TWELVE

Prosauropoda
Prosauropod dinosaurs were medium- to large-sized (approximately 2.5 to 10 m), bipedal, facultatively bipedal or quadrupedal sauropodomorphs with long necks and tails (fig. 12.1A, C). They were obligate or facultative herbivores and are usually the most common terrestrial vertebrates in the beds in which they occur. The skull is small, less than half the length of the femur; the jaw articulation is situated slightly to well below the level of the maxillary tooth row; and the dentition consists of small, homodont or weakly heterodont, spatulate teeth with coarse, obliquely angled marginal serrations. Digit I of the manus bears an enormous trenchant ungual. The ilium is low, and the distal part of the pubes forms a broad flat apron. The femur was not fully erect in the parasagittal plane. Ungual phalanx I of the pes is the largest.
Prosauropods have been found on all the major continents (table 12.1), including Antarctica (Hammer and Hickerson 1994; Rich et al. 1997:fig. 5), but the Australian record (Agrosaurus) is based on mislabeled material of Thecodontosaurus from England (Vickers-Rich et al. 1999b; Galton 2000a).
The identification of prosauropod bones from the Middle Triassic (Anisian—Ladinian) of England by Huene (1907–8) was based on the misidentification of rauisuchian and phytosaurian archosaur and dinosauriform remains (Galton and Walker 1996a, 1996b).
The oldest possible prosauropod is Azendohsaurus (Gauffre 1993a), represented by jaws with teeth from the Argana Formation of Morocco (early late Carnian [Lucas 1998; Harris et al. 2002]). However, Azendohsaurus may be Ornithodira incertae sedis rather than dinosaurian (Jalil and Knoll 2002, in prep.). A premaxilla and the caudal part of a lower jaw (Chatterjee 1984:fig. 1a–c, f), originally thought to belong to the ornithischian dinosaur Technosaurus from the Dockum Formation (early late Carnian [Heckert and Lucas 2000]) of Texas, were identified as Prosauropoda indet. by Sereno (1991; see also Hunt and Lucas 1994). An Azendohosaurus-like tooth from the Chinle Group of Texas is slightly younger (latest Carnian [Harris et al. 2002]); the material of Long and Murry (1995:191, fig. 193A–E, H–K) is not prosauropod. Two different kinds of jaws with teeth from the base of Isalo II of Madagascar (Flynn et al. 1999), one Azendohsaurus-like (Flynn 2000:47) and the other with more primitive-looking prosauropod teeth, are also Carnian. The best Carnian record (Heckert and Lucas 2000; Lucas and Heckert 2001; Harris et al. 2002) consists of several articulated skeletons from the Santa Maria Formation of Brazil; those of Saturnalia include impressions of the dentary and teeth (Langer et al. 1999b), whereas those of described but unillustrated forms from a slightly higher Caturrita Formation include skull material (Azevedo et al. 1999; Kellner et al. 1999). The presence of material from three continents indicates a Pangaean-wide distribution of Prosauropoda soon after the first fossil record (Harris et al. 2002), as is also the case for Theropoda (Heckert and Lucas 2000). The earliest prosauropod fauna is from the Lower Elliot Formation (age as Carnian, Shubin and Sues 1991; Carnian and/or Norian, Olsen and Galton 1984; Norian, Lucas and Hancox 2001; early to mid-Norian, Harris et al. 2002) of southern Africa (Euskelosaurus, Melanorosaurus, Blikanasaurus [Heerden 1979; Heerden and Galton 1997; Galton and Heerden 1998]); an earlier record from southern Africa is a proximal femur from the underlying equivalent of the Molteno Formation in Zimbabwe (Raath 1996).
Prosauropods are common in the Norian of Argentina (Coloradisaurus, Mussaurus, Riojasaurus [Bonaparte 1972; Bonaparte and Vince 1979; Bonaparte and Pumares 1995]) and Germany (upper Löwenstein Formation, Sellosaurus [Huene 1907–8, 1932]; Trossingen Formation ?into Rhaetic, Plateosaurus [Huene 1926a, 1932; Galton 2001a, 2001b]; Ruehleia [Galton 2001a, 2001b]); complete skeletons of Plateosaurus also occur in the Norian of Greenland (Jenkins et al. 1995). The incomplete skeleton of Camelotia is from the Rhaetian of England (Galton 1998b), and the first prosauropod to be described, Thecodontosaurus Riley and Stutchbury, 1836, is represented by many bones from fissure fills that are probably the same age (Benton et al. 2000b). Buffetaut et al. (1995c) describe the distal part of a robust prosauropod ischium from the Nam Phong Formation (Rhaetian) of Thailand, but it might be from the Nam Phong sauropod Isanosaurus, the ischium of which is unknown (Buffetaut et al. 2000b).
It has long been assumed that prosauropods were restricted to the Triassic, but based on the transfer of several important Upper Triassic beds that have yielded remains to the Lower Jurassic, prosauropods continued to be important until at least the Pliensbachian or Toarcian (Olsen and Galton 1977, 1984; Olsen and Sues 1986; Shubin and Sues 1991; Luo and Wu 1994; Sues et al. 1994a). These beds include the Upper Newark Super group and the Glen Canyon Series of North America (Ammosaurus, Anchisaurus, Massospondylus [Galton 1976b; Attridge et al. 1985]), the Upper Elliot and Clarens formations and equivalents of southern Africa (Massospondylus [Cooper 1981b]), the Cañón del Colorado Formation of San Juan, Argentina (Massospondylus [Martínez 1999, 2002a]), and the Lower Lufeng Series of Yunnan, People's Republic of China (“Gyposaurus” sinensis, Jingshanosaurus, Lufengosaurus, Yimenosaurus, Yunnanosaurus [Young 1941a, 1941b, 1942a, 1947, 1948a, 1951; Bai et al. 1990; Zhang and Yang 1995]). Kutty (1969) reported prosauropods from the Upper Triassic Dharmaram Formation of India, but these beds are now regarded as Lower Jurassic (Bandyopadhya and Roy-Chowdhury 1996); fragmentary remains of a Massospondylus-like form and almost complete skeletons of a melanorosaurid-like form are represented (Kutty et al., in prep.).
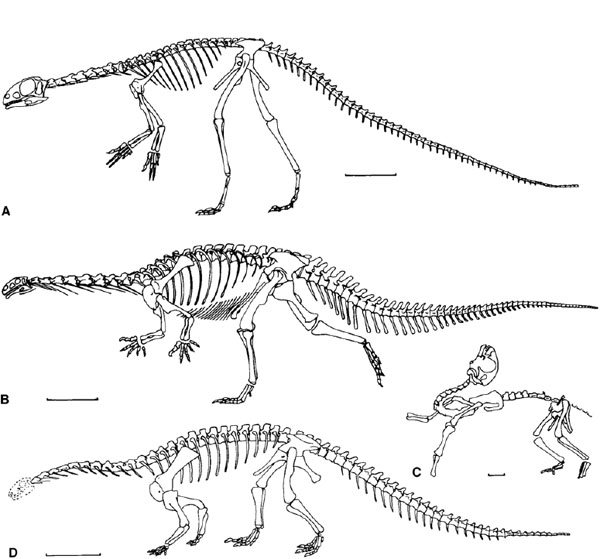
FIGURE 12.1. Prosauropod skeletons: A, Thecodontosaurus, juvenile individual; B, Plateosaurus; C, Mussaurus, juvenile individual; D, Riojasaurus. Scale = 10 cm (A), 50 cm (B, D), 1 cm (C). (A after Kermack 1984; B after Huene 1926a; C after Bonaparte and Vince 1979; D after Bonaparte 1972.)
Anchisauridae and Plateosauridae were erected by Marsh (1885, 1895b), but it was not until 1920 that Huene recognized Prosauropoda, which he combined with Sauropoda to form Sauropodomorpha Huene, 1932. The various systematic positions and constituents of Prosauropoda were reviewed by Colbert (1964a) and Charig et al. (1965) and summarized by Steel (1970), who also reviewed all the species of prosauropods. Prosauropoda has long been regarded as a paraphyletic taxon in which some genera are more closely related to sauropods than others (Romer 1956; Colbert 1964a; Charig et al. 1965; Charig 1982; Bonaparte 1986; Gauthier 1986; Benton 1990a; McIntosh 1990a; Heerden 1997; Benton and Storrs in Benton et al. 2000b), but it is here considered to be monophyletic, as it is in most other recent studies (Sereno 1989, 1997, 1998, 1999a; Galton 1990a; Upchurch 1993, 1995, 1998; Gauffre 1995a, 1996; Novas 1996a; Wilson and Sereno 1998; Galton and Upchurch 2000; Hinic 2002a, 2002b; Martinez 2002a, 2002b; Upchurch and Galton, in prep.).
Definition and Diagnosis
The stem-based name Prosauropoda is defined as all taxa more closely related to Plateosaurus than to Sauropoda. Prosauropoda can be diagnosed by the following synapomorphies: a lateral lamina on the maxilla; a straplike ventral process of the squamosal; a ridge on the lateral surface of the dentary; a caudally inset first dentary tooth; elongate caudal dorsal centra (the length-height ratio greater than 1.0; highly variable, possibly size-related); the absence of the prezygadiapophyseal lamina on the caudal dorsals; the deltopectoral crest oriented at right angles to the long axis through the distal humeral condyles (reversed in “Gyposaurus” sinensis, Jingshanosaurus, and Melanorosaurus); the transverse width of distal carpal 1 greater than the width of metacarpal I (reversed in Sellosaurus); phalanx I on manual digit I having a proximal “heel”; phalanx I of manual digit I having its proximal and distal articular surface long axes twisted at 45° to each other; a large pubic obturator foramen (multiple reversals).
TABLE 12.1
Prosauropoda



Anatomy
Skull and Mandible
Articulated and disarticulated cranial material of Plateosaurus (figs. 12.2, 12.5E) was redescribed by Galton (1984a, 1985a; see also Huene 1926a, 1932). Unless indicated to the contrary, the following references were used for particular genera: Anchisaurus (figs. 12.3A–C, 12.5A; Huene 1914a; Galton 1976), Azendohsaurus (Dutuit 1972; Gauffre 1993a), Coloradisaurus (figs. 12.3G, H, 12.5C; Bonaparte 1978b), “Gyposaurus” sinensis (Young 1941b, 1948a:fig. 33), Jingshanosaurus (fig. 12.4J–L; Zhang and Yang 1995), Lufengosaurus (fig. 12.3N–P; Young 1941a, 1947, 1951), Massospondylus (figs. 12.3J–M, 12.4E–I; Cooper 1981b; Attridge et al. 1985; Crompton and Attridge 1986; Gow 1990a; Gow et al. 1990; MacRae 1999:203), Mussaurus (fig. 12.3I; Bonaparte and Vince 1979; Casamiquela 1980), Riojasaurus (fig. 12.4A–D; Bonaparte and Pumares 1995; Wilson and Sereno 1998:fig. 36A), Saturnalia (Langer et al. 1999b), Sellosaurus (Galton 1985b, 2001a; Galton and Bakker 1985), Thecodontosaurus (figs. 12.3D–F, 12.5B, D; Huene 1907–8, 1914a; Kermack 1984; Benton et al. 2000b), Yimenosaurus (fig. 12.4M; Bai et al. 1990), and Yunnanosaurus (fig. 12.3Q, R; Young 1942a, 1951).
The skull of Plateosaurus is usually characterized as being narrow relative to its height, whereas that of Massospondylus is proportionally wider and lower (figs. 12.1A, B; 12.3J–L); this is the basis for the node-based Plateosauridae and Massospondylidae Huene, 1914b, of Sereno (1998, 1999a, 1999b). However, these differences may be more the result of differential preservation of a readily deformable framework. Skulls preserved on their side were subjected to lateral compression, resulting in narrow skulls, whereas those preserved “upright” were dorsoventrally compressed to give wide skulls. Thus one skull of Plateosaurus is wide and low (Galton 1985a:fig. 4), and skulls of Massospondylus are narrow and high (figs. 12.3M, 12.4E–G; Gow et al. 1990: figs. 1–3). Indeed, Gow et al. (1990:52) conclude that Massospondylus has a proportionally larger orbit and a deeper skull than Plateosaurus (ratio of skull height to skull length 35% in the former, 25% in the latter, but note the proportionally shorter snout in Massospondylus).
The external naris is large (more than 60% of the orbit diameter) in all prosauropods except Anchisaurus, Coloradisaurus, and Massospondylus (where they are 50% or less). The opening is oval to elliptical in Coloradisaurus, Jingshanosaurus, and Massospondylus; in the other prosauropods it is subtriangular with a right angle at the caudoventral corner, a result of the more upright ascending process of the maxilla. The opening is enclosed by the elongate dorsal and caudal processes of the premaxilla and by the nasal in Plateosaurus and Sellosaurus, whereas in other prosauropods the maxilla borders this opening caudoventrally. There is a shelflike area lateral to the external nares and extending onto the rostral end of the maxilla. The transverse width of the internarial bar is less than its rostrocaudal width in all prosauropods except Plateosaurus and Sellosaurus, and distally the dorsal process maintains or increases its transverse width. The sutural surface between the pre-maxillae of Plateosaurus is flat except for a prominent excavation into which the rostral processes of the maxillae and vomers fit (fig. 12.2C). The dorsal process of the maxilla meets the lacrimal, but the contact is hidden by an overlapping part of the nasal in Lufengosaurus, Massospondylus, and Sellosaurus; the condition is variable in Plateosaurus. The low lateral lamina of the maxilla extends caudally from the base of the dorsal process of the maxilla. This lamina is short and merges with the body of the maxilla in Anchisaurus, Jingshanosaurus, Lufengosaurus, Thecodontosaurus, and Yunnanosaurus. It is long in Coloradisaurus, Massospondylus, Plateosaurus, Riojasaurus, and Yimenosaurus. The medial lamina of the maxilla backs the rostral part of the antorbital fenestra to form an antorbital fossa that is small in all prosauropods except Coloradisaurus, Jingshanosaurus, Plateosaurus, Sellosaurus, Yimenosaurus, and especially Riojasaurus. There is one vascular foramen directed caudally, and there are five or six directed rostrally. The maxillary tooth row terminates just rostral to the orbit in Anchisaurus, Thecodontosaurus, and Yunnanosaurus, underlaps the orbit to a slight extent in Jingshanosaurus, Massospondylus, Riojasaurus, and Yimenosaurus, and extends caudally to a level approximately below the middle of the orbit in Coloradisaurus, Lufengosaurus, Mussaurus, and Plateosaurus.
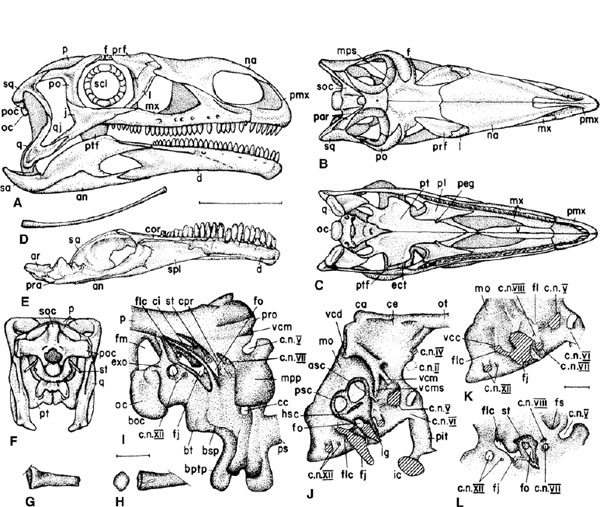
FIGURE 12.2. Cranial anatomy of Plateosaurus. A-C, restoration of skull in A, right lateral, B, dorsal, and C, ventral views. D, right ceratobranchial in lateral view. E, left mandible in medial view. F, reconstruction of skull in caudal view. G, H, proximal part of stapes: G, left in caudomedial view; H, right in ventromedial view. I, braincase in right caudolateral view to show foramina. J, K, endocranial cast in right lateral view: J, with semicircular canal restored; K, with inner ear and adjacent structures removed. L, left medial wall of braincase in oblique view to show foramina. Scale = 10 cm (A-F), 1 cm (G, H), 4 cm (I-L). (After Galton 1984a, 1985a.)
In all prosauropods except Plateosaurus the length of the nasal is less than half that of the skull roof. The rostral end is forked to border the external nares; the bases of the rostroventral and rostral processes are equal in width in Jingshanosaurus, but in all other prosauropods the former is 50% wider than the latter. There is a median nasal depression caudal to the external nares in Lufengosaurus, Massospondylus, Plateosaurus, Sellosaurus, and Yunnanosaurus but not in Jingshanosaurus; the region is not preserved in other prosauropods. The nasolacrimal canal extends from the orbital margin to the rostral end of the lacrimal. The lateral lamina of the lacrimal encloses the caudolateral corner of the antorbital fenestra, while the medial lamina backs the antorbital cavity. These laminae are narrow except in Plateosaurus, which has a wide lateral lamina, and Coloradisaurus, which has a wide medial lamina. As in other archosaurs, the antorbital cavity probably enclosed a paranasal air sinus (Witmer 1997a, 1997b).
