ONE

Origin and Relationships of Dinosauria
Dinosauria is a well-diagnosed clade, and since birds are included within it the group is clearly significant among terrestrial vertebrates. Dinosaurs belong within Archosauria, a broader clade that also includes crocodilians as well as pterosaurs and various basal groups of Triassic age. Over the past thirty years considerable effort has been devoted to disentangling the phylogeny of archosaurs: some relationships have been firmly established, while others have yet to be discovered. This chapter considers the origin of the dinosaurs in terms of phylogeny and the timing of events. The primary evidence comes from a cladistic analysis of the Triassic archosaurs, a paraphyletic group formerly known as thecodontians. That analysis is followed by an account of the evolutionary events that led to the radiation of the most astonishing animals of all time, the dinosaurs themselves.
Previous Cladistic Analyses of Dinosauria
Paleontologists have been interested in the phylogeny of the dinosaurs since their discovery in the 1820s. However, confusion reigned over many aspects of dinosaurian phylogeny until the 1980s, when cladistics began to be applied. The use of cladistic methods led to the solution of many phylogenetic conundrums, but others have continued to defy resolution.
Cladistics as applied to dinosaurs and their close relatives began in the early 1980s with a number of basic studies in which cladograms were compiled by hand from incomplete lists of sometimes poorly delimited characters. In the late 1980s and 1990s data matrices were analyzed by computer algorithms, and evidence was often given of the goodness of fit of particular parts of the tree to the data.
Further developments in cladistic practice have occurred since the publication of the first edition of The Dinosauria in 1990. Some workers have advocated the use of formalized systems for naming and diagnosing taxa. Both before 1990 and since, names have been applied to nodes in cladograms according to the whim of the systematists. Some workers name every node in a cladogram, while others prefer to name only those nodes that are robust (i.e., supported by a great deal of character evidence). In addition, there are differences in the choice of names for clades: some systematists prefer to retain well-known group names that most closely match the traditional understanding of contents and/or diagnostic characters, while others advocate the migration of widely used names to clades subtended by extant groups, the crown-clade concept.
There have also been important developments in the definition and diagnoses of taxa. Until 1990 most cladists equated taxa with their characters (e.g., Aves is diagnosed as those organisms that possess primary flight feathers and wings). Since then, a clear distinction has been proposed between the definitions and diagnoses of clades (de Queiroz and Gauthier 1992, 1994). Clades are diagnosed by characters that evolve at or immediately prior to their origin but do not make individual clades “what they are.” Instead, individual clades are named on the basis of their membership. Named taxa are then fixed on the basis of their ancestry.
There are three kinds of definitions of taxon names: apomorphy-based, stem-based, and node-based. An apomorphy-based definition is founded on one or more derived characters (e.g., “Aves consists of all archosaurs with feathered wings used for powered flight”). Apomorphy-based taxa combine definition and diagnosis. Stem- and node-based definitions include ancestors as the basal member of the clade (fig. 1.1). A stem-based taxon name refers to a clade that includes all the descendants from a particular cladogenetic event. An example of stem-based definition for Ornithischia is “all dinosaurs more closely related to Triceratops than to Tyrannosaurus.” A node-based taxon name is one that is defined by a basal node. For example, a node-based definition for Aves takes the form “Archaeopteryx, Neornithes, their most recent common ancestor, and all descendants.” Because we do not know all of the stem taxa in stem-based taxa, their diagnoses are not possible. Node-based taxa can be diagnosed on the basis of the derived features that have evolved in or immediately prior to the ancestor of the clade (this practice has been applied to stem-based clades as well). A crown clade is a node-based clade defined solely on the basis of extant forms. For example, the crown group Mammalia is defined as the common ancestor of Ornithorhynchus (platypus) and Homo (humans) and all the descendants of this common ancestor.
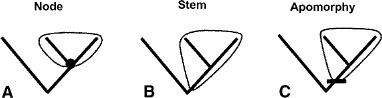
FIGURE 1.1. The three kinds of clade definitions: A, node-based; B, stem-based; C, apomorphy-based.
Archosauria
The crown-group issue affects the discussion of basal archosaurs and the origin of dinosaurs since certain names have different meanings for different people. Cope (1869a) introduced the name Archosauria for a wide group of extinct and extant reptiles, including anomodonts and rhynchosaurs, and no one has proposed a strict return to his view. However, for most of the twentieth century the name was used to refer to a group that includes modern crocodilians, as well as dinosaurs, pterosaurs, and a variety of Triassic forms, back to the Early Triassic proterosuchids and erythrosuchids, the earliest representative being Archosaurus, from the Late Permian (Tatarian) of Russia (e.g., Romer 1945, 1956, 1966; Hughes 1963; Charig and Reig 1970; Cruickshank 1972; Bonaparte 1982; Carroll 1988; Juul 1994; Gower and Wilkinson 1996; Benton 1997). Archosauria in this sense turns out to include those diapsids that possess an antorbital fenestra—an apomorphy-based taxon.
Gauthier (1986), urging use of the crown-clade concept, defined Archosauria as the clade subtended by living archosaurs (birds and crocodilians), in other words, excluding proterosuchids, erythrosuchids, proterochampsids, and euparkeriids. He argued that this realignment of the name had the advantage that all archosaurs, including fossil forms, would then have predictable soft-part characters (based on extrapolation from living forms). The apomorphy-based Archosauria was then renamed Archosauriformes (Gauthier 1986). Archosauria could also be defined as a stem-based taxon by extending membership down the stem to the next known node, but this approach has not yet been proposed.
Archosauria is treated as an apomorphy-based clade here, with note made to node- or stem-based definitions where relevant. Phylogenetic studies of dinosaurs in their archosaur context have led to the following generally accepted conclusions:
1. Archosauria is monophyletic. This view has been held generally for a long time, although the clade has been regarded as hard to diagnose anatomically (e.g., Romer 1956, 1966) and hence possibly polyphyletic.
2. Archosauria includes a number of basal Triassic forms that are sister groups to Avesuchia sensu Benton (1999), also known as crown group Archosauria sensu Gauthier (1986). The clade consists of two lines, one leading to crocodilians, the other to birds. This split was hinted at by Bonaparte (1975a), Krebs (1976), Cruickshank (1979), and Chatterjee (1982), and it has been confirmed in all subsequent cladistic analyses.
3. The crocodile line, Crurotarsi (Sereno 1991a), consists of Phytosauridae, Ornithosuchidae, Prestosuchidae, Rauisuchidae, Poposauridae, and Crocodylomorpha, but the relationships among those groups are contentious (Gauthier 1986; Benton and Clark 1988; Sereno 1991a; Parrish 1993; Juul 1994; Benton 1999).
4. The bird line, Avemetatarsalia (Benton 1999), consists of Scleromochlus, Pterosauria, and Dinosauromorpha. The South American forms, Lagerpeton and Marasuchus, are close outgroups of Dinosauria (Gauthier 1986; Benton and Clark 1988; Sereno 1991a; Sereno and Arcucci 1993, 1994; Juul 1994; Novas 1996a; Benton 1999). Avemetatarsalia is similar but not equivalent to Ornithosuchia sensu Gauthier (1986) because the former does not include Ornithosuchidae, a clade thought to be included in Crurotarsi (Benton 1999).
5. Dinosauria is monophyletic. Up to 1985, dinosaurs were usually seen as polyphyletic, with as many as three to six ancestors (e.g., Romer 1966; Charig and Reig 1970; Charig 1976a; Krebs 1976; Cruickshank 1979; Thulborn 1980; Chatterjee 1982). Bakker and Galton (1974) and Bonaparte (1976) argued for dinosaurian monophyly before such views became generally accepted (e.g., Gauthier 1986; Benton and Clark 1988; Sereno 1991b, 1997; Sereno and Arcucci 1993, 1994; Juul 1994; Novas 1996a; Benton 1999).
6. Dinosauria includes two clades, Saurischia and Ornithischia. This view has been generally held since Seeley's (1887a) invention of the two names, although he based Saurischia on what would now be seen as a plesiomorphy, namely, the “reptilian” pelvic arrangement. Apomorphies for both clades were given by Gauthier (1986).
7. Saurischia comprises several basal taxa, as well as Theropoda and Sauropodomorpha. These two clades were formerly regarded as having arisen from two or more independent sources among the basal archosaurs (e.g., Charig et al. 1965; Charig 1976b; Cruickshank 1979; Chatterjee 1982), a pattern for which there is no evidence (Gauthier 1986).
