11
Internal Transport: Plants and Invertebrates
Chemical reactions of all living cells occur in an aqueous environment, which explains why the materials within metabolically active cells are suspended or dissolved in water. In addition to the aqueous internal environment, the cells of many plants and animals are bathed in a nutritious extracellular environment, eliminating the need for specialized systems to move substances to and from these cells. The requirements of larger organisms, however, may differ considerably. Most large plants and animals have an internal transportation system that suits their particular life history. In general, the larger or more mobile the animal, the more complex and faster moving is the internal transport capability.
Both plants and animals have tubular transport systems for distribution of materials within their bodies. Most plants have two major pathways for internal transport: the phloem, which carries carbohydrates, and the xylem, which carries water and ions. Many animals have a system of arteries, veins, and lymphatic vessels, which carry blood and lymph, the protein-containing fluid that escapes from the blood capillaries. These transport systems are described in more detail below, after a discussion of some organisms that do without such special transport systems.
ORGANISMS WITHOUT INTERNAL TRANSPORT SYSTEMS
The internal contents of many small, relatively simple organisms can be moved around without the complex of internal structures such as tubes, vessels, and special mechanisms observed in most higher organisms. Single-celled organisms such as bacteria and protists rely on diffusion as one of their major transport systems. It may or may not be fast, but, with such small distances involved, the random distribution resulting from diffusion of most microscopic substances throughout the internal space available seems to be an important, if not a primary, internal transport mechanism. Even in multicellular organisms, diffusion plays an important role at the cellular level.
While diffusion accounts for much of the movement of fluids and solutes within cells, intercellular diffusion accounts for movement between cells. The solute is the component that is dissolved in a solution (the solution is the solvent).
This important mechanism helps move material from one cell to another in multicellular organisms. In plants, intercellular diffusion may be facilitated by plasmodesmata, the strands of protoplasm that penetrate cell walls and connect the cytoplasm of adjacent cells (see Chapter 2).
Often, diffusion is a very slow process, one that is supplemented by other mechanisms. One process, known as cytoplasmic streaming, has been observed in many cells; the cytoplasm flows along what appears to be a definite route throughout the cell, moving substances many times faster than would otherwise be possible. Food vacuoles often move throughout a cell, distributing digested material to different parts of the cytoplasm.
Some plants, such as the bryophytes – which include the liverworts, hornworts, and mosses – lack vascular tissues, the efficient long-distance internal transport systems that are responsible for moving fluids throughout the bodies of higher plants. The absence of such tissues has probably limited the size attained by most of these plants for two reasons. First, when the vascular tissue xylem is present, it can also function as a major supportive tissue in vascular plants. Second, without an efficient internal transport system, the plants are unable to move materials rapidly around great distances, thus restricting the overall size.
There are other internal transport mechanisms possessed by multicellular organisms. These represent major alternatives to those found among the larger, more dominant plant and animal groups. Jellyfish, hydra, and planaria lack true circulatory systems, but they have other mechanisms that free them from relying completely on diffusion and intracellular transport. For instance, the gastrovascular cavity of the hydra penetrates each tentacle, enabling food particles to be absorbed by each tentacle cell. Planaria have branched gastrovascular cavities extending throughout their body; this system moves food particles to all their cells without the aid of a circulatory system.
TRANSPORT IN PLANTS
Plants need carbon dioxide and water to produce carbohydrates (plant tissues). Water is often a limiting factor, so many plants have developed effective systems that facilitate the absorption, movement, conservation, storage, and utilization of water. The term “vascular,” when referring to specific plant tissues, has to do with those tissues concerned with tubular internal transport, such as the xylem and phloem in plants. The evolution of these tissues has enabled plants to develop greater heights, more specialized parts, and more highly integrated functions. It is widely believed that the successful exploitation of terrestrial environments by plants followed the evolution of these complex internal transport systems.
The vascular tissue is continuous throughout the plant, extending through the roots, stems, twigs, and leaves, as well as other parts of the organism. Here, for the sake of simplicity, the major components of this system will be considered individually.
STEMS
The outermost tissue layer of the stem of herbaceous plants, those plants without woody parts, is called the epidermis. Inside this is the cortex, which sometimes is divided into two layers: the one just under the epidermis, called the collenchyma, and the innermost, the parenchyma. Just internal to the cortex is the vascular tissue.
