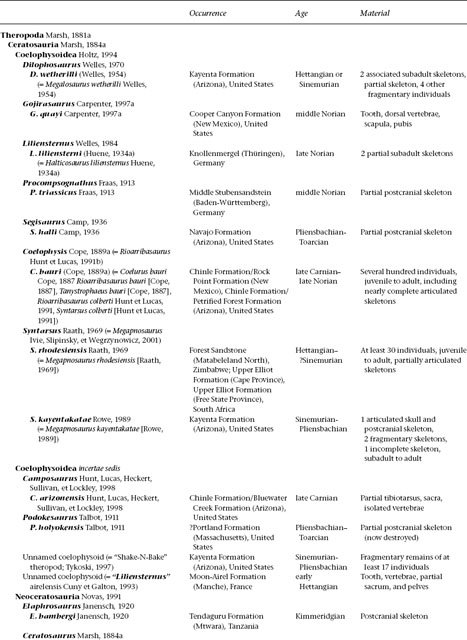
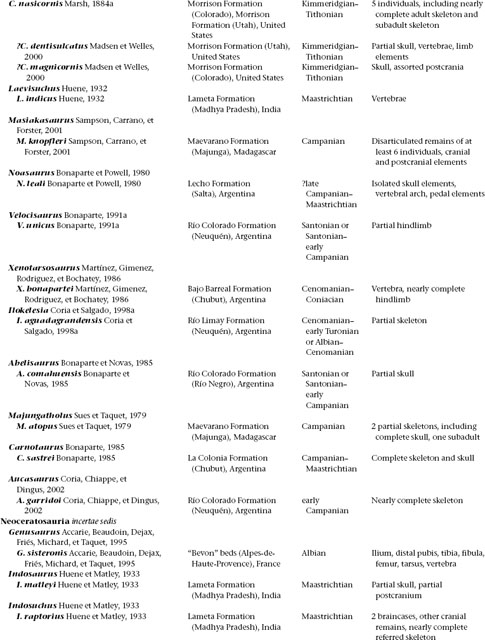
THREE

Ceratosauria
Ceratosauria represents the first widespread and diverse radiation of theropod dinosaurs (table 3.1). The remains of these predators are the most common theropod fossils recovered from Upper Triassic and Lower Jurassic deposits worldwide, and members of the clade evidently became dominant predators on the Gondwanan landmasses during Cretaceous time. Their fossils are known from Africa, India, Madagascar, North America, South America, and Europe. There is considerable debate over the phylogenetic relationships of the group (Gauthier 1986; Rowe 1989; Rowe and Gauthier 1990; Holtz 1994, 1998a; Rowe et al. 1997a; Rauhut 1998, 2000a; Tykoski 1998; Carrano and Sampson 1999; Forster 1999; Sereno 1999a; Carrano et al. 2002). Our phylogenetic analysis yields a monophyletic Ceratosauria comprising two main sister clades, Neoceratosauria and Coelophysoidea (Novas 1991, 1992b; Holtz 1994, 1998a; Padian et al. 1999).
Our knowledge of neoceratosaurs increased substantially over the past decade of research and discovery on Gondwanan continents. Most neoceratosaurs are known from Upper Cretaceous sediments, although the earliest records of the clade come from the Late Jurassic (Kimmeridgian–Tithonian) of western North America and eastern Africa. Several neoceratosaurs are large (up to 10 m) with a powerful build, and their large skulls are often adorned with pronounced cranial ornamentation (fig. 3.1). Some derived forms have extremely abbreviated forelimbs, a condition analogous to that seen in tyrannosaurids, alvarezsaurids, and some flightless birds.
The earliest coelophysoid record comes from the Late Triassic (Carnian–Norian) of North America and Europe. The group was diverse and widely distributed by Norian time, and their remains last appear in Lower Jurassic (Pliensbachian–Toarcian) deposits. Coelophysoids possessed slender overall proportions, with long necks and tails, narrow pelves, powerful forelimbs with grasping hands, long hindlimbs, and narrow, compact feet. They ranged in size from diminutive taxa 1 m long to forms greater than 6 m long. Some taxa sported elaborate cranial Ornamentation in the form of parasagittal crests on the dorsal skull surface (fig. 3.2). Interestingly, the Late Triassic Coelophysis bauri is one of the most derived coelophysoids, yet it is stratigraphically lower than more basal members of the lineage, suggesting that coelophysoids and hence Ceratosauria have a long history not reflected in the fossil record and that ceratosaurs may eventually be discovered in pre-Carnian deposits.
Definition and Diagnosis
A stem-based taxon defined as those theropods more closely related to Ceratosaurus nasicornis than to birds (Rowe 1989: 132), Ceratosauria can be diagnosed based on the following unambiguous apomorphies: axial neural spine extending cranially beyond the prezygapophyses; postaxial neural spines dorsoventrally low; transverse processes of dorsal vertebrae caudally backswept and triangular in the dorsal view; sacral ribs fused with the ilia; M. caudofemoralis brevis fossa of the ilium broad; supracetabular crest of the ilium flaring laterally and ventrally, overhanging much of the craniodorsal half of the acetabulum in lateral view; pubic-shaft axis bowing cranially; dimorphism in the femoral cranial trochanter; femoral medial epicondyle well developed and crestlike; tibiofibular crest of the distal femur sharply separated from the fibular condyle; astragalus and calcaneum fused to form an astragalocalcaneum in adults; distal tarsal 4 having a large rectangular notch in the caudolateral margin. The presence of two pleurocoels in the postaxial cervical and cranial dorsal vertebrae is also diagnostic for Ceratosauria. There is uncertainty whether this feature arose directly from a condition in which these pneumatic structures were lacking or from a condition in which there was one pleurocoel.
Anatomy
Recognition of an ontogenetic stage is critical to the proper anatomical comparison and subsequent interpretation of ceratosaur character states. Following earlier work (Gauthier 1986; Rowe1989; Rowe and Gauthier 1990), we recognize three broad, if arbitrary, stages of posthatchling ontogenetic development in these taxa. The stages are based on the sequence and degree of fusion between skeletal elements, as well as other size-independent indicators of an ontogenetic stage in fossil organisms (Brinkman 1988; Bennett 1993; Sampson 1993; Brochu 1996; Carr 1999). Juveniles range from hatchlings to near-adult-sized individuals but lack signs of cessation of growth; subadults are at nearly full size and show some but not all of the skeletal trans-formations that mark cessation of growth; and in adults the cessation of growth is indicated by a number of features. Adult features include closure of sutures in the braincase and the occipital condyle; fusion between the atlantal centrum, the axial intercentrum, and the axial centrum; fusion of the midcervical ribs to their respective vertebrae; complete closure between vertebral neural arches and centra; a fused scapulocoracoid; fusion of the sacral and pelvic elements; fusion of the proximal tarsals to form an astragalocalcaneum; and in some coelophysoids, complete fusion of distal tarsal 3 to its metatarsal and proximal fusion of metatarsals II and III to each other.
TABLE 3.1
Ceratosauria
Most adult neoceratosaurs are medium- to large-sized theropods, some probably attaining a length of 10 m. Coelophysoids are generally small- to medium-sized theropods, with most of the well-represented taxa 1–3 m in length, although at least one taxon (Dilophosaurus) rivals some neoceratosaurs in length. Coelophysoid adults exhibit pronounced dimorphism in those taxa known from samples of adequate sizes. It has been suggested that this is an expression of sexual differences (Colbert 1990; Raath 1990). Robust individuals have shorter skulls and necks, more pronounced muscular insertions and processes on the limb bones, and more pronounced co-ossification in the pelves and tarsus. Gracile individuals have longer skulls and necks, lack hypertrophied muscular attachments, and exhibit less pronounced skeletal fusions. These differences are observed in adult individuals of comparable size, arguing against interpretation that gracile individuals are merely juveniles.
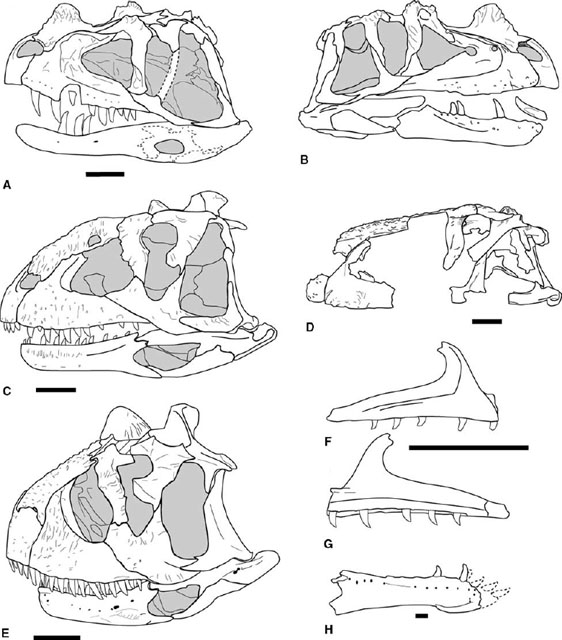
FIGURE 3.1. Neoceratosaur skulls. A, B, Ceratosaurus nasicornis: A, left lateral, and B, right lateral views; C, Majungatholus atopus; D, Abelisaurus comahuensis; E, Carnotaurus sastrei; F, G: right maxilla of Noasaurus leali in F, lateral, and G, medial views; H, right dentary of Masiakasaurus knopfleri. Scale = 10 cm (A–E), 5 cm (F, G), 1 cm (H). (A, B, after Gilmore 1920; C after Sampson et al. 1998; D after Bonaparte and Novas 1985; E after Bonaparte et al. 1990; F, G, after Bonaparte 1991b; H after Sampson et al. 2001.)
Skull and Mandible
Neoceratosaur and coelophysoid skulls differ in overall form. The external cranial bones of abelisaurids are marked by extensive pitting and sculpturing, but no such sculpturing is found in Ceratosaurus or coelophysoids (figs. 3.1, 3.2). Neoceratosaur skulls are large and dorsoventrally deep with short preorbital proportions and a broad snout. The infratemproal fenestra is also large, often twice or more the size of the orbit. Coelophysoids have low, long skulls that taper rostrally to a narrow tip. The length of the antorbital fenestra is equal to 25% or more of the length of the skull in Coelophysis, Syntarsus, and Dilophosaurus wetherilli. Many, if not most, ceratosaurs bore some cranial ornamentation. A medial, flattened nasal horn is known in Ceratosaurus, as are hornlets arising from the lacrimals (Marsh 1884a; Gilmore 1920). Carnotaurus bears a pair of robust supraorbital horns (Bonaparte et al. 1990). Majungatholus atopus possesses a single median frontal dome (Sampson et al. 1998). Dilophosaurus has some of the most flamboyant of theropod headgear in the form of delicate, paired parasagittal crests (Welles 1984). Smaller parasagittal crests arise from the nasals of Syntarsus kayentakatae (Rowe 1989). Coelophysis lacked dorsally located cranial ornamentation, but it bore laterally raised ridges along the dorsolateral margin of the lacrimal and nasal (Colbert 1989).
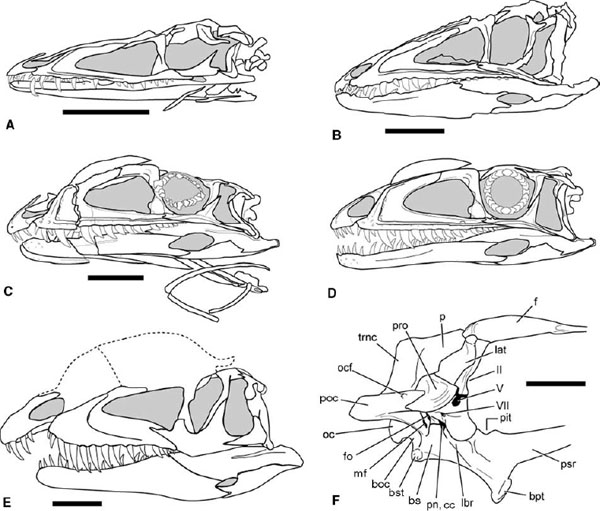
FIGURE 3.2. Coelophysoid skulls in lateral view: A, B, Coelophysis bauri (B is reversed); C, D, Syntarsus kayentakatae; E, Dilophosaurus wetherilli; F, Syntarsus rhodesiensis, braincase in right lateral view. Scale = 5 cm (A–D), 10 cm (E), 2 cm (F). (A, B, after Colbert 1989; C, D, after Tykoski 1998; E after Welles 1984; F after Raath 1977, 1985.)
The premaxilla in neoceratosaurs is deep below the external naris, especially in the abelisaurids, in which the rostral and caudal margins of the bone can be nearly parallel. The maxillary and palatal processes are reduced in size. The premaxilla of Ceratosaurus contains only three teeth, an unusual number for theropods. In coelophysoids the premaxilla is long and low, with an elongate maxillary process that loosely overrides the premaxillary process of the maxilla. There may have been some degree of mobility between these elements. The body of the premaxilla and hence the alveoli lie entirely rostral to the border of the external naris in the latter clade.
The maxilla is dorsoventrally deep in the larger neoceratosaurs (fig. 3.1) but less so in the noasaurids (fig. 3.1F, G). In lateral view the alveolar border is ventrally convex, except in Noasaurus, in which the alveolar border is concave (Gilmore 1920; Bonaparte and Powell 1980; Bonaparte and Novas 1985; Bonaparte et al. 1990; Bonaparte 1991b; Chatterjee and Rudra 1996; Sampson et al. 1998; Madsen and Welles 2000; Lamanna et al. 2002). The premaxillary (= rostral) process of the maxilla is short and dorsoventrally deep in Ceratosaurus and Majungatholus but virtually nonexistent in Carnotaurus and Abelisaurus, as well as in a maxilla from the Lameta Formation of India and a maxilla from the Bajo Barreal Formation of Argentina (fig. 3.1; Gilmore 1920; Chatterjee 1978b; Bonaparte and Novas 1985; Sampson et al. 1998; Bonaparte et al. 1990; Madsen and Welles 2000; Lamanna et al. 2002). The maxillary dorsal process is short and nearly vertical in some neoceratosaurs, with little or no caudal direction (fig. 3.1; Lamanna et al. 2002). Abelisaurids have a strongly reduced maxillary antorbital fossa, but large fossae are found in Ceratosaurus and Noasaurus. The ventral border of the antorbital fossa is marked by a short, caudoventrally dipping ridge in Noasaurus that is superficially similar to the alveolar ridge of some coelophysoids (fig. 3.1F). The rostrodorsal margin of the maxillary antorbital fossa in Ceratosaurus is penetrated by distinct, deep pneumatic fossae. There is a small promaxillary fenestra (Witmer 1997a, 1997b) partially hidden in lateral view in Carnotaurus and Abelisaurus, in a maxilla from the Bajo Barreal Formation of Argentina, and perhaps in Majungatholus (Bonaparte and Novas 1985; Bonaparte et al. 1990; Sampson et al. 1998; Lamanna et al. 2002).