8. Ornithischia comprises several basal forms, as well as Thyreophora and Cerapoda (Sereno 1986, 1997, 1998; Weishampel and Witmer 1990a). Thyreophora includes the armored Stegosauria and Ankylosauria, as well as several basal taxa. Cerapoda splits into Marginocephalia (Pachycephalosauria and Ceratopsia) and Ornithopoda (Heterodontosauridae and Euornithopoda).
Archosauria and Its Evolutionary Context
Among living vertebrates, birds and crocodilians are linked as sister groups within the Avesuchia/crown group Archosauria (fig. 1.2). Although seemingly different kinds of animals, these two groups share numerous derived characters of the skull, postcranial skeleton, and soft parts that are absent in other living vertebrates (Gauthier 1986; Benton and Clark 1988). Early molecular studies of the phylogeny of tetrapods were equivocal regarding the nature of Archosauria, and many analyses of protein sequences supported a close pairing of Aves and Mammalia (e.g., Bishop and Friday 1988; Hedges et al. 1990; reviewed in Benton 1990b). More recently, the validity of Archosauria has been accepted based on sequencing of nucleic acids (e.g., Janke and Arnason 1997; Hedges and Poling 1999), although this has been challenged by the suggestion that turtles belong to Archosauria.
Archosauria is included in the larger clade Diapsida (living crocodilians, birds, Sphenodon, lizards, snakes, and their extinct relatives), one of the major clades of tetrapods. The crown group Tetrapoda, four-limbed vertebrates, includes Amphibia and various basal groups, as well as Amniota (Reptilia + Aves + Mammalia). Amniota comprises Anapsida, Diapsida, and Synapsida, diagnosed by the nature of their temporal openings, among other characters (Laurin and Reisz 1995; Benton 1997). The stability of this division of Amniota has been challenged by some morphological and molecular results that suggest that Anapsida should be subsumed within Diapsida: Rieppel and de Braga found that turtles may have been close relatives of lepidosaurs (Sphenodon, lizards and snakes) based on morphological characters (Rieppel and de Braga 1996; de Braga and Rieppel 1997), although their results have been queried (Wilkinson et al. 1997), while complete mitochondrial DNA sequencing suggests that turtles are the sister group to Archosauria (Zardoya and Meyer 1998; Hedges and Poling 1999; Kumazawa and Nishida 1999). These dramatic proposals have not yet been fully tested (reviewed in Rieppel 2000).

FIGURE 1.2. Phylogeny of tetrapods showing the position of Archosauria. Synapomorphies at major nodes are listed.
1. TETRAPODA: single pair of nasals meeting in midline; compact oticoccipital region to skull; cheek with broad jugal-quadratojugal contact; absence of coronoid fangs; absence of operculars, median gular, and submandibulars; presence of pre- and postzygapophyses; well-developed ventrally directed ribs; large ornamented interclavicle; carpus, tarsus, and up to eight digits; iliac blade of pelvis extended dorsally and attached to vertebral column by sacral ribs; ischia contribute to puboischial symphysis; femur with adductor crest.
2. AMNIOTA: frontal enters orbital margin; narrow convex occipital condyle; absence of coronoid denticles; three scapulocoracoid ossifications (primitively); astragalus present.
3. EUREPTILIA: tabular absent; middle coronoid absent; cleithrum absent.
4. DIAPSIDA: supra- and intratemporal fenestrae; suborbital fenestra; paroccipital process reaches suspensorium; sternum mineralized.
5. NEODIAPSIDA: lacrimal does not contact nasal or naris; postparietal absent; contact between ectopterygoid and maxilla; gastralia present; medial epicondylar foramen on humerus absent; fifth distal tarsal absent.
6. ARCHOSAUROMORPHA: caudodorsal process on premaxilla; sagittal crest on parietal; slender tapering cervical ribs at a low angle to the vertebrae; notch on cranial margin of clavicle; dorsal margin of ilium composed of a small cranial process and a large caudal process; medial centrale of carpus absent.
7. UNNAMED CLADE: foramen for entrance of internal carotid arteries on ventral surface of parasphenoid; astragalocalcaneum articulation concavoconvex; lateral tuber on calcaneum; centrale of tarsus contacts tibia.
8. UNNAMED CLADE: ectopterygoid expanded caudally; upturned retroarticular process; second sacral rib bifurcate and with caudal process truncated sharply; cranial apron of pubis.
9. UNNAMED CLADE: Elongate snout (more than 50% of skull length); nasal longer than frontal; posttemporal fenestra small; teeth recurved; teeth laterally compressed; pterygoids remain separate; distal ends of cervical neural spines expanded as a spine table.
(After Benton 1985; Laurin 1991; Ahlberg and Milner 1994; and Dilkes 1997.)
Archosauria was established by Cope (1869a) for a broad grouping of amniotes: Crocodylia, Thecodontia, Dinosauria, Anomodontia (i.e., dicynodonts + dinocephalians), and Rhynchocephalia (i.e., sphenodontids + rhynchosaurs). Cope (1869b) then restricted Archosauria to include Dinosauria, Phytosauria, Crocodylia, and Rhynchocephalia, excluding anomodonts. In the 1890s Cope and Baur independently developed the “theory of fenestration,” according to which the major lines of amniote evolution could be identified by the numbers of temporal fenestrae, whether none (Anapsida), one (Synapsida), or two (Diapsida), the last two group names introduced by Osborn (1903) to reflect the new phylogenetic ideas.
Osborn (1903) rejected Cope's Archosauria, instead referring archosaur groups to a number of separate sections within Diapsida. This view was followed by many workers until the 1930s, with Broom, von Huene, Haughton, and others referring Triassic archosaurs to Pseudosuchia, Parasuchia, and Protorosauria. Romer (1933) resurrected Archosauria for Thecodontia, Crocodylia, Pterosauria, Saurischia, and Ornithischia but included proterosuchids in Eosuchia within Lepidosauria. Proterosuchids and erythrosuchids were replaced in Archosauria in later works (Romer 1945, 1956, 1966), and this has been the generally accepted viewpoint since (e.g., Hughes 1963; Charig and Reig 1970; Cruickshank 1972; Bonaparte 1982; Carroll 1988; Juul 1994; Gower and Wilkinson 1996; Benton 1997).
Archosauria sensu Benton 1999 is nested within larger clades, the most significant of which are Archosauromorpha and Diapsida. Diapsida comprises some basal taxa, principally Araeoscelidia (Petrolacosaurus, Araeoscelis, and relatives) from the Late Carboniferous and Early Permian, and two major clades, Lepidosauromorpha and Archosauromorpha, which diverged presumably early in the Permian (Benton 1983a, 1984b, 1985; Evans 1984, 1988; Gauthier 1986; Benton and Clark 1988; Laurin 1991; Laurin and Reisz 1995; Dilkes 1997).
Archosauromorpha has been given node- and stem-based definitions. According to the former, Archosauromorpha is defined as the most recent common ancestor of Neornithes (extant birds), Squamata (extant lizards and snakes), and all of the descendants of this common ancestor. Accordingly, the clade comprises Trilophosaurus, Rhynchosauria, Prolacertiformes, and Archosauria, as well as probably thalattosaurs, choristoderans, and drepanosaurids (Benton 1985; Benton and Clark 1988; Laurin 1991; de Braga and Rieppel 1997; Dilkes 1997). Trilophosaurus is an unusual Late Triassic herbivore with no infratemporal openings. Rhynchosaurs are a distinctive clade of Triassic herbivores that had multiple tooth rows and beaklike premaxillary projections. Prolacertiforms are long-necked insectivores and carnivores known from the mid-Permian to the Late Triassic that may comprise an unnatural grouping of diverse archosauromorph taxa (Dilkes 1997). Most cladistic analyses agree that among these archosauromorphs Prolacerta, a prolacertiform, is the sister group of Archosauria and that Rhynchosauria is a more distant outgroup. Other probable archosauromorphs include thalattosaurs, marine Triassic forms, choristoderans, superficially crocodilian-like aquatic animals known from the Triassic to the mid-Tertiary, and drepanosaurids, Late Triassic swimmers.
A stem-based definition for Archosauromorpha (the most recent common ancestor of Prolacerta, Trilophosaurus, Hyperodapedon, and archosaurs and all its descendants) was provided by Laurin (1991:90). However, this definition excludes Protorosaurus, Drepanosauridae, and Tanystropheidae, according to Dilkes's (1997) cladogram. Dilkes (1997:528) gave a revised stem-based definition for Archosauromorpha, namely, Protorosaurus and all other saurians that are related more closely to Protorosaurus than to Lepidosauria, but this definition is rejected here since it would refer to a much more restricted grouping according to other cladograms of basal diapsids: it is debated whether “Protorosauria,” Rhynchosauria, or Trilophosaurus is the basal archosauromorph taxon (e.g., Benton 1985; Chatterjee 1986a; Evans 1988; Laurin 1991). The present definition can cope with all the competing cladograms and still refers to the same clade contents (Trilophosaurus+ Rhynchosauria + “Protorosauria” + Prolacertiformes + Archosauria).