Vascular tissue in some plants is arranged in bundles. In others it occurs as a continuous layer, like a cylinder, lying inside the cortex with the pith filling the innermost part of the stem. The pith is a storage area. Those plants with vascular tissue arranged in bundles tend to have no clear distinction between the cortex and the pith. Rather, the tissue throughout the interior part of the stem appears to be quite homogenous.
In other types of plants, the phloem lies outside the xylem, with a layer of tissue in between the two that is primarily involved with producing new cells through mitotic cell division. This middle layer is the lateral meristematic tissue, or cambium. After the initial growth phase of many plants, the cambium of herbaceous species ceases to produce more phloem and xylem cells. However, in other species, the cambium reactivates at the beginning of the next growing season and continues to produce more phloem cells on the outside and xylem cells on the inside. In some plants (e.g. trees), the old phloem can be seen flaking off. The bark includes all the tissues outside the vascular cambium. On the inside, the old xylem continually forms additional rings of tissue that gradually add to the stem's or trunk's diameter. These are the growth rings, which appear in cross section when a tree is sawed down. As years go by, more xylem gets laid down, more phloem continually flakes off, and the stem increases in thickness; eventually the bulk of the stem of the older plant is made primarily of xylem. This tissue is commonly called wood. Some plants also have a cork cambium located outside the cortex, which divides mitotically to create the most external tissue, known as cork (see Figure 11.1).
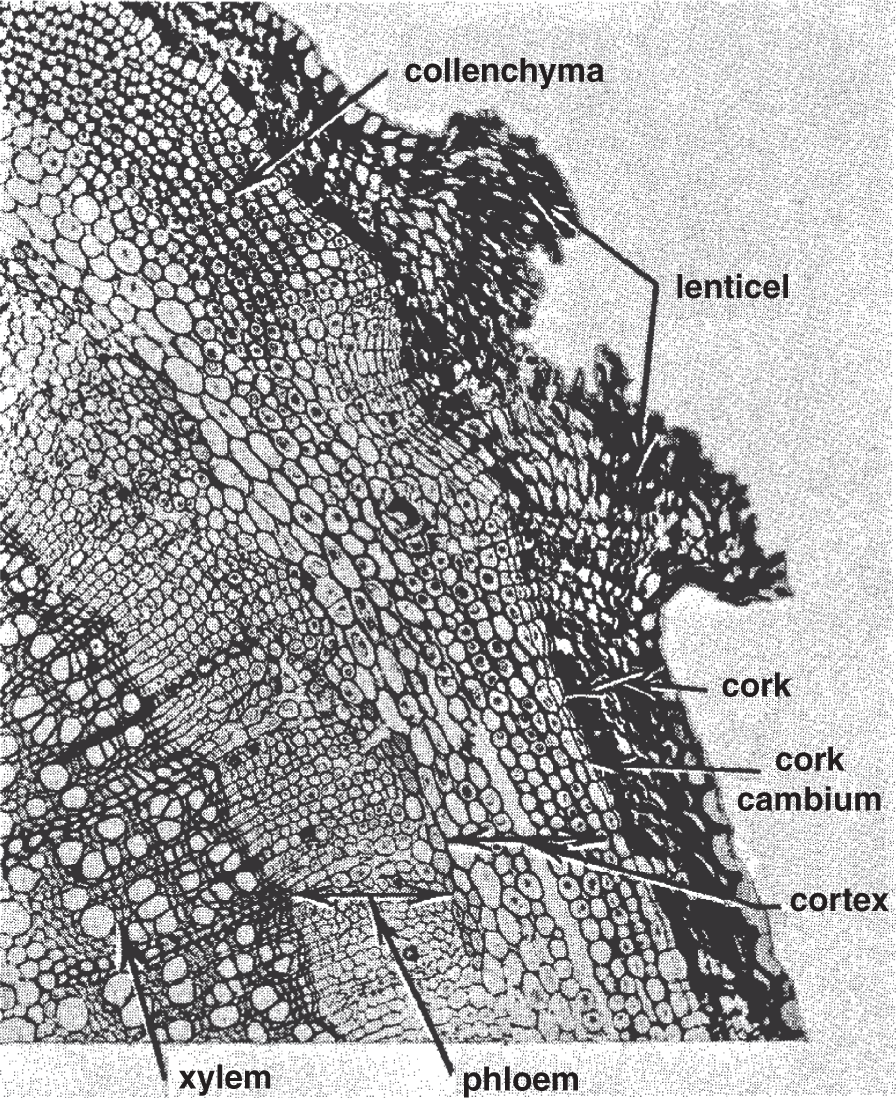
Figure 11.1 Going from the inside to the outside of the stem, the tissues shown in this elderberry stem cross section are xylem, phloem, cortex, cork cambium, and cork. The vascular cambium consists of the first two layers of thin-walled cells on the phloem side of the xylem.