The coelophysoid maxilla is long and low and bears a broad maxillary antorbital fossa (fig. 3.2; Raath 1977; Welles 1984; Colbert 1989; Rowe 1989). There is a small, low rostral process in Coelophysis and Syntarsus that protrudes sharply from the base of the dorsal process. The premaxillary process is upcurved rostrally in these taxa and in Dilophosaurus, meeting the maxillary process of the premaxilla in a high position. This relationship creates a subnarial gap, or diastema, in the upper tooth row (Welles 1984). The dorsal process is long and has a sharp caudal orientation. A small promaxillary fenestra is tucked into the rostroventral corner of the maxillary antorbital fossa in Dilophosaurus and Syntarsus kayentakatae (Welles 1984; Tykoski 1998). This opening is reportedly absent in Coelophysis and Syntarsus rhodesiensis (Raath 1977; Colbert 1989). Coelophysis, Liliensternus liliensterni, and Syntarsus bear a sharply raised alveolar ridge several millimeters above the tooth row (fig. 3.2A–D). The ridge begins at the rostroventral corner of the maxillary antorbital fossa, above the fourth maxillary tooth, and it continues for the length of the maxilla, paralleling the alveolar border. The ridge also marks the ventral border of the large external antorbital fenestra (sensu Witmer 1997a) in these taxa.
The lacrimal forms the caudal and part of the dorsal border of the antorbital fenestra in Ceratosaurus but only the caudal border in the abelisaurids. It is broadly exposed on the skull roof. In Ceratosaurus the lacrimal is dorsally expanded into a preorbital brow horn (fig. 3.1A, B). The rostral (= nasal) process of the lacrimal is greatly reduced or nearly absent in Majungatholus and Carnotaurus (fig. 3.1C, E). The abelisaurid lacrimal is also distinctive for possessing a suborbital flange that intrudes caudally into the orbital border. Caudally and dorsally the abelisaurid lacrimal makes contact with the postorbital, dorsally roofing the orbit (fig. 3.1C–E). In coelophysids the lacrimal has a long rostral process, as long as or longer than its ventral process (fig. 3.2). The ventral ramus bears a large, triangular lacrimal antorbital fossa in Syntarsus and in some Coelophysis. In Dilophosaurus wetherilli the lacrimal evidently contributes significantly to the parasagittal crests (Welles 1984).
The nasals of neoceratosaurs are modified in a variety of ways. Protuberances of the right and left nasals fuse along the midline to form the nasal horn of adult Ceratosaurus (fig. 3.1A, B; Madsen and Welles 2000). The rostral end of the nasal is laterally convex caudal to the external naris. In abelisaurids the nasals are fused, have a rugose external texture, and border the internal antorbital fenestra (fig. 3.1C–E). In at least Majungatholus the nasals are filled with pneumatic spaces that communicate with a large foramen in the rostrolateral edge of the bone (Sampson et al. 1998). Coelophysoid nasals are thin and rarely preserved intact. They abut the prefrontals and frontals in a squamous articulation in Coelophysis and Syntarsus (Raath 1977; Colbert 1989; Rowe 1989). In Dilophosaurus the nasals reportedly contribute in part to the cranial crests (Welles 1984). The smaller cranial crests of Syntarsus kayentakatae are derived entirely from the nasals (Tykoski 1998). In both Syntarsus species the nasal, prefrontal, and frontal bound a diamond-shaped nasal fenestra on the dorsal skull surface, an opening absent in other ceratosaurs (Raath 1977; Rowe 1989). Coelophysis lacks dorsally placed cranial crests. However, the lateral margins of the nasals and perhaps also the lacrimals form low ridges over the antorbital cavity.
Neoceratosaur frontals are fused to each other and also to the unified parietals. In Carnotaurus sastrei the frontal forms large, laterally projecting supraorbital horns (Bonaparte et al. 1990). The frontal of Majungatholus bears a median bony dome or horn core; the isolated holotypic Majungatholus frontal dome was originally misidentified as belonging to a pachycephalosaur (Sues and Taquet 1979). The parietals in abelisaurids are fused, and between the supratemporal fenestrae they narrow to form a small sagittal crest. Caudal to this the parietals project sharply upward, forming a large parietal eminence. The eminence rises high above the dorsal skull roof in Carnotaurus and Majungatholus but less so in Abelisaurus (Bonaparte and Novas 1985; Bonaparte et al. 1990; Sampson et al. 1998). The large parietal trans-verse crests form a powerful nuchal crest. Coelophysoids are less derived with regard to these elements. The frontals abut one another, and each interdigitates with a parietal behind the orbit in Coelophysis and Syntarsus. There is a strongly interdigitating suture with the postorbital. The parietals form the roof of the neurocranium, where they only abut the laterosphenoids, the prootics, and the opisthotics ventrally. Caudally the parietals flare out to form large transverse parietal crests that in turn form the dorsolateral margins of the occipital plate.
The suborbital process of the abelisaurid postorbital projects rostrally into the orbital border, partially flooring the orbit (fig. 3.1C–E). Together with the suborbital flange of the lacrimal, it creates a keyhole-shaped orbital opening. The squamosal is small, has a rodlike ventral process, and slopes caudoventrally. The quadrate of Ceratosaurus and abelisaurids is tall and slopes caudoventrally, placing the quadrate-articular contact far caudal to the occipital condyle (Britt et al. 2000), which increases the breadth of the infratemporal fenestra, especially ventrally. The quadratojugal fuses to the lateral margin of the quadrate, and there is no quadrate foramen in the abelisaurids. Coelophysoids have a looser connection between the bones of the cheek region. The postorbital is triradiate, with a long, narrow ventral (= jugal) process. The jugal is also a triradiate element. Its rostral process does not reach the border of the antorbital fenestra in Coelophysis and Syntarsus, but it contributes significantly to the antorbital opening in Dilophosaurus. The jugal does contact the rim of the antorbital fossa in Syntarsus and Coelophysis. The jugal is marked laterally in the latter taxa by a raised ridge that is continuous with the alveolar ridge of the maxilla. The caudal process of the jugal is long and rodlike in Coelophysis and Syntarsus, an unusual condition within Theropoda (Tykoski 1998). The quadrate is tall, but it does not slope as caudoventrally as in neoceratosaurs.
The neoceratosaur palate is little known, with good three-dimensionally preserved material only known from Majungatholus (Sampson et al. 1998). However, a detailed description of this material has yet to be published. The coelophysoid palate is not much better known. Most data come from disarticulated Syntarsus rhodesiensis or from flattened Coelophysis (Raath 1977; Colbert 1989). Vomers are definitively known only from Coelophysis, in which they fuse rostrally as in most other theropods. The vomer, the maxilla, and the palatine form the borders of the rostral palatal vacuity. The ectopterygoid is expanded and bears a fossa on its ventral surface. The pterygoid is longitudinally braced along the midline. It projects far rostrally to meet the palatine and the vomer. The pterygoids are separated medially from each other for most of their length, perhaps contacting only at their rostral tips. Contact with the basipterygoid processes was apparently synovial and probably allowed some degree of flexibility.
The braincase is preserved in several neoceratosaurs. Indeed, Indosuchus raptorius is based on isolated braincase material, although more complete material has been referred to it (Huene and Matley 1933; Chatterjee 1978b; Chatterjee and Rudra 1996). However, few neoceratosaur braincases have been described at this time. The rostral surface of the braincase is ossified in Ceratosaurus, and a large interorbital septum is known (Madsen and Welles 2000). The paroccipital processes are large and sharply backturned, and they have expanded distal ends (Madsen and Welles 2000). In abelisaurids the processes are short and laterally directed (Bonaparte et al. 1990; Sampson et al. 1998). The basisphenoid of Ceratosaurus is not as dorsoventrally tall as in the tetanurans Allosaurus fragilis and Acrocanthosaurus atokensis (Stovall and Langston 1950; Madsen 1976a; Chure and Madsen 1998). Shallow lateral excavations on the basipterygoid processes lead up and under the prootic crest (i.e., crista prootica). Ventrally, the basisphenoidal recess (= basisphenoidal fontanelle) is small and shallow. The basioccipital, in addition to forming most of the occipital condyle, forms a broad surface ventral to the condyle as it descends to the basal tubera.
Among coelophysoids, the braincase is known in Dilophosaurus, Coelophysis, and both Syntarsus taxa (fig. 3.2F; Raath 1977, 1985; Welles 1984; Rowe 1989; Tykoski 1998). Study of the region is difficult in Coelophysis because of crushing. To date, few useful systematic data have been derived from this region. The use of high-resolution X-ray computed tomography (CT) scanning techniques (Rowe et al. 1995, 1999; Rowe et al. 1997b; Tykoski 1998) should enable more detailed examination of brain-cases inaccessible by standard methods.
The paroccipital process is formed mainly by the opisthotic, with a small contribution from the prootic rostrally and by the exoccipital caudally. The tympanic cavity, recessed deeply beneath the paroccipital process, is divided into rostral and caudal chambers by a curved septum, the crista interfenestralis, which descends from the prootic. Caudal to the septum is the large metotic fissure, passageway for cranial nerves IX, X, and XI. Rostral to the septum is a smaller depression that houses a pair of small foramina, the foramen ovalis dorsally and the foramen pseudorotunda ventrally (Raath 1985; Tykoski 1998). The basisphenoid is the largest element in the braincase. In Syntarsus the basipterygoid process is marked by a shallow lateral sulcus leading dorsally to a large foramen partially hidden by the over-lapping prootic crest (fig. 3.2F; Raath 1977, 1985; Tykoski 1998). CT scans of Syntarsus kayentakatae show that the foramen opens into a large pneumatic cavity. The internal carotid probably passed through this foramen as well. Ventrally the basisphenoid is excavated by a large but shallow basisphenoidal recess. Although a possible epiotic was reported in the dorsolateral wall of the braincase in Dilophosaurus (Welles 1984), no such element has been reported in any other ceratosaur.
The lower jaw of neoceratosaurs possesses a number of distinctive features. The dentary curves rostrodorsally in Carnotaurus (fig. 3.1E) and in a large individual of Ceratosaurus referred to C. dentisulcatus by Madsen and Welles (2000; see also Bonaparte et al. 1990). The noasaurid Masiakasaurus possesses a highly derived condition of the dentary that includes procumbent rostral dentition and pronounced heterodonty in the lower jaw (fig. 3.1H; Sampson et al. 2001). The dentary of abelisauroids has a loose articulation with the postdentary bones, with little overlap between elements at the inframandibular joint. The dentary also receives a rostral prong from the surangular in a distinctive dorsocaudal socket (fig. 3.1C, E, H; Bonaparte et al. 1990; Sampson et al. 1998; Sampson et al. 2001; Carrano et al. 2002). The external mandibular fenestra is large in abelisauroids (fig. 3.1C, E). This region is not well preserved in Ceratosaurus nasicornis (fig. 3.1A, B). An ossified hyoid corpus and a single pair of ceratobranchial rods are preserved in Carnotaurus (Bonaparte et al. 1990).
Coelophysoids also possess unique features in the lower jaw. The rostral tip of the dentary is dorsally raised, elevating the mesial three to four dentary teeth relative to the remaining tooth row (fig. 3.2B–E). The third or fourth dentary tooth is slightly enlarged in Syntarsus, Dilophosaurus, and Coelophysis (Raath 1977; Welles 1984; Colbert 1989; Rowe 1989; Tykoski 1998), presumably fitting into the subnarial gap in the upper jaw of these taxa. The external mandibular fenestra is large in Coelophysis and Syntarsus but small in Dilophosaurus (Raath 1977; Welles 1984; Colbert 1989; Rowe 1989). A single pair of long, curved ceratobranchial rods is known in Coelophysis and Syntarsus (fig. 3.2C).
The upper teeth of Ceratosaurus are large and bladelike, proportionally larger than those of the contemporaneous tetanuran Allosaurus (Gilmore 1920; Madsen and Welles 2000). The lower teeth are also large. In contrast, the tooth crowns of some abelisaurids (Majungatholus, abelisaurid maxillae from India and Argentina) are surprisingly low, their height only about 1.5 times the mesiodistal base width (Lamanna et al. 2002). The interdental plates are fused to one another in neoceratosaurs (as in some tetanuran taxa), and in at least some abelisauroids they also bear conspicuous striae (Sampson et al. 1996; Lamanna et al. 2002; Carrano et al. 2002). Dorsal to the interdental plates of Noasaurus and Masiakasaurus is a distinct longitudinal groove (fig. 3.1G). Masiakasaurus is unique among known theropods in having strongly procumbent, asymmetrical mesial dentary teeth (fig. 3.1H). This taxon and Noasaurus are also unusual in possessing ten or fewer maxillary teeth (Bonaparte and Powell 1980; Bonaparte 1991b; Sampson et al. 2001; Carrano et al. 2002).
Coelophysoids also have heterodont dentition, but not nearly to the extreme seen in Masiakasaurus. The premaxillary teeth in Coelophysidae, elliptical to nearly circular in cross section, show little, if any, curvature and bear few to no serrations. In Syntarsus kayentakatae the mesial two premaxillary teeth completely lack carinae, while the third and fourth premaxillary teeth bear faint mesial and distal carinae but no serrations (Tykoski 1998). Similar conditions are described for S. rhodesiensis and Coelophysis (Raath 1977; Colbert 1989). The most mesial dentary teeth of coelophysids are generally similar to the premaxillary teeth. In Dilophosaurus, serrations are on at least the second and third premaxillary teeth but absent from the fourth (Welles 1984). The maxillary and more distal dentary teeth are linguolabially flattened, strongly recurved, and serrated. The largest maxillary tooth lies in or near the fourth alveolus, with crown height diminishing distally. The most mesial maxillary tooth projects slightly rostrally from its alveolus, a consequence of the upturned ventral border of the premaxillary process. As reconstructed, the maxillary tooth row terminates before the orbit in Dilophosaurus wetherilli and Syntarsus rhodesiensis, but these reconstructions were based on disarticulated material and have not been confirmed in articulated material (Raath 1977; Welles 1984). The tooth row terminates below the orbit in all articulated coelophysoid skulls. Dilophosaurus is reported to have 12 maxillary teeth and 17 or 18 dentary teeth (Welles 1984). Syntarsus rhodesiensis and Syntarsus kayentakatae have 20 maxillary teeth, and the former has 25 dentary teeth; the number of dentary teeth in S. kayentakatae is not known (Raath 1977; Rowe 1989; Tykoski 1998). Coelophysis bauri has up to 26 maxillary and 27 dentary teeth (Colbert 1989).
Postcranial Skeleton
AXIAL SKELETON
At least 20 presacral vertebrae are known in Ceratosaurus, but there are breaks in the column (Gilmore 1920; Madsen and Welles 2000). Carnotaurus has 22 presacrals, the last with coossification between its postzygapophyses and the following sacral vertebra (Bonaparte et al. 1990). Sixteen to 17 presacrals are known for Elaphrosaurus bambergi, with gaps in both the cervical and dorsal sections (Janensch 1925). Twenty-four presacrals are reported in Dilophosaurus wetherilli, but 23 may be the actual number, as Welles (1984) identified only 4 sacral vertebrae. There are 23 presacrals in both Coelophysis and Syntarsus rhodesiensis, identified as 10 cervicals and 13 dorsals (Raath 1969, 1977; Colbert 1989). Other ceratosaurs are too incomplete to allow an accurate presacral vertebral count. Welles (1984) labeled the tenth cervical and first four dorsal vertebrae in Dilophosaurus as pectorals, recognized by the position of the parapophysis straddling the neurocentral suture. The same convention was used by Madsen and Welles (2000). This convention has not been widely adopted, and we divide the theropod presacral series into only cervical and dorsal regions. The division between the two is recognized by the shift of the parapophysis from the ventrolateral margin of the centrum to a position nearly or completely on the neural arch. Marked changes in vertebral and rib morphology roughly correspond to this transition.