Laurin (1991:83) also gave a node-based definition of Diapsida: the most recent common ancestor of araeoscelidians, lepidosaurs, and archosaurs and all its descendants. However, in this definition it is unclear whether Petrolacosaurus or Araeoscelidia (i.e., Petrolacosaurus, Araeoscelis, and other taxa) is the basal taxon (e.g., Benton 1985; Evans 1988; Laurin and Reisz 1995). An apomorphy-based definition is preferred here since that indicates Osborn's (1903) intention that diapsids are those amniotes with two temporal openings, the view accepted ever since. Therefore, Diapsida refers to the clade stemming from the first amniote with a supratemporal fenestra homologous with that of Aves (Vultur gryphus Linnaeus, 1758).
Relationships of Triassic Archosauria
Triassic archosaurs, excluding crocodylomorphs, pterosaurs, and dinosaurs, used to be termed thecodontians (e.g., Romer 1966; Charig 1976a; Bonaparte 1982; Chatterjee 1982). The term thecodontian, literally “socket-tooth,” is inappropriate descriptively because virtually all archosaurs with teeth are thecodontian, as are mammals, whose teeth likewise reside in separate sockets within the bony jaws. More importantly, thecodontians have no cohesion or common characters, and the term has generally been abandoned because it is paraphyletic (Gauthier 1986).
Precladistic phylogenies of basal archosaurs offered a great range of hypotheses. Most authors (e.g., Romer 1956; Krebs 1976) favored a tripartite arrangement: Proterosuchia for basal forms (Proterosuchidae + Erythrosuchidae, but also at times Rauisuchidae, Prestosuchidae, and Proterochampsidae), Parasuchia (= Phytosauridae), and Pseudosuchia (= Stagonolepididae, “Rauisuchia,” Ornithosuchidae, some basal crocodylomorphs). Others separated Stagonolepididae as Aetosauria (e.g., Romer 1966; Bonaparte 1975a; Charig 1976a; Thulborn 1980), some of the early crocodylomorphs as Sphenosuchia (Bonaparte 1982), ornithosuchids as Ornithosuchia (Huene 1907–8; Chatterjee 1982), and rauisuchids and poposaurids as Rauisuchia (Chatterjee 1982).
Several precladistic analyses laid great emphasis on the structure of the ankle of basal archosaurs (e.g., Bonaparte 1975a, 1982; Cruickshank 1979; Chatterjee 1982; Cruickshank and Benton 1985; fig. 1.3). These authors distinguished a so-called primitive mesotarsal (PM) arrangement, in which there was a simple hinge between the astragalus-calcaneum unit and the rest of the foot (seen in proterosuchids and, in a modified version, in erythrosuchids and proterochampsids). The other basal archosaurs have a crocodiloid ankle in which the bending plane runs between the calcaneum and fibula, then zigzags down between the astragalus and calcaneum and extends across between the astragalus and foot. This pattern could occur in a crocodile-normal (CN) fashion, as in crocodylomorphs, rauisuchians, aetosaurs, and phytosaurs, or in a crocodile-reversed (CR) fashion, as in ornithosuchids. Pterosaurs and dinosaurs show the advanced mesotarsal (AM) ankle, in which the astragalus is united firmly with the tibia, the calcaneum is reduced and adheres firmly to the astragalus, and the ankle bends between those two proximal tarsals and the rest of the foot.
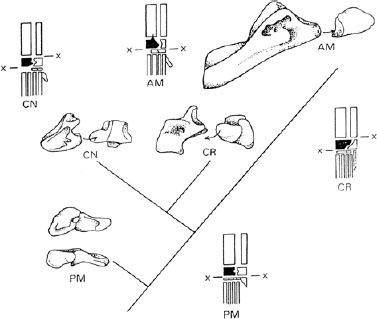
FIGURE 1.3. Ankle evolution in Triassic archosaurs. For each ankle pattern, a cranial view of the astragalus (left) and calcaneum (right) is shown. An additional proximal view is shown for the PM and NIPM types (upper drawing). For each ankle type, a diagram of the lower leg, ankle, and foot is shown (astragalus shaded black) with the main hinge line (x–x). The ankles illustrated here are the PM (primitive mesotarsal) of Proterosuchus, the MPM (modified primitive mesotarsal) of Chanaresuchus, the CN (crocodile-normal of Neoaetosauroides), the CR (crocodile-reversed) of Riojasuchus, and the AM (advanced mesotarsal) of a prosauropod dinosaur. (After Benton 1997.)
Sereno (1991a) showed that too much attention had been paid to the gross characters of the ankle, and restudy of the ankle characters and others showed that ornithosuchids formed part of the crocodile line, a group he termed Crurotarsi, comprising all archosaurs with a crocodiloid tarsus. Euparkeria is a basal form. Relationships of clades within Crurotarsi are uncertain, with a wide range of specific patterns having been proposed (Gauthier 1986; Benton and Clark 1988; Sereno 1991a; Parrish 1993; Juul 1994; Bennett 1996). A reanalysis and assessment of the robustness of nodes (Benton 1999) suggested only a weak indication that Ornithosuchidae might be basal to the other crurotarsans. On the other hand, relationships of taxa within the bird line have shown clearer resolution (Benton 1985, 1990a; Gauthier 1986; Benton and Clark 1988; Sereno 1991a, 1997; Juul 1994; Novas 1996a; Sereno and Arcucci 1993, 1994), with Pterosauria and Dinosauromorpha as sister groups and Lagerpeton and Marasuchus as successive outgroups to a monophyletic Dinosauria within Dinosauromorpha. The controversy concerned Scleromochlus, which might be the sister group of Pterosauria (Gauthier 1986; Sereno 1991a; Juul 1994; Novas 1996a) or an outgroup to Ornithodira, the basal member of Avemetatarsalia (Benton 1999).
Bennett (1996) offered an alternative view, extracting Pterosauria from Ornithosuchia sensu Gauthier (1986) and placing it as a basal archosaur clade, perhaps branching off between Proterosuchidae and Erythrosuchidae. He also found Ornithosuchidae to be the basal group on the bird line, resurrecting the view of Gauthier (1986), Benton and Clark (1988), and others. Bennett (1996) achieved this different view by recoding or excluding 11 characters of the hindlimb that he argued were convergences between pterosaurs and dinosauromorphs. These recodings are considered in the “Cladistic Analysis of Archosauria” section below.
It is important to review the robustness of previous efforts to establish the relationships of basal archosaurs and the position of Dinosauria among them. Seven independent cladograms of archosaurs are assessed: Gauthier (1986), Benton and Clark (1988), Sereno and Arcucci (1990), Sereno (1991a), Juul (1994), Bennett (1996), and Benton (1999). Benton and Clark (1988) did not publish their data matrix, but it exists in unpublished form and is resurrected here. The coding of the published data matrices used here has been neither checked nor changed; they are all available at http://palaeo.gly.bris.ac.uk/cladestore. The data matrices were analyzed using the branch-and-bound method of PAUP*4.0b3a (Swofford 1999), with all options set to default. The robustness of nodes was assessed using boot-strapping (heuristic 1,000 replicates), and the resulting trees are presented in figure 1.4. Nodes with less than 50% bootstrap support are collapsed. Nodes with greater than 50% bootstrap support are retained, and the bootstrap values are indicated. Basic tree statistics are given in figure 1.4. Gauthier's(1986) cladogram is shown simply as he gave it; it has not been recalculated because he did not offer a data matrix.
Replotting has modified some of the trees. In Benton and Clark (1988:fig. 8.1) Gracilisuchus is the outgroup to suchians (Crocodylomorpha + “rauisuchians” + Stagonolepididae). Here it forms an unresolved tritomy with suchians and Ornithosuchia sensu Gauthier (1986) as a result of assessment of the robustness of nodes. Reanalysis of the data of Sereno and Arcucci (1990) and Sereno (1991a) yielded no change in the results. Differences in the recalculated tree from Juul's (1994) data include the pairing of Youngina and Rhynchosauria in the outgroup, the collapse of Crocodylomorpha into a tritomy with Postosuchus and Gracilisuchus, and the collapse of Prestosuchidae and Stagonolepididae into a multitomy with Parasuchia and Suchia (Prestosuchidae and Stagonolepididae formed a distinct clade in Juul's results). The reanalysis of Bennett's (1996) data matrix similarly resulted in the collapse of some nodes because of low bootstrap values. Euparkeria joins a tetratomy with Parasuchia + Suchia, Ornithosuchidae, and Ornithodira: Bennett found that Ornithosuchidae was an outgroup to Ornithodira, contrary to Sereno's (1991a) finding that it fell within the crocodile line. Further, in the reanalysis Scleromochlus is the outgroup to Dinosauromorpha, although Bennett (1996:fig. 2) showed this genus as the sister group of Pterosauria. Benton's (1999) results are the same.