The prefrontal wraps around the lacrimal, overlaps the frontal, and forms the rostrolateral part of the orbital rim. The orbital part of the prefrontal is short and transversely narrow in Coloradisaurus and Massospondylus, longer but still narrow in Anchisaurus, Sellosaurus, and Thecodontosaurus, longer and transversely broad in Riojasaurus, and elongate and broad in Lufengosaurus, Plateosaurus, and Yunnanosaurus. The prefrontal has a long ventral process medial to the orbital margin of the lacrimal in Coloradisaurus, Massospondylus (Attridge et al. 1985), Plateosaurus, and Sellosaurus.
The frontal provides an important contribution to the orbital rim in Anchisaurus, Coloradisaurus, Massospondylus, Riojasaurus, Sellosaurus, Thecodontosaurus, and Yimenosaurus. However, the frontal is mostly excluded from the orbit by the overlapping prefrontal and postorbital in Plateosaurus (fig. 12.2B); it is likely similar in Lufengosaurus and Yunnanosaurus. The frontal is excluded from the supratemporal fossa by contact between the parietal and postorbital in all prosauropods except Plateosaurus and Sellosaurus. The frontal and the adjacent parts of the parietal and postorbital of Plateosaurus are deeply excavated to form the area of origin of M. pseudotemporalis. This attachment area is less prominent in other prosauropods but recognizable in Massospondylus and Thecodontosaurus. The parietals are separate bones in Anchisaurus, Riojasaurus, Sellosaurus, and Thecodontosaurus and are partly or completely fused in Coloradisaurus, Lufengosaurus, and Yunnanosaurus. Both conditions exist in Massospondylus and Plateosaurus; but as only parietals of juveniles are known for Sellosaurus and Thecodontosaurus, the condition is not known for adults of these genera. A parietal foramen is present in some individuals of Plateosaurus and absent in others.
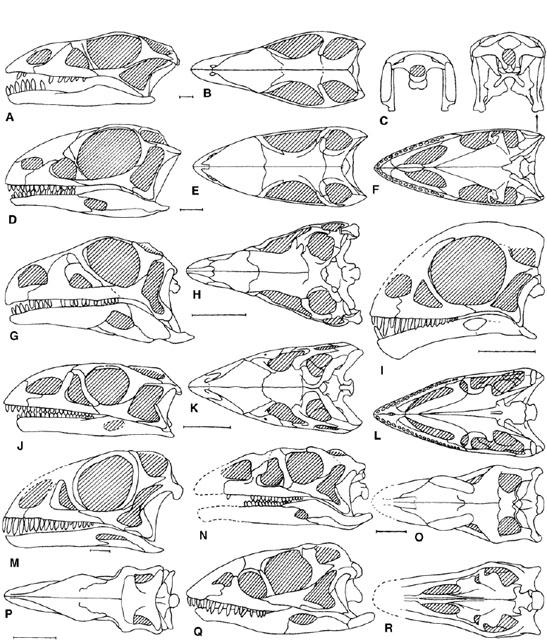
FIGURE 12.3. Reconstruction of prosauropod skulls in left lateral, dorsal, ventral, and caudal views: A–C, Anchisaurus in A, left lateral, B, dorsal, and C, caudal views; D–F, Thecodontosaurus, juvenile in D, left lateral, E, dorsal, and F, ventral views; G, H, Coloradisaurus in G, left lateral, and H, dorsal views; I, Mussaurus, juvenile in left lateral view; J–L, Massospondylus in J, left lateral, K, dorsal, and L, ventral views; M, Massospondylus, juvenile in left lateral view; N–P, Lufengosaurus in N, left lateral, O, dorsal, and P, ventral views; Q, R, Yunnanosaurus in Q, left lateral, and R, dorsal views. Scale = 1 cm (A–F, I, M), 5 cm (G, H, J–L, N–R). (A–C after Galton 1976b; D–F after Kermack 1984; G, H, after Bonaparte 1978b; I after Bonaparte and Vince 1979; J–L after Attridge et al. 1985; M after Cooper 1981b; N–P after Young 1941a, 1951; Q, R, after Young 1942a.)
The tetraradiate squamosal overlaps the parietal, the opisthotic, the postorbital, and the quadrate. In sauropods and theropods the ventral process of the squamosal in lateral view is formed by an extension and gradually narrowing of the main body of the squamosal (i.e., it is tablike), whereas in prosauropods the area caudal to the ventral process is more sharply distinguished from the main body, giving the bone a distorted T shape. A socket in the body of the squamosal is for the dorsal head of the quadrate. The quadrate is rostroventrally directed in Anchisaurus, Thecodontosaurus, and Yimenosaurus and vertical or caudoventrally directed in the remaining genera. The mandibular condyle is located slightly below the maxillary tooth row in Anchisaurus, Jingshanosaurus, Massospondylus, Riojasaurus, Thecodontosaurus, Yimenosaurus, and Yunnanosaurus and is located well below the level of the dentary tooth row in Coloradisaurus, Lufengosaurus, Plateosaurus, and Sellosaurus.

FIGURE 12.4. Reconstructions of prosauropod skulls in left lateral, dorsal, and caudal views and braincases in ventral, rostral, and left lateral views: A-D, Riojasaurus skull in A, left lateral, B, dorsal, and C, caudal views and braincase in D, ventral view; E-I, Massospondylus skull in E, left lateral, F, dorsal, and G, left lateral views and braincase in H, rostral, and I, left lateral views; J-L, Jingshanosaurus skull in J, left lateral, and K, dorsal views and braincase in L, left lateral view; M, Yimenosaurus skull in left lateral view. Scale = 5 cm (A-G, J, K, M), 1 cm (H, I, L). (A-D after Bonaparte and Pumares 1995 and Wilson and Sereno 1998; E-G after Gow et al. 1990; H, I, after Gow 1990a; J-L after Zhang and Yang 1995; M after Bai et al. 1990.)
The V-shaped quadratojugal overlaps the quadrate and jugal. The large pterygoid ramus of the quadrate overlaps the pterygoid medially. The rostral edge of the lateral wall of the quadrate is overlapped by the squamosal and quadratojugal, so the quadrate is excluded from the border of the infratemporal fenestra. The rostral and dorsal processes of the quadratojugal form an angle of 90° in Anchisaurus, Jingshanosaurus, Mussaurus, Riojasaurus, and Thecodontosaurus; this angle is 50°–60° in Massospondylus and Yunnanosaurus and less than 45° in Coloradisaurus, Lufengosaurus, and Plateosaurus. The apical part of the quadratojugal in Plateosaurus fits into a prominent depression on the main body of the quadrate. The Y-shaped jugal overlaps the maxilla, the lacrimal, and the ectopterygoid and is overlapped by the postorbital and the quadratojugal. The dorsal part bordering the orbit is transversely thick, whereas the ventral process is thin and slender, especially in Anchisaurus.
The pterygoids form the major part of the caudal palate. They meet along the midline in Lufengosaurus, Plateosaurus, and Thecodontosaurus, whereas in Massospondylus they apparently do not. In Plateosaurus the pterygoids are small compared with those of Lufengosaurus and especially those of Massospondylus and Thecodontosaurus. The central part of the pterygoid is thick but constricted. From it extend the caudodorsally and laterally directed broad quadrate ramus, the caudomedially and slightly dorsally directed short process that supports a socket for the basipterygoid process of the basisphenoid, the ventrally directed pterygoid ramus, and the rostrodorsally directed and elongate palatal ramus. The complete epipterygoid is known only in Plateosaurus (only the dorsal part is known in Massospondylus); it is a slender bone that overlaps the rostrodorsal part of the quadrate ramus of the pterygoid and the ventral part of the laterosphenoid.
The bar-shaped ectopterygoid is expanded to fit laterally against the jugal and medially against the pterygoid ramus of the pterygoid. The rostrocaudally expanded part of the palatine, which fits against the maxilla laterally, is small in Massospondylus, proportionally larger in Lufengosaurus and Thecodontosaurus, and largest in Plateosaurus. The thin medial processes of the palatine overlap the pterygoid and converge rostrally to meet ventrally before contacting the deep vomer. The palatines occupy half the length of the palate in Massospondylus, in which the vomers are presumably short and hidden by the palatines; a third in Thecodontosaurus; and a quarter in Lufengosaurus and Plateosaurus, in which the vomers are the longest. Ventromedially the palatine of Plateosaurus has a prominent, peglike projection that might have supported the caudal part of a soft secondary palate (also supported by the medial horizontal edge of the maxilla and the vomers).
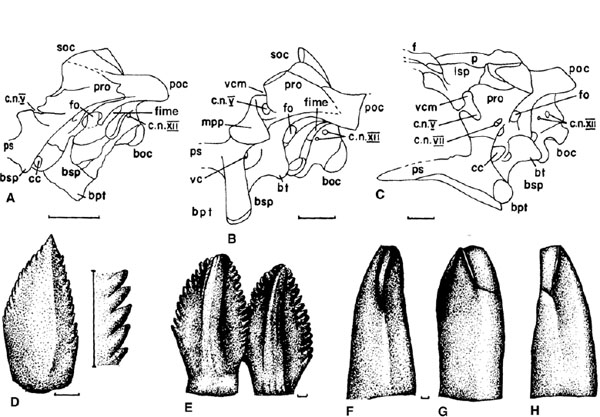
FIGURE 12.5. A-C, prosauropod braincases in left lateral view: A, Anchisaurus; B, Thecodontosaurus; C, Coloradisaurus. D, E, right dentary teeth in buccal view: D, Thecodontosaurus; E, Plateosaurus. F-H, Yunnanosaurus, right maxillary tooth in F, mesial, G, labial, and H, distal views. Scale = 1 cm (A-C), 1 mm (D-H). (A, B, after Galton and Bakker 1985; C after Bonaparte 1978b; D, E, after Huene 1907–8; F-H after Galton 1985c.)
The supraoccipital in Anchisaurus, Massospondylus, Riojasaurus, Sellosaurus, Thecodontosaurus, and Yunnanosaurus is steeply inclined at an angle of 75°, so that its apex is well caudal to the basipterygoid processes. In Coloradisaurus, Lufengosaurus, and Plateosaurus the supraoccipital slopes at 45°, and its apex is approximately above the basipterygoid process. The horizontal paroccipital process is presumably formed from the opisthotic. These processes are set at an angle of 40°–60° to the midline. The exoccipital forms the caudal part of the sidewall of the braincase. The occipital condyle is apparently formed only from the basioccipital in Coloradisaurus, whereas in all other prosauropods the exoccipital contributes dorsolaterally. In Anchisaurus, Jingshanosaurus, Massospondylus, Riojasaurus, Sellosaurus, and Thecodontosaurus the occipital condyle is in line with the parasphenoid, but in Anchisaurus the basal tubera extend farther ventrally than do the small basipterygoid processes (figs. 12.4C, I, L; 12.6A, B). The basipterygoid processes are elongate in Thecodontosaurus and distally expanded in Riojasaurus (figs. 12.4D, 12.6B). In Coloradisaurus, Lufengosaurus, and Plateosaurus the steplike ventral outline of the braincase provides a deep rostral floor to the braincase and the occipital condyle is above the level of the parasphenoid (figs. 12.2I, 12.4C). The basipterygoid processes converge dorsally to form a V-shaped depression in all prosauropods except Plateosaurus, in which this area is flat because of a subvertical transverse lamina with a central projection between the processes (fig. 12.2J). The parasphenoid is a slender rod, subtriangular in cross section, that extends rostrally from the basipterygoid processes and between the pterygoids. A rugose, rostrolaterally facing surface on the adjacent parts of the basisphenoid and prootic is the area of origin of M. protractor pterygoideus in Plateosaurus and Thecodontosaurus (figs. 12.2I, 12.5B). Vena cerebralis media exited through the trigeminal foramen in Thecodontosaurus; in all other prosauropods there is a notch in the portico, above the opening for c.n. V, for this vein (the former condition in juvenile Massospondylus, the latter in adult individuals [Gow 1990a]). The laterosphenoid of Coloradisaurus (fig. 12.5C), Massospondylus (fig. 12.4H, I), Plateosaurus, and Sellosaurus is triangular in lateral view and tapers from the prootic to the transversely expanded rostral end. The orbitosphenoid is only known for Massospondylus (fig. 12.4H, I) and Plateosaurus, in which it is a rib-shaped bone situated immediately rostral to the laterosphenoid.
Information from endocranial casts of Thecodontosaurus (Benton et al. 2000b) and Plateosaurus (Galton 1985a; see also the braincase of Massospondylus, fig. 12.4H, I) suggests that the brain (fig. 12.2J) is short and deep with a short and deep medulla oblongata, a steeply inclined caudodorsal edge to the metencephalon, and prominent cerebral and pontine flexures. In the telencephalon the slightly differentiated cerebral hemispheres form the widest part of the brain. These taper rostrally into elongate olfactory tracts that then widen into large olfactory bulbs. In the diencephalon there is no dorsal diverticulum. The dorsal apex of the diencephalon probably represents a cartilaginous infilling between the ossified supraoccipital and the overlying part of the parietal. C.n. II arose from the diencephalon, the ventral part of which was within part of the pituitary fossa. The extent of the mesencephalon is uncertain because the optic lobes are not differentiated and the points of origin of c.nn. III and IV cannot be determined, only their exit points (fig. 12.4H, I). The metencephalon has no dorsal cerebellar expansion, but the prominent flocullar lobes of the cerebellum do project caudodorsally to occupy the fossa subarcuata in the medial wall of the prootic (fig. 12.2K, L). C.n. V originates from this region, whereas c.nn. VI to XII originate from the myelencephalon (fig. 12.2K). In the inner ear (fig. 12.2K), the rostral semicircular canal is the longest, the lateral canal is the shortest, the sacculus is small, and the lagena is short, as in sauropods. The thin, indented dorsal part of the myelencephalon becomes widest at the level of vena cerebralis caudalis.
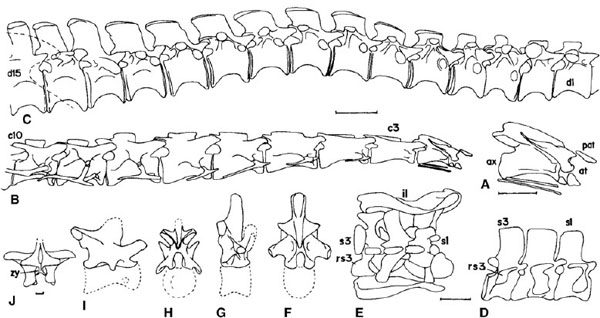
FIGURE 12.6. Prosauropod vertebrae. A-E, Plateosaurus: A, proatlas, atlas, and axis in right lateral view; B, cervical vertebrae 1 to 10 with ribs in right lateral view; C, dorsal vertebrae 1 to 15 in right lateral view; D, sacrum in right lateral view; E, sacrum with ilia in dorsal view. F-I, neural arches of Lessemsaurus: F, cranial dorsal in caudal view; G, cranial dorsal in right lateral view; H, caudal cervical in caudal view; I, caudal cervical in right lateral view. J, Massospondylus, neural arch of mid-dorsal in caudal view. Scale = 5 cm (A), 10 cm (B-E), 1 cm (J); unknown for F-I. (A-E after Huene 1907-8, 1926a, 1932; F-I after Bonaparte 1986; J after Cooper 1981b.)
The dentary curves slightly ventrally toward its rostral tip, with the coarsely ridged medial surface indicative of a firm immovable suture, and the first tooth is inset a short distance. The dentary makes up more than half of the length of the mandible in all prosauropods except Thecodontosaurus. A prominent ridge continues from the lateral surface of the overlapping surangular onto the dentary and passes diagonally across the latter such that the more distal teeth are slightly inset. This ridge is possibly the attachment site for a cheek functionally analogous to that of mammals (Paul 1984a; Galton 1984a, 1985a) and ornithischian dinosaurs (Galton 1973a). In addition, the vascular foramina of the dentary and maxilla are large and few in number in prosauropods, rather than small and numerous as in reptiles without cheeks (Paul 1984a). Cheeks were probably developed to a varying degree in all prosauropods.
The dentary, surangular, and angular border an external mandibular fenestra that is prominent (10%–15% of the length of the mandible) in all prosauropods except Jingshanosaurus, in which it is small (only 5%). The coronoid eminence of the surangular is low, and the jaw articulation is in line with the dentary tooth row in Anchisaurus, Jingshanosaurus, Massospondylus, Mussaurus, Riojasaurus, Thecodontosaurus, Yimenosaurus, and Yunnanosaurus. In Coloradisaurus, Lufengosaurus, Plateosaurus, and Sellosaurus the coronoid eminence is deeper and the jaw articulation is well offset ventrally. The jaw joint is rostrocaudally short. The medial aspect of the mandible is described only in Plateosaurus. The elongate intercoronoid described by Brown and Schlaikjer (1940) and Galton (1984a) is probably the rostral part of the small coronoid that medially overlaps the dentary. The large splenial overlaps the dentary and the prearticular, more caudally overlapping the surangular and the articular, the middle part of which is constricted dorsoventrally. The funnel-like caudal part of the prearticular overlaps the articular, the transversely broad central part of which articulated with the quadrate. The stout retroarticular process of the articular (and overlapping sheet of surangular) is short in Anchisaurus, Massospondylus, Riojasaurus, and Thecodontosaurus and long in the remaining genera.