Given the criteria above, ceratosaurs possess ten cervical vertebrae, with strongly offset cranial and caudal articular surfaces forming an S-curved neck. Masiakasaurus may be an exception, as the articular faces are not offset in that taxon (Sampson et al. 2001). The cervical centra of Ceratosaurus, abelisaurids, and Dilophosaurus are deeply concave caudally (Gilmore 1920; Welles 1984; Bonaparte et al. 1990). The cranial articular surfaces are flat to weakly convex in Ceratosaurus and Dilophosaurus (fig. 3.3A–D, M, N). The cranial convexity of the cervicals is more pronounced in Carnotaurus and Majungatholus, but the articulations between the cervical vertebrae are not as strongly developed as in many large tetanurans (fig. 3.3F–H; Gilmore 1920; Madsen 1976a; Welles 1984; Bonaparte et al. 1990; Britt 1991; Sampson et al. 1998). The cervical centra are weakly amphicoelous in Coelophysis, Elaphrosaurus, Liliensternus, and Syntarsus (fig. 3.3E, R).
Ceratosaurs possess a double pair of pleurocoels (= pneumatic fossa and/or foramen) in the cervical centra, one pair positioned cranially and the other caudally. Neoceratosaurs and coelophysoids differ in the structure and degree of development of the pleurocoels. Neoceratosaurs (except Elaphrosaurus) have two small foramina penetrating the lateral centrum surface leading to cavities within the centrum (fig. 3.3B–D, F–H). This structure is generally similar to that in tetanuran theropods, although there is only a single, larger foramen in the latter. The paired foramina are larger in the axis of Ceratosaurus and Carnotaurus than in subsequent cervicals (fig. 3.3B, F). No pneumatic foramina or fossae are found in the cervical centra of Masiakasaurus (Sampson et al. 2001; Carrano et al. 2002). In coelophysoids and Elaphrosaurus the pleurocoels are deep pockets and fossae excavated into the sides of the centra (fig. 3.3E, M–N, R). In at least cervicals 4–6 of Dilophosaurus the right- and left-side pleurocoels communicate with each other (Welles 1984; Rowe and Gauthier 1990). The borders of the caudal pleurocoel can sometimes be indistinct, leading to claims that this character is absent in some taxa (Colbert 1989; Cuny and Galton 1993). The cranial pleurocoel is often obscured in lateral view in individuals with fused cervical ribs (fig. 3.3H, R).
No proatlas arch is reported in any ceratosaur. The atlantal intercentrum is crescentic, bearing a deep fossa on its cranial surface for articulation with the occipital condyle. Neurapophyses are known in Carnotaurus, Ceratosaurus, Coelophysis, Majungatholus, Dilophosaurus, and Syntarsus (Gilmore 1920; Raath 1977; Welles 1984; Colbert 1989; Rowe 1989; Bonaparte et al. 1990; Madsen and Welles 2000). They are fused to the atlantal intercentrum in Carnotaurus and Majungatholus (Bonaparte et al. 1990). In adults the atlantal centrum fuses to the axial centrum to form a strong odontoid process, and the axial intercentum fuses to the axial centrum and the atlantal centrum. The axial transverse processes and diapophyseal facets are small in Carnotaurus and Ceratosaurus and absent in coelophysoids (Gilmore 1920; Bonaparte et al. 1990). Ceratosaurus and the abelisaurids also possess a pneumatic foramen (or foramina) in the axial neural arch caudal or caudoventral to the diapophysis, as do Tetanurae (fig. 3.3B, F; Madsen 1976a). Axial pleurocoels are absent in coelophysoids (fig. 3.3M). The axial neural spine is broad and bladelike, and there is no spine table. The axis bears a strong ventral keel in Ceratosaurus, Coelophysis, and Syntarsus (Gilmore 1920; Raath 1977; Colbert 1989; Tykoski 1998).
The postaxial cervical vertebrae differ greatly between most neoceratosaurs and coelophysoids. In Ceratosaurus the midcervical centra are roughly as long as they are tall, with sharply downturned transverse processes and tall neural spines (fig. 3.3C, D). The zygapophyses are large, and there are robust epipophyses projecting above and beyond the postzygapophyses. The centra become craniocaudally shorter, and the transverse processes become larger and more laterally directed along the cervical series. In Carnotaurus, Majungatholus, Noasaurus, and Masiakasaurus the cervical neural spines are small, but the epipophyses are exceptionally large, extending caudodorsally above the top of the neural spines (fig. 3.3G, H). Carnotaurus, Noasaurus, and reportedly Ilokelesia also possess cranially directed spines on the tips of the epipophyses (fig. 3.3H; Bonaparte and Powell 1980; Bonaparte et al. 1990; Coria and Salgado 1998b). The cervical neural arches of Ceratosaurus and the abelisauroids are perforated by additional pneumatic foramina. The degree of arch pneumaticity increases caudally into the dorsal vertebrae of abelisaurids, as expressed by increasing numbers of foramina and fossae in the arch ventral to the transverse processes.
The postaxial cervicals in Coelophysoidea and Elaphrosaurus are proportionally longer and lower than in most other theropods. The centra increase in length caudally to the sixth or seventh vertebra, which in Coelophysis and Syntarsus are three or more times longer than tall. The third cervical retains a strong ventral keel, but this feature weakens or disappears in successive vertebrae. The neural arches are low and elongate, with low neural spines that may run the length of the arch. In coelophysoids the transverse processes are directed ventrally and lateroventrally, forming broad triangular sheets lateral to the centrum. Large, pronglike epipophyses project caudally beyond the postzygapophyses (fig. 3.3R). In Coelophysis and Syntarsus large triangular openings on the lateral surfaces of the arch pedicles are craniolateral to the postzygapophyses (Colbert 1989; Tykoski 1998). Each opening leads to a large pneumatic cavity within the arch pedicle that is both lateral and parallel to the neural canal. Coelophysis bauri also has a cranial opening for the cavity, caudolateral to the prezygapophysis (Colbert 1989).
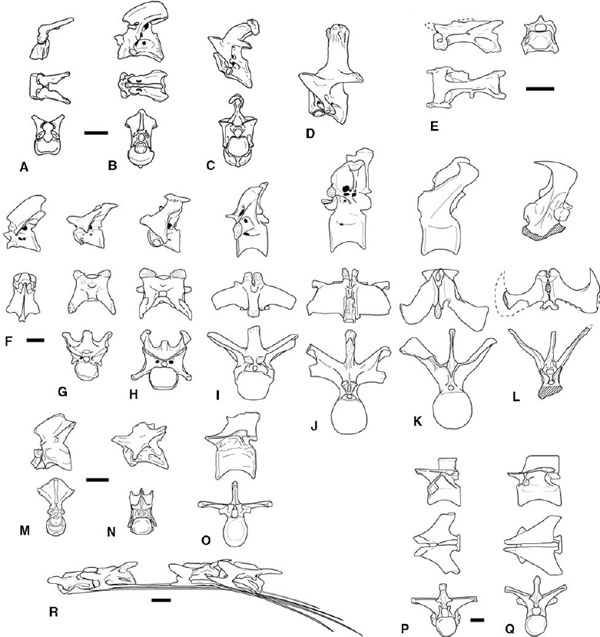
FIGURE 3.3. Vertebrae. A–D, Ceratosaurus nasicornis: A, atlas; B, axis; C, third cervical; D, sixth cervical. E, Elaphrosaurus bambergi, seventh cervical. F–L, Carnotaurus sastrei: F, axis; G, third cervical; H, sixth cervical; I, fourth dorsal; J, eighth dorsal; K, first caudal; L, sixth caudal arch. M–O, Dilophosaurus wetherilli: M, atlas and axis; N, sixth cervical; O, seventh dorsal. P–Q, Syntarsus rhodesiensis: P, second dorsal; Q, fifth dorsal; R, Syntarsus kayentakatae, third, fourth, sixth, and seventh cervicals and ribs. Scale = 5 cm (A–O), 1 cm (P, Q), 2 cm (R). (A–D after Gilmore 1920; E after Janensch 1925; F–L after Bonaparte et al. 1990; M–O after Welles 1984; P–Q after Raath 1977.)
Atlantal and axial ribs are found in Carnotaurus, Coelophysis, and Syntarsus (Rowe 1989; Bonaparte et al. 1990; Rowe and Gauthier 1990). They are single-headed, lack a tuberculum, and articulate weakly with the parapophyses. The atlantal and axial ribs are not preserved in Ceratosaurus, but the parapophysis is as developed as in Carnotaurus. The postaxial cervical ribs are bicapitate, with a large capitulum and smaller tuberculum. A sharp spine projects a short distance cranially beyond the rib heads. The cervical ribs are long and straight, extending caudally at least three vertebrae beyond their origin in Coelophysis, four in Carnotaurus, Majungatholus, and Syntarsus rhodesiensis, and five to six in Syntarsus kayentakatae (fig. 3.3R; Raath 1977; Colbert 1989; Bonaparte et al. 1990; Sampson et al. 1998; Tykoski 1998). The cervical rib shaft may be only 1–2 mm thick for most of its length. As many as three of these delicate shafts can over-lap to form rib bundles paralleling the cervical column. There is a distinct morphological change in the more caudal cervical ribs: they become shorter, thicker, and caudoventrally angled. The midcervical ribs fuse to their respective centra by adult-hood in Coelophysis, Syntarsus, Elaphrosaurus, and Ceratosaurus (fig. 3.3H, R; Janensch 1925; Raath 1977; Colbert 1989; Rowe 1989; Tykoski 1998; Madsen and Welles 2000:pls. 6, 15).
In addition to the dorsal shift of the parapophysis, dorsal vertebrae are marked by loss of the epipophyses, more laterally or dorsally directed transverse processes, and increased size of the neural spines. Ceratosaurus dorsal centra are about as long as the neural spines are tall. The parapophyses are borne at the ends of laterally projecting stalks in the cranial and mid-dorsal series of Ceratosaurus, Carnotaurus, and Majungatholus (fig. 3.3J; Gilmore 1920; Bonaparte et al. 1990; Britt 1991; Sampson et al. 1998; Madsen and Welles 2000). The dorsal transverse processes are subtriangular to strongly triangular in dorsal view in coelophysoids (fig. 3.3P, Q), probably Ceratosaurus (Britt 1991:fig. 25), and Majungatholus (O'Connor and Sampson 1998). The expression of this morphology varies through the dorsal series, especially caudally, where the parapophysis moves from the infradiapophyseal lamina to a position on the cranial edge of the transverse process. The centra of the caudal dorsals in Syntarsus and Coelophysis are elongate, being twice as long as tall (fig. 3.3Q). The dorsal ribs have a thin web of bone connecting the capitulum and tuberculum. In Ceratosaurus the proximal end of the rib is pierced medially by apneumatic foramen (Madsen and Welles 2000). Similar pneumatic foramina are found in Majungatholus (Sampson et al. 1998).
There are five vertebrae in the coelophysoid sacrum, six in Ceratosaurus and Elaphrosaurus, and either six or seven in Carnotaurus (fig. 3.4A, B, D; Gilmore 1920; Janensch 1925; Raath 1977; Colbert 1989; Rowe 1989; Bonaparte et al. 1990; Rowe and Gauthier 1990; Tykoski 1998). As in tetanuran theropods, the sacral count is increased from three to five by incorporation of a second caudosacral and a dorsosacral. The low presacral vertebral count in Carnotaurus (only 22 vertebrae) indicates that the most caudal dorsal was incorporated into the sacral series as a second dorsosacral (fig. 3.4A). The sacral count likely increased in a similar fashion in Ceratosaurus and Elaphrosaurus (Gilmore 1920; Janensch 1925). However, the position of vertebrae bearing both sacral ribs and transverse processes suggests that the sacral count was increased by incorporation of a third caudosacral (fig. 3.4A). The postzygapophyses of the last dorsal (presacral 22) in Carnotaurus show some co-ossification with the first in the sacral series, but the rest of the neural arch, the neural spine, and the centrum remain free (Bonaparte et al. 1990). Small juvenile Coelophysis and Syntarsus may have only four sacrals (Raath 1977, 1990; Colbert 1989). Reports of only four sacrals in Dilophosaurus and Liliensternus (Huene 1934a; Welles 1984; Cuny and Galton 1993) are based on subadult or incomplete individuals as indicated by other size-independent ontogenetic criteria.
Sacral ribs project from the craniodorsal margin of the centra (Raath 1969, 1977; Welles 1984; Tykoski 1998). Four sacral ribs are known in Carnotaurus, the first being the largest and the most ventrally positioned (fig. 3.4A; Bonaparte et al. 1990). In Syntarsus, Coelophysis, and an as yet unnamed taxon from the Kayenta Formation of Arizona the third sacral rib (from caudosacral 1) is the largest and most ventrally positioned (fig. 3.4B; Tykoski 1998). The transverse processes contact the ilia on all the sacral vertebrae (fig. 3.4A–C). The sacral ribs and transverse processes are connected by vertical, transversely oriented laminae that further strengthen the synsacrum (fig. 3.4A, B). The sacral neural arches are perforated by large pneumatic foramina between the vertical laminae in Carnotaurus (fig. 3.4A).
Adult sacrals are so extensively fused in ceratosaurs that sutures between centra are obliterated and only swellings indicate where articular surfaces meet. The diameters of the midsacral centra are strongly reduced in Carnotaurus and Ceratosaurus, and the middle of the sacrum arches dorsally in these taxa and Elaphrosaurus (Gilmore 1920; Janensch 1925; Bonaparte et al. 1990). The neural arches, neural spines, transverse processes, and sacral ribs coalesce over the length of the sacrum. In Elaphrosaurus, Syntarsus, and the aforementioned Kayenta Formation taxon the transverse processes of the sacrals form a single horizontal sheet of bone (fig. 3.4B, C; Janensch 1925; Raath 1969, 1977, 1990). The sacral ribs and transverse processes fuse extensively to the medial wall of the ilium. The resulting synsacrum is analogous to the condition in ornithurine birds.
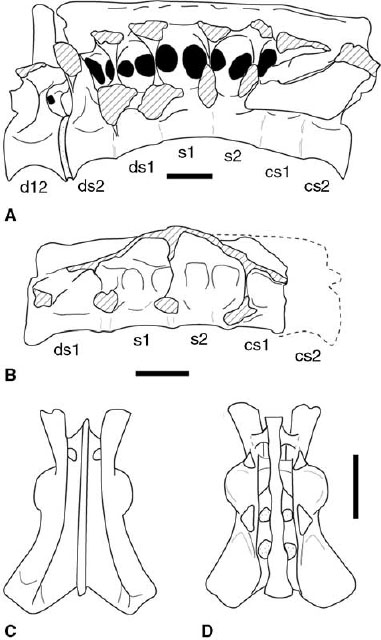
FIGURE 3.4. Sacra. A, Carnotaurus sastrei in lateral view; B, unnamed coelophysoid taxon from the Kayenta Formation of Arizona in left lateral view; C, D, Syntarsus rhodesiensis: sacrum and ilia in C, dorsal, and D, ventral views. Crosshatching indicates sectioned or broken bone. Scale = 10 cm (A), 1 cm (B), 5 cm (C, D). (A after Bonaparte et al. 1990; C, D, after Raath 1977, 1990.)