Despite the variations in the data sets used by the different authors, the cladograms all confirm that Archosauria comprises Proterosuchidae, Erythrosuchidae, Euparkeria, and Proterochampsidae, in that order, as outgroups to Avesuchia/crown group Archosauria. The position of Ornithosuchidae is controversial, as are the arrangement of “rauisuchians” and its relationship to Crocodylomorpha. Scleromochlus also has a variable position in Avemetatarsalia, whether as sister group to Pterosauria, sister group to Dinosauromorpha, or basal to Avemetatarsalia overall.
Cladistic Analysis of Archosauria
Of the 100 or more named species of Triassic archosaurs, certain terminal taxa have been selected for the new cladistic analysis reported here. Some of these terminal taxa are individual genera (in most cases individual species), while others are suprageneric groups. Species are used where there is dispute about their assignment to larger clades. Suprageneric taxa are employed where there is little dispute about taxon membership (e.g., Phytosauridae, Stagonolepididae, Pterosauria, Sauropodomorpha). A total of 22 ingroup taxa were analyzed.
Outgroup taxa for the cladistic analysis are the rhynchosaur Hyperodapedon gordoni (Benton 1983a) and the prolacertiform Prolacerta broomi (Gow 1975b; Evans 1986). These two taxa are generally accepted as archosauromorphs (Benton 1985; Evans 1988; Laurin 1991; Dilkes 1997) but not as archosaurs, and their anatomy is well known.
Ingroup terminal taxa include a diverse array of forms. Proterosuchus fergusi (Cruickshank 1972; fig. 1.5A), Erythrosuchus africanus (Gower 2003; fig. 1.5B), and Euparkeria capensis (Ewer 1965; fig. 1.5C) loosely represent their respective clades (i.e., Proterosuchidae, Erythrosuchidae, and Euparkeridae) since the bounds of these clades cannot be defined on the basis of a robust cladogram. In addition, Proterochampsidae (Sill 1967; Romer 1971a, 1972a; fig. 1.5D), Phytosauridae (Chatterjee 1978a; Hungerbühler 1998a, 2002; fig. 1.6A), Ornithosuchidae (Walker 1964; Bonaparte 1975b; Sereno 1991a; fig. 1.6B), and Stagonolepididae (Walker 1961; fig. 1.6C) are distinctive in terms of membership, although the precise validity and relationships of included species have yet to be fully established and the basal most members of each clade are unclear. Proterochampsidae includes those taxa that stem from the last common ancestor of Proterochampsa barrionuevoi and Chanaresuchus bonapartei and sharing a more recent common ancestor with those species than with Erythrosuchus africanus, Euparkeria capensis, Crocodylia, or Neornithes. Phytosauridae includes those taxa stemming from the last common ancestor of Mystriosuchus planirostris and Paleo rhinus bransoni and sharing a more recent common ancestor with those species than with Ornithosuchus longidens, Stagonolepis robertsoni, or Crocodylia. Ornithosuchidae includes those taxa stemming from the last common ancestor of Ornithosuchus longidens and Riojasuchus tenuiceps and sharing a more recent common ancestor with those species than with Mystriosuchus planirostris, Stagonolepis robertsoni, or Crocodylia. As indicated earlier, Ornithosuchidae is no longer the basal taxon in Ornithosuchia sensu Gauthier (1986). Stagonolepididae includes those taxa stemming from the last common ancestor of Stagonolepis robertsoni and Aetosaurus ferratus and sharing a more recent common ancestor with those species than with Mystriosuchus planirostris, Ornithosuchus longidens, or Crocodylia.
“Rauisuchia,” variously classified as Rauisuchidae, Prestosuchidae, Poposauridae, and Chatterjeeidae, cannot be treated at a suprageneric level since not all of the divisions among the various subgroups are clear (Gower 2000). Instead, six “rauisuchian” taxa from different positions in cladograms (Benton and Clark 1988; Sereno 1991a; Parrish 1993; Juul 1994) were chosen for these analyses. They include Ticinosuchus ferox (Krebs 1965), Prestosuchus chiniquensis (Huene 1942; Barberena 1978), Saurosuchus galilei (Sill 1974; Alcober 2000; fig. 1.6D), Fasolasuchus tenax (Bonaparte 1981), Gracilisuchus stipanicorum (Romer 1972b; Brinkman 1981), and Postosuchus kirkpatricki (Chatterjee 1985; Long and Murry 1995).
Crocodylomorpha comprises all archosaurs closer to Eusuchia than to Ornithosuchus longidens or Postosuchus kirkpatricki. These latter two species have been selected for outgroup comparison since the cladistic relationships of the nearest outgroups to Crocodylomorpha are unresolved. A stem-based definition is preferred since a number of basal forms (e.g., Trialestes, Pseudhesperosuchus, Saltoposuchus) are close to the origin of the clade, and new finds and studies might alter our understanding of the cladogram.
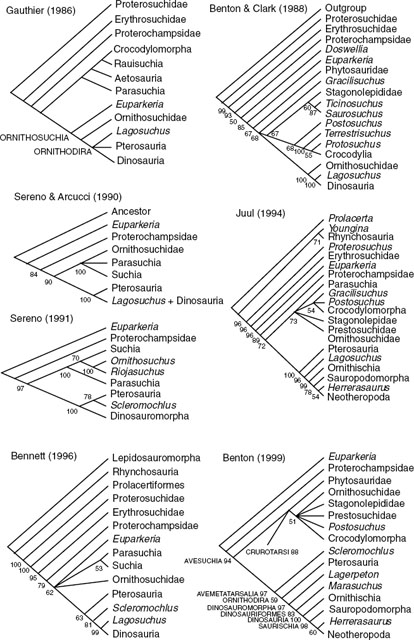
FIGURE 1.4. Previous cladograms of Triassic archosaurs. All cladograms except that of Gauthier (1986) are recalculated from the original data matrices with PAUP*4.0b3a (Swofford 1999) using the branch-and-bound method (all options set to default). The robustness of nodes was assessed using bootstrapping (heuristic 1,000 replicates): nodes achieving bootstrap values of less than 50% are collapsed; nodes supported in more than 50% of bootstrap replicates are retained, and the percentage occurrence is indicated. Note: The 50% bootstrap value has no particular significance; it is merely a convention returned by the default settings on PAUP. Nodes with bootstrap values less than 80% should probably be regarded as not robust.
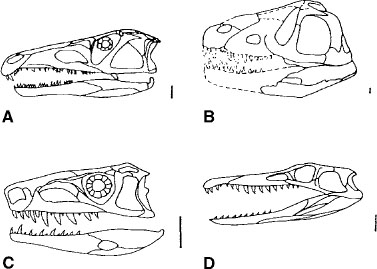
FIGURE 1.5. Skulls of basal archosaurs: A, Proterosuchus; B, Erythrosuchus; C, Euparkeria; D, Chanaresuchus. The skulls are drawn to uniform length. Scale = 20 mm. (From Benton and Clark 1988.)
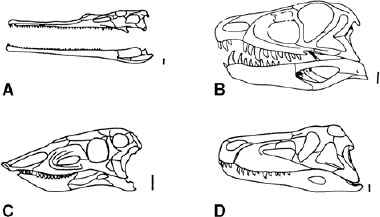
FIGURE 1.6. Skulls of Triassic archosaurs: A, Parasuchus; B, Ornithosuchus; C, Stagonolepis; D, Saurosuchus. The skulls are drawn to uniform length. Scale = 20 mm. (From Benton and Clark 1988.)
In the bird line, a number of genera and suprageneric taxa are treated as terminal taxa. Among them are Scleromochlus (fig. 1.7A; Benton 1999) and Pterosauria (Wellnhofer 1978; Wild 1978a). Sereno (1991a) gave a node-based definition of Pterosauria in which he listed nine genera and Pterodactyloidea, as well as all descendants of their common ancestor. However, such a node-based definition cannot accommodate wing-bearing forms more basal than Eudimorphodon, Peteinosaurus, and Preondactylus, pterosaurs known from the Late Triassic. Hence, an apomorphy-based definition takes into account the fact that pterosaurs have always been recognized on the basis of many clear apomorphies, and such a definition will maintain the concept that is accepted by all. Thus, Pterosauria includes those taxa stemming from the first archosaur with a long fourth finger (manual digit IV) supporting a wing membrane. Several dinosauromorph species follow: Lagerpeton chanarensis (Sereno and Arcucci 1994) and Marasuchus talampayensis (Sereno and Arcucci 1994; fig. 1.7B). The latter is the sister group to Dinosauria.