The teeth are set in sockets that are bordered lingually by small interdental plates alternating with special foramina of varying sizes. Spacing between Zahnreihen ranges between 2.0 and 3.0 tooth positions. This alternating pattern is found in Plateosaurus, but because the length of the replacement wave decreases from 5 to 4 to 3 tooth positions passing distally along the tooth row, the Z-spacings show a corresponding increase from 2.5 to 2.66 to 3.0.
The dental formulae for the premaxilla, maxilla, and dentary are as follows: Anchisaurus, 5, 11, ?; Coloradisaurus, 3, 23 or 24, 22; Jingshanosaurus, 4, 16, 21 or 22; Lufengosaurus, 5, 19, 25; Massospondylus juvenile, 4, 15, ?; Massospondylus adult from Arizona, 4, 16, 20; Massospondylus adult from South Africa, 4, 18, 17; Mussaurus juvenile, 4, 10–12, ?; Plateosaurus, 5–6, 24–30, 21–28; Riojasaurus, 5, 23 or 24, ?; Sellosaurus, 4 or 5, 25, 22; Thecodontosaurus juvenile, 4, 10, 14, adult dentary 20 or 21; Yimenosaurus, 4, 17–18, 21–23; and Yunnanosaurus, 4, 15, 17 or 18.
The crowns of the teeth of the premaxilla and the rostral part of the dentary of most prosauropods taper apically and are slightly recurved with little mesiodistal expansion, whereas those of the maxilla and the rest of the dentary are more spatulate, transversely compressed, more expanded mesiodistally, and symmetrical in mesial and distal views (figs. 12.5E, 12.7A); all teeth of Thecodontosaurus are recurved (fig. 12.5D). The crowns are oriented slightly obliquely relative to the long axis of the maxilla and dentary, so that the distal edge of one tooth slightly overlaps the mesial edge of the tooth behind it, an en echelon arrangement. The lingual surfaces of the crowns are convex or nearly flat mesiodistally, and the base is wide. The root is circular in cross section. The labial surface of the crown bears a central thickening, so that it is slightly more convex mesiodistally than the lingual surface, but there are no prominent vertical ridges as seen in many ornithischians. The marginal serrations are prominent (high notches, or Spitzkerben, of Huene 1926a) and set at an angle of 45° to the cutting edge. The mesial edge has fewer serrations, which are found farther from the root than on the distal edge. In Riojasaurus the crowns are more conical and lack the constriction at the base found in many other prosauropods. The teeth are slender and elongate in Anchisaurus, Azendohsaurus, Massospondylus, Mussaurus, and Yimenosaurus, slender in Coloradisaurus, Jingshanosaurus, Lufengosaurus, Saturnalia, Sellosaurus, and Thecodontosaurus, and broad in Plateosaurus. Tooth wear is usually absent, but food-wear facets, in the form of a smooth flattish arc, are described for the apices of in situ teeth of Massospondylus (Gow et al. 1990) and isolated teeth referred to Plateosaurus (fig. 12.7C; Buffetaut and Wouters 1986; Cuny 1996; Galton 1998a).
The maxillary and dentary teeth of Yunnanosaurus (Young 1942a, 1951) are transversely asymmetrical, the apices being directed slightly lingually, and the mesial teeth are the largest, decreasing in size distally. The labial surface is uniformly convex vertically, whereas the lingual surface is concave except near the root. The teeth of Y. robustus are “advanced spatulated, almost given every detail of the teeth of a sauropod” (Young 1951:61, fig. 2). Vestiges of the coarse, 45° inclined marginal denticles are retained on the distal edge of dentary teeth and on both edges of a few maxillary teeth (Young 1942a, 1951). There is a distinct groove developed along the labiodistal edge on both sets of teeth and also on the linguomesial edge of dentary teeth. A maxillary tooth referred to Yunnanosaurus has wear facets on the distal and mesial edges (fig. 12.5F–H; Galton 1986a:fig. 16.3B–D, unworn teeth fig. 16.3E–I). The distal edge, formed by tooth-food wear, is slightly curved with a smooth, slightly raised enamel rim bordering the dentine. The mesial edge is a large, flat, obliquely inclined wear surface that faces mesiolingually (fig. 12.5G, H). Similar surfaces on the maxillary teeth of the sauropod Brachiosaurus are thought to have resulted from wear against labiodistal surfaces of reciprocal dentary teeth (Janensch 1935–36). However, these teeth were not found with any diagnostic material of Yunnanosaurus (Simmons 1965). Wilson and Sereno (1998:38; see also Salgado and Calvo 1997) noted that these teeth were incorrectly referred to Yunnanosaurus, in which the in situ teeth are not as strongly medially curved and the roots are not as robust, and are not prosauropod. Barrett (1999, 2000a) agreed, noting that the crowns of in situ teeth are also labiolingually compressed, not almost cylindrical in cross section, and lack wear facets. However, Dong (1992:42) noted that the “teeth are cylindrical but flattened from side to side, presenting a chisel-like appearance. The tip of the tooth tends to be worn off at an angle forming a sharp cutting edge, similar to the teeth of sauropod dinosaurs.” Christiansen (1999, 2000) accepted the referral to Yunnanosaurus, pointing out that the development of cylindrical teeth like those of diplodocids and titanosaurids does not occur in basal sauropods. The isolated “Yunnanosaurus” teeth may belong to an as yet unknown eusauropod, a group already represented by Gongxianosaurus, “Kunmingosaurus,” and Zizhongosaurus from the Early Jurassic of the People's Republic of China (Dong et al. 1983; Zhao 1985; Dong 1992; He et al. 1998; Upchurch et al., this vol.). However, the “Yunnanosaurus” teeth do not possess V-shaped wear facets (interdigitating occlusion), an autapomorphy for Sauropoda (Wilson and Sereno 1998:38), because the distal surface is formed by tooth-food wear, not tooth-tooth wear. These teeth also lack the wrinkled texture of the enamel, a plesiomorphy for Eusauropoda (Wilson and Sereno 1998). Consequently, these teeth may be from an as yet unknown prosauropod. An isolated referred tooth of Plateosaurus from the Norian of France has a continuous wear surface on the apex and adjacent parts of the mesial and distal edges that is in a plane subparallel to the labiolingual tooth axis (fig. 12.7D; Cuny and Ramboer 1991:fig. 3g; Galton 1998a), as well as tooth-tooth wear on an isolated tooth from Switzerland (fig. 12.7B; Sander 1992:fig. 10F). A comparable wear surface is found in sauropods having interdigitating, spoonlike teeth.
The stapes is a slender bony rod, most of which is preserved in a skull of Plateosaurus (fig. 12.2F; also in Anchisaurus and Massospondylus [fig. 12.4H]). The medial footplate, which is pierced by a foramen in Massospondylus (Barrett, pers. comm.), is only slightly expanded, and a short diagonal ridge passes distally onto the shaft (fig. 12.2G, H).
Only three complete sclerotic rings are described for Plateosaurus (fig. 12.2A; Galton 1984a); those of Massospondylus have not yet been described. Each ring is composed of 18 plates grouped into four unequal quadrants. The partial ring of Riojasaurus (fig. 12.4A) is similar. Otherwise, only a few plates are preserved in other prosauropod taxa (e.g., Coloradisaurus).
The hyoid apparatus is represented by a pair of elongate, asymmetrical, rodlike first ceratobranchials preserved caudoventral to the mandibles in Massospondylus and Plateosaurus. These elements vary in length from 50% to 60% of the length of the mandible (fig. 12.2D).
Postcranial Skeleton
The postcranial skeleton tends to be more uniform than the skull among prosauropods, but there is more variability than commonly thought (e.g., Sereno 1997). The main overall difference is that the bones are slender and lightly built in smaller genera (e.g., Ammosaurus, Anchisaurus, Massospondylus, Saturnalia, Sellosaurus, and Thecodontosaurus) and thicker and more heavily built in larger genera (e.g., Euskelosaurus, Jingshanosaurus, Lufengosaurus, Plateosaurus, Riojasaurus, and Yunnanosaurus). Illustrations are given of the vertebral column (figs. 12.6, 12.7E–K), the pectoral girdle and forelimb (figs. 12.7L, 12.8), and the pelvic girdle and hindlimb (figs. 12.9, 12.10). Unless indicated to the contrary, the following references were used for particular genera: Ammosaurus (fig. 12.9A; Huene 1906, 1914a; Galton 1976b), Anchisaurus (figs. 12.7N, 12.9B, G; Huene 1906, 1914a; Galton 1976b), Camelotia (Huene 1907–8, Galton 1998b), Euskelosaurus (Heerden 1979; Gauffre 1996), “Gyposaurus” sinensis (Young 1941b, 1948a; Galton and Cluver 1976:fig. 12C), Jingshanosaurus (fig. 12.7K, L; Zhang and Yang 1995), Lessemsaurus (fig. 12.6F–I; Bonaparte 1986, 1999a), Lufengosaurus (fig. 12.7P; Young 1941a, 1947, 1951; Dong 1992:fig. 29), Massospondylus (figs. 12.6J, 12.7J, 12.8Q; Cooper 1981), Melanorosaurus (Heerden 1977, 1979; Heerden and Galton 1997; Galton et al., in press), Plateosaurus (figs. 12.1B, 12.6A–E, 12.7E, F, 12.8A–K, 12.9; Huene 1907–8, 1926a, 1932; Jaekel 1913–14; Galton 2000b, 2001b), Riojasaurus (figs. 12.8L, 12.10C–F, L; Bonaparte 1972; Bonaparte and Pumares 1995), Ruehleia (Galton 2001b), Saturnalia (Langer et al. 1999b), Sellosaurus (figs. 12.7G, H, 12.10H, I; Huene 1907–8, 1932; Galton 1973b, 1984b, 1985c, 1999b, 2001a, 2001c), Thecodontosaurus (figs. 12.1A, 12.8C; Huene 1907–8; Kermack 1984; Benton et al. 2000b), Yimenosaurus (Bai et al. 1990), and Yunnanosaurus (figs. 12.8M, 12.10J; Young 1942a, 1951).
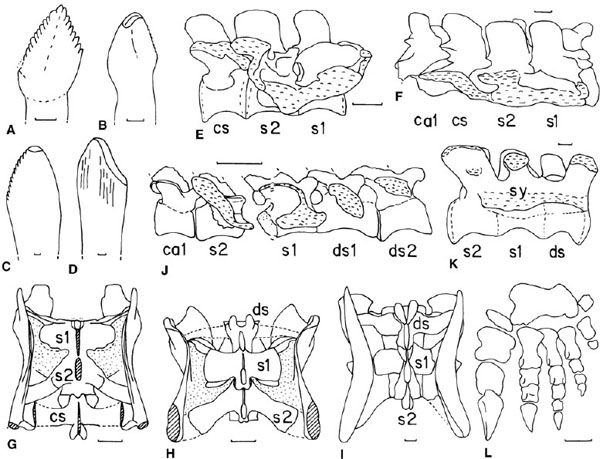
FIGURE 12.7. A, Azendohsaurus, dentary tooth in buccal view. B–D, Plateosaurus, isolated maxillary or dentary teeth showing B, D, tooth-tooth, and C, tooth-food wear. E, F, Plateosaurus longiceps: E, vertebrae sacra of subadult with a caudosacral vertebra in right lateral view; F, vertebrae sacra with adjacent vertebrae of adult with a caudosacral vertebra in right lateral view. G, H, Sellosaurus gracilis, reconstructed: vertebrae sacra with G, a caudosacral vertebra, and H, a dorsosacral vertebra, in dorsal view. I, “Sinosaurus” postcrania, vertebrae sacra with a dorsosacral vertebra in dorsal view. J, Massospondylus, incipient dorsosacral 2, dorsosacral 1, sacrals 1 and 2, and unmodified caudal 1. K, Jingshanosaurus, vertebrae sacra with a dorsosacral vertebra in right lateral view. L, Jingshanosaurus, left manus in cranial view, showing massive carpal block. Cross hatching = broken bone; stipple = matrix; wavy shading = sutural surface for ilium. Scale = 1 mm (A–D), 5 cm (E–L). (A after Dutuit 1972; B after Sander 1992; C, D, after Cuny 1996 and Buffetaut and Wouters 1986; E after Galton 2000b; F after Plieninger 1857; G, H, after Galton 2001c; I after Young 1948a; J after Cooper 1981b; K, L, after Zhang and Yang 1995.)
AXIAL SKELETON
The complete vertebral series of Plateosaurus (figs. 12.1B, 12.6A–E; for presacrals see Bonaparte 1999a:figs. 9–12, also figs. 4–8 for Riojasaurus) comprises the proatlas plus 10 cervicals, 15 dorsals, 3 sacrals, and 50 caudals. The proatlas consists of a pair of thin plates that overlap the zygapophysis-like processes of the supraoccipital and the neural arch of the atlas (fig. 12.6A). Each neural arch of the atlas has an elongate caudodorsal process. The length-height ratio of the centrum of the axis is 3.0 or more, and the postzygapophyses project caudally beyond the end of the centrum. The cervical vertebrae are low and elongate cranially but become higher and eventually slightly shorter caudally. The length-height ratio of the longest postaxial cervical centrum is less than 3.0 in “Gyposaurus” sinensis, Riojasaurus, Saturnalia, and Thecodontosaurus, in which the cervical vertebrae are proportionally shorter, and more than 3.0 in the other genera. In Thecodontosaurus the epipophyses are overhanging and planar and there are unique fossa, possibly pneumatic in origin, on cervical vertebrae 6–8 of the referred juvenile skeleton (Yates 2001). The centra of Plateosaurus are strongly compressed transversely and markedly amphicoelous, especially cranially. The neural spine of the axis is large, while that of the third cervical vertebra is long and low. Passing caudally, the neural spines of cervical vertebra 4 to dorsal vertebra 3 become progressively shorter and taller. The neural spines of the remaining dorsal vertebrae continue to increase slightly in height and especially in length. However, the last three dorsals show a decrease in length of the neural spine. The transverse processes of the cranial dorsal vertebrae are directed strongly dorsolaterally in “Gyposaurus” sinensis and Lufengosaurus but laterally or slightly upward in the remaining genera. The central part of the deep, platycoelous dorsal centra is strongly compressed transversely, and the last five dorsals have lateral pleurocentral indentations. The length-height ratios for caudal dorsal centra are less than 1.0 in Camelotia, Euskelosaurus, Jingshanosaurus, and Lufengosaurus but more than 1.0 in the remaining genera. The middle and especially the caudal dorsal vertebrae have hyposphene-hypantrum accessory articulations for the zygapophyses (fig. 12.6J). The four principal diaphyseal laminae of saurischians (Wilson 1999), the pre- and postzygodiaphyseal and cranial and caudal centrodiapophyseal laminae, are strongly developed in the dorsal vertebrae, much less conspicuous in the cervical region, and absent in the caudals (Bonaparte 1999a). In the first six dorsal vertebrae the thin lamellae are prominent. Prezygapophyseal laminae are absent on the cranial dorsals of Anchisaurus, Massospondylus, and Melanorosaurus but present in other prosauropods. Three cavities below the transverse process are delimited by the two ventral lamellae. The remaining dorsals have only two cavities because of the convergence and fusion of the two cranial lamellae. In smaller prosauropods such as Ammosaurus, Anchisaurus, Massospondylus, Saturnalia, Sellosaurus, and Thecodontosaurus the centra of the dorsal vertebrae are proportionally lower and the lamellae ventral to the transverse process are less prominent. Bonaparte (1986,1999a:figs. 13–16) has described caudal cervical and cranial dorsal vertebrae of Lessemsaurus (fig. 12.6F–I); the tall dorsal neural arches and dorsal neural spines taller than wide axially are sauropodlike characters.
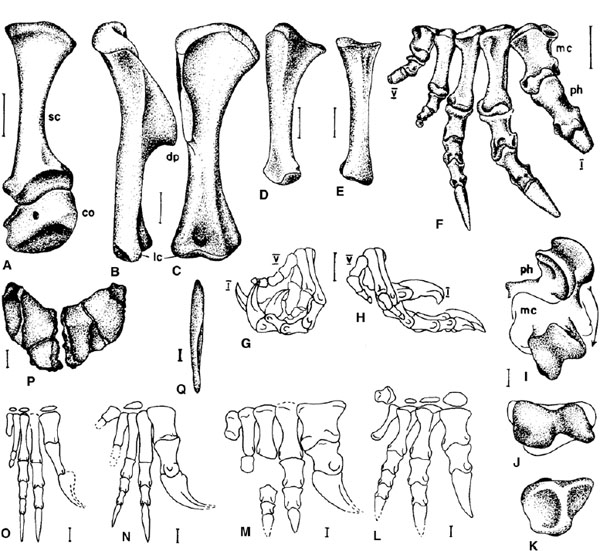
FIGURE 12.8. A–K, pectoral girdle and forelimb of Plateosaurus, from right side. A, scapula and coracoid in lateral view. B, C, humerus in B, lateral, and C, cranial views. D, ulna in lateral view. E, radius in lateral view. F–H, manus: F, in cranial view with digit I partly flexed and digits II to V straight; G, in lateral view with digits fully flexed; H, in medial view with digits in full extension in weight-supporting pose during quadrupedal walking. I–K, metacarpal I and first phalanx: I, phalanx shown in full extension and full flexion on distal end of metacarpal I; J, metacarpal I in distal view; K, phalanx in proximal view. L–O, right manus in cranial view with all digits straight: L, Riojasaurus; M, Yunnanosaurus; N, Anchisaurus; O, Thecodontosaurus. P, Lufengosaurus, sternal plates in dorsal view. Q, Massospondylus, interclavicle in ventral view. Scale = 10 cm (A), 5 cm (B–H, P), 1 cm (I–O, Q). (A–K after Huene 1926a, 1932; L after Bonaparte 1972; M, P, after Young 1947; N after Galton 1976b; O after Galton and Cluver 1976; Q after Cooper 1981b.)