The exact number of caudal vertebrae is not known for most ceratosaurs. Coelophysis and Syntarsus are reported to have 40 caudals, 44 were preserved and three reconstructed in Dilophosaurus, and Ceratosaurus is estimated to have 50 (Gilmore 1920; Raath 1977; Welles 1984; Colbert 1989). The ventral surface of the proximal caudals bears a sharp longitudinal groove in Ceratosaurus, Dilophosaurus, Coelophysis, Syntarsus, Liliensternus, and Elaphrosaurus (Janensch 1925; Huene 1934a; Madsen 1976a; Raath 1977; Welles 1984; Rowe and Gauthier 1990; Britt 1991; Tykoski 1998). The neural spines and arches of Ceratosaurus are tall, and the chevrons are proportionally long, resulting in a deep lateral profile to the tail. The caudals of Ilokelesia and the abelisaurid Carnotaurus are highly distinctive in the form of the transverse processes. In the proximal caudals of these taxa and perhaps other abelisauroids the transverse processes are dorsolaterally directed, and the lateral tips bear cranially directed, spinelike processes (fig. 3.3K, L; Bonaparte et al. 1990; Coria and Salgado 1998a). When articulated, these tips may have over-lapped the transverse processes of preceding vertebrae, helping to brace the tail in a manner unique among theropods. The first caudal also bears these proximal projections, which, given the location of the vertebra, may have articulated with or over-lapped the notched, concave caudal rim of the ilium seen in Carnotaurus and Majungatholus (Bonaparte et al. 1990; Sampson et al. 1998).
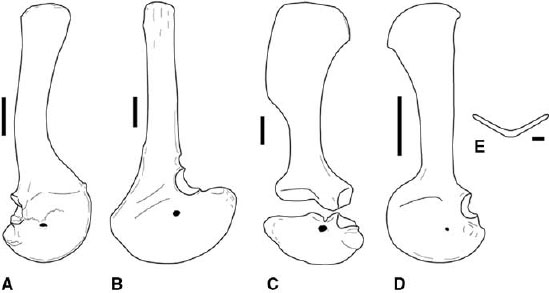
FIGURE 3.5. Pectoral girdles: A, Ceratosaurus, right scapulocoracoid in lateral view; B, Carnotaurus, left scapulocoracoid in lateral view; C, Dilophosaurus wetherilli, left scapulocoracoid in lateral view; D, Syntarsus kayentakatae, left scapulocoracoid in lateral view; E, Syntarsus kayentakatae, reconstructed furcula in cranial view. Scale = 10 cm (A, B), 5 cm (C, D), 1 cm (E). (A after Madsen and Welles 2000; B after Bonaparte et al. 1990; C after Welles 1984.)
Among coelophysoids the diameter of the caudal vertebrae diminishes distally, but the overall length of the centrum remains nearly the same. This results in middistal to distal caudals that can be as much as four times longer than tall. The neural arches and spines become reduced in size after approximately the fifteenth vertebra, and the neural spines shift to a position on the caudal portion of the arch. Midway down the caudal series the spines disappear, and the arches are composed of only a low neural canal and the zygapophyses. The proximal transverse processes are broader laterally than at their base, but distally they become craniocaudally narrower and shorter, diminishing and eventually disappearing, marking the transition point roughly halfway down the tail.
APPENDICULAR SKELETON
Among neoceratosaurs, complete pectoral girdles are described for Ceratosaurus, Carnotaurus, and Elaphrosaurus (Gilmore 1920; Janensch 1925; Bonaparte et al. 1990; Madsen and Welles 2000). The cranial margins of the scapula and coracoid and the distal end of the scapula are missing in Elaphrosaurus. The reconstruction of the missing parts (Janensch 1929a) is unusually broad and probably does not accurately reflect the morphology of these elements. The scapula and coracoid are fused in Ceratosaurus and Carnotaurus (fig. 3.5A, B). The cranial margin of the resulting scapulocoracoid is smoothly rounded, lacking any notch or interruption in the margin at the scapulocoracoid suture. The scapular blade is long and increases in craniocaudal width only slightly toward the distal end. In Ceratosaurus the axis of the scapular blade curves cranially, so it is concave cranially and convex caudally in lateral view (fig. 3.5A). There is a small acromion process in Ceratosaurus. The preglenoid fossa is strongly expressed, and the coracoid foramen is large and passes dorsomedially through the coracoid. The caudoventral process is short and bluntly rounded (fig. 3.5A). The scapulocoracoid of Carnotaurus also has an unexpanded blade (fig. 3.5B; Bonaparte et al. 1990). The proximal scapula and coracoid are greatly broadened, resulting in a large acromion process. The coracoid also has a large, pointed caudoventral process below the glenoid. A small piece of bone associated with the right scapulocoracoid of Carnotaurus was identified as a clavicle (Bonaparte et al. 1990).
The coelophysoid scapula and coracoid also fuse in adults, with visible sutures or separation of elements indicating immaturity (fig. 3.5C, D). The scapula has an expanded distal blade, but in Liliensternus, Gojirasaurus, Coelophysis, and Syntarsus the caudal margin of the blade is straight for most of its length, curving caudally only at the distal tip (fig. 3.5D; Huene 1934a; Raath 1969, 1977; Parrish and Carpenter 1986; Carpenter 1997a). The coracoid has a short, rounded caudoventral process below the glenoid fossa. The distal scapular expansion is uniquely rectangular in Dilophosaurus wetherilli (fig. 3.5C; Welles 1984). The clavicles are fused at the midline in Syntarsus, Segisaurus, and reportedly some Coelophysis, forming a furcula (fig. 3.5E; Tykoski 1998; Downs 2000; Senter and Hutchinson 2001; Tykoski et al. 2002). The presence of the furcula in coelophysoids represents the earliest known occurrence of this element in Theropoda.
The humerus is highly variable among neoceratosaurs (fig. 3.6A–C). In general, humeri are straight-shafted and lack most of the sigmoid curvature and torsion seen in coelophysoids and more derived tetanurans, such as Allosaurus (Janensch 1925; Madsen 1976a; Welles 1984; Bonaparte et al. 1990; Madsen and Welles 2000; Sampson et al. 2001). However, this straight-shafted condition is reminiscent of that in some basal tetanuran taxa, such as Torvosaurus and Spinosauridae. The humerus is narrow, with a small deltopectoral crest and a bulbous proximal head in Elaphrosaurus and Masiakasaurus (Janensch 1925; Sampson et al. 2001). The proximal and distal ends are not strongly expanded. The humerus was not recovered with the original material of C. nasicornis. However, later discoveries produced nearly complete humeri, although these have been assigned to two new Ceratosaurus taxa (fig. 3.6A, B; Madsen and Welles 2000). The proximal and distal ends are mediolaterally expanded, but the proximal end does not have the subspherical head seen in abelisaurids and to a lesser extent in Elaphrosaurus. The deltopectoral crest is longitudinally oriented and extends distally 40%–45% of the length of the humerus. An articulated lower forelimb element is known in Ceratosaurus nasicornis (fig. 3.6H; Gilmore 1920). The radius is robust throughout its length, with little additional expansion at either end. It does turn slightly medially toward its distal end. The ulna is missing most of its middle section as preserved but bears a strong olecranon process.
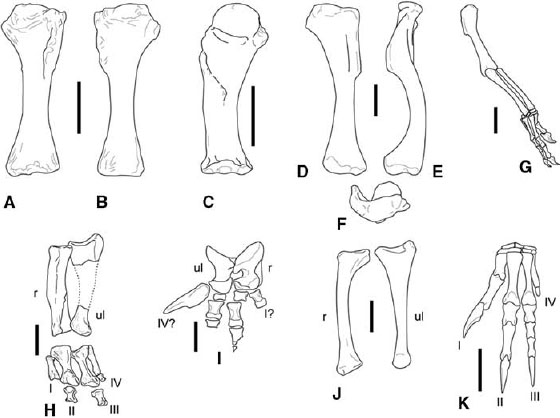
FIGURE 3.6. Forelimbs. A, B, Ceratosaurus: left humerus in A, cranial, and B, caudal views; C, Carnotaurus, right humerus in cranial view; D–F, Dilophosaurus wetherilli: left humerus in D, cranial, E, lateral, and F, proximal views; G, Coelophysis bauri, small right forelimb in craniolateral view; H, Ceratosaurus nasicornis, articulated left forearm and manus elements in dorsal view; I, Carnotaurus sastrei, forearm and manus elements in caudal view; J, Dilophosaurus wetherilli, left radius in lateral view and left ulna in medial view; K, Syntarsus rhodesiensis, left manus in dorsal view. Scale = 10 cm (A–C), 5 cm (D–F, H, I), 2 cm (G, J, K). (A, B, after Madsen and Welles 2000; C, I, after Bonaparte et al. 1990; D–F, J, after Welles 1984; H after Gilmore 1920; K after Galton 1971a.)
An articulated partial manus of Ceratosaurus nasicornis (fig. 3.6H; Gilmore 1920) does not include ossified carpals. The first metacarpal is short and narrow, unlike the proportions seen in the same element of coelophysoids and tetanurans. The second and third metacarpals are the most robust in the hand, with the second slightly longer than the third. The fourth metacarpal is reduced to only about 75% the length of metacarpal III. Its proximal end is closely appressed to metacarpal III. Proximal phalanges were recovered on the second, third, and fourth digits. The phalanges on digits II and III are short, blocky, and subequal in length. The first phalanx of digit IV is reduced to a small nubbin of bone, although it apparently retains a distinct proximal condyle for articulation with its metacarpal (Gilmore 1920).
The entire forelimb of Carnotaurus is highly apomorphic, but future discoveries will show whether its form is shared by other abelisaurids. The humeral shaft is straight (fig. 3.6C; Bonaparte et al. 1990). The proximal end is large, subspherical, and set off from the shaft by a short neck. The deltopectoral crest is low but powerfully constructed, traverses the cranial surface of the shaft obliquely, and extends more than half the length of the humerus. The distal end bears well-defined, flattened articular facets for the highly derived radius and ulna. The radius and ulna are short, only about one-fourth the length of the humerus (fig. 3.6I). The ulnar proximal end is deeply concave for articulation with the humerus, and the olecranon process is strongly developed. The ulna constricts to a short shaft, then widens distally to form a strongly rounded condyle for articulation with the carpus. The radius is slightly shorter than the ulna, and its proximal surface is flattened. The shaft is deeply constricted ventrally (caudally), resulting in a hooklike overhanging lip on the proximal end. Distally the radius is slightly convex but not nearly as strongly so as the ulna. There is no consensus on the structure of the manus in Carnotaurus. Bonaparte et al. (1990) tentatively identified a pair of irregularly shaped bones as carpals, but this interpretation is open to question. The manus has four metacarpals. The first three are short but bear strong distal articular surfaces. The element identified as metacarpal IV is the largest in the manus, and it tapers distally to a blunt point. Small, fragmentary pieces have also been identified as phalanges.
Whereas the neoceratosaur forelimb is short, coelophysoids retain plesiomorphically long forelimbs with a powerful, grasping hand (fig. 3.6D–G). The coelophysoid humerus shows sigmoid curvature, as well as torsion (fig. 3.6D–F). The deltopectoral crest is pronounced but does not exceed 45% of the humeral length (fig. 3.6D–G). Large samples of Coelophysis bauri and Syntarsus rhodesiensis provide the opportunity to examine proportional differences between forelimbs within populations of these taxa. There is dimorphism in the size and structure of the forelimb in both taxa (Colbert 1990; Raath 1990). The robust form has longer forelimbs, a larger deltopectoral crest, a broad humeral head, expanded and well-defined distal humeral epicondyles, and in Syntarsus a greatly enlarged olecranon process of the ulna (Raath 1977, 1990). The gracile form has a shorter forelimb with a less pronounced humeral head, deltopectoral crest, epicondyles, and olecranon process. The coelophysoid radius is shorter than the ulna and is bowed laterally (fig. 3.6J).
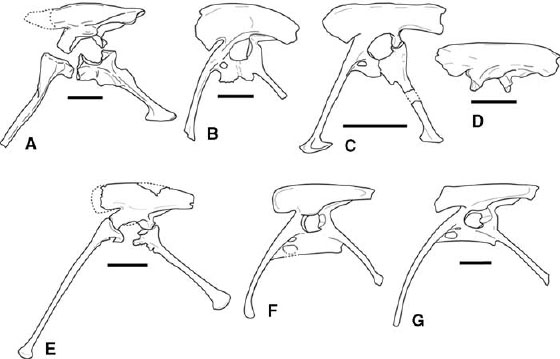
FIGURE 3.7. Pelves: A, Elaphrosaurus bambergi; B, Ceratosaurus nasicornis; C, Carnotaurus sastrei; D, Majungatholus atopus; E, Liliensternus liliensterni; F, Coelophysis bauri; G, Syntarsus rhodesiensis. Scale = 10 cm (A, E), 20 cm (B, D); 50 cm (C), 5 cm (F, G). (A after Janensch 1925; B after Gilmore 1920; C after Bonaparte et al. 1990; D after Sampson et al. 2001; E after Huene 1934a; F after Rowe and Gauthier 1990; G after Raath 1969, 1977.)
The coelophysoid manus measures less than two-thirds the combined lengths of the radius and the humerus (fig. 3.6G). As many as five ossified carpals have been reported in Coelophysis, and as many as six in Syntarsus rhodesiensis (fig. 3.6G, K; Raath 1969, 1977; Galton 1971a; Colbert 1989). Proximally digits I and II both articulate against an enlarged distal carpal 1 formed by fusion of the first and second distal carpals (fig. 3.6K). There is no ossified fifth digit. Metacarpal I is short and has asymmetrical distal condyles, facilitating some ability for digit I to oppose digits II and III during grasping (Galton 1971a). The proximal end is appressed against the side of metacarpal II, further strengthening the first digit. This contact is confined to the proximal third of metacarpal I, unlike in tetanurans, in which the contact may extend half the length of the first metacarpal. The second metacarpal is the largest in the hand, although the more slender metacarpal III is slightly longer in Dilophosaurus. Metacarpal III is not shifted to the palmar surface of II, unlike in tetanurans. There remains an ossified metacarpal IV, which is shorter than either metacarpals II or III and retains a nubbinlike phalanx. Given its small size and close association with metacarpal III, digit IV was possibly bound up in the palmar aponeurosis in life. As in tetanurans, the penultimate phalanges are elongate on all three functional digits. The unguals are laterally compressed and recurved and have large flexor tubercles. The manual phalangeal count is 2-3-4-1.