The cladistic analysis conducted here takes the same form as that presented previously. The 95-character data matrix was analyzed with PAUP*4.0b3a (Swofford 1999) using the branch-and-bound method, with the characters unweighted and unordered. The resulting 18 most parsimonius trees are 172 steps long and have a consistency index of 0.587, a retention index of 0.819, and a rescaled consistency index of 0.481. The 18 trees were combined to produce a strict consensus tree, a 50% majority-rule consensus tree, and an Adams consensus tree (fig. 1.8A), which was assessed by bootstrapping (1,000 replicates) for the robustness of nodes. Much of the crurotarsan clade collapsed at this point (fig. 1.8B) because most nodes were supported only in fewer than 50% of bootstrap replicates. All other nodes survived in 50% or more of bootstrap replicates, but only an unnamed node (Erythrosuchus plus more derived archosaurs) (100%), Archosauria (99%), Dinosauromorpha (99%), Dinosauria (98%), Avemetatarsalia (98%), Crurotarsi (93%), Saurischia (88%), Avesuchia/crown group Archosauria (84%), and Dinosauriformes (84%) achieved values greater than 80%. The other clades, an unnamed node comprising Proterochampsidae plus more derived archosaurs (75%), Prestosuchidae (74%), an unnamed node comprising Euparkeria plus more derived archosaurs (62%), Ornithodira (60%), and an unnamed node comprising “Rauisuchia” (53%) were not so well supported. Nodal supports (i.e., diagnoses) are provided in the appendix.
Monophyly of Archosauria
Eight diagnostic characters for Archosauria were noted by Benton (1990a), derived from Benton (1985), Gauthier (1986), and Benton and Clark (1988). Five of these are now rejected; one, bilaterally compressed marginal teeth, is seen also in Prolacerta and probably applies to a larger group, and the other four are hard to code (Sereno 1991a; Juul 1994). Benton's (1990a) character 6 (loss of trunk intercentra) applies to a subclade of Archosauria because Proterosuchus and erythrosuchids retain intercentra in the trunk region (Gower 2003). Benton's (1990a) character 8 (fourth trochanter on the femur) also refers to a more restricted group since Proterosuchus lacks this feature, as do erythrosuchids (Juul 1994; Gower 2003), contrary to Parrish's (1992) claim. Benton's (1990a) character 7 (lateral epicondylar foramen absent on humerus) is also uncertain; Juul (1994:3–4) notes a lateral epicondylar groove in a proterosuchid and in Stagonolepis. Three additional characters (4–6), from Juul (1994) and Gower and Sennikov (1996), are added.
The following are the hard-part synapomorphies of Archosauria sensu Benton 1999:
1. An antorbital fenestra
2. The postfrontal reduced to less than half the dimensions of the postorbital or absent
3. An ossified laterosphenoid
4. The prootic midline contact on the endocranial cavity floor
5. Teeth with serrated margins
6. A lateral mandibular fenestra
Proterosuchus is the basal archosaur (fig. 1.8), as indicated by nearly every previous author in the field. Erythrosuchus, Euparkeria, and Proterochampsidae are confirmed as outgroups of Avesuchia/crown group Archosauria, the conclusion of all previous authors except Gauthier (1986), who included Euparkeria in Ornithosuchia. Nearly all previous authors placed Erythrosuchus as more basal than Euparkeria and Proterochampsidae, but there has been uncertainty about the order of the last two. Benton and Clark (1988) and Bennett (1996) found Proterochampsidae to be the more basal outgroup, while Sereno (1991a) and Juul (1994) found Euparkeria to be more basal, the conclusion reached here. Characters pertaining to each of the successively more inclusive clades between Archosauria and Avesuchia are listed in the appendix.
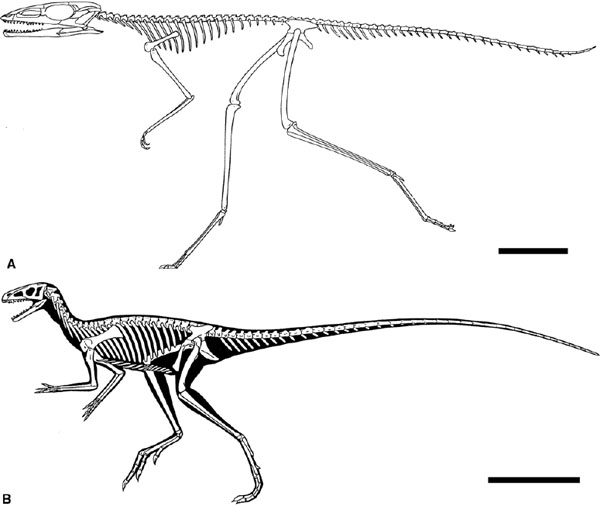
FIGURE 1.7. Basal avemetatarsalians: A, Scleromochlus; B, Marasuchus. Scale = 20 mm. (From Benton 1999; and Sereno and Arcucci 1994.)
Avesuchia/crown group Archosauria is the taxon comprising Avemetatarsalia and Crurotarsi (and sister taxa of Crurotarsi that are closer to Crocodylia than to Aves) and all their descendants. It is a strong clade, supported in 84% of bootstrap replicates (fig. 1.8B). The clade has six postulated synapomorphies: teeth on the palatine and vomer absent; manual digit IV having four or fewer phalanges; the ventral astragalocalcaneal articular facet larger than the dorsal articulation; the calcaneal tuber oriented more than 45° caudolaterally; the articular surfaces for the fibula and distal tarsal 4 on the calcaneum continuous; and the articular facet for metatarsal V on distal tarsal 4 less than half of the lateral surface of distal tarsal 4 (Sereno 1991a; Juul 1994). Avesuchia was given a node-based definition by Benton (1999: 1440) that is modified by the parenthetical insertion because both included taxa should be stem-based.
Crurotarsi is supported even more convincingly, surviving in 93% of bootstrap replicates. The clade has seven postulated synapomorphies: a scapulocoracoid notch at the cranial junction of the scapula and coracoid; the pubic acetabular margin recessed; the astragalar tibial facet flexed; the calcaneal tuber shaft broader than tall; the calcaneal tuber distal end rounded; the calcaneal tuber distal end having a vertical median depression; and a hemicylindrical calcaneal condyle for articulation with fibula. Crurotarsi was given a node-based definition by Sereno (1991a:27) that is retained with many reference taxa in view of the uncertainties about relationships within Crurotarsi.
Crurotarsi includes Phytosauridae, Ornithosuchidae, Prestosuchus, Suchia, and all descendants of their common ancestor. Relationships within Crurotarsi, however, are not well established. The Adams consensus tree (fig. 1.8A) shows a pairing of Phytosauridae + Gracilisuchus, which forms an unresolved tritomy with Ornithosuchidae and Suchia. Within Suchia, Postosuchus is paired with Crocodylomorpha, a relationship discovered by Benton and Clark (1988), and these two form an unresolved tritomy with Stagonolepididae and “Rauisuchia.” “Rauisuchia” includes Fasolasuchus as the outgroup to Prestosuchidae (Ticinosuchus, Prestosuchus, Saurosuchus). After boot-strap assessment of robustness, most of this structure within Suchia is lost, the only survivors being “Rauisuchia,” at a low bootstrap value of 53%, and Prestosuchidae, at 74% (fig. 1.8B). This phylogeny confirms that Ornithosuchidae are crurotarsans, as postulated by Sereno (1991a) and followed by Juul (1994) and Gower and Wilkinson (1996), rather than ornithodirans, as initially proposed by Gauthier (1986) and Benton and Clark (1988). There is no clear evidence that Phytosauridae is the basal crurotarsan group as suggested previously (Gauthier 1986; Benton and Clark 1988, Sereno 1991a; Parrish 1993). It should be noted that the crurotarsan portions of previous cladograms are equally poorly supported, although these earlier authors did not provide bootstrap or other tests (fig. 1.4). Parrish (1993) provided a detailed cladogram of Crurotarsi, but bootstrap testing and Bremer support values (Gower and Wilkinson 1996) show similar weaknesses.
