfilament
- microtubule
- cilium
- flagellum
- centriole
- Compare and contrast eukaryotic and prokaryotic cells.