The ceratosaur pelvic girdle is distinctive in that the ilium, ischium, and pubis fuse to one another in adults, analogous to the condition later achieved in ornithurine birds and some other coelurosaurs. Fusion between pelvic elements is not complete in Carnotaurus, with co-ossification between the pubis and ilium and to only a partial degree between the ilium and ischium (Bonaparte et al. 1990). The ilium of Majungatholus shows no fusion to the other pelvic elements (fig. 3.7D; Sampson et al. 1998:fig. 3.2F). There is also no fusion between the pelvic elements of Elaphrosaurus (fig. 3.7A). However, the pelvic bones are thoroughly fused in Ceratosaurus nasicornis, Coelophysis, Syntarsus rhodesiensis, Syntarsus kayentakatae, and an unnamed taxon from the Kayenta Formation of Arizona (fig. 3.7B, F, G; Gilmore 1920; Raath 1969, 1977, 1990; Rowe 1989; Tykoski 1998).
The ceratosaur ilium is generally elongate, but with a low iliac blade (fig. 3.7). The postacetabular process is particularly long. The preacetabular process is ventrally expanded in Ceratosaurus, Carnotaurus, and Majungatholus (fig. 3.7B–D). The fossa for M. caudofemoralis brevis (= brevis fossa) is broad and deep, the ilium flaring laterally caudal to the acetabulum. Ceratosaurs lack a second fossa along the base of the preacetabular process for M. cuppedicus (Rowe 1986), a condition in many tetanurans. Syntarsus rhodesiensis and Coelophysis are derived in possession of a distinct, caudally rimmed fossa for M. iliofemoralis on the postacetabular process of the ilium (fig. 3.7F, G; Raath 1969, 1977; Rowe and Gauthier 1990; Hutchinson 2001a). The supracetabular crest is large, and particularly in coelophysoids it forms a dorsal hood overhanging the acetabulum. The pubic and ischial peduncles are roughly equal in size in coelophysoids, while the pubic peduncle is much larger than the ischial peduncle in neoceratosaurs (fig. 3.7A–D). The pubic and ischial peduncles in Ceratosaurus and Majungatholus bear stout projections that are inserted into corresponding sockets in the proximal pubis and ischium, respectively (fig. 3.7D; Sampson et al. 1998; Britt et al. 1999).
The proximal pubic plate is perforated by two fenestrae in several ceratosaur taxa (fig. 3.7B, F, G). A small obturator foramen opens ventrolaterally through the puboischial plate. There is a derived second opening ventromedial to the obturator foramen, the pubic fenestra. Owing to the exceptionally delicate nature of the puboischial plate, the borders of these openings are rarely preserved. The pubic shaft is cranially bowed to varying degrees in Ceratosaurus nasicornis, Masiakasaurus, Dilophosaurus, Liliensternus, Gojirasaurus, Coelophysis, and Syntarsus (fig. 3.7B, E–G; Gilmore 1920; Huene 1934a; Raath 1969; Colbert 1989; Carpenter 1997a; Tykoski 1998; Sampson et al. 2001). The pubis was restored without cranial bowing for Dilophosaurus (Welles 1984). The pubic shaft is shorter and broader in Segisaurus halli and Procompsognathus triassicus (Camp 1936; Ostrom 1981; Sereno and Wild 1992). This may be an illusion caused by breakage, as at least one individual of Syntarsus kayentakatae shows artificially broad pubic shafts caused by crushing of the pubic apron (Tykoski 1998). The distal end is tipped by a small, craniocaudal expansion or a knoblike swelling in coelophysoids but is not expanded into a “foot.”
The pubis is straight-shafted in Carnotaurus and in an Indian abelisaurid (Bonaparte et al. 1990; Chatterjee and Rudra 1996). The proximal pubis of Masiakasaurus possesses a socket to receive a peg from the ilium, much as in Majungatholus (Sampson et al. 2001). A large distal expansion (“foot”) was restored by Marsh (1892a) and Gilmore (1920) in illustrations of Ceratosaurus nasicornis, but as stated (Gilmore 1920:128), the restorations were based on Allosaurus. Recent finds show that the size and shape of the pubic foot in Ceratosaurus may change through ontogeny (Britt et al. 1999, 2000). The small pubic foot is expanded more caudally than cranially in Carnotaurus (fig. 3.7C). The distal foot is also caudally directed in Ceratosaurus and Masiakasaurus (Britt et al. 2000; Sampson et al. 2001:fig. 3.1g). The pubic foot of neoceratosaurs is smaller than that in some avetheropod tetanurans (e.g., Allosaurus), but it is as large as or larger than that seen in more basal tetanurans (e.g., Torvosaurus, Spinosauridae).
The ischium is plesiomorphic in possessing an obturator process that is not separated from the rest of the ischial plate (caudal half of the puboischial plate), unlike the derived condition in avetheropod tentanurans (Hutchinson 2001a). The ischium of Segisaurus is perforated by a large ischial foramen proximally, and the shaft is flattened and laterally expanded in a unique way (Camp 1936). However, it is possible that this flattening is the result of postmortem processes. Among neoceratosaurs the ischial shaft is straight and terminates in a craniocaudally expanded ischial foot in Elaphrosaurus and Carnotaurus (fig. 3.7A, C). In coelophysoids the ischial shaft is long, curves slightly ventrally, and terminates in a knoblike swelling (fig. 3.7E–G). In Dilophosaurus and Liliensternus the distal ischial expansion is much larger than the corresponding structure on the distal pubis (Huene 1934a; Welles 1984). Coelophysids and Liliensternus exhibit a prominent antitrochanter that straddles the ischium-ilium contact (fig. 3.7E–G). The ischial portion of the antitrochanter juts sharply into the acetabulum, forming a strong notch in the ischial border of the acetabulum.
The hindlimb bones are all hollow and thin-walled. The femoral neck and head angle craniomedially from the shaft, and the head is declined below the level of the greater trochanter, questionable in Carnotaurus (fig. 3.8). In Ceratosaurus the femoral head is large but still craniocaudally narrow (fig. 3.8A–C; Gilmore 1920; Madsen and Welles 2000). The femoral head of abelisaurids is more subspherical (Martínez et al. 1986; Bonaparte et al. 1990). The cranial trochanter is positioned low on the femur, staying below or just at the level of the femoral head (fig. 3.8A, B). It has been proposed that the cranial trochanter represents the insertion for M. puboischiofemoralis (Rowe 1986) or M. iliotrochantericus caudalis (Hutchinson 2001b). In Carnotaurus, Xenotarsosaurus, and some Ceratosaurus the cranial trochanter approaches the aliform condition seen in basal tetanurans (Gilmore 1920; Martínez et al. 1986; Bonaparte et al. 1990; Madsen and Welles 2000). A conspicuous trochanteric shelf (Andrews 1921) is known in Ceratosaurus, Elaphrosaurus, and Xenotarsosaurus, but little or no shelf is figured in Carnotaurus (fig. 3.8A, B; Gilmore 1920; Janensch 1925; Martínez et al. 1986; Bonaparte et al. 1990). The medial epicondyle (= craniomedial crest, entocondylar ridge) is enlarged and distinct in ceratosaurs (fig. 3.8), but it is exceptionally developed in abelisauroids such as Carnotaurus and Masiakasaurus (Bonaparte et al. 1990; Sampson et al. 2001). In Ceratosaurus and Xenotarsosaurus the crista tibiofibularis (also known as the ectocondylar tuber or tuberous process) is sharply set off from the lateral (= fibular) condyle on the caudal surface of the distal femur (fig. 3.8C; Gilmore 1920; Martínez et al. 1986; Madsen and Welles 2000).
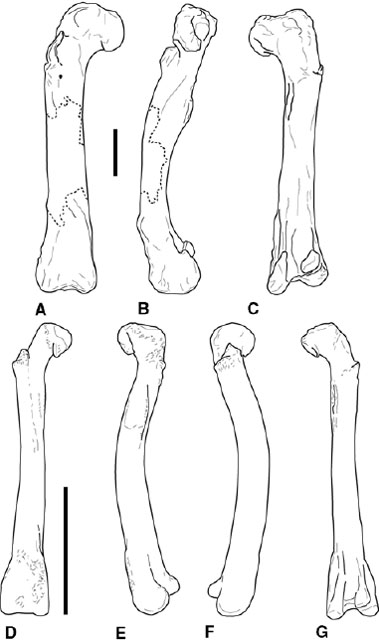
FIGURE 3.8. Femora. A–C, Ceratosaurus nasicornis: right femur in A, cranial, B, medial, and C, caudal views; D–G, Syntarsus kayentakatae: composite femur in D, cranial, E, lateral, F, medial, and G, caudal views. Scale = 10 cm.
The proximal femur in coelophysoids is small and wedge-shaped in proximal view, narrowing laterally into the greater trochanter (fig. 3.8D). Sample sizes are large enough for some coelophysoid taxa to determine a bimodal distribution in the development of proximal muscle scars and processes. The two morphologies are referred to as gracile and robust, respectively. In gracile forms the cranial trochanter is small and subconical or flangelike (Raath 1977, 1990; Welles 1984; Tykoski 1998). The trochanteric shelf is weakly developed, if discernable at all. In robust forms the cranial trochanter is a rugose, pyramidal mound that rises proximally from a strong, cranially jutting trochanteric shelf (fig. 3.8D, E). The trochanteric shelf differs in morphology and in its strong degree of development from the condition plesiomorphically found in Herrerasaurus and basal dinosauromorphs. The coelophysoid distal femur also bears an enlarged medial epicondyle (fig. 3.8D–F). Caudally, the crista tibiofibularis is distinctly set off from the lateral condyle in Dilophosaurus and perhaps Liliensternus (Huene 1934a; Welles 1984). In the coelophysids Coelophysis and Syntarsus the crest is further separated from the body of the lateral condyle by a distinct sulcus along the lateral margin of its base (fig. 3.8F; Rowe 1989). The popliteal fossa is crossed by a low infrapopliteal ridge between the lateral and medial condyles.
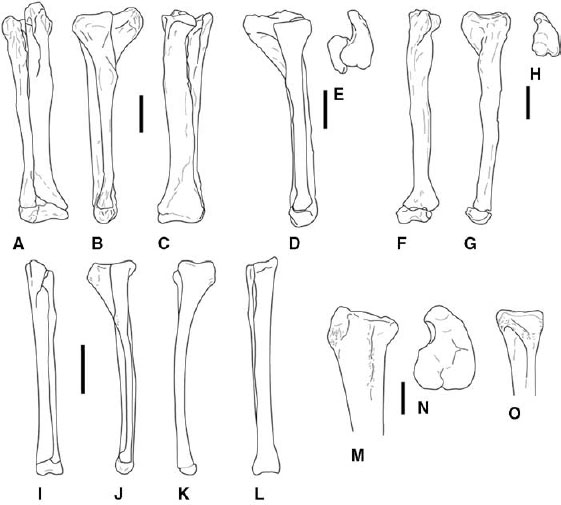
FIGURE 3.9. Tibiae and fibulae. A–C, Ceratosaurus nasicornis: right tibiotarsus and fibula in A, cranial, B, lateral, and C, caudal views. D, E, Xenotarsosaurus bonapartei: tibiotarsus in D, lateral, and E, proximal views. F–H, Elaphrosaurus bambergi: left tibia and astragalocalcaneum in F, cranial, G, lateral, and H, proximal views. I–L, Syntarsus rhodesiensis: left tibiotarsus and fibula in I, cranial, J, lateral, K, medial, and L, caudal views. M–O, Syntarsus kayentakatae: subadult left tibia in M, lateral, and N, proximal views; O, proximal left fibula in medial view. Scale = 10 cm (A–H), 5 cm (I–L), 2 cm (M–O). (A–C after Gilmore 1920; D, E, after Martínez et al. 1986 and Bonaparte et al. 1990; F–H after Janensch 1925; I–L after Raath 1977.)
The neoceratosaur tibia has an enlarged cnemial crest that rises proximally higher than the femoral condyles (fig. 3.9A, B, D, E). The craniodorsal end of the cnemial crest may be expanded to form a tuberosity. The tibia is shorter than the femur in Ceratosaurus and especially in abelisaurids (contra Bonaparte et al. 1990), but the tibia is longer than the femur in Elaphrosaurus (Gilmore 1920; Janensch 1925; Madsen and Welles 2000). There is a large crista fibularis on the lateral side of the tibia, and there is a deep dorsolaterally angled fossa on the cranial surface of the distal end for receipt of the ascending process of the astragalus. The medial surface of the proximal end of the fibula bears a strong sulcus in Ceratosaurus, much as in coelophysoids. The fibula of Ceratosaurus and abelisaurids bears a large cranial process for insertion of M. iliofibularis (fig. 3.9B). Distally the astragalus is firmly locked to the tibia, or it may even fuse, forming a tibiotarsus (Gilmore 1920; Martínez et al. 1986; Madsen and Welles 2000; see also below).
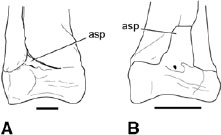
FIGURE 3.10. Astragalocalcaneum: A, Syntarsus kayentakatae, right distal tibiotarsus and fibula in cranial view; B, Masiakasaurus knopfleri, left distal tibiotarsus and fibula in cranial view. Scale = 1 cm. (B after Sampson et al. 2001.)
The tibia is shorter than the femur in Dilophosaurus and Liliensternus (Huene 1934a; Welles 1984) but longer in the smaller coelophysid taxa. The cnemial crest is not as large as in neoceratosaurs, and it does not project proximally beyond the level of the femoral condyles (fig. 3.9F, G). In Dilophosaurus, Syntarsus, and Coelophysis the tibia and fibula are closely appressed. Indeed, S. kayentakatae and Dilophosaurus have a longitudinal groove on the lateral surface of the tibia in which the fibula rests, but there is no evidence of fusion between the two. The proximal fibula in coelophysids and perhaps other mature coelophysoids is distinctive for the presence of a deep caudoventrally opening sulcus on the medial surface (fig. 3.9H; Rowe 1989; Rowe and Gauthier 1990). As mentioned, the same feature is also known in Ceratosaurus.
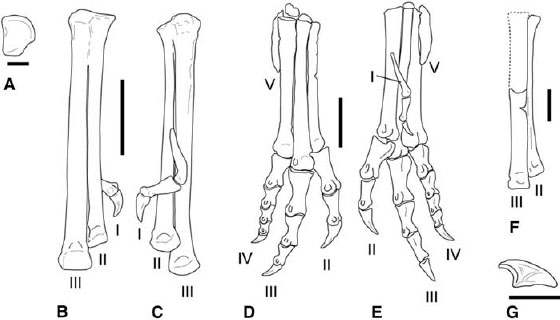
FIGURE 3.11. Pes. A–C, Syntarsus kayentakatae: A, right distal tarsal 4 in proximal view; B, C, right tarsometatarsus (missing unfused metatarsal IV) in B, cranial, and C, caudal views. D, E, Dilophosaurus wetherilli: articulated right pes in D, cranial, and E, caudal views. F, G, Masiakasaurus knopfleri: F, partial right pes (metatarsals II and III) in cranial view; G, pedal ungual in lateral view. Scale = 1 cm (A, F, G), 5 cm (B, C), 10 cm (D, E). (F, G, after Sampson et al. 2001.)