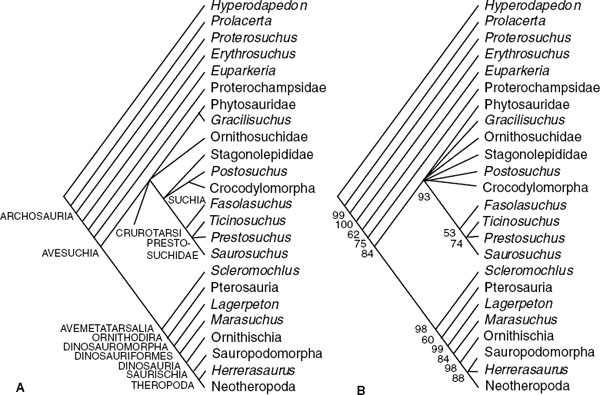
FIGURE 1.8. Cladogram of Triassic archosaurs, based on 24 taxa and 95 characters. Tree length = 172, CI = 0.5872, RI = 0.8193, RCI: 0.4811. A, Adams consensus of the 18 most parsimonious trees, showing major clades. B, Bootstrapping (heuristic 1,000 replicates) determined weak nodes (less than 50% of replicates; collapsed) and stronger nodes (greater than 50% of replicates; marked with bootstrap percentage). Note: The 50% bootstrap value has no particular significance; it is merely a convention returned by the default settings on PAUP. Nodes with bootstrap values less than 80% should probably be regarded as not robust.
Analyses have been much more informative with regard to the bird line. The clade made up of Scleromochlus+ Ornithodira, named Avemetatarsalia by Benton (1999), comprises all avesuchians/crown-group archosaurs closer to Dinosauria than to Crocodylia. It is well supported (98% of bootstrap replicates), and relationships within the clade are clearer than those within Crurotarsi (fig. 1.8). There are seven postulated synapomorphies: the interclavicle absent; the ratio of forelimb length to hindlimb length less than 0.55; manual digits I–III having a long penultimate phalanx with trenchant unguals; the ratio of tibia length to femur length more than 1.0; the fibula tapering and the calcaneum reduced in size; a compact metatarsus, with metatarsals I–IV tightly appressed; and the length of metatarsals II–IV more than 50% that of tibia. In addition, dermal armor is absent. The elongate hindlimb with elongate tibia and foot and the appressed elongate metatarsus are striking characters of Scleromochlus (fig. 1.7A) and other avemetatarsalians. An alternative name, Ornithosuchia Huene, 1907–8, defined by Gauthier (1986) as referring to the taxon that includes extant birds and all extinct archosaurs that are closer to birds than they are to crocodiles, might have been used here. However, Gauthier and de Queiroz (2001) reject that definition since Ornithosuchia would now have to exclude the species on which the name was based, namely, Ornithosuchus longidens (Huxley 1877), which is closer to crocodiles than to birds, according to current cladograms. Hence, they erect Panaves to refer to the most inclusive clade containing Aves (Vultur gryphus Linnaeus, 1758) but not Crocodylia (Crocodylus niloticus Lauenti, 1768), but this clade is equivalent to Avemetatarsalia, introduced as a stem-based taxon by Benton (1999) for Ornithodira and its close outgroups.
Ornithodira—Pterosauria, Dinosauromorpha, their most recent common ancestor, and all descendants—is supported in 60% of bootstrap replicates. It is supported by three apomorphies: the deltopectoral crest on the humerus subrectangular; the astragalar caudal groove absent; and the calcaneal tuber rudimentary or absent. The clade has been given a node-based diagnosis by Sereno (1991a:34), but he included Scleromochlus, seen by him as possibly the sister group of either Pterosauria or Dinosauromorpha. In the diagnosis given here Scleromochlus may be included or excluded, depending on its position in the cladogram: here it lies outside Ornithodira.
Dinosauromorpha refers to the clade comprising Neornithes and all ornithodirans closer to Neornithes than to Pterosauria. It is a more robust clade, supported in 99% of bootstrap replicates. The clade has at least six synapomorphies (appendix): the subrectangular and distinctly offset femoral head; the astragalar craniomedial corner acute; the astragalar ascending process on the cranial face of the tibia; the calcaneal distal articular face less than 35% of that of the astragalus; the articular facet for metatarsal V less than half of the lateral surface of distal tarsal 4; the midshaft diameters of metatarsals I and V less than those of II–IV; and metatarsal V having no “hooked” proximal end, with the articular face for distal tarsal 4 subparallel to the shaft axis. There are many equivocal synapomorphies at this node and the next two (appendix) because of incomplete knowledge of Lagerpeton and Marasuchus and uncertainty about the location in the cladogram where a number of dinosauromorph features were acquired. Dinosauromorpha was given a node-based definition by Sereno (1991a:23, modified slightly): Dinosauromorpha includes Lagerpeton chanarensis, Marasuchus talampayensis, Pseudolagosuchus major, Dinosauria, and all descendants of their common ancestor. A stem-based definition might be more appropriate because the basal forms are not well known.
Dinosauriformes is the name given the clade comprising Neornithes and all ornithodirans closer to Neornithes than to Lagerpeton. It is less well supported (in only 84% of bootstrap replicates), but Marasuchus and Dinosauria show parallelogram-shaped cervical centra, an acetabular antitrochanter, an articular surface on the femur that extends under the proximal head, and a weakly developed cranial trochanter on the femur, features that are absent in Lagerpeton. Dinosauriformes was given a node-based definition by Novas (1992b:56), who defined it as comprising the most recent common ancestor of Lagosuchus, Dinosauria, and all taxa stemming from it. Again, because the basal taxa are not well known, a stem-based definition is more appropriate.
Monophyly of Dinosauria
Dinosauria is a well-supported clade, present in 98% of boot-strap replications. It is diagnosed by many features, including loss of the postfrontal; an elongate deltopectoral crest on the humerus; a brevis shelf on the ventral surface of the postacetabular part of the ilium; an extensively perforated acetabulum; and a tibia with a transversely expanded subrectangular distal end, as well as a caudolateral flange, a depression for the astragalus, and an ascending astragalar process on its cranial face. There are probably further synapomorphies, but incomplete knowledge of Marasuchus makes the placement of many of these equivocal (appendix). These characters include several given by Gauthier (1986), Benton and Clark (1988), Novas (1996a), and others, but the lists have been reduced and revised. The elongate vomers, elongate scapula, symmetrical hand, and three or more sacral vertebrae were eliminated from the analysis because they are seen in other basal archosaurs or because they are not convincingly present in all dinosaurs (Herrerasaurus has two sacral vertebrae, while ornithosuchids, Scleromochlus, and Pterosauria apparently have three). Dinosauria has been given a node-based diagnosis by Sereno (1998:61).
Dinosauria consists of Triceratops, Neornithes, their most recent common ancestor, and all descendants. Within Dinosauria, Saurischia is defined as comprising all dinosaurs that are closer to birds than to Ornithischia (Gauthier 1986:15). Ornithischia comprises those dinosaurs closer to Triceratops than to Neornithes (Sereno 1998:61).
The Triassic and the Origin of Dinosaurs
The Triassic was a time of major upheaval in land faunas. After the massive extinctions at the end of the Permian period, some 251 million years ago, when as many as 50%–60% of tetrapod “families” died out (corresponding to perhaps 90% of species), ecosystems were devastated (Erwin 1993; Benton 2003). The complex Late Permian food webs, well known from both South Africa and from Russia, each comprised numerous herbivores and carnivores. Large herbivores such as pareiasaurs, tapinocephalid dinocephalians, and dicynodonts disappeared completely, and only two or three dicynodont species survived into the Triassic. Likewise, large carnivores such as gorgonopsians and titanosuchid dinocephalians also disappeared. Some of the smaller diapsids survived into the Triassic.
One of these was Archosaurus from the latest Permian of Russia. It is known from only fragmentary remains, but these show at least one diagnostic archosaur character, namely, the presence of an antorbital fenestra. Archosaurus is usually thought to be a proterosuchid, best represented by Proterosuchus from the Early Triassic of South Africa (fig. 1.5A), a 1.5 m long-snouted semiaquatic form. Proterosuchids were rare in the earliest Triassic, but the dicynodont Lystrosaurus dominated faunas to an astonishing extent, comprising more than 90% of specimens in faunas from South Africa and Antarctica and known also from South America, China, and Russia. Proterosuchids survived into the Early Triassic.
In the Early Triassic came two other basal archosaur groups: erythrosuchids, represented by Erythrosuchus (fig. 1.5B) and Euparkeria (fig. 1.5C), both from South Africa. Erythrosuchus and its relatives, such as Shansisuchus from China and Garjainia and Vjushkovia from Russia, were up to 5 m long, and they were clearly the top predators of their day. Euparkeria was a small animal that may have adopted a quadrupedal or bipedal posture. The final basal group, Proterochampsidae, represented by Chanaresuchus (fig. 1.5D) and Proterochampsa from the Middle Triassic of Argentina, comprises superficially crocodile-like fish eaters. These were restricted geographically and in numbers, and they did not radiate extensively.