CONDUCTING TISSUES IN PLANTS
Phloem
As stated earlier, phloem carries carbohydrates, the products of photosynthesis, generally in the form of sucrose, from the leaves to the nonphotosynthetic parts of the plant. The conducting cells of the phloem are the sieve elements, which are joined together in vertical columns, creating the sieve tubes, each of which is joined at the end, at the sieve plates. Where the sieve elements meet, the sieve plates are perforated, facilitating the movement of intracellular contents from cell to cell.
Next to the sieve elements are companion cells that seem to be involved in maintaining the sieve elements. The companion cells are living when they mature, though they lose many of their organelles, including the nucleus, which disintegrates. For the sieve elements to remain functional, they have to retain an intact cell membrane, as well as their watery cytoplasm. The absence of most organelles in the phloem cells may promote the mass movement of water and solutes through the sieve tubes.
Most of the movement of the contents in the sieve tubes occurs from the regions where plant food is produced, to the sites of food storage or utilization, where the solutes and water move from the sieve tubes into the adjacent cells and into the xylem. In addition to carbohydrates, organic nitrogen compounds move through the phloem, being conducted not only downward but, depending on where the nutrients are needed, laterally and upward as well.
Xylem
Xylem is the plant tissue that carries water and ions from the roots to other parts of the plant. It is laid down along the inside of the cambium (lateral meristematic tissue). Unlike phloem, the xylem's conducting cells, the tracheids and vessel elements are water-conducting cells with thickened cell walls. They die at maturity and become empty shells consisting of primary and secondary cell walls without a cell membrane, cytoplasm, nucleus, or any organelles. (The additional types of xylem cells are rarely mentioned in most introductory texts.)
Tracheids are long, narrow cells tapering at the ends where they come in contact and overlap with each other. The first tracheids to be laid down by the plant are characterized by ring-like or annular secondary cell wall thickenings and/or helical or spiral secondary cell wall thickenings that are capable of being stretched as the stems grow. In addition to having meristematic cells (capable of active cell division and differentiation into specialized tissues) that develop into phloem and xylem cells, the vascular cambium also has meristematic cells that develop into a system of cells oriented at a right angle to the stem's axis. These structures, known as vascular rays, carry food from the sieve tubes radially into the cambium and xylem. And sometimes the rays store large quantities of carbohydrates.
ROOTS
Most roots are storage organs, holding minerals and carbohydrates for future use, but they also function as a holdfast, stabilizing the plant (see Figure 11.2).
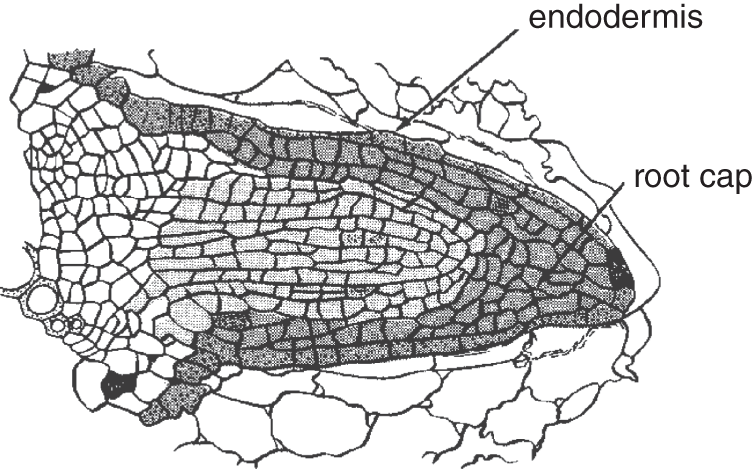
Figure 11.2 At the start, a young wheat root begins growing by pushing out through the cells of the cortex.