Ceratosaurs fuse the astragalus and calcaneum (fig. 3.10). This condition is not known in Dilophosaurus (Welles 1984), but based on a wide range of features throughout the skeleton all the known individuals of this taxon represent subadult individuals. In most adult ceratosaurs the two elements are so thoroughly co-ossified as to eliminate the line of suture to form an astragalocalcaneum, although the suture is still visible in Liliensternus (Huene 1934a). Furthermore, the astragalocalcaneum is fused to the distal of the tibia in most adult coelophysoids, Ceratosaurus, Masiakasaurus, and Xenotarsosaurus, creating a tibiotarsus (Gilmore 1920; Raath 1969, 1977; Martínez et al. 1986; Colbert 1989, 1990; Rowe 1989; Rowe and Gauthier 1990; Long and Murry 1995:fig. 3.192F–J; Hunt et al. 1998; Tykoski 1998; Sampson et al. 2001). Fusion is strongest caudally, where the suture is first obliterated. There is also fusion between the distal fibula and the calcaneum in Syntarsus kayentakatae, in Xenotarsosaurus, and in an isolated distal tibiotarsus of Camposaurus arizonensis (Hunt et al. 1998). The cranial face of the astragalus is traversed by a shallow groove in neoceratosaurs, a condition shared with tetanurans. A less pronounced groove is described on the astragalus of Dilophosaurus (Welles 1984), and one is also discernible in Syntarsus kayentakatae (fig. 3.10A). As seen in Dilophosaurus wetherilli, the ascending process of the astragalus may be a separate ossification center (Welles 1983, 1984). The ascending process of the astragalus is a low triangular wedge nestled within a fossa on the distal end of the tibia in Elaphrosaurus, Ceratosaurus, and coelophysoids (figs. 3.9A, 3.10A). In coelophysoids the process can be obscured cranially by an overlapping flange of the distal end of the fibula, which may have led some to believe it was absent (e.g., Colbert 1964b, 1989). The condition in abelisauroids more closely resembles that in tetanurans. In Masiakasaurus the ascending process is a tall rectangular plate (fig. 3.10B; Sampson et al. 2001), but in other abelisauroids it is a triangular plate (Martínez et al. 1986).
The pes is virtually undescribed for abelisaurids, although partial pedes of Elaphrosaurus and Ceratosaurus are known (Gilmore 1920; Martínez et al. 1986; Bonaparte et al. 1990; Madsen and Welles 2000). No distal tarsals are known for Elaphrosaurus, but a fourth distal tarsal was described and figured for Ceratosaurus (Madsen and Welles 2000). It is virtually identical in overall form to that in coelophysoids. Rather than a simple disc, the fourth distal tarsal is rectangular with a large notch in its caudolateral corner, giving the impression that the tarsal has a tuberous caudal process (fig. 3.11A). Contrary to some reports (Colbert 1989; Rowe 1989; Rowe and Gauthier 1990), an ossified distal tarsal 2 is not known in Coelophysis bauri or Syntarsus kayentakatae. Raath (1969) identified such an element in S. rhodesiensis but later (1977) decided that it was really distal tarsal 3. Distal tarsal 3 is large, caps its respective metatarsal, and fuses to it in subadult stage. Distal tarsal 4 evidently does not co-ossify with the metatarsals at any point in ontogeny.
Ceratosaurus nasicornis exhibits fusion between metatarsals II, III, and IV. The rough, pitted condition opens the possibility that fusion between these elements is pathological. Indeed, a large, isolated metatarsal IV assigned to Ceratosaurus shows no sign of fusion with other pedal elements (Madsen and Welles 2000). In adult coelophysids distal tarsal 3 and metatarsals II and III fuse to create a tarsometatarsus (fig. 3.11B). Articulated coelophysoid metatarsi are narrow and long (fig. 3.11B, D). The first metatarsal is restricted to the distal half of metatarsal II, affixed to its caudal surface (fig. 3.11C, E). Metatarsal V is reduced to a splint and lacks phalanges. The proximal end lies within the notch in the caudolateral corner of distal tarsal 4. The metatarsi of noasaurids are apomorphic in the great reduction in thickness of the proximal two-thirds of metatarsal II (Bonaparte and Powell 1980; Bonaparte 1991b; Sampson et al. 2001). In Velocisaurus unicus metatarsal IV is also reduced in size (Bonaparte 1991b). The pedal unguals of abelisauroids are distinctive in having a triangular set of lateral grooves, in contrast to the single groove found on other theropods (fig. 3.11G; Coria and Salgado 1998b; Sampson et al. 2001).
Systematics and Evolution
Marsh (1884a) originally coined the name Ceratosauria to encompass Ceratosauridae (at the time composed of only Ceratosaurus nasicornis) and Ornithomimidae (Marsh 1884a). Subsequent authors recognized either Ceratosauridae or Ceratosauria, disagreeing on the taxonomic content of each (Gilmore 1920; Huene 1932, 1948, 1956; Welles 1984). Others rejected both terms (e.g., Lydekker 1888a; Cope 1892b; Abel 1919; Romer 1933, 1956, 1966; Lapparent and Lavocat 1955; Colbert 1964a; Steel 1970; Charig 1979). Gauthier (1986) resurrected the name Ceratosauria to designate the sister taxon to the stem group Tetanurae. However, the clade still lacked a stable phylogenetic definition as the name Ceratosauria was defined only by enumeration of its taxonomic content.
The first stable, stem-based definition for Ceratosauria was provided by Rowe (1989:132): “Tetanurae includes those theropods more closely related to birds (and includes birds), whereas Ceratosauria includes taxa more closely related to Ceratosaurus nasicornis.” Birds (= Aves of Gauthier 1986; Neornithes of Sereno 1998; Padian et al. 1999) and the eponymous Ceratosaurus nasicornis are thus the reference taxa on which the name is established, and Ceratosauria so designated is a stem lineage (sensu de Queiroz and Gauthier 1990, 1992). Rowe and Gauthier (1990) provided a node-based definition for the name, and Sereno (1998) redefined the name to refer to all neotheropods closer to Coelophysis bauri than to Aves (i.e., Neornithes). however, Rowe (1989) has priority. Also, the name Ceratosauria was originally brought into the phylogenetic system to represent the sister taxon to the stem clade Tetanurae (Gauthier 1986), so we recognize the stem definition of the clade here.
In addition to the diagnostic features of Ceratosauria listed in the “Definition and Diagnosis” section above, the phylogenetic placement of several other potentially informative apomorphies is ambiguous at this nexus, as they depend on the character-optimization criterion used, for example, ACCTRAN or DELTRAN (Swofford 1998). These apomorphies include the absence of the subnarial foramen on the premaxilla-maxilla suture; the craniodorsal border of the axial neural spine being convex in the lateral view; the postaxial cervical pleurocoels being deep ovoid pockets or well-defined fossae excavated into the lateral surfaces of the centra; the cervical ribs being fused to vertebrae in adults; the cervical ribs being two to three times as long as the centrum; fused sacral vertebrae; elements of the sacral neural arch (transverse processes, arches, neural spines) and sacral ribs being fused to one another; a sharp ventral groove on at least the proximal caudal centra; a broad scapular blade; the dorsal margin of the ilium being linear, dipping caudoventrally above the ischial peduncle; an iliac postacetabular process with a concave caudal margin; the cranial trochanter being a conical or pyramidal prominence; an infrapopliteal ridge between the fibular and medial condyles of distal femur; the proximal fibula being excavated medially by a deep caudoventrally opening sulcus; and the ascending process of the astragalus being low, triangular, and wedgelike.
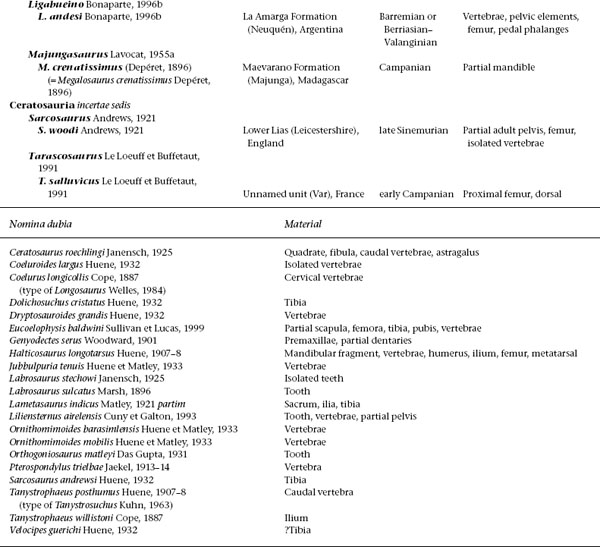
Many ceratosaurs have long histories of problematic taxonomy and assignment. Noncladistic classifications placed most in taxonomic wastebaskets (e.g., Podokesauridae, Halticosauridae, Procompsognathidae, Megalosauridae [Romer 1956, 1966; Welles 1954; Carroll 1988]). These groups became repositories for nearly any primitive or incomplete theropod from Upper Triassic or Jurassic sediments. To prevent Ceratosauria from becoming a modern wastebasket group for nontetanuran theropods, we recognize only those taxa that possess unequivocal ceratosaur apomorphies or are diagnosable at some less inclusive level within the lineage. We also recognize only those higher taxonomic names that have node- or stem-based definitions.
Within Ceratosauria are two main sister lineages, Coelophysoidea and Neoceratosauria (Novas 1991, 1992b; Holtz 1994). Coelophysoidea comprises all theropods closer to Coelophysis than to Ceratosaurus (Holtz 1994). The only well-supported higher-level clade named within the coelophysoid lineage is Coelophysidae, comprising Coelophysis, Syntarsus, and all descendants of their most recent common ancestor (Holtz 1994). Sereno (1998) defined Coelophysidae in reference to Coelophysis, Procompsognathus triassicus, and all descendants of their most recent common ancestor and coined the names Coelophysinae and Procompsognathinae for all coelophysids (sensu Sereno 1998) closer to Coelophysis or Procompsognathus than to the other, respectively. However, insofar as our analysis finds the phylogenetic position of Procompsognathus triassicus to be unsteady under certain conditions (see below), we do not employ Coelophysidae, Coelophysinae, or Procompsognathinae sensu Sereno (1998).
Neoceratosauria has received considerable attention over the past decade as new material and taxa have been described. Neoceratosauria comprises all theropods closer to Ceratosaurus than to Coelophysis (Holtz 1994, 1998a). Within it are several named subclades. Abelisauroidea is defined as comprising all theropods closer to Carnotaurus sastrei than to Ceratosaurus nasicornis (Holtz 1994, 1998a; Padian et al. 1999). The name Abelisauria has been used by some for the node-based clade comprising Noasaurus, Carnotaurus, and all descendants of their most recent common ancestor. Abelisauridae has been defined as comprising Abelisaurus comahuensis and Carnotaurus sastrei and all descendants of their most recent common ancestor (Holtz 1998a; Sereno 1998; Padian et al. 1999). Abelisaurids have been further divided into Carnotaurinae (i.e., all abelisaurids closer to Carnotaurus than to Abelisaurus) and Abelisaurinae (i.e., all abelisaurids closer to Abelisaurus than to Carnotaurus) (Sereno 1998). Bonaparte and Powell (1980) erected the name Noasauridae solely for Noasaurus leali. A handful of fragmentary taxa show some affinities with Noasaurus, and together they may form a diagnosable clade (Huene 1932; Huene and Matley 1933; Bonaparte 1991b, 1996b; Sampson et al. 2001).
What follows is the result of a numerical cladistic analysis of 19 ingroup taxa, including all ceratosaurs and three basal tetanurans (Torvosaurus tanneri, Spinosauridae, Allosaurus fragilis). The taxa were scored for 175 characters assembled from several previously published or in-press works, as well as a number of new characters. Successively closer outgroup taxa were Ornithischia, Prosauropoda, and Herrerasaurus. All characters were treated as unordered characters. The analysis was conducted using PAUP*4.0 (Swofford 1998) and resulted in five most parsimonious trees, all with a length of 386, a consistency index (CI) of 0.588, and a retention index (RI) of 0.739. Character distribution was evaluated under both ACCTRAN and DELTRAN optimization criteria. Figure 3.12A represents an Adams consensus tree of the five hypotheses generated by our analysis. The five trees differ only in the position of Segisaurus with respect to Coelophysis and Syntarsus.
As discussed earlier, our analysis supports previous hypotheses of ceratosaurian monophyly, uniting coelophysoids and neoceratosaurs as sister lineages within Ceratosauria (fig. 3.12A; Gauthier 1986; Rowe1989; Rowe and Gauthier 1990; Holtz 1994, 1998a; Sereno 1999a). Other analyses have resulted in hypotheses of ceratosaur relationship that remove coelophysoids from the Ceratosaurus + Abelisauridae + Tetanurae clade (fig. 3.12B, C; Rauhut 1998; Forster 1999; Carrano and Sampson 1999; Sampson et al. 2001; Carrano et al. 2002). These alternative hypotheses do not agree on the position of Ceratosaurus and abelisaurids with respect to Tetanurae and each other. Some hypotheses shift Coelophysoidea outside the neotheropod clade (= Ceratosauria + Tetanurae) but still find a monophyletic Ceratosauria with abelisauroids the sister taxon to Ceratosaurus (fig. 3.12B; Sampson et al. 2001; Carrano et al. 2002). Others find coelophysoids, Abelisauroidea, and Ceratosaurus to be successively closer outgroups to Tetanurae (fig. 3.12C; Rauhut 1998; Carrano and Sampson 1999). Fewer hypotheses find abelisaurids to be tetanurans, derived relative to Torvosaurus and other basal tetanuran taxa (Forster 1999). The temporal gap between coelophysoids and neoceratosaurs adds to this debate. Coelophysoids are not known from deposits younger than the Early Jurassic, and definitive neoceratosaurs are unknown from sediments older than the Late Jurassic. The temporal gap between the two lineages can be viewed in two ways. If hypotheses supporting the position of coelophysoids within Ceratosauria are correct, the temporal gap is evidence of substantial missing data in our record of ceratosaur diversity (i.e., ghost lineages). However, the gap could instead be evidence that coelophysoids lie outside the ceratosaurian clade and represent a radiation of theropods prior to the neotheropod split into Ceratosauria and Tetanurae (Rauhut 1998, 2000a; Carrano and Sampson 1999; Forster 1999).
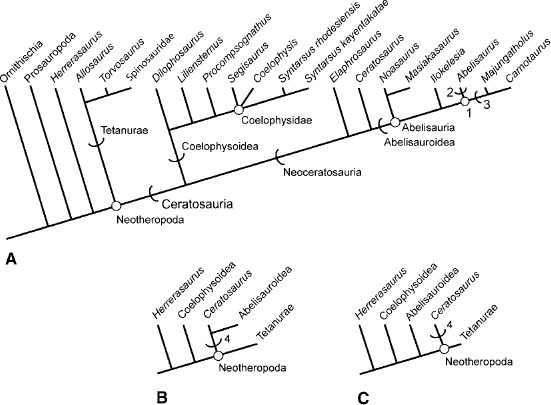
FIGURE 3.12. Phylogenetic hypotheses of ceratosaurian relationships: A, Adams consensus of five equally most parsimonious trees generated by our analysis; tree length = 386, CI = 0.588, RI = 0.739. Stem-defined clades indicated by arcs. Node-defined clades indicated by open circles at nodes. B, alternative phylogenetic hypothesis supported by Sampson et al. (2001). C, alternative hypothesis of ceratosaur relationships from Rauhut (1998) and Carrano and Sampson (1999). 1 = Abelisauridae; 2 = Abelisaurinae; 3 = Carnotaurinae; 4 = Ceratosauria.