Archosaur diversity increased substantially in the Middle Triassic (fig. 1.9) as the group expanded into new niches. Prestosuchids became ever larger, some of them reaching a length of 5 m, with high skulls armed with sharp teeth and an additional opening in the upper jaw between the premaxilla and maxilla, just below the external naris (fig. 1.6D). Some “rauisuchians” also show remarkable modifications in the pelvis and hindlimb, where the pillarlike femur fits straight up into a nearly horizontal acetabulum. Poposaurids, such as Postosuchus, were also quadrupeds, of similar large size but with unusual high, narrow skulls that apparently had some crocodile-like features in the caudal portion and a much-modified pelvis. “Rauisuchians” continued as major predators through the Late Triassic.
Other groups appeared in the Late Triassic (fig. 1.9). Phytosaurids were long-snouted animals measuring 2–4 m from the Late Triassic of Europe, North America, parts of Asia, and North Africa (fig. 1.6A). They looked superficially like modern crocodiles, but their crocodile-like features evolved convergently; for example, their external nares are contained in a bony mound between the eyes instead of at the tip of the snout as in crocodylomorphs. Ornithosuchidae, from the Late Triassic of Scotland (Ornithosuchus) and Argentina (Riojasuchus, Venaticosuchus), were facultatively bipedal carnivores measuring 1–3 m in length. The skull (fig. 1.6B) shows a bulbous snout and a gap in the tooth row between the premaxilla and maxilla. Stagonolepididae are a further distinctive group, unusually of herbivorous basal archosaurs, from the Late Triassic. They had low, blunt-ended snouts, peglike teeth (fig. 1.6C), and heavily armored, 1–3 m long bodies.
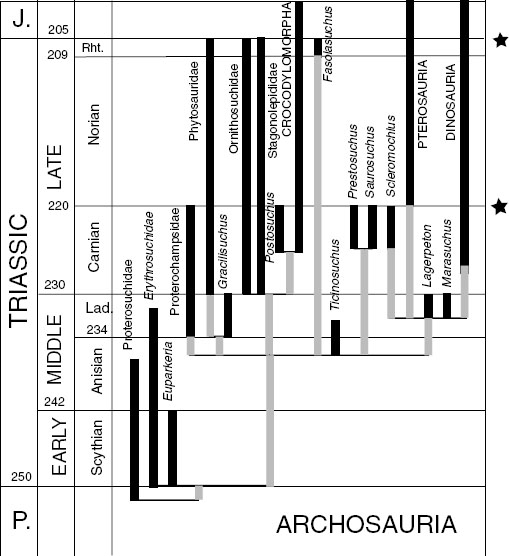
FIGURE 1.9. Evolution of the major archosaur groups through the Triassic, showing the major groups plotted against their occurrences in time (after Benton 1993), with postulated relationships based on the cladogram in figure 1.8. Stars mark the two postulated extinction events that affected archosaurian evolution.
Crocodylomorpha arose in the Late Triassic initially as small bipedal terrestrial forms, such as Terrestrisuchus and Saltoposuchus. These superficially uncrocodile-like animals already had the crocodylomorph hallmarks of elongate wrist bones and pneumatic cranial bones. A Late Triassic and Early Jurassic clade, sphenosuchids such as Sphenosuchus from South Africa (Walker 1990) were quadrupedal and probably fed on a mixed carnivorous diet of land animals and fishes.
The bird line of archosaur evolution includes principally the dinosaurs and the pterosaurs. The most basal avemetatarsalian, Scleromochlus from the Late Triassic of Scotland (Benton 1999), looks like a small dinosaur (fig. 1.7A), but it lacks the specialized ankle joint of pterosaurs, dinosaurs, and Marasuchus. It does have the characteristic digitigrade foot and tightly bound subequal metatarsals I–IV seen in all avemetatarsalians.
Pterosaurs are a distinctive clade of flying archosaurs with wings supported on their arms and an elongate fourth finger (Wellnhofer 1978). The oldest pterosaurs, such as Eudimorphodon from the Late Triassic of northern Italy, already show the wing fully developed, as well as numerous other pterosaur characters, such as the large, lightly built head, the caudally shifted orbit and bulbous temporal region, the shoulder girdle fused to the axial skeleton, the massive ossified keelless sternum, and the reduced pelvic girdle (Wild 1978a).
Basal Dinosauromorpha includes some small bipeds from the Middle Triassic of Argentina, Lagerpeton and Marasuchus. Lagerpeton is known only from the hindlimbs, the pelvis, and the trunk and caudal vertebrae (Sereno and Arcucci 1993). It has an unusual, functionally two-toed foot and was evidently an agile hopper or runner. Marasuchus is similar in size and adaptations (fig. 1.7B). It is known from partial skull remains and from much of the postcranial skeleton of several individuals (Bonaparte 1975a; Sereno and Arcucci 1994), which indicate an animal 1.3 m long that shares many features with dinosaurs. The oldest known dinosaurs are Eoraptor, Herrerasaurus, and Pisanosaurus from the Ischigualasto Formation of Argentina, dated as mid- to late Carnian, some 228 Ma old (Rogers et al. 1993). A number of other Carnian dinosaurs of similar age are known also from Brazil, India, Morocco, the southwestern United States (Coelophysis), and possibly Scotland (Saltopus).
There are currently two ways of viewing the radiation of the dinosaurs in the Late Triassic. Either they “took their chances” after a mass extinction event and radiated opportunistically or they competed over a longer timespan with the mammal-like reptiles, rhynchosaurs, and crurotarsan archosaurs, eventually prevailing. I suggest that the former was the case.
Most authors (e.g., Bakker 1972; Charig 1984) assumed that the competitive model was correct for four reasons. First, many considered dinosaurs to be a polyphyletic assemblage and hence believed that dinosaurs had arisen several times as a result of similar competitive pressures. Second, the origin of dinosaurs was seen as a drawn-out affair that started well down in the Middle Triassic and involved extensive and long-term competition. Third, dinosaur ancestors were regarded as competitively superior animals compared with the other members of their biota, with advanced locomotor adaptations (erect gait) or physiological advances (warm-bloodedness or cold-bloodedness: both cases have been argued), that progressively competed with all the mammal-like reptiles and crurotarsan archosaurs and caused their extinction. Fourth, it was generally assumed that major replacements in time were progressive, that evolution led ever onward and upward. Perhaps in some ways it does, but this assumption of progress meant that an a priori assertion was made that the new animals were better than the old ones and thus the replacement had happened competitively.
All four assumptions fail on close scrutiny: (1) Dinosauria is not polyphyletic; (2) dinosaurs diversified rapidly in the mid-Carnian, and not over a span of 10–15 Ma; (3) dinosaurs did not become abundant as soon as they appeared, but perhaps some 10 Ma later, and hence evidently did not demonstrate their competitive superiority; and (4) the argument that progress equaled competitive replacement is circular and requires some supporting evidence.
Several lines of evidence (Benton 1983b, 1986, 1994) suggest that dinosaurs radiated after ecospace had been cleared during an extinction event and that they did not establish their preeminence after a long period of competition with precursor groups:
1. The pattern in the fossil record does not support the competitive model. Dinosaurs appeared in the mid-Carnian or earlier and were rare elements in their faunas (1%–2% of individuals) until a major extinction event at the end of the Carnian, when various basal archosaurs and the dominant herbivore groups—dicynodonts, herbivorous cynodonts, and rhynchosaurs—all died out. Dinosaurs became more diverse and more abundant during the Norian, and all the major lineages appeared during this time. In Norian faunas dinosaurs represented 50%–90% of individuals, a dramatic increase from their pre-Norian totals. Dinosaurs diversified further in the Early Jurassic, after a second mass extinction at the end of the Triassic, when the remaining basal archosaurs and other groups died out.
2. The so-called superior adaptations of dinosaurs were probably not so profound as was once thought. Many other archosaurs (e.g., aetosaurs, “rauisuchians,” ornithosuchids, and some crocodylomorphs) also evolved erect gait in the Late Triassic, and yet they died out. The physiological characters of dinosaurs—whether they were warm-blooded or not, for example—cannot be determined with confidence. Dinosaurs were unique in being bipedal, but such a functional argument has not been put forth as the key driving force of active competitive replacement.