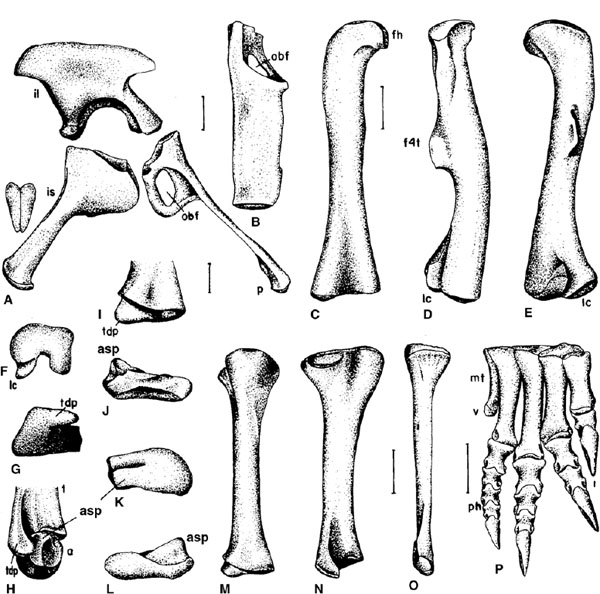
FIGURE 12.9. Pelvic girdle and hindlimb of Plateosaurus, all from right side. A, pelvic girdle in lateral view with distal end of ischia; B, pubis in dorsal view; C-F, femur in C, cranial, D, lateral, E, caudal, and F, distal views; G-I, distal end of tibia in G, distal, H, lateral (with astragalus), and I, cranial views; J-L, astragalus in J, cranial, K, proximal, and L, caudal views; M, N, tibia in M, cranial, and N, lateral views; O, fibula in lateral view; P, pes in cranial view. Scale = 10 cm (A-F, M-P), 5 cm (G-L). (After Huene 1926a.)
Each cervical rib of Plateosaurus is thin, delicate, and more than twice as long as the supporting vertebra (figs. 12.1B, 12.6B) but shorter in Thecodontosaurus (fig. 12.1A). The atlantal rib lacks a tuberculum (fig. 12.1A). In the remaining cervical ribs the line of the shaft is continued beyond the point of divergence of the capitular and tubercular processes as a spinous cranial process. These processes are short in Massospondylus and long in Plateosaurus. The first eight dorsal ribs are especially strong, and in life the slightly thickened distal end was continued ventrally by cartilage. The remaining ribs taper gradually to a point and show a progressive decrease in length. The tuberculum forms a prominent process on the first four ribs, but on the rest it is a small facet dorsolateral to the long capitular process.
Gastralia consisting of slender subparallel rods form a basket-like support for the ventral abdominal wall (fig. 12.1B).
A three-vertebra sacrum with a caudosacral occurs in Ammosaurus, Euskelosaurus, Melanorosaurus (with additional dorsosacral), Plateosaurus, Saturnalia, Sellosaurus, Thecodontosaurus, and Yimenosaurus (figs. 12.6D, E, 12.7E–G; Galton 1999b, 2001b, 2001c). In Ammosaurus the diapophysis and the rib of the caudosacral diverge from each other distally, as also occurs in the last caudosacral of a sacrum of the sauropod Apatosaurus (Wilson and Sereno 1998:fig. 14). However, the third sacral vertebra is a dorsosacral (figures in Galton 1999b) in “Gyposaurus” sinensis, Jingshanosaurus (fig. 12.7K), Lufengosaurus, Massospondylus (fig. 12.7J; also incipient dorsosacral 2), Riojasaurus (Novas 1996a), Ruehleia (Galton 2001b), Sellosaurus (fig. 12.7H; Galton 2000a, 2001a, 2001c), referred “Sinosaurus postcrania” (fig. 12.7I), and Yunnanosaurus. There are 25 presacral vertebrae (15 as free dorsals) in Plateosaurus with a caudosacral but only 24 (with 14 as free dorsals, the number in Ruehleia) in Jingshanosaurus and Lufengosaurus with a dorsosacral, which confirms the identification of the third sacral vertebra as a dorsosacral in the latter genera. The adjacent ends of the ribs of sacrals 1 and 2 commonly fuse together (fig. 12.7E). However, the ribs of the dorsosacral or caudosacral remain unfused except in large individuals, in which the rib fuses to the adjacent sacral rib to form a sacrocostal yoke (fig. 12.7F) that is massive in Jingshanosaurus (fig. 12.7K) and Yimenosaurus. However, the yoke does not form a medial extension to the acetabular surface as occurs in sauropods (see Wilson and Sereno 1998).

FIGURE 12.10. Prosauropod pelvic girdles and hindlimbs, all from right side. A-C, pelvic girdles in lateral view: A, Ammosaurus; B, Anchisaurus; C, Riojasaurus. D-F, femur of Riojasaurus in D, cranial, E, medial, and F, caudal views. G-K, pes: G, Anchisaurus, in caudal view; H, I, Sellosaurus, in cranial view from H, juvenile, and I, larger individuals; J, Yunnanosaurus, in cranial view; K, Riojasaurus, in cranial view. Scale = 10 cm (A, B, G, H, K), 5 cm (C-F, I, J). (A, B, G, after Galton 1976b; C-F, K, after Bonaparte 1972; H, I, after Galton 1973b, 1984b; J after Young 1947.)
The slender, oblique neural spines and transverse processes of the caudal vertebra of Plateosaurus progressively decrease in size distally and disappear at caudal vertebrae 36 and 28, respectively. The neural spines on proximal caudal vertebrae of Sellosaurus and Thecodontosaurus are narrower axially than in other prosauropods. Height is greater than length in the proximal centra, and more distally the centra are lower and longer. Ventral facets on adjoining centra together support a single chevron. The first chevron is a nubbin of bone between the centrum of the caudosacral and the first caudal centrum, with chevrons 2 to 6 long, slender, and increasing in length, but distal to this the chevrons progressively decrease in length (fig. 12.1B). However, in other individuals of Plateosaurus the second chevron is short (Galton 2001a, 2001b). In Riojasaurus the first chevron is borne between caudals 3 and 4 (fig. 12.1D). The chevrons have a club-shaped distal end in Yimenosaurus.
APPENDICULAR SKELETON
The scapula is long and slender with expanded ends, especially ventrally. The suboval coracoid has a prominent notch ventral to the glenoid cavity in all prosauropods except Riojasaurus. In this genus the caudal border is convex and projects ventrally as a prominent process. A slender rodlike clavicle contacts the cranial part of the coracoid in Plateosaurus (Galton 2001b). In Massospondylus the interclavicle is long, moderately broad, and spatulate. It is found adjacent to the ventral edge of the coracoid; the sternal plates caudal to the coracoids consist of a pair of thick, irregular, platelike bones that in life were probably suboval in outline. In Jingshanosaurus and Lufengosaurus the sternal plates fit together to form a heart-shaped shield (fig. 12.8P).
The proximal and distal ends of the humerus are transversely expanded, their axes at an angle of 45° to each other. The large deltopectoral crest points cranially and is perpendicular to the plane of the distal condyles. The apex of the deltopectoral crest is at midlength except in Ammosaurus, Anchisaurus, and especially Thecodontosaurus, in which it is more proximally placed. The radius has an oval, saddle-shaped proximal articular surface and a flat, square obliquely facing distal articulation. The expanded ends of the ulna are at an angle of 40° to each other. The proximal end is triangular in outline, while the distal end has a convex articular surface. In Jingshanosaurus, Lufengosaurus, Riojasaurus, and Yunnanosaurus the proportions of the humerus, radius, and ulna are the same as those of Plateosaurus, but the articular ends are much larger.
The structure of the manus is similar in most prosauropods (figs. 12.7L, 12.8F–O; Galton and Cluver 1976; Galton 2001a, in press). Metacarpal I is slender in Thecodontosaurus, and metacarpals II–V are slender in Anchisaurus, Thecodontosaurus, and a juvenile individual of Sellosaurus. The proximal carpals are rarely preserved in prosauropods, occurring occasionally in Plateosaurus and Sellosaurus, and they were probably represented by cartilage rather than bone. A possible intermedium is pyramidal with a three-sided (Massospondylus) or four-sided (Jingshanosaurus) base. Distal carpals 1 and 2 are preserved in most prosauropods and consist of large and small thin plates with a gently convex proximal surface that cap metacarpals I (and distal carpal 2, Massospondylus) and II, respectively. Occasionally a small distal carpal 3 is also preserved, and rarely, small distal carpals 4 and 5 (Plateosaurus [Huene 1932:pl. 11, fig. 1]). In Jingshanosaurus distal carpal 1 caps metatarsal I, and the massive, irregularly shaped bone (three to four times the size of metacarpal I, their width equal to that of the rest of the metatarsus) represents fused distal tarsals 2–4, with distal tarsal 5 free (fig. 12.7L). In Ruehleia the large distal carpals 1–3 have complex articular surfaces. Metacarpal I is short but stout with a triangular proximal end that fits against distal carpal 1 and the side of distal carpal 2, so that it is wedged into the carpus. The distal ginglymus is asymmetric, extending well onto the extensor surface, so that marked hyperextension was possible. The proximal articular surfaces of the first phalanx of digit I are also unequally developed, and the rotational axis of the distal condyles forms an angle of 45° with that of the proximal articulation (which has a prominent “heel” ventral to it). The trenchant, raptorial ungual of digit I is large, exceeding the size of the largest ungual of the pes. Metacarpal II is slightly longer and more robust than metacarpal III. Digits II and III are subequal in length, cannot be hyperextended, and bear much smaller unguals that are only slightly recurved. Metacarpal IV is slightly shorter and more slender than metacarpal III and bears a vestigial series of phalanges, as does the small metacarpal V.
The body of the ilium is low compared with that of sauropods and theropods. The dorsal margin is smoothly convex except in Melanorosaurus and Riojasaurus, in which it has a “steplike” profile in lateral view. The concave area on the lateral surface extends ventrally to just above the acetabular margin in most prosauropods but is restricted to the dorsal half in Lufengosaurus, Plateosaurus, and Riojasaurus. The preacetabular process is short except in Ammosaurus and Anchisaurus, in which it is proportionally elongate (figs. 12.8A, 12.9A–C). In Ammosaurus, Anchisaurus, and Melanorosaurus it lacks a scar that is present in “Gyposaurus” sinensis, Lufengosaurus, Massospondylus, Plateosaurus, and Sellosaurus (possibly for M. iliofemoralis). The angle the preacetabular process makes with the pubic peduncle is gently curved in all prosauropods except Ammosaurus, Anchisaurus, and Riojasaurus (adults), in which it is acute. The ventral extent of the pubic and ischial peduncles is subequal, so that the inferior ends are nearly parallel to the horizontally oriented long axis of the iliac blade. The brevis shelf is narrow, and the sacral rib of the caudosacral and/or sacral vertebra 2 (figs. 12.6E, 12.7I) articulates with it, while the other sacral rib or ribs suture more cranially against the body of the ilium. The thin supracetabular rim projects laterally and extends along the craniodorsal part of the acetabulum. The acetabulum itself is backed by the ilium extensively in Thecodontosaurus and juveniles of Sellosaurus but only slightly in most prosauropods. The pubis is twisted along its length, with its proximal part forming a deep, thin, oblique subacetabular region that bears an obturator foramen (figs. 12.9A, B; 12.10A–C). This region is shallow in Riojasaurus and still shallower in Anchisaurus, in which the obturator foramen opens ventrally. The obturator foramen is small in “Gyposaurus” sinensis, Jingshanosaurus, Massospondylus, Riojasaurus, and Yunnanosaurus, variable in Plateosaurus, and large in Ammosaurus, Anchisaurus, Lufengosaurus, Melanorosaurus, Saturnalia, Sellosaurus, and Thecodontosaurus. The foramen is partly obscured in cranial view in Jingshanosaurus, Massospondylus, Plateosaurus, Sellosaurus, and Thecodontosaurus but completely visible in the other prosauropods. The main part of the pubis is transversely expanded, and because there is an extensive median symphysis, the two pubes form a platelike apron. This apron is broad in all prosauropods except Anchisaurus, in which it is narrow. In cranial view the lateral margin of the pubis is straight or bows laterally in all prosauropods except “Gyposaurus” sinensis, Lufengosaurus, and Massospondylus, in which it has a concave profile. The ratio of the ischium length to the pubis length is less than 0.9 in Jingshanosaurus, Lufengosaurus, Massospondylus, Melanorosaurus, Plateosaurus, Riojasaurus, and Yunnanosaurus and more than 0.9 in the remaining genera. The ratio of the lengths of the pubic and ischial subacetabular margins is 1.0 in Ammosaurus, Anchisaurus, “Gyposaurus” sinensis, Jingshanosaurus, Riojasaurus, and Yunnanosaurus but 0.5 in the remaining genera. The deep (except in Ammosaurus), obliquely inclined subacetabular part of the ischium sweeps back to the more distal median symphysis (fig. 12.9A). The distal end is only slightly expanded in lateral view relative to the rest of the shaft in Anchisaurus and Thecodontosaurus; in the remaining genera the thickness of the shaft is double at the distal end. The distal end is rounded or flattened in end view in Camelotia, Plateosaurus, and Thecodontosaurus but subtriangular in outline in the other prosauropods.
The femur of many prosauropods is sigmoid in the cranial and lateral views and lacks a distinct neck (fig. 12.9C–E), and the femoral head merges smoothly with the lateral margin of the shaft in cranial view. However, in Anchisaurus, Camelotia, “Gyposaurus” capensis, Jingshanosaurus, Lufengosaurus (Young 1947), Melanorosaurus, Riojasaurus, and Yunnanosaurus (Young 1947) the distal part of the shaft is straight in caudal view, as in sauropods, and the head meets the lateral margin at an abrupt angle in Camelotia, Melanorosaurus, and Riojasaurus. However, in lateral view the cranial face of the femur is convex in all prosauropods. The greater trochanter of prosauropods is represented by a thick lateral edge adjacent to the head. The cranial trochanter consists of a low ridge, all of which lies medial to the lateral edge of the femur, except in Riojasaurus, Melanorosaurus, and Camelotia, in which it becomes a progressively more prominent sheet whose lateral edge is visible in caudal view. The proximal end terminates below the head except in Euskelosaurus, Lufengosaurus, Massospondylus, and Yunnanosaurus, in which it terminates level with the femoral head. Saturnalia has a subhorizontal trochanteric shelf, a structure lost in all other prosauropods (see Novas 1996a). The fourth trochanter is a prominent, platelike structure with a pendant shape in Massospondylus and in some individuals of Lufengosaurus and Yunnanosaurus. It lies close to the midline of the shaft in Euskelosaurus, “Gyposaurus” sinensis, Jingshanosaurus, Massospondylus, Plateosaurus, Sellosaurus, Thecodontosaurus, and Yunnanosaurus. The apex is usually located on the proximal half of the femur, but it extends below midlength in Camelotia, Euskelosaurus, Melanorosaurus, and Riojasaurus. The shaft, which is usually subcircular in cross section, is twisted about its long axis, so that the articular axis of the subequal distal condyles forms an angle of 25° with the axis of the femoral head. In melanorosaurids the femoral shaft is transversely widened and craniocaudally compressed as in sauropods.
The ratio of tibia length to femur length is 0.70 or higher in all prosauropods except Lufengosaurus, in which it is 0.65. The tibia has a prominent, cranially directed cnemial crest, the shaft is triangular in cross section, and the distal end is slightly expanded transversely. A prominent groove deepens laterally to accommodate the ascending process of the astragalus (fig. 12.9G–N). This groove is backed by a prominent, thick descending process formed by the medial malleolus of the tibia. The tibiae of smaller prosauropods such as Anchisaurus, “Gyposaurus” sinensis, Massospondylus, Saturnalia, Sellosaurus, and Thecodontosaurus are proportionally more slender, whereas those of melanorosaurids are more robust with a much larger cnemial crest. The expanded ends of the fibula are set at 40° to each other, so that the distal cranial edge also faces medially. In Massospondylus there is a muscle scar on the lateral surface at midlength.