And, of course, they have the vascular tissues, xylem and phloem. Much of the absorption of water and minerals takes place through the root hairs, which are thin outgrowths of epidermal cells. They lack the waxy cuticle that protects other thin structures, such as leaves. In many species, a symbiotic relationship exists between the plant's root hairs and the filamentous elements of specific fungi, which form the mycelium. The product of such a close association is known as mycorrhizae.
Root Pressure
There seem to be several mechanisms that account for the upward movement of fluids and solutes from one cell to the next. Root pressure is responsible for some of the movement of water across the root tissues. It appears to be a function, in part, of simple diffusion along a concentration gradient. However, more seems to be involved. Some other pressure besides simple diffusion is present since the water in the xylem is more concentrated than that in the protoplasm of the endodermal tissue (the tissue surrounding the vascular cylinder). In other words, if simple diffusion alone were involved, the water probably wouldn't just move into the xylem, but also out of it. In addition, the downward hydrostatic pressure exerted by the column of water standing in the xylem would be expected to force the water out of the vascular tissue and into the surrounding root tissues. Instead, as it actually happens, water goes in and up the xylem.
The force exerted by the roots disappears when the roots are killed or deprived of oxygen. It has been found that the respiratory production of adenosine triphosphate (ATP) is necessary to provide the energy to create a solute gradient that is at least partially responsible for the movement of water through the root. In addition to the osmotic pressure and the use of semipermeable membranes, facilitated diffusion and active transport may also be involved.
Cohesion Theory
Another hypothesis, the cohesion theory, sometimes called the transpiration theory, states that water is pulled from above. According to this concept, water is lost through transpiration, the escape of water vapor from the aerial parts of the plant, and is replaced by the fluids in adjoining cells, creating an osmotic gradient that draws water from the xylem in the veins of the leaf to the adjacent tissues. The removal of water from the xylem for other uses, such as in photosynthesis, growth, or other metabolic processes, would also contribute to the draw, bringing more water up the xylem. This theory is strengthened by knowledge of the great cohesive forces that exist between individual water molecules.
This system is not, however, run simply by suction, which usually results from the removal of air, but by the removal of water, creating a pull that affects all the water below. Suction alone could not pull so much water to such heights.
The inside of the thin tubes may be made of a material to which water molecules adhere. This, in conjunction with the annular and the spiral secondary cell wall thickenings, probably accounts for part of the necessary architecture that allows water to rise, in some cases as high as hundreds of feet, defying most projections of what would otherwise be possible. In addition, the internal design may also reduce the water's downward hydrostatic pressure.
Another factor is the considerable weight of the atmosphere resting on the ground. The atmosphere creates enough pressure to account for pushing water up the xylem some 32 ft (almost 10 m). Together, atmospheric pressure, combined with the effects related to transpiration, the adhesive forces that prevent the water column from breaking under the pressure of gravity, capillary action that draws water up the thin tubes resulting from the adhesion of the water molecules to the inside of the xylem, the cohesion of the water molecules to each other, and the intricate design of the vessel's interior seem to account for a good portion of the upward movement of water observed in the xylem. The cohesive strength of water has been experimentally shown to be equivalent to the pressure exerted by 25–30 atmospheres.
MOVEMENT OF INORGANIC SOLUTES
Through the xylem, such inorganic ions as calcium, phosphorus, and sulfur are transported up from the roots to the leaves. Although phosphorus ions are capable of moving up through the xylem and down through the phloem, calcium ions are not nearly so mobile in the phloem and cannot be transferred from dying leaves to newer ones. Plants therefore have to obtain a steady supply of new calcium ions from the soil.
Different substances move through the sieve tubes at different speeds; sugar moves through some phloem 40,000 times faster than when diffusing through a liquid. The direction of movement through any given sieve tube reverses periodically, sometimes with the contents in neighboring sieve tubes moving in the opposite direction. Obviously, the transport through living phloem cells is unlike that through the dead xylem cells.