The neoceratosaur lineage is diagnosed by the following unambiguous apomorphies: a sacrum composed of six vertebrae by addition of either a second dorsosacral or a third caudosacral; a midsacral centra strongly reduced in size; the ventral margin of the sacrum arching dorsally; the distal end of the ischium expanded into an ischial foot; and the cnemial crest of the tibia craniocaudal as long as or longer than the articular condyles, the crest hooking sharply laterally and rising well above the proximal condyles.
A large number of ambiguous apomorphies may also diagnose the neoceratosaur clade. Many are located in the skull and jaws, the atlas, and the axis, all of which are missing in Elaphrosaurus, the most basal neoceratosaur in this analysis. Therefore, these cranial apomorphies are mapped as present for Neoceratosauria under ACCTRAN optimization, but most are diagnostic of a Ceratosaurus + Abelisauroidea clade under DELTRAN optimization. The ambiguous apomorphies for the neoceratosaur lineage are as follows: the skull length less than three times the height of the caudal skull height; the premaxilla dorsoventrally taller than it is rostrocaudally long below the external naris; the maxillary process of the premaxilla short, allowing the maxilla to contribute significantly to the external naris; the nasals fused; the rostral end of the nasal laterally convex, overhanging the caudal portion of the external naris; the nasal contributing to the antorbital cavity; the lacrimal antorbital recess having a single opening; the frontals fused; the frontals and parietals fused; the infratemporal fenestra about twice the size of the orbit in lateral view; the quadrate and quadratojugal fused; the quadrate foramen absent; the height of the quadrate dorsal ramus greater than that of the orbit; a pronounced, sharply defined median ridge on the supraoccipital; the interdental plates fused to one another; a pleurocoel in the cranial end of the axis; the pneumatic foramen or foramina in the axis neural arch caudodorsal to the diapophysis; the sacral centra fused to an extreme degree, sutures being difficult to discern and only swellings marking the articular surfaces of the centrum; the cranial to mid-dorsal centra having two pairs of pleurocoels in the centrum; the cranial margin of the scapulocoracoid smoothly curved and uninterrupted by a notch at the scapulocoracoid contact; the proximal head of the humerus bulbous; the distal humeral condyles flattened; the deltopectoral crest greater than 45% the length of the humerus; the deltopectoral crest oriented obliquely on the humeral shaft; the proximal end of metacarpal I loosely appressed to metacarpal II; the iliac postacetabular process having a concave caudal margin; a peg-in-socket ilium-pubis articulation; the ischial peduncle of the ilium oriented ventrally; the distal end of the pubis terminating in a caudally expanded foot; the craniocaudal length of the pubic foot 10%–30% of the length of the pubic shaft; the ischial antitrochanter large or markedly developed; a shallow extensor groove on the cranial surface of the distal femur; the tibia expanded caudal to the fibula; pedal digits with two lateral grooves.
The position of Elaphrosaurus bambergi varies among different analyses. This Late Jurassic taxon from Africa was once considered a basal ornithomimid, based in large part on its low cervical vertebrae and straight-shafted humerus (Galton 1982c; Barsbold and Osmólska 1990). Cladistic studies show Elaphrosaurus to be a member of the neoceratosaur lineage, although its position varies from basal Abelisauroidea (Holtz 1994) to basal Neoceratosauria (this study). Thus far, the skull, atlas, and axis, some cervical and dorsal vertebrae, and most of the forearm and manus of Elaphrosaurus are unknown (Janensch 1925). Nevertheless, it possesses a straight-shafted humerus with a rounded proximal head, six vertebrae in the sacral series, and an ischial foot, characters shared by other neoceratosaurs. However, its cervical vertebrae strongly resemble those of derived coelophysoids, possessing broad fossalike lateral pleurocoels and elongate midcervical centra.
Ceratosaurus nasicornis has received considerable attention in recent years as more material of this Late Jurassic taxon has been found, including a juvenile (Britt et al. 1999). Two new Ceratosaurus taxa were proposed recently: C. magnicornis and C. dentisulcatus (Madsen and Welles 2000). Both of the purported new taxa are substantially larger than C. nasicornis, an individual generally assumed to be of adult ontogenetic stage (Gilmore 1920; Rowe and Gauthier 1990; Madsen and Welles 2000). However, this assumption has now been questioned (Britt et al. 2000); it is possible that material assigned to C. magnicornis and C. dentisulcatus represents later ontogentic stages of C. nasicornis. However, fusion of the sacrum, pelves, and neurocentral sutures indicates an adult stage for C. nasicornis. In addition to their larger size, both new Ceratosaurus forms possess premaxillary and mesial dentary teeth that bear longitudinal ridges on their lingual surface (Madsen and Welles 2000).
Ceratosaurus nasicornis and both of the purported additional Ceratosaurus taxa possess a large, median nasal horn, as well as lacrimal hornlets (fig. 3.1A, B). The median horn is formed by fusion of the left and right nasals, and all three of these cranial ornamentations increase in size through ontogeny (Britt et al. 2000). A large pneumatic recess marks the rostrodorsal corner of the maxillary antorbital fossa (Gilmore 1920; Madsen and Welles 2000). Ceratosaurus is also distinctive in possessing a row of median osteoderms, the only such osseous ornamentation reported for any theropod (Gilmore 1920; Madsen and Welles 2000).
Additional material attributed to Ceratosaurus includes a fragmentary specimen from the Morrison Formation of Oklahoma (Stovall 1938). However, the material possesses no characters diagnosable beyond the level of Theropoda. Janensch (1925) named Ceratosaurus roechlingi for material collected from the Tendagaru deposits of Tanzania. A referred femur has a trochanteric shelf and a tibiofibular crest sulcus, but the structures are not explicitly described. Although this form does not possess features diagnostic of Ceratosaurus, a large neoceratosaur is known for the Late Jurassic of Tanzania.
Our understanding of Abelisauridae has increased greatly in the past decade. New finds from South America, Madagascar, and India show that the clade was widespread on the Gondwanan landmasses during the Cretaceous. Abelisaurids are notable for their short, deep skulls, extensive sculpturing on the cranial bones, contact between the lacrimal and postorbital that forms pronounced brows over the orbit, constriction of the orbital fenestra by processes of the lacrimal and postorbital, exceptionally large epipophyses on the cervical vertebrae, short hindlimb proportions, and in at least Carnotaurus extremely reduced forelimbs. The preorbital region of the skull is taller than long in Carnotaurus but longer than tall in other abelisaurids (Bonaparte and Novas 1985; Bonaparte et al. 1990; Chatterjee and Rudra 1996; Sampson et al. 1998).
Our analysis found both Carnotaurus and Majungatholus to be more closely related to each other than to Abelisaurus, although given the lack of postcranial material for the latter, this hypothesis of relationship may change with future discoveries. Both Carnotaurus and Majungatholus have dorsal cranial ornamentation in the form of hypertrophied supraorbital horns in the former and a median frontal dome in the latter (Bonaparte et al. 1990; Sampson et al. 1998). Smaller supraorbital eminencies are known in Aucasaurus (Coria et al. 2002b). The skull of Abelisaurus (Bonaparte and Novas 1985). The skull lacks the extravagant cranial ornamentation seen in carnotaurine abelisaurs. Ilokelesia aguadagrandensis is incompletely known but may be a basal abelisaurid or the closest outgroup to Abelisauridae (Coria and Salgado 1998a).
Indosuchus raptorius and Indosaurus matleyi were both based on isolated cranial remains, now lost (Chatterjee 1978b) from the Lameta Formation (Maastrichtian) of India (Huene and Matley 1933). These taxa were considered allosaurids or even tyrannosaurids (Huene and Matley 1933; Walker 1964; Chatterjee 1978b), but closer examination revealed abelisaurid features in each (Bonaparte et al. 1990; Molnar 1990; Chatterjee and Rudra 1996). A number of cranial remains and a complete postcranial skeleton of a large abelisaurid from the Lameta Formation were assigned to Indosuchus raptorius (Chatterjee 1978b; Chatterjee and Rudra 1996). However, these specimens do not share any preserved braincase material with the latter and therefore cannot be assigned to that taxon with certainty.
Xenotarsosaurus bonapartei was named for a nearly complete right hindlimb and a dorsal vertebra from the Bajo Barreal Formation of Chubut Province, Argentina (Martínez et al. 1986). The remains possess a number of ceratosaurian synapomorphies (a deep femoral ligament sulcus, an infrapopliteal ridge, fused tibia, and an astragalocalcaneum), as well as neoceratosaur features, such as a hypertrophied epicondylar crest on the distal femur and a craniodorsally enlarged cnemial crest. An isolated maxilla from near where Xenotarsosaurus was discovered shows other abelisaurid traits, such as a nearly vertical dorsal (nasal) process, external sculpturing, a reduced maxillary antorbital fossa, and heavily striated interdental plates (Lamanna et al. 2002).
It has been suggested that there is a close relationship between abelisaurids and Carcharodontosauridae, a clade of gigantic theropods from the Early Cretaceous of northern Africa and South America (Novas 1997b). Abelisaurids share several cranial features with carcharodontosaurids, including sculpturing of some cranial bones, fusion between the nasals, a highly reduced maxillary antorbital fossa, supraorbital contact between the lacrimal and postorbital, and constriction of the orbital fenestra by processes of the lacrimal and postorbital (Novas 1997b; Lamanna et al. 2002). However, cladistic analyses show that carcharodontosaurids are members of Carnosauria (sensu Padian et al. 1999) and not closely related to Abelisauridae (Holtz 1998a; Sereno 1998).
Noasaurid abelisauroids are little known, the exception being Masiakasaurus knopfleri from the Maevarano Formation (Campanian) of Madagascar (Sampson et al. 2001). These small abelisauroids may be united by the presence of ten or fewer maxillary teeth, a sharply marked and caudoventrally dipping ventral border of the maxillary antorbital fossa, cervical neural spines positioned over the cranial half of the centrum, a dorsal vertebral centrum length greater than twice the height of the cranial surface, and a metatarsal II with a reduced midshaft diameter. In addition to Noasaurus and Masiakasaurus, Velocisaurus unicus from the Late Cretaceous of Argentina and Laevisuchus indicus from the Lameta Formation of India (Bonaparte 1991b; Huene and Matley 1933) also exhibit one or more of these characters. Like abelisaurids, noasaurids were a more diverse and more geographically widespread clade than previously thought.
Genusaurus sisteronis is based on vertebral, pelvic, and hind-limb elements of a small theropod from Lower Cretaceous (Albian) sediments of France (Accarie et al. 1995). Some of the features are an elongate dorsal centrum, fusion of the pelvic elements, a low trochanteric shelf and cranial trochanter, a hypertrophied medial epicondyle on the distal femur, and a pronounced M. iliofibularis tubercle on the fibula. The position of Genusaurus is highly variable in the cladistic analysis, so we treat it as Neoceratosauria incertae sedis. Additional theropod fossils from southern France were the basis for Tarascosaurus salluvicus (Le Loeuff and Buffetaut 1991). The incomplete dorsal vertebra known from this species has numerous pneumatic foramina in the neural arch reminiscent of those in abelisaurids. Few definitive data can be had from the poorly preserved proximal femur of this taxon. We tentatively consider Tarascosaurus as Abelisauroidea incertae sedis.
The phylogeny of coelophysoids has remained stable since the first cladistic studies of the group (fig. 3.12A; Gauthier 1986; Rowe 1989; Rowe and Gauthier 1990). The coelophysoid lineage is diagnosed here by 21 unambiguous apomorphies: the craniocaudal length of the internal antorbital fenestra greater than 25% of the maximum skull length; the premaxilla nasal process forming half or less of the rostrodorsal narial border; the maxillary process of the premaxilla loosely overlapping the premaxillary process of the maxilla, resulting in flexible articulation; a subnarial diastema in the tooth row at the premaxilla-maxilla contact; the maxilla alveolar border sharply upcurved rostrally to the degree that the first maxillary tooth is oriented rostroventrally; the dentary rostral end dorsally raised over the length of the first three to four alveoli; serrations reduced or absent on premaxillary teeth; the mesial premaxillary teeth subcircular to circular in cross section and straight or only slightly recurved (nearly conical); the premaxillary tooth row not extending caudally below the external naris; most mesial dentary teeth nearly straight, with subcircular cross section and reduced or absent serrations; the lacrimal rostral ramus longer than the ventral ramus; the axial diapophysis absent; the axial parapophysis reduced; the largest sacral rib articulating with the first caudosacral; the distal scapular blade expanding relative to the base of the blade; the humerus sigmoid in lateral view; the iliac-pubic articulation smaller than the iliac-ischial articulation; the distal end of the iliac-pubic peduncle having distinct cranial and ventral articular faces separated by a sharp angle; the distal pubis terminating with a small expansion, or “knob”; the femoral head ligament (ligamentum capitus femoralis) sulcus on the caudal surface of the proximal femur deep, giving the femur a caudally hooked profile in proximal view; and the medial flange on the distal fibula partly overlapping the ascending process of the astragalus.
Equivocal coelophysoid apomorphies include a skull length greater than three times the caudal skull height; the subnarial foramen absent; nasals bearing low ridges along lateral margins; a single opening for the trigeminal nerve; the mesial premaxillary teeth circular to subcircular in cross section and straight or only slightly recurved; postaxial the cervical and cranial dorsal pleurocoels deep ovoid pockets or well-defined fossae excavated into the lateral surfaces of centra; the length of the midcervical centra about twice the diameter of the cranial face; the cervical ribs long and extremely thin, four or more times as long as the centrum; a sharp ventral groove on at least the proximal caudal centra; pubic shafts separated by a short rectangular notch in the pubic apron at the distal extremity; the distal ischial shaft terminating in a small knob; the proximal end of the femur transversely elongate and wedge-shaped in proximal view; the distal end of the tibia not expanded caudal to the fibula; distal tarsal 3 fused to metatarsal III.
Dilophosaurus wetherilli is a large coelophysoid known only from the Kayenta Formation (Sinemurian–Pliensbachian) of northeastern Arizona (Welles 1954, 1970, 1984). The sister taxon to the other coelophysoids, it is recognized by its highly arched, paired nasolacrimal crests, the cruciform distal extremities of its cervical neural spines, and the squared-off distal expansion of its scapular blade (Welles 1984). All individuals known so far are juveniles and subadults showing gracile features (Welles 1984). Welles (1984) considered an incomplete robust individual found near the locality where D. wetherilli was originally found to be a separate taxon, but it is now recognized as a larger, more ontogenetically developed individual. Hu (1993) assigned a second species to Dilophosaurus: D. sinensis, from the Lower Lufeng Series (Lower Jurassic) of China. Although the skull of this taxon sports large nasolacrimal crests superficially like those reconstructed in D. wetherilli (Welles 1984), features elsewhere in the skeleton ally it to tetanuran theropods. The taxonomic assignment of the Chinese species remains in question (Lamanna et al. 1998).