The large astragalus has a craniolateral ascending process that fits into the grooved distal end of the tibia. In most prosauropods two distal tarsals are preserved capping metatarsals III and IV. The calcaneum is a small, subtriangular bone that contacts the fibula proximally and the astragalus medially.
The form of the pes is uniform. Metatarsals I and V are short, but I is robust and bears a short but strong digit, whereas V is paddle-shaped, being broad and thin proximally but tapering rapidly to a rodlike distal part that bears a vestigial phalanx. Metatarsals I and II have proximal width-length ratios of 0.25 in Anchisaurus, “Gyposaurus” sinensis, Saturnalia, and Thecodontosaurus and more than 0.25 in the remaining genera; in Sellosaurus the former is true for juveniles, and the latter for adults. The proximal end of metatarsal II is subquadrangular in Jingshanosaurus but hourglass-shaped in the remaining genera. Digits II and IV are subequal, and digit III is only slightly longer. Apart from the unguals, all phalanges are longer proximodistally than they are wide transversely except in Camelotia and Melanorosaurus, in which the width exceeds the length in at least some phalanges. Unguals decrease in length from digits I to IV in all prosauropods except Anchisaurus, in which the ungual of digit I is shorter than that of digit II (fig. 12.9G). In smaller forms, such as Anchisaurus, Saturnalia, and Thecodontosaurus, the pes is slender, the plesiomorphic condition. However, the robustness can change ontogenetically, as indicated by the pes of different-sized individuals of Massospondylus (Cooper 1981b) and Sellosaurus (fig. 12.9H, I; Huene 1907–8, 1915, 1932; Galton 1973b, 1985c).
Systematics and Evolution
Basal sauropodomorph relationships have been reconstructed using a data matrix of 137 characters for 1 outgroup and 23 ingroup taxa. The analysis includes prosauropods plus the basal sauropods Barapasaurus (Jain et al. 1975, 1979), Blikanasaurus (Galton and Heerden 1985, 1998), Kotasaurus (Yadagiri 1988, 2001), Shunosaurus (Dong et al. 1983; Zhang 1988; Zheng 1991), and Vulcanodon (Raath 1972; Cooper 1984). Most prosauropod genera were included, but information on Ruehleia (Galton 2001a) and Yimenosaurus (Bai et al. 1990) was obtained too late for inclusion in the current analysis. The basic outgroup framework for Sauropodomorpha follows that of Novas 1996a. The first two outgroups—Theropoda (represented by Eoraptor [Sereno et al. 1993], Herrerasaurus [Novas 1993; Sereno 1994; Sereno and Novas 1993], Staurikosaurus [Colbert 1970; Galton 1977a, 2000a], Coelophysis [Colbert 1989], Dilophosaurus [Welles 1984], and Allosaurus [Madsen 1976a]) and Ornithischia (Lesothosaurus [Sereno 1991], Hypsilophodon [Galton 1974a], and Iguanodon [Norman 1980, 1986])—agreed on most polarity determinations, but when they did not, we used the following supplementary outgroups: nondinosaurian Dinosauromorpha (Lagerpeton, Marasuchus [Sereno and Arcucci 1993, 1994]), Ornithosuchus (Walker 1964), and Scleromochlus (Benton 2000). These outgroups were used to create a hypothetical “Ancestor,” which has been assigned a state of zero for all characters.
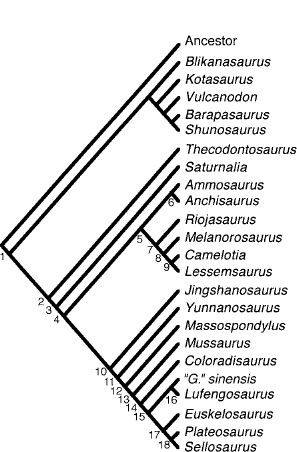
FIGURE 12.11. Cladistic analysis of Prosauropoda: one of the two most parsimonious trees found by a Heuristic Search using PAUP*4.0 (Swofford 1998). The other most parsimonious tree is identical to this except that Massospondylus and Yunnanosaurus have swapped positions. Tree length = 279 steps, CI = 0.541, RI = 0.635, RCI = 0.355.
The data matrix was analyzed using the Heuristic Search in PAUP*4.0 (Swofford 1998). This analysis produced two most parsimonious trees (fig. 12.11) with the following statistics: tree length = 279 steps; CI = 0.541; RI = 0.635; RCI = 0.355.
The following clade diagnoses are based on the delayed transformation optimization (DELTRAN) in PAUP*4.0 (Swofford 1998), which favors convergence over reversal. Character numbers correspond to those in the character list. Character numbers preceded by an asterisk have an equivocal distribution (i.e., accelerated transformation optimization would place the character transformation at a different node). This ambiguity may reflect missing data or the impact of homoplasy. The abbreviation “Con.” followed by the names of one or more taxa indicates that the character has been independently derived in that taxon. Similarly, the abbreviation “Rev.” indicates character reversal. The characters were mapped onto one of the two most parsimonious trees (fig. 12.11) produced by the cladistic analysis. The two most parsimonious trees are so similar in topology that the arbitrary decision to use one of them for character mapping makes little difference to the character-state distributions at most nodes. The two most parsimonious trees differ only in the positions of Massospondylus and Yunnanosaurus (fig. 12.11), and characters relevant to this instability are discussed below.
Node 1, Sauropodomorpha, is defined as all taxa more closely related to Saltasaurus than to Theropoda. Diagnosis of this clade includes the following: the skull length is less than 50% of the femur length (1: Rev. Mussaurus); the diameter of the external naris is at least 60% of the maximum orbit diameter (3: Rev. Anchisaurus and Coloradisaurus); the maxilla enters the margin of the external naris (5: Rev. at node 18); the internarial bar is transversely compressed (6); the infratemporal opening extends forward to a point beneath the orbit (17: Con. Coloradisaurus and at node 18); the supratemporal fenestra is visible in lateral view (23: Rev. Riojasaurus and at node 14); the rostral end of the dentary is deflected ventrally in lateral view (31); the tooth crowns overlap in lateral view (37: Rev. Riojasaurus); tooth serrations are oriented at 45° to the crown long axis (38); there are ten cervical vertebrae (44); the number of sacral vertebrae increases to three via the addition of a caudosacral (62: Rev. Riojasaurus and at node 10, Con. at node 17 [Sellosaurus is dimorphic]); the proximal carpals are absent or fail to ossify (80: Rev. at node 10, Con. Lufengosaurus); the ascending process of the astragalus keys into the distal surface of the tibia (122); metatarsal V is funnel- or paddle-shaped (132); and the pedal ungual on digit I is longer than the other pedal unguals (135: Rev. Anchisaurus).
The definition and diagnosis of node 2, Prosauropoda, is provided in the “Definition and Diagnosis” section above. Basally placed in this clade is Thecodontosaurus antiquus, a small (length ca. 2.5 m), fully bipedal form represented by many isolated bones and a skeleton from a juvenile individual ca. 1 m long from the Late Triassic (Rhaetian) of Britain (figs. 12.1A, 12.3D–F, 12.5B, D, 12.8O; Huene 1907–8, 1914a; Kermack 1984; Benton et al. 2000b). Autapomorphies of Thecodontosaurus include elongate basipterygoid processes; a short dentary (Benton et al. 2000b); over-hanging, planar cervical epipophyses; and a short, dorsoventrally deep preacetabular process of the ilium (Yates 2001).
Node 3, so far unnamed, can be diagnosed based on the following features: the medially and distally located teeth are lanceolate or unrecurved (*42: Con. Sauropoda; alternatively a synapomorphy of Sauropodomorpha that reverses in Thecodontosaurus); the deltopectoral crest terminates at or below the midlength of the humerus (73: Rev. at node 6 and “Gyposaurus” sinensis); and the distal end of the ischium is expanded dorsoventrally (*101: Rev. Anchisaurus and Con. Sauropoda; alternatively a synapomorphy of Sauropodomorpha that reverses in Anchisaurus and Thecodontosaurus).
Saturnalia tupiniquim is a small (1.5 m long), slender, fully bipedal form represented by three partial skeletons lacking skulls (except for an impression of a dentary with teeth) from the Late Triassic (?late Ladinian or early Carnian) of Brazil (Langer et al. 1999b). Although described as a sauropodomorph, it is a basal prosauropod. The femur is extremely plesiomorphic in retaining a prominent trochanteric shelf proximally (cf. Novas 1996a). The position of Saturnalia in the cladogram is controversial insofar as the only other study to consider this taxon (Langer et al. 1999b) argued that it represents a “basal sauropodomorph”; i.e., it is the sister taxon to Sauropoda + Prosauropoda. The latter view is supported on the basis of “overall morphology” (Langer et al. 1999b:4) rather than a suite of synapomorphies uniting prosauropods and sauropods to the exclusion of Saturnalia. As demonstrated by Langer et al. (1999b), it is not disputed that Saturnalia is some form of sauropodomorph. However, our cladistic analysis places Saturnalia within Prosauropoda on the basis of two synapomorphies: an elongate caudal dorsal centra (ratio of length to height greater than 1.0 [53: highly variable, possibly size-related]) and a large pubic foramen (98: multiple reversals [because of missing elements and the requirement for further preparation, the other prosauropod synapomorphies were not examined in Saturnalia]). Placement of Saturnalia “above” Thecodontosaurus is based on three synapomorphies (see diagnosis for node 3, above). Current knowledge of the anatomy of Saturnalia is poor, and it is possible that further preparation and discoveries will reveal new data that support the original view of Langer et al. (1999b). The character-state distributions listed above are clearly equivocal, and therefore the proposed relationships of Saturnalia should be regarded as tentative. However, it is safer to regard Saturnalia as a basal prosauropod.
Node 4, so far unnamed, can be diagnosed on the following synapomorphies: the maxilla bears a foramen directed caudally and five to six directed rostrally (*9); the vena cerebralis media has a separate opening above the foramen for c.n. V (*27); the axial postzygapophyses terminate flush with the caudal end of the centrum (*46); the length-height ratio of the axial centrum is 3.0 or greater (*47); metacarpal I is short and robust (*82); the acetabulum is not backed medially by a sheet of bone (90: Rev. Euskelosaurus, Con. Sauropoda); the distal end of the ischium is subtriangular in outline (*103: Rev. Camelotia and Plateosaurus, Con. Vulcanodon; alternatively a synapomorphy of Sauropodomorpha that reverses in Camelotia, Plateosaurus, Thecodontosaurus, and Eusauropoda [sensu Upchurch, 1995, 1998]); the robustness of metatarsals II and III increases (130: Rev. Anchisaurus and “Gyposaurus” sinensis, Con. Sauropoda); and the ratio of the transverse width to the dorsoventral height of the proximal end of metatarsal IV is 3.0 (*131).
Node 5, Anchisauria, is a new taxon defined as Anchisaurus and Melanorosaurus, their common ancestor, and all its descendants. It is diagnosed based on the following characters: the length of the prefrontal is subequal to that of the frontal (*19: Con. at node 15; alternatively a synapomorphy at node 4 [or more basal nodes] and Rev. Coloradisaurus and Massospondylus); the frontal is excluded from the margin of the supratemporal fossa (*22: Con. Massospondylus, Coloradisaurus, and Lufengosaurus); at least five teeth are found in the premaxilla (35: Con. Sellosaurus [polymorphic] and at node 15); the ratio of forelimb length to hindlimb length is greater than 0.60 (*70: Con. Sauropoda; alternatively a synapomorphy of Sauropodomorpha that reverses at node 10); the pubic foramen is completely visible in the cranial view of the pubis (99: Con. at node 11, Rev. at node 18); the femoral shaft is straight in cranial or caudal view (112: Con. Sauropoda and at node 15); the femoral fourth trochanter is displaced to the caudomedial margin of the shaft (115: Con. Sauropoda and Lufengosaurus); and the proximal end of metatarsal II is hourglass-shaped (*129: Con. at node 10; alternatively a synapomorphy at node 4 that reverses in Jingshanosaurus).
Node 6, Anchisauridae, is defined as all taxa more closely related to Anchisaurus than to Melanorosaurus. Synapomorphies include the following: the length-height ratio of the longest postaxial cervical centrum is at least 3.0 (*48: Con. Eusauropoda [sensu Upchurch, 1995, 1998] and at node 10; alternatively a synapomorphy at node 4 that is convergently acquired by eusauropods and reverses in Riojasaurus and “Gyposaurus” sinensis); the deltopectoral crest terminates at above 50% of the humerus length, a character state reversal (73: reversal Con. “Gyposaurus” sinensis); the preacetabular process of the ilium terminates in front of the distal tip of the pubic process in lateral view (87: Con. Sauropoda); and the angle between the preacetabular and pubic processes of the ilium is acute in lateral view (89: Con. Riojasaurus [polymorphic], Barapasaurus, and Kotasaurus).
Anchisaurus polyzelus is a small animal, ca. 2.5 m long (figs. 12.5A, 12.10B, G; Huene 1906, 1914a; Lull 1953; Galton 1976b), from the Pliensbachian or Toarcian of the Connecticut Valley. An unusual plesiomorphic condition is the transverse narrowness of the distal apronlike part of the pubis. Autapomorphies include large basisphenoid tubera that project farther ventrally than the small basipterygoid processes; ventral emargination of the proximal part of the pubis, resulting in an open obturator foramen; and reduction in the size of the first ungual of the pes, so that it is smaller than the second ungual, a character reversal.
Sereno (1999a) synonymized Ammosaurus with Anchisaurus. The fact that these two taxa are sister groups is consistent with such a synonymy, but clear non-age-related differences between these two taxa (e.g., the emarginated proximal portion of the pubis in Anchisaurus versus emargination of the proximal end of the ischium in Ammosaurus; and the reduced ungual on pedal digit I in Anchisaurus, which is not seen in Ammosaurus) suggest that it is premature to regard them as the same genus with “Ammosaurus” as the adult. Also, a 1 m long juvenile referred to Ammosaurus major has a broad metatarsus, while it is slender in the larger Anchisaurus (Galton 1976b).
Ammosaurus major, from the Pliensbachian or Toarcian of Connecticut, is an animal ca. 4 m long whose skull is unknown (fig. 12.10A; Huene 1906, 1914a; Lull 1953; Galton 1976b). Autapomorphies include the distal separation of the transverse process and the rib of the caudosacral vertebra and the ventral emargination of the proximal part of the ischium. Articulated material from the Pliensbachian or Toarcian of Arizona was referred to Ammosaurus major by Galton (1971b, 1976b) but may represent Massospondylus; other unillustrated partial skeletons from the Early Jurassic of Nova Scotia were described as cf. Ammosaurus sp. by Shubin et al. (1994).
Node 7, Melanorosauridae, is defined as all taxa more closely related to Melanorosaurus than to Anchisaurus. Apomorphies supporting this taxon include the following: a dorsosacral is added to the sacrum (*63: multiple independent acquisitions, including Sauropoda; alternative, equally complex set of transformations present under accelerated optimization); the dorsal margin of the ilium has a steplike sigmoid profile in lateral view (92); the cranial trochanter projects beyond the lateral margin of the femur, thus visible in caudal view (105); the cranial trochanter develops into a prominent sheetlike structure (108: potentially not logically independent from 105); the proximal and lateral margins of the femur meet at an abrupt right angle in cranial view (111: Con. Sauropoda above level of Kotasaurus); the distal end of the fourth trochanter lies at or below the femoral midlength (114: Con. Euskelosaurus and Sauropoda above level of Kotasaurus); and the femoral shaft is transversely wide and craniocaudally compressed (116: Con. Sauropoda).
Riojasaurus incertus is represented by several relatively long (8 m) partial skeletons (fig. 12.1D), one with a skull (fig. 12.4A–D), from the Norian of Argentina (fig. 12.10C–F, K; Bonaparte 1972, 1999a; Bonaparte and Pumares 1995; Wilson and Sereno 1998: fig. 36A). Autapomorphies include a rostral prominence on the premaxilla; a low, rounded lacrimal-prefrontal crest; an expanded distal end to the basipterygoid process; and a coracoid with a cranial process.
Node 8, another unnamed clade, is diagnosed based on the following features: the prezygadiapophyseal lamina is absent on the cranial dorsals (*58: Con. Anchisaurus and Massospondylus); the proximal caudal centra are high relative to their axial length (66: Con. Sauropoda, Lufengosaurus, and Massospondylus [polymorphic]); and at least some pedal phalanges, excluding unguals, are broader transversely than long proximodistally (134: Con. Shunosaurus).
Melanorosaurus readi is represented by a nearly complete skeleton (length 7.5 m) lacking the skull and two other partial skeletons from the early Norian (Lucas and Hancox 2001; Harris et al. 2002), of South Africa (Haughton 1924; Heerden 1977, 1979; Heerden and Galton 1997; Galton et al., in press). However, additional Melanorosaurus material was probably collected, because Kitching and Raath (1984) arbitrarily listed all large prosauropods from the Lower Elliot Formation as Euskelosaurus sp. An autapomorphy is the incorporation of a dorsal vertebra into the sacrum as a fourth sacral vertebra (with retention of the caudosacral).