It has been suggested that materials are carried through the phloem cells by cytoplasmic streaming. Materials move from phloem cell to phloem cell through the sieve plates by diffusion; active transport may also be involved. Then they are carried to the other end of the cell by the cytoplasmic streaming, and then diffuse out again. Cytoplasmic streaming involves movement in both directions. This could explain the suggestion that some materials move in one direction, while others move in the opposite direction, through any given cell, though this has yet to be proven. Facilitated diffusion may also be involved.
Another hypothesis involves pressure flow of water and solutes through the sieve tubes along a turgor-pressure gradient. Known as the mass-flow hypothesis, it states that cells such as those in the leaves contain high concentrations of osmotically active substances such as sugar. So water diffuses into these and adjacent cells, raising the internal pressure, or turgor. This pressure forces substances from one cell to the next, pushing solutes farther up the plant. Since the sugars are being used up in the metabolically active areas, such as those that are actively growing, as well as in the cells that are sequestering the sugars in storage organs, osmotic concentrations are lowered in the nearby sieve tubes. Therefore, they lose water and turgor pressure. This results in a mass flow of the contents of the sieve tubes from the regions under high pressure. Such regions include leaves, where sugars are being made, and storage organs such as the roots, from which sugars exit during the spring when they are in transit to where they are needed. The contents then move to areas of low pressure, such as those regions where reserves are being rapidly consumed – where cells are growing or where materials are being put into storage. As presented, this system depends on the massive uptake of water at one end, because of high osmotic concentrations, and on the massive loss of water at the other end, because of the low osmotic concentrations that are due to the constant depletion of sugar and other solutes.
KEY TERMS
| annular secondary cell walls | epidermis |
| bark | food vacuoles |
| cambium | gastrovascular cavity |
| cohesion theory | herbaceous |
| collenchyma | inorganic ions |
| cork | intercellular diffusion |
| cork cambium | lateral meristematic tissue |
| cortex | mass-flow hypothesis |
| cytoplasmic streaming | meristematic cells |
| diffusion | mycelium |
| endodermal tissue | mycorrhizae |
| parenchyma | sieve tubes |
| phloem | spiral secondary cell walls |
| pith | tracheids |
| plasmodesmata | transpiration |
| pressure flow | transpiration theory |
| root hairs | vascular cambium |
| root pressure | vascular rays |
| roots | vessel elements |
| sieve elements | wood |
| sieve plates | xylem |
SELF-TEST
Multiple-Choice Questions
Internal Transport Systems
- Most plants have the following major pathways for internal transport:
- arteries and veins
- arteries, veins, and lymphatic vessels
- xylem, phloem, arteries, and veins
- trachea, dorsal longitudinal vessels, xylem, and phloem
- xylem and phloem
- Many animals have the following major pathways for internal transport:
- xylem and phloem
- arteries, veins, xylem, and phloem
- arteries, xylem, and phloem
- arteries, veins, and lymphatic vessels
- lymphatic vessels, xylem, and phloem
- Single-celled organisms such as bacteria and protists rely on ___________ as their major transport system(s).
- diffusion
- xylem and phloem
- arteries and veins
- cilia and flagella
- all of the above
- In plants, intercellular diffusion may be facilitated by ___________.
- food vacuoles
- microtubules
- cytoplasmic streaming
- plasmodesmata
- all of the above
- The following often move throughout a cell, distributing digested material to different parts of the cytoplasm:
- endoplasmic reticulum
- microtubules
- food vacuoles
- cytoplasmic streaming
- all of the above
Transport in Plants
- Some plants, such as the bryophytes, which include the liverworts, hornworts, and mosses, ___________.
- have vascular tissues
- lack vascular tissues
- have gastrovascular cavities
- have a circulatory system
- have xylem and phloem
- When referring to specific plant tissues, the following term concerns those involved with internal transport:
- cortex
- collenchyma
- parenchyma
- epidermis
- vascular
- It is thought that the evolution of vascular tissue enabled plants to develop ___________.
- greater height
- more specialized parts
- more highly integrated functions
- more successfully on land
- all of the above
- When plants have vascular tissue, it is usually found in the ___________.
- roots
- stems
- twigs
- leaves
- all of the above
- The outermost tissue layer of an herbaceous plant's stem is called the ___________.