3. There is good evidence for other extinctions at the end of the Carnian and for environmental changes that may have mediated the changeover to dinosaur-dominated faunas. The Dicroidium flora of the Southern Hemisphere gave way to a worldwide conifer flora about this time. There were turnovers in marine communities, particularly in reefs, and there was a shift from pluvial climates to arid climates throughout much of the world (Simms and Ruffell 1990). The climatic and floral changes may have caused the extinctions of the dominant herbivorous tetrapods, opening the way for dinosaurian herbivores.
4. Many critical adaptations of the various dinosaur lineages were already established before dinosaurs diversified as the common large-bodied fauna. For example, the saurischians Eoraptor and Herrerasaurus and the ornithischian Pisanosaurus are known from the Carnian, thereby establishing the rudiments of the two great dinosaurian clades in the Carnian, well before cladal diversification took place.
5. The idea that simple competition can have major long-term effects in evolution is probably an oversimplification of a complex set of processes. Competition between clades is different from competition within or between species. In paleontological examples like this, competition has often been assumed to have been the mechanism, but the evidence has generally turned out to be weak.
The interval from the Carnian to the Norian has long been recognized as critical in the evolution of terrestrial vertebrates. Charig (1984), for example, noted the changeover from so-called paleotetrapods, essentially Paleozoic groups, to neotetrapods, or more modern forms, at this time. The last of the dicynodonts, basal archosauromorphs (rhynchosaurs, prolacertiforms, basal archosaurs), gave way to the new dinosaurs, pterosaurs, crocodylomorphs, turtles, and mammals. These new groups lived side by side with some surviving basal archosaurs—phytosaurs, stagonolepidids, ornithosuchids, and “rauisuchians”—and indeed the last two groups were the top predators, but they succumbed to the mass extinction at the end of the Triassic (fig. 1.9), after which larger carnivorous dinosaurs arose.
Appendix: Characters Diagnosing Clades
The following characters diagnose the clades in the cladogram shown in figure 1.8. The apomorphic condition (1) usually is not indicated, but in the case of multistate characters (numbers 9, 14, 41, 49, 53, 63, 64), the condition (1–4) is shown. Character state reversals are indicated by a negative sign. Equivocal character placements are indicated by an asterisk (*) at each possible node. The clade diagnoses include only the unequivocal characters.
Archosauria + Prolacerta: 13*, 26
Marginal teeth bilaterally compressed.
Archosauria: 5, 9 (1), 18, 23, 25, 29
Antorbital fenestra, postfrontal reduced to half the size of the postorbital, ossified laterosphenoid, prootic midline contact on endocranial cavity floor, teeth with serrated margins, lateral mandibular fenestra.
Erythrosuchus+ Euparkeria+ Proterochampsidae + Avesuchia: 5, 14, 15, 19, 24, 28, 35*, 40, 50, 58, 85, 86, 87
Antorbital fossa, parietal foramen absent, supratemporals absent, basisphenoid vertical, external abducens foramina on rostral surface of prootic, interclavicle lateral processes reduced, preacetabular process on iliac blade, ischium longer than iliac blade, centrale absent, distal tarsal 1 absent, distal tarsal 2 absent.
Euparkeria+ Proterochampsidae + Avesuchia: 20, 41 (l), 57, 62
Basisphenoid intertuberal plate absent, scapula more than twice the maximum width, pubic tuber strongly downturned, intertrochanteric fossa on ventral proximal femur absent.
Proterochampsidae + Avesuchia: −19, 21, 22, 34, −35*, 71
Basisphenoid horizontal, foramina for cerebral branches of internal carotid arteries lateral on basisphenoid, basisphenoid tubera with semilunar depression on lateral face, postaxial intercentra absent, crural facets on astragalus continuous.
Avesuchia: 27, 49 (l), 70, 79, 83, 88
Teeth on palatine and vomer absent, manual digit IV with four or fewer phalanges, ventral astragalocalcaneal articular facet larger than dorsal articulation, calcaneal tuber oriented more than 45° caudolaterally, articular surfaces for fibula and distal tarsal 4 on calcaneum continuous, articular facet for metatarsal V on distal tarsal 4 less than half of lateral surface of distal tarsal 4.
Crurotarsi: 16*, 33*, 35*, 42, 56, 72, 80, 81, 82, 84, 95*
Scapulocoracoid notch at cranial junction of scapula and coracoid, pubic acetabular margin recessed, astragalar tibial facet flexed, calcaneal tuber shaft broader than tall, calcaneal tuber distal end rounded, calcaneal tuber distal end with vertical median depression, hemicylindrical calcaneal condyle for articulation with fibula.
Phytosauridae + Gracilisuchus: 8*
Stagonolepididae + Crocodylomorpha + Ornithosuchidae + “Rauisuchia”: 19*, 68, −88
Fibular cranial trochanter robust and pendent; distal tarsal 4 broader than distal tarsal 3.
Ornithosuchidae + Crocodylomorpha + “Rauisuchia”: −49, −95
Five phalanges in manual digit IV, loss of osteoderm sculpture.
Ornithosuchidae + Postosuchus + Crocodylomorpha: 53 (1)*, 55*, 64 (2)*
Postosuchus + Crocodylomorpha: 13 (1)*, 17*, −35*
“Rauisuchia”: 3 Maxillary-premaxillary kinetic joint.
Prestosuchidae: 2*, 6, 10*, 38*, 52*
Antorbital fenestra triangular with elongate narrow rostral point.
Avemetatarsalia: 1*, 38*, 39, 41 (2)*, 43, 47, 59, 69, 90, 92*, 93
Interclavicle absent, forelimb-hindlimb ratio less than 0.55, manual digits I–III with a long penultimate phalanx with trenchant unguals, tibia-femur ratio equal to or more than 1.0, fibula tapering and calcaneum reduced in size, compact metatarsus with metatarsals I–V tightly bunched, metatarsals II–V more than 50% tibial length.
Omithodira: 32*, 44, 74, 78
Deltopectoral crest on humerus subrectangular, astragalar caudal groove absent, calcaneal tuber rudimentary or absent.
Dinosauromorpha: 9 (2)*, 53 (1)*, 60, 75, 77, 89, 91, 94
Femoral proximal head subrectangular and distinctly offset, astragalar craniomedial corner acute, calcaneal distal articular face less than 35% that of astragalus, articular facet for metatarsal V less than half of lateral surface of distal tarsal 4, midshaft diameters of metatarsals I and V less than those of II–V, metatarsal V has no “hooked” proximal end and articular face for distal tarsal 4 is subparallel to shaft axis.
Dinosauriformes: 31, 54, 61, 63, 64 (l)
Centrum shape of cervical centra 6–9 parallelogram-shaped, acetabular antitrochanter on ilium and ischium, femoral head articular surface extends under head, fossa trochanterica on proximal face of femoral head, femoral cranial trochanter weakly developed.
Dinosauria: 9 (2), 13*, 35*, 45, 49 (2)*, 51, 53 (2), 55*, 64 (2)*, 65*, 66, 67, 73
Postfrontal absent, deltopectoral crest elongate with apex situated at a point corresponding to more than 38% down the length of the humerus, brevis shelf on ventral surface of postacetabular part of ilium, acetabulum extensively perforated, tibial distal end transversely expanded with subrectangular end, tibia with caudolateral flange and receiving depression on dorsal aspect of astragalus, astragalar ascending process on cranial face of tibia.
Saurischia: 2*, 7, 8*, 36*, 46, 48, 76
Jugal overlaps lacrimal, robust saurischian thumb (metacarpal I robust and half or less the length of metacarpal II, first phalanx longer than metacarpal I or any other phalanx in the hand, ungual phalanx of digit I much larger than other unguals), metacarpal IV and V bases lie on palmar surfaces of manual digits III and IV, respectively, calcaneal proximal articular face concave.
Theropoda: 16*, 30, −69*
Intramandibular joint well developed.
Prolacerta: 32*
Proterosuchus: 17*
Erythrosuchus: 33*
Proterochampsidae: 9 (2)*, −58
Gracilisuchus: 8*, −9, 13 (1)*, 17*, 65*
Stagonolepididae: 11 *, 49 (2)*, 52*
Phytosauridae: 1*, 8*, 12 (1)*, −14, −26, 41 (2)*, −56, −82*
Fasolasuchus: 3*, 36*, 64 (1)*
Ornithosuchidae: −42, 65*, −82*
Postosuchus: 2*, 10*, 11*, 12 (2), −37, 38*, 39*, −68
Crocodylomorpha: 9 (2)*, −23, 92*, 95*
Scleromochlus: 1*, −58*, −69*, −78, 92*
Pterosauria: 1*, −58*, 92*
Marasuchus: 32, −41 (1)*, −60
Ornithischia: 19*, −47*
Herrerasaurus:−41 (1)*, −46, −51
Sauropodomorpha: 19*, −47*