Melanorosaurus thabanensis is based on a femur from the Hettangian–?Sinemurian of Lesotho (Gauffre 1993c). Autapomorphies include an oblique fourth trochanter far from the medial edge and a cranial trochanter far from the lateral edge. Heerden (1977; see also Charig et al. 1965) noted that the postcranial elements referred to the theropod dinosaur Sinosaurus triassicus (Hettangian to Sinemurian, People's Republic of China) by Young (1948b, 1951) include a mixture of plateosaurid and melanorosaurid elements. He has not elaborated further on this, but if true, it would represent another Early Jurassic record of a melanorosaurid. Welman (1999) described the braincase of a complete skull of a basal prosauropod from the Lower Elliot Formation of South Africa. This specimen, which represents a nearly complete skeleton, was identified originally as Euskelosaurus (Welman 1998; photo in MacRae 1999:202), but it is re-identified as Melanorosaurus by Galton et al. (in press). The skull is characterized by a wide basal fissure between the basioccipital and the basisphenoid and a medially expanded retroarticular process (Welman 1999).
Node 9, as yet unnamed, is diagnosed by a length-height ratio for the caudal dorsal centra of less than 1.0, a character reversal (53: highly variable). Included taxa are Camelotia and Lessemsaurus.
Camelotia borealis is based on a partial skeleton (length 10 m) from the Rhaetian of England (Seeley 1898; Huene 1907–8; Galton 1985d, 1998b). An autapomorphy is the large, sheetlike cranial trochanter.
Lessemsaurus sauropoides is a large animal (length 9 m) based on three individuals represented only by neural arches of 3 cervicals, 14 dorsals, and 2 sacrals plus the centra from 3 cervicals and numerous dorsals from the Norian of Argentina. A caudal cervical and a cranial dorsal vertebra (fig. 12.6F–I) were originally referred to an unnamed melanorosaurid (Bonaparte 1986), but these and other vertebrae from the same series were described later as L. sauropoides (Bonaparte 1999a). Autapomorphies include middle and caudal cervical vertebrae with tall neural arches having a strong transverse infrapostzygapophyseal constriction, deep postspinal fossa, and high infrapostzygapophyseal fossa; and the middle and caudal dorsal vertebrae with neural spines higher than wide (height-width ratio 1.5–2:1). These vertebrae are much more sauropodlike than are those of other prosauropods.
Node 10 represents a clade named Plateosauria (Tornier, 1913), which is defined as Jingshanosaurus, Plateosaurus, their most recent common ancestor, and all of its descendants. Features that diagnose Plateosauria include the following: the retroarticular process is long (35: Rev. Massospondylus); the length-height ratio of the longest postaxial cervical centrum is at least 3.0 (*48: Con. Eusauropoda [sensu Upchurch, 1995, 1998] and at node 6, Rev. “Gyposaurus” sinensis; alternatively a synapomorphy at node 4 convergently acquired in eusauropods and reversed in Riojasaurus and “G.” sinensis); one of the caudosacrals is lost, a character reversal (62: original derived state would have to be reacquired in Plateosaurus and Sellosaurus [polymorphic]); a dorsosacral is added to the sacrum (63: Con. Sauropoda and Melanorosauridae, Rev. at node 17); proximal carpals are ossified, a character reversal (90: Rev. at node 16); and the obturator foramen in the pubis is large, a character reversal (98: Rev. at node 18).
Jingshanosaurus xinwaensis is a medium-sized animal (length ca. 5 m) known from a single partial skeleton (figs. 12.4J–L, 12.7K, L; Zhang and Yang 1995, cervicals 4–10 missing) from the lower Dull Purplish Beds of the Lower Lufeng Series (Hettangian–Pliensbachian [Luo and Wu 1994, 1995; Dong 1995]). Autapomorphies include a small external mandibular fenestra, a massive sacrocostal yoke (developed independently in sauropods), and the massive carpal block formed by the fusion of distal carpals 2–5. The postcrania referred to Sinosaurus by Young (1948b, 1951) from the upper Dark Red Beds of the Lower Lufeng Series were reassigned to this taxon by Zhang and Yang (1995), but this assignment is unlikely because of differences in horizon and in the form of the sacrum (Galton 1999b).
Nodes 11 and 12, so far unnamed, are not diagnosed here because of uncertainty about the relationships of prosauropods. The two most parsimonious trees (fig. 12.11) are identical except for the positions of Massospondylus and Yunnanosaurus. These two genera diverge from the lineages leading to plateosaurids at nodes 11 and 12, swapping places with each other between the two cladograms.
Massospondylus carinatus, from the Hettangian to Pliensbachian of southern Africa, is a medium-sized animal ca. 5 m long (figs. 12.3J–M, 12.4E–I, 12.6J, 12.7J, 12.8Q; Cooper 1981b; Crompton and Attridge 1986:fig. 17.4, 17.5; Gow 1990a; Gow et al. 1990; MacRae, 1999:203 for color photos of skull and of complete skeleton as preserved; Hinic 2002a, 2002b). Autapomorphies include a skull that is at least 10% wider than high with the width of the base of the cultriform process at least 20% of its length (Hinic 2002a) and a prominent muscle scar on the lateral surface of the fibula at midlength. Material referred to Ammosaurus major from the Pliensbachian or Toarcian of Arizona by Galton (1971b, 1976b) may be referable to Massospondylus, a complete skull of which is described from Arizona (Attridge et al. 1985; Crompton and Attridge 1986:fig. 17.6). Based on several partial skeletons, including the skull, Massospondylus is also recognized from the Cañon del Colorado Formation (Lower Jurassic) of San Juan, Argentina (Martínez 1999, 2002a).
Curiously, even though the anatomy of Massospondylus is well known, it is one of the two unstable taxa in the current analysis (indicating that character conflict, rather than missing data, is having an important effect on cladogram topology). To explore this issue further, the character-state distributions relevant to the position of Massospondylus have been examined under delayed and accelerated optimization transformation for both of the most parsimonious trees. Under both delayed and accelerated transformation optimization only one character supports the sister-group relationship between Massospondylus and the plateosaurid clade (Mussaurus, Coloradisaurus, “Gyposaurus” sinensis, Lufengosaurus, Euskelosaurus, Plateosaurus, and Sellosaurus). The synapomorphy is the presence of a long lateral lamina on the maxilla (13: independently acquired in Riojasaurus and lost in Lufengosaurus + “Gyposaurus” sinensis). All other derived characters supporting this node under delayed transformation have a different distribution under accelerated transformation. The only unequivocal character supporting the placement of Yunnanosaurus closer to plateosaurids than is Massospondylus is the presence of a pubic obturator foramen that is completely visible in cranial view (99: Con. Anchisauria, Rev. at node 18). Thus, the evidence supporting each of the alternative placements of Massospondylus and Yunnanosaurus is extremely weak.
Yunnanosaurus huangi, a large animal ca. 7 m long (figs. 12.3Q, R, 12.8L, 12.10J; Young 1942a, 1951) from the upper Dark Red Beds of the Lower Lufeng Series (?Rhaetian–Pliensbachian [Luo and Wu 1994, 1995; Dong 1995]), of the Lufeng Basin, Yunnan, People's Republic of China. The maxillary and dentary teeth are autapomorphic, being weakly spatulate and noticeably asymmetrical, with the apices directed slightly labially and at most only a few coarse apically directed marginal denticles.
Node 13, Plateosauridae, a taxon named by Marsh (1895b), is defined as all taxa more closely related to Plateosaurus than to Yunnanosaurus or Massospondylus. (Ideally only one of the two latter reference taxa would be employed in this definition, but both are used here because of the current uncertainty in their precise relationships.) Plateosauridae is diagnosed by an infratemporal fenestra that does not extend ventrally beneath the orbit, a character reversal (17: Rev. Lufengosaurus).
Mussaurus patagonicus, from the ?Norian of southern Argentina, is represented by extremely small skeletons ca. 25–30 cm long (figs. 12.1C, 12.3I; Bonaparte and Vince 1979). However, a series of intermediate-sized skeletons link these juveniles to larger individuals (length ca. 3 m) from the same site that were briefly described by Casamiquela (1980) as Plateosaurus sp. (Bonaparte, pers. comm.). The latter referral is incorrect because the third sacral vertebra is a dorsosacral in the Argentinean material (Novas, pers. comm.) rather than a caudosacral as in Plateosaurus, the angle between the rami of the quadratojugal is 65° rather than 45°, and there are differences in the size and shape of the antorbital fenestra (Casamiquela 1980, pl. 1). In addition, the dorsal process of the maxilla is narrow and vertically oriented with a small medial lamina, the mandibular symphysis lacks a ventral projection, and the parasphenoid process is located at almost the same level as the occipital condyle (Martínez, pers. comm.). The diagnosis of Mussaurus patagonicus must await a full description of the adult individuals.
Node 14, an unnamed clade, can be diagnosed by the following apomorphies: the caudal half of the premaxillary ascending process is horizontal (*7); the supratemporal fenestra is obscured laterally by the upper temporal bar, a character reversal (23: Con. Riojasaurus); supraoccipital is inclined at 45° in lateral view (26); the parasphenoid rostrum lies below the level of the occipital condyle (*29: Rev. Sellosaurus); and the jaw articulation lies below the tooth row (*34).
Coloradisaurus brevis, from the Norian of northern Argentina, is represented by a deep, wide skull with a short snout and an articulated neck (figs. 12.3G, H, 12.5C; Bonaparte 1978b; Bonaparte and Pumares 1995). Autapomorphies include a wide dorsal process and a large medial lamina to the maxilla and a large medial lamina to the lacrimal.
Node 15, as yet unnamed, is diagnosed by three features: the nasal obscures the dorsal contact between the maxilla and the lacrimal in lateral view (10: Con. Massospondylus); the ratio of prefrontal length to frontal length is greater than 0.95 (19: Con. Anchisaurus and Riojasaurus); and five teeth are found in the premaxilla (36: Con. Anchisauria).
Node 16, also unnamed, is diagnosed by the following features: the transverse processes of the cranial dorsal vertebrae are directed dorsolaterally (56: Con. Shunosaurus); the proximal carpals are absent or fail to ossify, a character reversal (80); the lateral margin of the pubis is concave in cranial view (*96: Con. Sauropoda and Massospondylus); the pubic obturator foramen is completely visible in cranial view (*99: Con. Anchisauria, Euskelosaurus, and Yunnanosaurus); the femoral shaft is straight in cranial view (112: Con. Sauropoda and Anchisauria); and the ungual on pedal digit I is subequal to or longer than other pedal phalanges (136: Con. Sauropoda).
Lufengosaurus huenei is a medium-sized animal (length ca. 5 m) known from several articulated skeletons (figs. 12.2N–P, 12.8P; Young 1941a, 1947, 1951; Dong 1992:figs. 26, 27) from the lower Dull Purplish Beds of the Lower Lufeng Series (?Rhaetian–Bajocian [Luo and Wu 1994, 1995; Dong 1995]). Derived characters include a skull with large prefrontals and an expanded top to the dorsal process of the maxilla, as well as a 0.65 ratio of tibia length to femur length.
“Gyposaurus” sinensis is represented by several partial skeletons 2.3 m in length (Young 1941b, 1948a) that are also from the lower Dull Purplish Beds of the Lower Lufeng Series. It is characterized by short cervical vertebrae, a character reversal. “G.” sinensis has been regarded as a junior synonym of Lufengosaurus huenei, the former being a juvenile of the latter (Rozhdestvensky 1966; Galton and Cluver 1976; Galton 1990a). However, Sereno (1997, 1999a) has argued that the short cervicals of “G.” sinensis, along with other unspecified features, mean that it is a separate taxon and that it probably belongs close to the base of Prosauropoda, along with the other short-necked forms (Saturnalia, Thecodontosaurus, and Riojasaurus, as against the long-necked prosauropods). Sereno (1999a) provided character data for 11 prosauropod taxa, although his cladogram only shows 6 of these (one of the taxa excluded is “G.” sinensis). Rerunning the full data set gives six most parsimonious trees, in each of which “G.” sinensis is variably positioned close to the base of the tree. However, the analysis in this chapter places “G.” sinensis as the sister taxon to Lufengosaurus huenei, a result consistent with synonymizing these two taxa. Changes from the plesiomorphic to the derived condition associated with continued growth could include the changes in proportions of the caudal dorsal centra (change in length-height ratio from less than 1.0 to greater than 1.0 [53]) and the proximal caudal centra (from 1.0 to 0.5 [66]); the more distal shift in position of the deltopectoral crest on the humerus (as occurs in Euskelosaurus and Riojasaurus [73]); and the increased robustness of the pes (as occurs in Massospondylus and Sellosaurus [130]). This explanation is less likely for the proximal shift in the termination of the cranial trochanter (from below the femoral head to level with it [104]); the more medial shift in position of the fourth trochanter on the femur (from the midline to close to the caudomedial margin of the shaft [115]), and the reduction in the ratio of tibia length to femoral length from 0.89 to 0.63–0.66 (117). The ratio of ischium length to pubis length is 0.73–0.77 in Lufengosaurus, compared with 0.9–1.07 in “Gyposaurus” sinensis, the derived condition (97). Also, the third cervical centrum, the longest in the series, is more elongate in a smaller individual of “G.” sinensis than it is in a larger one (Young 1941b:pl. 1, figs. 3, 4); it should be the reverse if “G.” sinensis is a juvenile Lufengosaurus huenei. “G.” sinensis probably represents a distinct species from L. huenei, but further study of the morphology of these two taxa is required to determine if these differences warrant a generic separation. Unfortunately, the skull is poorly represented in “G.” sinensis, but a photograph is given in Dong (1992:fig. 33) of an undescribed skull from the Dafung Basin, Guizhou Province.
Node 17 constitutes an unnamed clade that can be diagnosed by the loss of a dorsosacral (63) and the addition of a caudosacral (62), a character reversal.
Euskelosaurus browni, from the late Carnian or early Norian of southern Africa (Heerden 1979; Gauffre 1993b, 1996), has long been referred to Melanorosauridae (including Cooper 1980), but Heerden (1979) showed that it is a large plateosaurid ca. 10 m long. The large maxilla described by Seeley (1894a) is not prosauropod; it was referred to the basal saurischian Aliwalia rex Galton, 1985c, by Galton and Heerden (1998). Euskelosaurus is characterized by a deltopectoral crest of the humerus that is sigmoid in cranial view.
Node 18 is an unnamed clade diagnosable by the following features: the maxilla is excluded from the external narial margin by contact between the premaxilla and nasal, a character reversal (*5); the frontal is excluded from the margin of the supratemporal fossa, a character reversal (*22); and the proximal end of the cranial trochanter terminates below the level of the femoral head, a character reversal (104).
Plateosaurus longiceps, the common species of Plateosaurus, is the best-represented prosauropod in the world. It is represented by numerous skeletons, some with complete skulls, occurring in mass accumulations in the middle to late Norian at Trossingen (figs. 12.1B, 12.2, 12.5E, 12.6A–E, 12.7E, 12.8A–K, 12.9; Huene 1926a, 1932; Galton 1984a, 1985a, 2001a, 2001b; Weishampel 1984a; Weishampel and Westphal 1986; Sander 1992) and Knollenmergel (Trossingen Formation) of Halberstadt (Jaekel 1913–14; Huene 1932; Galton 1985a, 2001a, 2001b), plus equivalent beds in other parts of Germany (fig. 12.7F), France (Huene 1907–8; Galton 1998a), Switzerland (Huene 1907–8; Galton 1986b), and probably the Norian of Greenland (Jenkins et al. 1995). It is large (length up to ca. 9 m), and the skull has an elongate snout. Autapomorphies include nasal length greater than half the length of the skull roof; a medially directed, peglike process from the middle of the palatine; and a transverse subvertical lamina of the basisphenoid between the basipterygoid processes.
Plateosaurus engelhardti is based on fragmentary remains from the Feuerletten Formation (middle to upper Norian) of Bayern, Germany (Huene 1907–8; Galton 2000a, 2001a, 2001b). The distal part of the femur is possibly straight in craniocaudal view (rather than curved as in P. longiceps), and autapomorphies include sacral rib 1 and 2 originating from the complete length of centrum 1 and from the caudal 75% of centrum 2 (cf. plesiomorphic cranial 50% and 75% in P. longiceps). Additional Fuerletten material with more massive hindlimb bones and a proportionally shorter metatarsus than in P. longiceps may be referable to this species (Wellnhofer 1993b; Galton 2000a); if so, then additional autapomorphies are slightly wedge-shaped centra to caudal vertebrae 3–7 and an open obturator foramen in the pubis (also in Anchisaurus). This species may also be represented by single specimens from Poligny, France, and Halberstadt, Germany (Galton 1998a, 2000a, 2001a, 2001b).
Sellosaurus gracilis is known from several partial skeletons (length up to ca. 6.5 m) from the middle Norian of Germany (figs. 12.7G, H, 12.10H, I; Huene 1907–8, 1915, 1932; Galton 1973b, 1984b, 1985b, 1985c, 1999b, 2000b, 2001a, 2001c, in prep. b; Galton and Bakker 1985). The skull of S. gracilis lacks several of the derived characters of the more recent Plateosaurus longiceps, so it is no longer referred to that genus (Galton 1985b). Sacra have three vertebrae, with the third one either a dorsosacral or a caudosacral (fig. 12.7G, H; Galton 2000b, 2001a, 2001c).