- epidermis
- cortex
- collenchyma
- parenchyma
- pith
- Directly inside the vascular cambium is the ___________.
- cortex
- pith
- xylem
- collenchyma
- parenchyma
- Just outside the cork cambium is the ___________.
- cork
- collenchyma
- parenchyma
- vascular tissue
- pith
- Those tissues concerned with tubular internal transport, such as xylem and phloem, are called ___________.
- epidermis
- collenchyma
- parenchyma
- pith
- vascular
- In between the phloem and the xylem lies the ___________.
- lateral meristematic tissue
- vascular cambium
- apical meristematic tissue
- a and b
- a, b, and c
- In some plants, especially trees, the old ___________ can be seen flaking off, as the old bark.
- xylem and phloem
- phloem, cortex, and cork
- xylem
- a and b
- all of the above
- Next to the sieve elements are ___________ that seem to be involved in maintaining the sieve elements.
- sieve tubes
- sieve plates
- companion cells
- all of the above
- none of the above
- ___________ carries carbohydrates, usually in the form of sucrose, from the leaves to the nonphotosynthetic parts of the plant.
- xylem
- phloem
- arteries
- veins
- lymphatics
- ___________ is the plant tissue that carries water and ions from the roots to other parts of the plant.
- xylem
- phloem
- sieve tube
- sieve plate
- companion cell
- ___________ is the vascular tissue laid down on the inside of the cambium (lateral meristematic tissue).
- xylem
- phloem
- sieve tube
- sieve plate
- companion cell
- Unlike the phloem, the xylem's conducting cells, the ___________ and ___________ die at maturity and become empty shells consisting of primary and secondary cell walls, without a cell membrane, cytoplasm, nucleus, or any organelles.
- sieve tubes, sieve elements
- sieve plates, sieve tubes
- companion cells, sieve elements
- vessel elements, sieve tubes
- tracheids, vessel elements
- ___________ are long, narrow cells with tapering ends that overlap where they come in contact with each other. The first to be laid down are characterized by annular and/or spiral secondary cell walls that are involved in moving fluids up through the xylem tissue.
- tracheids
- vessel elements
- companion cells
- sieve tubes
- sieve elements
- ___________ that grow later in the plant's life have numerous pits which allow water and dissolved substances to move from cell to cell.
- tracheids
- vessel elements
- companion cells
- sieve tubes
- sieve elements
- ___________ are more specialized than tracheids, from which they probably evolved, and are found primarily in the more advanced, flowering plants.
- tracheids
- vessel cells
- companion cells
- sieve tubes
- sieve elements
- In the xylem are clusters of parenchyma cells forming ___________, which instead of running laterally up and down the plant, as do the other internal transport cells, run radially, and facilitate lateral movement of materials.
- tracheids
- vessels cells
- vascular rays
- sieve elements
- companion cells
Roots, Movement of Fluids, and Cohesion Theory
- Much of the absorption of water and minerals takes place through the ___________, which are thin outgrowths of epidermal cells.
- vascular rays
- tracheids
- companion cells
- sieve tubes
- root hairs
- When the roots are killed or are deprived of oxygen, the force that seems to be pushing fluids up, known as ___________, is no longer present.
- root pressure
- cohesion pressure
- transpiration pressure
- endodermal pressure
- atmospheric pressure
- The ___________ theory states that water is pulled from above.
- cohesion
- root
- transpiration
- a and b
- a and c
- According to the cohesion theory, water is lost through ___________, and it is replaced by the fluids in adjoining cells, creating an osmotic gradient that draws water from the xylem in the veins of the leaf to the adjacent tissues.
- cohesion
- transpiration
- insulation
- circulation
- pressurization
- ___________ can only account for pushing water up xylem about 32 feet.
- cohesion pressure
- transpiration pressure
- endodermal pressure
- root pressure
- atmospheric pressure
ANSWERS
- e
- d
- a
- d
- c
- b
- e
- e
- e
- a
- a
- a
- d
- d
- a
- c
- b
- a
- a
- e
- a
- a
- b
- c
- e
- b
- e
- b
- e
Questions to Think About
- Describe the conducting tissues of plants.
- What are the theories concerning how materials are conducted throughout plants?
- What is the function of the roots, and how do they work?