Another large coelophysoid, Liliensternusliliensterni, was found to be more derived than Dilophosaurus in this and other analyses (Rowe 1989; Rowe and Gauthier 1990). It is known from two incomplete subadult skeletons from the Knollenmergel (Norian) of Germany. Apart from those characters grouping them with other coelophysoids, and in particular the coelophysids, the two individuals do not preserve any shared apomorphies that might be considered diagnostic of Liliensternus. Until such data are found Liliensternus should be viewed as a metataxon. Huene (1934a) described the two skeletons under the name Halticosaurus liliensterni, but Welles (1984) erected the name Liliensternus to emphasize their distinctness from Halticosaurus longotarsus (Huene 1907–8) and Halticosaurus orbitoangulatus (Huene 1932). This decision is justified in that the material referred to Liliensternus preserves data permitting its placement within Coelophysoidea, whereas the Halticosaurus material possesses no ceratosaurian features. Cuny and Galton (1993) erected a second species of Liliensternus, L. airelensis, based on fragmentary remains from northwestern France. The material possesses one unambiguous ceratosaurian character (two pairs of cervical pleurocoels), and the pleurocoels are well-defined fossae on the lateral surface of the centra, a coelophysoid feature. As Liliensternus has no diagnostic apomorphies, the name cannot be applied to the French material, and we consider it referable only to Coelophysoidea indet.
Procompsognathus triassicus is based on a small, incomplete skeleton from the Stubensandstein (Norian) of Germany. It preserves the dorsal through the most proximal caudal vertebrae, the humerus, partial pelves, and hindlimbs (Fraas 1913, 1914; Huene 1921a; Ostrom 1981; Sereno and Wild 1992). A small skull once referred to P. triassicus is that of a small crocodylomorph (Sereno and Wild 1992). P. triassicus itself shares features with Segisaurus halli and Coelophysidae, including dorsal vertebral centra twice as long as tall and a markedly protruding cranial trochanter. The proximal tarsals are fused to one another and to the tibia, suggesting that this small individual may have been an adult.
Segisaurus halli is based on another small, incomplete skeleton lacking the skull and most of the dorsal and cervical vertebral series (Camp 1936) from the Navajo Sandstone (Pliensbachian–Toarcian) of northeastern Arizona; thus it is the geologically youngest known, unequivocal coelophysoid. An element first identified as one of a pair of clavicles (Camp 1936) is actually a furcula (Senter and Hutchinson 2001), a feature shared with Syntarsus and possibly Coelophysis (Tykoski 1998; Downs 2000; Tykoski et al. 2002). A structure unique to Segisaurus is a large opening in the ischial portion of the puboischial plate, named the ischial fenestra (Camp 1936). New preparation and restudy of S. halli promise to reveal much more about this taxon (Senter and Hutchinson 2001).
As described elsewhere, Coelophysidae comprises Coelophysis and Syntarsus and all descendants of their most recent common ancestor. Our analysis found that Segisaurus may belong to this clade, although given the incomplete nature of known material, its phylogenetic position is likely to remain uncertain. There are large amounts of missing data for Segisaurus and the successive outgroups to Coelophysidae (e.g., Procompsognathus, Liliensternus). However, in those phylogenetic hypotheses that found Segisaurus within Coelophysidae, the clade is unambiguously diagnosed by fusion of the pelvic bones and the presence of the pubic fenestra ventromedial to the obturator foramen. Regardless of the position of Segisaurus, it is clear that Coelophysis and Syntarsus are closely related. However, calls for synonymy between Coelophysis and Syntarsus (Paul 1988a, 1993) are academic arguments rooted in rank-based (i.e., noncladistic) classification and are not reflective of phylogeny.
Coelophysis bauri is the best-represented Triassic (Carnian–Norian) theropod (Colbert 1989, 1990), although it has been the center of a heated debate over the diagnosability of its original material and its relationship to theropod material from the prolific Ghost Ranch Quarry locality (Padian 1986; Hunt and Lucas 1991a). Hunt and Lucas (1991a) urged restricting the name to the original material and proposed a new name for the Ghost Ranch theropods, Rioarribasaurus colberti. However, the International Commission on Zoological Nomenclature was petitioned to erect a neotype (Colbert et al. 1992), which it did (ICZN 1996). All the coelophysids from the Ghost Ranch locality are now considered Coelophysis bauri. Surprisingly, this well-known theropod has not been diagnosed by unique apomorphies and is currently a metataxon that possesses only those characters shared within Coelophysidae.
Fragmentary remains of a small theropod were recently collected from the Chinle Formation, near the Ghost Ranch locality, and were proposed to be a new taxon distinct from Coelophysis (Sullivan and Lucas 1999). The material represents a juvenile or subadult and cannot be unequivocally diagnosed from other coelophysoids. Two specimens from the Connecticut Valley, in the eastern United States, probably from the Port-land Formation (Lower Jurassic), were assigned to Coelophysis. One was composed solely of the lost specimen of Podokesaurus holyokensis (Talbot 1911). Colbert (1964b) referred this specimen to Coelophysis (C. holyokensis) on the basis of what can now be seen as shared plesiomorphic features. While Podokesaurus may possess coelophysoid characters (e.g., the small knoblike distal expansion of the pubis), it preserves no derived characters uniting it with Coelophysis. We agree with others (Olsen 1980c; Padian 1986; Rowe and Gauthier 1990) that the name Podokesaurus holyokensis be restricted to the lost specimen. The other Connecticut Valley specimen is a natural cast of a partial tibia, pubis, and rib (Colbert and Baird 1958). This specimen may be a coelophysoid, but it possesses no characters diagnosable beyond the level of Theropoda.
Syntarsus includes two taxa, S. rhodesiensis from the Forest Sandstone and Upper Elliot formations (Hettangian–Sinemurian) of southern Africa and S. kayentakatae from the Kayenta Formation (Sinemurian–Pliensbachian) of Arizona (Raath 1969, 1977; Rowe 1989; Tykoski 1998). Syntarsus is diagnosed by the presence of a small pit below the base of the nasal process of the premaxilla, nasal fenestrae in the skull roof, and a rostrally directed spur of the basioccipital roofing the basisphenoidal recess. It may also be distinguished by the presence of a large pneumatic foramen in the lateral surface of the basisphenoid that is laterally overlapped by the crista prootica. Both taxa are represented by multiple individuals spanning a range of ontogenetic stages.
An early skull reconstruction of S. rhodesiensis based on disarticulated material exhibited an unusually large antorbital fenestra (40% of the skull length) and a lacrimal that overlapped the maxilla and jugal laterally, reaching the alveolar border (Raath 1977). However, comparison with the articulated skull of S. kayentakatae shows that the lacrimal was reversed in the earlier reconstruction. When corrected the lacrimal abuts the maxilla and jugal ventrally, and it also has the effect of shortening the preorbital length of the skull. With these changes S. rhodesiensis lacks unambiguous apomorphies beyond those of Syntarsus ancestrally.
Syntarsus kayentakatae is unambiguously diagnosed by the caudal margin of the frontals being separated on the midline by a short rostral projection of the parietals and the presence of a faint groove across the cranial surface of the astragalus, as in Dilophosaurus wetherilli (Welles 1983, 1984). It may also be diagnosable by its low nasal crests and the presence of a promaxillary fenestra (both reportedly absent in S. rhodesiensis and Coelophysis). It is unlikely that two crested coelophysoids would coexist in the North American southwest during the time represented by Kayenta sediments, but material of both taxa has been collected within a few hundred meters of one another at virtually the same stratigraphic level. Adults of S.kayentakatae are slightly larger than ontogenetically comparable individuals of S. rhodesiensis and have a greater dorsoventral profile to the preorbital region of the skull (Raath 1977; Rowe 1989; Tykoski 1998).
Gojirasaurus quayi is one of the largest theropods known from the Late Triassic, reaching nearly the size of some Dilophosaurus (Parrish and Carpenter 1986; Carpenter 1997a). It is based on incomplete remains from the Cooper Canyon Formation (Norian) of the Dockum Group, New Mexico. The pubis shows slight cranial bowing, and the caudal border of the scapula is straight for most of its length, as in coelophysids (Carpenter 1997a). Our analysis shows Gojirasaurus to be more derived than Dilophosaurus, but it is not diagnosable beyond this level.
Camposaurus arizonensis is based on a small distal tibiotarsus and other fragmentary bones from the upper Carnian Placerias Quarry (lower Petrified Forest Member of the Chinle Formation), near St. Johns, Arizona (Hunt et al. 1998). The astragalus has a low, triangular ascending process as in coelophysoids and Elaphrosaurus. The distal fibula cranially overlaps the ascending process and is fused to the calcaneum, a condition also present in Syntarsus kayentakatae and Xenotarsosaurus bonapartei (Martínez et al. 1986; Rowe 1989). The proximal femur of C. arizonensis bears remnants of a prominent trochanteric shelf and the partial sacrum is composed of four fused centra. This taxon is a coelophysoid, but at this time it cannot be diagnosed to a more specific level within the clade.
Paleobiology
Ceratosaurs had an essentially global distribution, their remains being found on every continent except Australia and Antarctica. This is especially true of the coelophysoids, which evidently enjoyed a Pangaean distribution during the Late Triassic and Early Jurassic. Ceratosaur fossils are almost always found associated with terrestrial faunas and in a range of depositional environments. Genusaurus sisteronis was collected from marine lime-stones, but faunal and paleobotanical evidence suggest that it was buried in a nearshore setting (Accarie et al. 1995). The fossil record for Ceratosauria spans a minimum of 155 million years, from the late Carnian of the Late Triassic to the end of the Cretaceous. Ceratosaurs evolved into a broad range of sizes and body forms, from lightly built, diminutive taxa such as Segisaurus (1–1.5 m in length) to the large abelisaurids, such as Carnotaurus (10–11 m). The Kayenta Formation (Sinemurian–Pliensbachian) of Arizona has produced the remains of three coelophysoid taxa of different body size, the most diverse ceratosaur fauna yet known (Tykoski 1998). This indicates that ceratosaurs had adapted to a wide range of predatory opportunities and roles in their respective environments.
The distribution of noninterdigitating sutures and contacts in the skull of some reptilian groups was interpreted by some authors as indicating the presence of a system of levers driven by jaw muscles as an aid to predation (Bradley 1903; Versluys 1910; Frazzetta 1962; Iordansky 1968). Bakker (1986) rejected the lever-system hypothesis but argued that the looseness of skull elements in Ceratosaurus allowed the animal to ingest prey items larger than its own head. Welles (1984) also rejected the lever-system hypothesis and interpreted the potential mobility in the skull of Dilophosaurus as a sign of weakness, arguing that the loose connection of the premaxilla precluded the capture and subduing of live prey.
The potential for movement between some cranial bones exists, but it is more likely that any movement was passive in nature, just a bending of the skull to accommodate the stresses imposed while biting and manipulating prey. The fibrous connections between skull elements would have provided shock-absorbing elasticity, ample tensile strength, and flexibility. This is close to Bakker's (1986) hypothesis of cranial flexibility, but there is no evidence for the degree of mobility between elements that would have allowed the swallowing of prey items as large as he envisioned. The unique coelophysoid premaxilla-maxilla articulation and straight, unserrated premaxillary and rostral dentary teeth may have provided the ability for deft manipulation of smaller prey items. The rest of the coelophysoid dentition is powerful, and there is no reason to suppose that the premaxillary flexibility would have prevented these animals from acquiring live prey (contra Welles 1984).
With the exception of Carnotaurus, abelisaurids have unusually low-crowned teeth, different in proportion from those of Ceratosaurus and most tetanurans of comparable size (Chatterjee and Rudra 1996; Sampson et al. 1998; Lamanna et al. 2002). It may be safe to assume that the extremely short forelimbs and exceptionally short manus of Carnotaurus (and possibly other abelisaurids) were not used to aid in catching prey. Abelisaurid skulls bear a large nuchal crest and a dorsally projecting parietal eminence. These would have provided large areas and lever arms for powerful neck musculature. It is possible that the large nuchal crest and parietal eminence evolved in response to reduced forelimbs and smaller tooth crowns.
Most ceratosaurs and many tetanurans for which adequate cranial material is known possess some sort of cranial ornamentation. Ceratosaurus nasicornis, Dilophosaurus wetherilli, and Syntarsus kayentakatae possess autapomorphic cranial crests. Those of Dilophosaurus and S. kayentakatae are too fragile for anything but display purposes. The medial nasal horn and lacrimal hornlets of Ceratosaurus were more robust structures, but these and its median dermal osteoderms might have served a similar display function. The grotesque supraorbital horns of Carnotaurus sastrei may have been able to absorb more physical stresses than the cranial structures mentioned above, presenting the possibility of more physical infraspecific contests in this taxon. X-ray CT scans reveal the median frontal dome of Majungatholus atopus to be hollow, reducing the likelihood that the structure was used in excessively physical infraspecific fighting (Sampson et al. 1998). The fused nasals of abelisaurids are notable for their rugose, pitted external texture (Bonaparte and Novas 1985; Bonaparte et al. 1990; Sampson et al. 1998). It is possible that in life the nasals were covered by rough, keratinous skin or other non-bony tissue. Widespread cranial ornamentation, the presence of sexual dimorphism in coelophysoids, and taphonomic evidence (see below) all suggest some degree of complex social behavior among ceratosaur taxa.
Several coelophysoid taxa were collected from mass burials, where multiple individuals were preserved together. Syntarsus rhodesiensis is known from at least thirty individuals found at localities in Zimbabwe and South Africa (Gow and Raath 1977; Raath 1977, 1980). The Kayenta Formation has produced several mass burial sites, including one that included the remains of at least three individuals of Syntarsus kayentakatae (Rowe 1989). Dilophosaurus wetherilli was first collected from a locality that yielded three individuals (Welles 1984). Another Kayenta locality has yielded the remains of at least sixteen individuals of a new coelophysoid taxon (Tykoski 1997, 1998). The prolific Ghost Ranch Quarry of New Mexico has yielded thousands of specimens that may represent hundreds of individuals of Coelophysis bauri, and there are still large blocks from this site awaiting preparation (Colbert 1989; Schwartz and Gillette 1994). Liliensternus liliensterni comes from a site that produced at least two individuals (Huene 1934a).
The depositional setting of coelophysoid mass-burial sites is variable. A taphonomic study of the Ghost Ranch site suggested that large numbers of Coelophysis and other taxa were concentrated and died near dwindling water supplies during severe drought and that subsequent refilling and overbank flow transported and buried the multitude of Coelophysis carcasses (Schwartz and Gillette 1994). Syntarsus kayentakatae was collected from a meter-thick sandstone that may have been deposited during flash floods in an ephemeral stream channel. Some Dilophosaurus wetherilli material was preserved in overbank deposits. Raath (1980) reported Syntarsus rhodesiensis burials in thin fluvial lenses within aeolian deposits. The monospecific (or paucispecific) nature of these sites led Welles (1984) to suggest that group behavior contributed to the circumstances that led to these mass accumulations. Among neoceratosaurs only Masiakasaurus knopfleri is known from a site producing multiple individuals—at least six individuals, recovered from the same locality (Sampson et al. 2001).