A number of less well known taxa are grouped here under the rubric Prosauropoda incertae sedis. They include Azendohsaurus, Ruehleia, and Yimenosaurus.
Azendohsaurus laaroussi, from the Late Triassic (middle Carnian) of Morocco, was originally described by Dutuit (1972) as an ornithischian dinosaur. Originally known from a dentary, teeth (fig. 12.7A; Dutuit 1972:figs. a, b), and maxilla (Gauffre 1993a:fig. 2), A. laaroussi was first regarded as a prosauropod, but one of the teeth (Dutuit 1972:fig. c) is similar to those of the ornithischian Lesothosaurus (Galton 1984b, 1985e, 1986a, 1990a; Sereno 1991). Gauffre (1993a) noted two tooth forms for Azendohsaurus with the following contrasting sets of characters: 8–12 or 4–6 mesial and distal denticles; the ratio of crown height to mesiodistal width 1.5–2 or 1–1.4; a central vertical ridge present on both faces or absent; the mesial and distal edges either asymmetrical with different denticle counts or symmetrical with the same denticle count; and cingula on both faces or only on the labial face. Most dentary teeth are of the first kind (fig. 12.7A), and most maxillary teeth of the second form, but both are sometimes associated in the same tooth row with teeth of intermediate forms (Gauffre 1993a). Although heterodonty occurs along the tooth rows in prosauropods, this marked difference between maxillary and dentary tooth rows is unique to Azendohsaurus (as is the ornithischian-like form of the second kind of tooth). However, the dentary lacks a lateral ridge, and the first tooth is not caudally inset—two prosauropod synapomorphies (32, 39). Disarticulated unillustrated postcranial bones found with Azendohsaurus show no dinosaurian synapomorphies but, instead, an assortment of plesiomorphic character states (e.g., an ilium with an imperforate acetabulum and no brevis fossa and a femur with a proximally situated fourth trochanter and a head that is not distinctly set off from the shaft) indicating that this taxon is Ornithodira incertae sedis (Jalil and Knoll 2002, in prep.), not dinosaurian. If these associations prove to be correct, then the presence of Azendohsaurus-like teeth (fig. 12.7A) and jaws is not indicative of an early prosauropod and some of the isolated teeth of Triassic ornithischians may have been incorrectly identified by Hunt and Lucas (1994).
Ruehleia bedheimensis is known from a reasonably complete skeleton (cervicals 4–10, dorsals 1–14, a dorsosacral, sacrals 1 and 2, representative sample of 20 caudals, and the appendicular skeleton lacking the pes; length 6.5 m), from the Norian of Thüringen, Germany (Galton 2001a, 2001b). The three-vertebra sacrum has a dorsosacral rather than a caudosacral as in Plateosaurus. The manus has three large carpals with complex articular surfaces; the ilium is plesiomorphic with a large pubic peduncle and an extremely short and small preacetabular process; and for the proximal end of the pubis the length of the articular surface for the ilium is much longer than wide with a short acetabular part (equal to half the width of the iliac surface).
Yimenosaurus youngi is based on four partial skeletons (length 9 m) with partial skulls from the Pliensbachian or Toarcian of Yunnan, People's Republic of China (fig. 12.4M; Bai et al. 1990, only skull illustrated). Autapomorphies include an elongate ascending process of the maxilla (50% of the length of the maxilla; the height of the skull at the antorbital opening 50% of the length), chevrons with club-shaped distal ends, and a large inflated crest on the distal pubis.
Biogeography
Few studies have investigated the biogeographic distribution of prosauropods. Until recently a rigorous cladistic approach to this issue has not been possible because of the lack of a detailed, well-resolved cladogram for the group. Upchurch et al. (2002) applied the cladistic biogeographic method known as component analysis to a large data set on dinosaurian phylogeny and distribution that included information for prosauropods. The results of the analyses for the Late Triassic and Early Jurassic revealed no statistically significant area relationships. One possible explanation for these results is that prosauropod phylogeny did not have a continent-level vicariance pattern imposed on it by Pangean fragmentation. This would not be surprising, given that the major fragmentation events occurred after the extinction of prosauropods (i.e., from the Middle Jurassic onward). Alternatively, prosauropod biogeographic distributions may have displayed a consistent pattern, but that pattern could be obscured by missing data and problematic taxonomic identifications.
Taphonomy
Prosauropods often occur as complete or partial skeletons, but complete skulls are rare. The remains of these animals usually occur in marls, mudstones, and siltstones (e.g., Euskelosaurus, Lufengosaurus, Massospondylus, Melanorosaurus, Plateosaurus, Riojasaurus, Yunnanosaurus), in sandstones (e.g., Ammosaurus, Anchisaurus, Massospondylus, Saturnalia, Sellosaurus), and in eolian sandstones (e.g., Ammosaurus from Arizona). Articulated material of Massospondylus from Zimbabwe occurs in thin, water-laid sediments within an eolian-dominated basin (Raath 1980). Disarticulated remains of Euskelosaurus and Massospondylus from South Africa occur in separate thin beds of mudstone (1–3 m and 2 m thick) with numerous calcareous concretions that represent conditions of subaerial exposure with the production of calcareous soil or caliche (Kitching and Raath 1984). Remains of Thecodontosaurus from Britain occur in Rhaetic fissure fills in Carboniferous limestone as mostly disarticulated bones of adults in a conglomerate (see Huene 1907–8, 1914a; and Benton et al. 2000b) and as articulated bones of juveniles in a stratified marl (see Kermack 1984).
The bones may be distorted, and surfaces are commonly sun-cracked (e.g., Euskelosaurus, Lufengosaurus, Plateosaurus, Yunnanosaurus). Moser (2000) pointed out that the shapes of some of the bones of Plateosaurus from the upper bone bed at Trossingen were modified by pedogenesis. He noted that the increase in size resulted from the swelling of clay minerals within the bone. In this way specimens became unnaturally enlarged, leading to “adult” bones with the proportions of juvenile bones (Huene 1932).
Some prosauropods occur as isolated skeletons (e.g., Ammosaurus, Anchisaurus, Blikanasaurus, Coloradisaurus, Euskelosaurus, Massospondylus), whereas others occur as monospecific mass accumulations. Examples of the latter include Lufengosaurus huenei (Young 1951), Mussaurus patagonicus (Bonaparte and Vince 1979; Casamiquela 1980), Plateosaurus longiceps (Jaekel 1913–14; Huene 1928, 1932; Weishampel 1984a; Galton 1986b, 2001b; Weishampel and Westphal 1986; Sander 1992, 1999a), Riojasaurus incertus (Bonaparte 1982b), Sellosaurus gracilis (Huene 1907–8, 1932; Galton 1985b, 1985c; Hungerbühler 1998b), Thecodontosaurus antiquus (Huene 1907–8, 1914a; Benton et al. 2000b), and Yunnanosaurus huangi (Young 1951). Detailed taphonomic information is available for Plateosaurus longiceps, including a map of their occurrence at Halberstadt (Jaekel 1913–14) and an analysis of 1930s quarry data from Trossingen, Germany (Weishampel 1984a). On the basis of age-class and completeness distributions, Weishampel (1984a) interpreted the lower frequency and less complete remains from two marl bone beds as the result of normal background turnover in an animal that was common in the region. However, he suggested that a concentration of more complete carcasses in the top 1 m of the lower bone bed indicates catastrophic mortality related to the overlying intermediate bed, a layer dominated by bone shards that represents a rapid mudflow. However, Sander (1992, 1999a) and Hungerbühler (1998b, also for Sellosaurus) noted that the remains consist of an almost monospecific assemblage, the caudal parts of the skeletons are preferentially preserved, and in some cases there is an identical burial pose, lying flat on the belly with the limbs projecting downward into the sediment. Their interpretation involves single individuals becoming mired in mudholes situated on an ancient floodplain, with the cranial part or most of the axial skeleton of some of the carcasses being removed by the scavenging activities of predators (smaller theropods [see Hungerbühler 1998b]). Over time this miring resulted in large, local accumulations of prosauropods, but smaller elements of the fauna were absent because they did not become mired and preserved. A quarry diagram of an almost complete skeleton from Trossingen demonstrates disarticulation of the tail, which was slightly scattered as four separated segments by predators (Galton 2001b). This accumulation of carcasses at Trossingen should not be used as evidence for herding, as has been done in the past (Huene 1928), because the period of time involved is not known (Weishampel 1984a; Sander 1992); in addition, one individual every 28 square meters is not exactly what would be expected for a traveling herd (Sander 1999a).
Paleoecology and Paleobiology
Prosauropods are a rare constituent in the late Carnian faunas in which they occur. In the Lower Elliot Formation (Norian) of South Africa and Lesotho, Euskelosaurus is common (Heerden 1979; Kitching and Raath 1984), while Melanorosaurus and especially Blikanasaurus are rare. Other elements of this fauna include a capitosaurid amphibian, a rauisuchian, and a traversodontid therapsid (Anderson et al. 1998b; Galton and Heerden 1998; Lucas and Hancox 2001). This paleofauna is the earliest occurrence of the Prosauropod Empire of Benton (1983a). In the Los Colorados Formation (Norian) of Argentina 40% of individuals are the large (up to 10 m) Riojasaurus (Bonaparte 1982b), while the large Lessemsaurus and the smaller Coloradisaurus are rare (Bonaparte 1978b, 1999a); there is also an undescribed form with a slender manus and pes (Arcucci and Coria 1996). The remainder of this fauna consists of primitive theropods, stagonolepids, rauisuchians, ornithosuchids, sphenosuchid and protosuchid crocodilomorphs, and pachygenelid therapsids (Bonaparte 1982b). Studies of prosauropod-carnivore teeth assemblages from the Norian of Germany show that the large basal archosaurs (heterodont, deep-snouted phytosaurs and rauisuchians) preying on Sellosaurus of the Löwenstein Formation (middle Norian) were replaced by theropod dinosaurs preying on Plateosaurus of the Trossingen Formation (upper Norian [Hungerbühler 1998b]). The numerical predominance of prosauropods in the fossil record continues for the rest of the Norian and into the Early Jurassic (to the Toarcian). Massospondylus from the Upper Elliot and Clarens formations (Lower Jurassic) shared the scene with primitive ornithischians, primitive theropods, protosuchid and pedeticosaurid crocodilomorphs, tritylodontid, tritheleodontid, and diarthrognathid therapsids, and morganucodontid mammals (Olsen and Galton 1984; Anderson et al. 1998b). Based on data given by Benton (1983a), Plateosaurus represents at least 75% of the individual animals from the Trossingen Formation (upper Norian) of Trossingen, Germany, and Lufengosaurus and Yunnanosaurus represent 82% of the individuals from the Lower Lufeng Series (Lower Jurassic) of the People's Republic of China (see Luo and Wu 1994 for associated faunas). In both cases prosauropods are by far the largest animals and certainly formed a majority of the original biomass.
Both raw taxic and phylogenetically corrected estimates of prosauropod diversity indicate that the greatest number of prosauropod lineages existed during the Norian. From this peak, diversity drops rapidly during the Rhaetian and then gradually throughout the Early Jurassic. The factors controlling this diversity pattern are not understood. It is conceivable that competition with radiating sauropod lineages resulted in the decline of the prosauropods. However, given the discovery of Triassic sauropod bones (Isanosaurus, Rhaetian Thailand [Buffetaut et al. 2000b]) and the recent reidentification of Triassic bones (Blikanosaurus, mid-Carnian [Upchurch et al., this vol.]) and footprints (Tetrasauropus, Norian–Rhaetian, United States, southern Africa, Greenland, western Europe [Lockley et al. 2001c]) as those of sauropods, these two groups effectively overlapped geographically for at least 30 million years during the Late Triassic and Early Jurassic. Benton (1996) has argued that long overlaps between groups are unlikely to occur when competitive replacement is an important factor, suggesting that the sauropods were not responsible for the final extinction of the prosauropods. It is interesting, however, that those prosauropods that most closely resemble sauropods in terms of anatomy and body size (melanorosaurids) are a short-lived group and, except for Melanorosaurus thabanensis, do not survive beyond the Triassic–Jurassic boundary. Although speculative, a competitive interaction between sauropods and melanorosaurids remains feasible.
Although some prosauropods were regarded as carnivorous, they are usually considered to represent the first radiation of herbivorous dinosaurs (Colbert 1951; Charig et al. 1965; Romer 1966; Galton 1976b; Bonaparte 1982b; Benton 1983a; Weishampel 1984b; Carroll 1988). The evidence for a herbivorous versus a carnivorous diet is discussed by Galton (1984a, 1985e, 1986a, 1990a, 2001a; see also Attridge et al. 1985; Crompton and Attridge 1986; Gow et al. 1990; and Barrett 2000a); Galton also rejected a scavenging predatory diet with occasional cannibalism in these animals (Cooper 1981b). Part of the reason for the debate concerning the diet of prosauropods is that their masticatory apparatus appears to be poorly adapted for dealing with resistant plant material, especially when compared with that of ornithischian dinosaurs and other Carnian and Norian herbivores (e.g., chelonians, procolophonids, trilophosaurids, rhynchosaurs, mammal-like reptiles such as dicynodonts and tritylodontids, and the small mammals [Galton 1986a; Crompton and Attridge 1986]). Compared with the skulls of most of these herbivores, the skulls of prosauropods are lightly constructed, with noninterdigitating, overlapping, or abutting sutures that tended to separate prior to preservation (see Galton 1985a); there is proportionally much less room for the adductor muscles (Crompton and Attridge 1986); and apart from referred teeth of Plateosaurus and “Yunnanosaurus” (figs. 12.5F–H, 12.7A–C), well-developed tooth-tooth wear facets are not found on the teeth. However, Barrett (2000a) was probably correct in regarding them as opportunistic or facultative omnivores rather than strict herbivores (see below).
The cranial characters of many prosauropods clearly match those of herbivorous reptiles rather than those of carnivorous reptiles (Galton 1984a, 1985e, 1986a, contra Swinton 1934 and 1970; Cooper 1981b). The jaw articulation is set below the line of the tooth row, and the degree of ventral offset ranges from small in Anchisaurus, Jingshanosaurus, Massospondylus, Riojasaurus, Thecodontosaurus, Yimenosaurus, and Yunnanosaurus (figs. 12.3A, D, J, M, Q; 12.4A, E, G, J, M) to large in Coloradisaurus, Mussaurus, Plateosaurus, and Lufengosaurus (figs. 12.2A; 12.3G, N). Similarly, ornithischian dinosaurs (Galton 1973a) represent another group of herbivorous dinosaurs that develop a ventral jaw articulation within the group (also in aetosaurs [Charig et al. 1976]). However, whereas nearly all ornithischians have a well-offset jaw articulation, in prosauropods this occurs only in Plateosauridae, so in this respect most prosauropods are less well adapted for herbivory. It is true that transverse movement of the mandibles was not possible in prosauropods, but two functions proposed for the dorsally offset articulation of herbivorous mammals that do not involve transverse movements also apply to the ventrally offset jaw articulation of prosauropods. First, offsetting increases the angle between the lever arm of the bite force and the plane of the teeth, which is important in dealing with resistant plant material (Crompton and Hiiemae 1969). Second, it allows a more even distribution of the biting force by ensuring that the tooth rows are almost parallel at occlusion, so that contact is made along the complete length of the tooth row by a nutcracker-like action (Colbert 1951:89). Crompton and Attridge (1986) reconstructed a predentary for Massospondylus (fig. 12.3J) because the lower jaw is shorter than the upper jaw in two skulls. However, Gowet al. (1990) note that a predentary is unnecessary because the discrepancy results from dorsoventral crushing and is absent in laterally compressed skulls (figs. 12.3M; 12.4E, F).
The coarsely serrated, spatulate crowns of the maxillary and dentary teeth of most prosauropods are transversely compressed and mesiodistally expanded, so that the maximum width of the crown is greater than that of the root (figs. 12.5D, E; 12.7A). This crown shape is nearly identical to that of extant herbivorous iguanids and agamids. Such an expansion, with consequent reduction of the space between adjacent teeth, results in a more nearly continuous cutting edge than occurs in insectivorous and carnivorous lizards (Hotton 1955; Ray 1965; Montanucci 1968; Throckmorton 1976; Auffenberg 1981). The teeth of most prosauropods also resemble those of Iguana iguana in the en echelon arrangement of the maxillary and dentary tooth crowns (fig. 12.5E), which bear prominent serrations set at 45° to the cutting edges (Throckmorton 1976).
The lack of wear facets on the teeth of most prosauropods indicates that the teeth did not come into contact with each other, so there was no tooth-tooth occlusion. Throckmorton (1976) showed that the teeth of Iguana iguana also lack wear facets and that they were used only to bite off a piece of a plant and not for food processing. Unlike lizards, prosauropods probably had cheeks, so food was retained, and an extensive soft secondary palate in Plateosaurus may have allowed breathing and chewing at the same time. Oral processing probably involved a piercing and puncturing action of the teeth in most prosauropods. This also produced the tooth-food wear surfaces on the apices of teeth of Massospondylus and Plateosaurus (fig. 12.7C) and on one edge of a maxillary tooth of “Yunnanosaurus” (fig. 12.5F). The edges of the flat, obliquely inclined second wear surface on the maxillary tooth of “Yunnanosaurus” (fig. 12.5G, H) were self-sharpened by wear against the corresponding surface on the opposing dentary tooth much like that in the sauropod Brachiosaurus (Janensch 1935–36). The continuous tooth-tooth wear surface on the mesial and distal edges of an isolated referred tooth of Plateosaurus (fig. 12.7D; Cuny and Ramboer 1991:fig. 3g; Galton 1998a:fig. 7C) is comparable to those in a variety of sauropods. These wear surfaces could have been caused by propalinal movements, which were widespread in sauropods, but they could also have been produced just by orthal movements (Christiansen 2000; Upchurch and Barrett 2000; Upchurch et al., this vol.), as was the case in prosauropods.
Barrett (1996a, 2000a) pointed out that herbivorous iguanines are not exclusively herbivorous, showing severe growth disorders if meat is excluded from their diet, and are better characterized as opportunist or facultative omnivores rather than strict herbivores. The more rostral part of the tooth row is adapted for the capture of small prey because the teeth are subconical in cross section, with fine serrations on the distal edges, are recurved, and terminate in a sharp point. Consequently, the serrated tooth morphology in the remainder of the tooth row is not an a priori adaptation to herbivory but instead provides the animal with an opportunity to engage in herbivory while continuing to be carnivorous or omnivorous. The striking similarities in the heterodont dentitions of prosauropods and herbivorous iguanines suggest that prosauropods were also opportunistic or facultative omnivores who supplemented their mostly herbivorous diet with small prey or carrion (Barrett 1996a, 2000a). Interestingly enough, a badly worn partial maxilla of the sphenodontid Clevosaurus was found in direct association with numerous worn gastroliths within the ribcage of a larger individual of cf. Ammosaurus sp. (Shubin et al. 1994). The enormous ungual phalanx of the prosauropod manus may even have been used to rip open carrion.
Once the plant food was ingested, a gastric mill provided further mechanical breakdown of the plant (and a little animal) material. Indeed, well-preserved gastric mills consisting of a concentrated mass of small stones have been found associated with the stomach contents of several Massospondylus (Bond 1955; Raath 1974), one Sellosaurus (Huene 1932; Galton 1973b), and one cf. Ammosaurus (Shubin et al. 1994). The occurrence of gastroliths is well documented in sauropods (Christiansen 1996a; Sanders et al. 2001). Modern birds, many of which are herbivorous, lack teeth but use gastroliths for the mechanical breakdown of food (Welty and Baptista 1988).
Because of their relative lack of oral processing compared with most ornithischian dinosaurs and their reliance on mechanical breakdown of food in the gizzard (Christiansen 1999), prosauropods probably had low turnover times in their fermentation chamber for the digestion of poor-quality plant material, although it is possible that they opted for high passage rates and concentrated on the most nutritive components of the food (Farlow 1987a). If so, they probably had long, large intestines and short, small intestines, partitions across the colon, and a symbiotic microflora in a hindgut fermentation chamber to degrade dietary fiber as in modern herbivorous reptiles (Guard 1980; Iverson 1980, 1982; Farlow 1987a). The gastrointestinal situation for prosauropods is unknown, but the barrel-like rib cage (see photos in Huene 1932; Weishampel and Westphal 1986; and Galton 2001b) and broad, apronlike pubis indicate a capacious digestive tract.
Osmólska (1979) pointed out that the external nostrils are enlarged in most herbivorous dinosaurs, including prosauropods such as Plateosaurus. She suggested that the nostrils housed a large lateral nasal gland that functioned as a salt gland to unload the excess potassium ions in the food of large herbivores. However, Whybrow (1981) questioned whether there was sufficient room for the nostrils to house the salt gland and its associated duct system and still perform a respiratory function.
Bakker (1978) suggested that prosauropods could assume a tripodal feeding posture in order to browse at high levels. However, as Coe et al. (1987) noted, elephants can easily stand on their hind legs but do not usually feed in this position. The long neck probably extended the vertical feeding range, so that higher vegetation could be reached, much like the situation in giraffes. Food could not have been conveyed directly to the mouth by the prosauropod manus. However, the manus was probably used to hold on to branches to assist balancing while the animal reared up on its hindlimbs to reach higher levels of vegetation, much as suggested for the large first ungual of sauropods (Upchurch 1994b; Galton, in press). In addition to foliage, prosauropods probably fed on fleshy nilssonialian fruits and possibly on bennettitalian inflorescences at lower levels (0–1 m) and, as the only Triassic browsers capable of feeding at higher levels (1–3 m [Parrish 1998]), on lycopsid fructifications as well (Weishampel 1984b).
According to Sereno (1997:451), “Most variation [in prosauropods] occurs within the skulls.” In fact, postcranial characters do make a significant contribution: of the 79 characters relating to potential phylogenetic signal within Prosauropoda, 45 (i.e., 57%) pertain to the postcrania. The majority (75%) of these postcranial characters concern the appendicular skeleton. Under both delayed and accelerated optimization (PAUP*4.0 [Swofford 1998]), the early phase of prosauropod evolution (as diagnosed by character-state changes near the base of the cladogram) involves both cranial and postcranial modifications. It is noticeable, however, that later character-state changes (e.g., those within Plateosauria) are strongly biased toward cranial modifications alone. This pattern may reflect early dominance of locomotor adaptation, perhaps related to increasing body size.
Neck elongation in basal sauropodomorphs is particularly interesting because it reflects an adaptation to a previously unexploited high-browsing niche (Parrish 1998). Sereno (1997:450) stated that “marked lengthening of the neck does not characterize sauropodomorphs as a whole but instead arose several times within Sauropodomorpha.” Sauropodomorphs lengthened their necks in three main ways (Upchurch 1994a): (1) elongation of cervical vertebrae; (2) addition of new cervical vertebrae; and (3) conversion of dorsal vertebrae to cervical vertebrae. Within Sauropoda, neck elongation always involves lengthening of individual cervicals, often combined with one or both of the other methods of neck elongation. The distribution of neck-related characters in the basal sauropodomorph cladogram indicates that the common ancestor of sauropods and prosauropods would have possessed an additional cervical vertebra (i.e., 10 instead of 9). Cervical elongation, however, has an equivocal distribution: under delayed optimization cervical centra acquire the derived 3:1 length-height ratio independently in more derived sauropods and prosauropods. Under accelerated optimization cervical elongation occurs in the common ancestor of all sauropodomorphs and is reversed in Riojasaurus and Kotasaurus. “Gyposaurus” sinensis also possesses short cervicals, but this does not alter the distribution of this character state because this taxon lies far from the base of the tree. A comparison of neck-trunk ratios (based on the summed lengths of all cervical and dorsal centra) implies that sauropodomorph neck elongation is perhaps more complex than previously considered. Basal ornithischians and theropods have ratios of neck length to trunk length of 0.40 (e.g., Eoraptor [Sereno et al. 1993]). In prosauropods this ratio lies between 0.70 and 0.90 (even in short-necked forms such as Riojasaurus), and in basal sauropods it rises to at least 1.2 (e.g., Shunosaurus [Zhang 1988]). In summary, some limited neck elongation probably occurred prior to the divergence of separate prosauropod and sauropod lineages. Prosauropods and sauropods may have then convergently developed even longer necks, with the latter carrying this trend to a greater extreme than the former.
The best example of sexual dimorphism in a prosauropod probably occurs in the numerous, mostly disarticulated bones of Thecodontosaurus antiquus from the Late Triassic of England, in which there are gracile and rarer robust morphs for the scapula, humerus, femur, and tibia (Galton 1997a; Benton et al. 2000b). Similar differences in scapula, femur, and tibia are found in Melanorosaurus readi (Heerden and Galton 1997; Galton et al., in press). More marked differences are found between the gracile and robust morphs of the theropod dinosaurs Syntarsus rhodesiensis (Early Jurassic, southern Africa [see Chinsamy 1990; Raath 1990]) and Tyrannosaurus rex (Late Cretaceous, western United States [see Carpenter 1990a; Larson 1994), in which several lines of evidence indicate that the robust form represents the female (see also Chapman et al. 1997; Galton 1999b). A morphometric analysis by Weishampel and Chapman (1990) of the femora of Plateosaurus longiceps from the quarry at Trossingen indicates the presence of one species with two cluster groups that were interpreted as individual variation, probably sexual (Galton 1997a). Gow et al. (1990) suggested that a Trossingen skull of P. longiceps (fig. 12.2A) and one of Massospondylus carinatus (fig. 12.5E) with a thicker dorsal orbital rim and a deeper maxilla caudally represent males and that the more lightly built skulls represent females (fig. 12.5F). However, these differences in the skull of P. longiceps, which do not correlate with the femoral differences, probably represent individual variations (Galton 1997a, 2001b), as do variations in the size of the chevron bone between caudal centra 1 and 2 and the sacral differences in Sellosaurus gracilis (Galton 2001c).
Eggs are reported for Mussaurus (Late Triassic [Bonaparte and Vince 1979]) and Massospondylus (Early Jurassic [Kitching 1979]). Under the SEM the shell structure of the latter is similar to those of birds and crocodiles but distinctly unlike the rigid eggshells of chelonians and squamates (Grine and Kitching 1987). The palisade layer of the prosauropod shell consists of broadly wedge-shaped but ill-delimited units, resembling crocodilian shells in this respect, but the prosauropod shell lacks the distinct horizontal laminae of crocodilians. The configuration of the calcite crystals of the prosauropod shell is tabular rather than spherulitic, and the packing and cleavage of the crystals results in an apparent herringbone pattern as in avian and dinosaur eggs. The inner surface has an undulating contour but lacks a mammillary layer; because it was probably at least partly decalcified during incubation, the mammillary layer was not preserved in the fossil (Grine and Kitching 1987). These eggs were reidentified as crocodilian by Carpenter (1999:30), but this is incorrect (Sues, pers. comm.).
The role in the evolution of dinosaurs of heterochrony, or changes in the timing or rate of developmental events relative to the ancestral condition, is discussed by Weishampel and Horner (1994) and Long and McNamara (1997). Prosauropods, like other dinosaurian groups for which there are ontogenetic samples (see Weishampel and Horner 1994), possessed fibro-lamellar primary bone and often dense Haversian secondary bone through-out their lives (Ricqlès 1980; Chinsamy 1993), so they appear to have been fast-growing animals. The clutch size is apomorphically reduced in prosauropods, as indicated by nest material with at least two eggs for Mussaurus patagonicus (Bonaparte and Vince 1979) and at least six eggs for Massospondylus carinatus (Kitching 1979). In both cases the hatchlings are small (weight ca. 1 kg, length ca. 0.2%–0.4% of adult length), so there is an apomorphic increase in adult size relative to hatchlings (less than 500 to 1,000 times larger than hatchlings [Weishampel and Horner 1994]).
The midshaft cross-sectional histology of femora of Massospondylus carinatus indicates that the physiology was at an intermediate level on the continuum from ectothermy to endothermy (Chinsamy 1993). The cortex is stratified into growth rings, so growth was seasonally cyclic, with one zone and one annulus representing an annual cycle. Slowed growth is indicated by the poorly vascularized, lamellated annuli, whereas fast growth is represented by the highly vascular zonal bone tissue. The presence of highly vascularized fibro-lamellar tissue suggests that the bone deposition rate during the favorable growth season was more similar to that of living endotherms. However, as in living ectotherms, growth was indeterminate because it continued indefinitely, although at a decreasing rate, with the largest femur representing an individual 15 years old (Chinsamy 1993). Long and McNamara (1997) note that if the onset of maturity is delayed (hypermorphosis), the extrapolation of rapid juvenile growth trajectories will result in attainment of a larger body size and more “developed” adults (peramorphosis; it can also occur by an increase in the growth rate [acceleration] or an earlier onset of growth [predisplacement]). They compare a juvenile skull of Mussaurus with that of an adult Plateosaurus (figs. 12.2A, 12.3I; see Carpenter et al. 1994a:fig. 24.1C, D, for comparison with adult Mussaurus maxilla and dentary) and note several differences. They note that the trends in the skulls of sauropods are “indicative of peramorphic development of the skull beginning with moderate ontogenetic change in prosauropods and extending to increased ontogenetic development throughout the lineages of camarasaurs, brachiosaurs, diplodocids, and nemegtosaurids”; they also attribute to peramorphosis the massive increase in size in sauropodomorphs from the Late Triassic to the Late Jurassic (Long and McNamara 1997:120, fig. 10).
Among prosauropods, Saturnalia and Thecodontosaurus are considered fully bipedal. The forelimbs are half the length of the hindlimbs as in other prosauropods, but the hindlimbs are proportionally much longer (fig. 12.1A). The ratio of trunk length to hindlimb length is 1:1.6 in Thecodontosaurus, compared with a ratio of 1:1.22 to 1:1.9 in other fully bipedal dinosaurs (Galton 1970b). The remaining prosauropods were probably only facultatively bipedal (ratios of trunk length to hindlimb length ranging from 1:0.95 to 1:1.15, not including the additional leverage from the proportionally longer neck). However, Riojasaurus (and other melanorosaurids) must have been fully quadrupedal (ratio of 1:0.71, compared with ratios of 1:0.69 to 1:0.9 in undoubtedly quadrupedal dinosaurs (Galton 1970b). The increase in the proportional length of the trunk provided storage space for the elongate viscera necessary for a herbivorous diet. On the basis of an analysis of the bending moments and compressive forces on the vertebral column, Christian and Preuschoft (1996: 808) stated that Plateosaurus “appears to have been a habitual quadruped, which may have used a bipedal gait infrequently at its fastest locomotion, or during standing.”
In prosauropods with sigmoid femora (fig. 12.9C–E) the hindlimb was held more upright than in sprawlers, but the femur was probably at an angle of 20° to the parasagittal plane, with the head pointing craniomedially, so the parasagittal tibia could be moved craniocaudally (Cooper 1981b; Heerden 1979 suggested an angle of 30°–45°). Cooper (1981b) suggested that during retraction the femur was twisted such that the head faced medially and perforated the open acetabulum. Christian et al. (1996) concluded that the limb posture of Plateosaurus during quadrupedal locomotion was more extended than in modern lizards, especially in the only slightly sprawling hindlimbs. However, as in lizards the adducting muscles of the pectoral and pelvic girdles were used for balancing the hip and shoulder joints, and there was at least moderate lateral undulation of the vertebral column. The femur was probably closer to parasagittal in Anchisauria, “Gyposaurus” sinensis, Jingshanosaurus, Lufengosaurus, and Yunnanosaurus, in which the head is more medially set and the bone is distally straighter in cranial and caudal views (fig. 12.10D, F). However, even in mammals a fully erect limb posture is found only in cursorial and graviportal mammals, and the femur is found at an angle of 20°–50° to the parasagittal plane in many other mammals (Jenkins 1971).
The locomotor capabilities of dinosaurs can be estimated by comparing plots of the hindlimb proportions (tibia:femur/metatarsal III:tibia) with those of living mammals (see Coombs 1978a). On this basis, small- to medium-sized prosauropods such as Ammosaurus, Anchisaurus, Massospondylus, Saturnalia, Sellosaurus, and Thecodontosaurus were low-grade subcursorial runners, and large prosauropods such as Jingshanosaurus, Lufengosaurus, Melanorosaurus, Plateosaurus, and Yunnanosaurus were mediportal. Consequently, prosauropods were probably the slowest of the bipedal dinosaurs but better runners than most other quadrupedal dinosaurs (Coombs 1978a; Carrano 1998).
The constancy of the structure of the prosauropod manus correlates well with the development of an enormous, trenchant first ungual that was used for offense or defense while the animal was in a bipedal stance (Galton 1971a, 1971b, 1990a, 2001a, in press). With the first digit in full extension, the ungual would clearly have been a formidable weapon (fig. 12.8H, L–O). During quadrupedal locomotion, with the digits of the manus in full extension, the weight of the forequarters was taken by digits II to IV (in particular subequal digits II and III), while the first ungual was held clear of the ground (fig. 12.8H). Only if the substrate were irregular or soft would the lateral surface of this phalanx have touched the ground, and even then the point of the claw would not have been damaged. Such contact is shown by the trackway of Navahopus falapollex that Baird (1980) thought was made by a prosauropod walking quadrupedally. The first ungual was usually held clear off the ground as a result of two specializations of the first digit (fig. 12.8I–K). First, the ginglymus of metacarpal I extends well above the dorsum of the shaft and the articular surface extends proximally onto the dorsal surface, so that the first phalanx was capable of marked hyperextension. Second, the first phalanx is twisted along its length (45° between the rotational axes). In full extension the ungual phalanx is directed ventromedially. During flexion the first digit of the prosauropod manus rotates laterally around its long axis, so that the first ungual comes to lie more or less parallel with the unguals of digits II and III. This rotation results from the asymmetrical structure of the ginglymus of metacarpal I, whose lateral condyle is larger than the medial condyle (fig. 12.8J). When flexed, the prosauropod manus must have acted as an efficient grasping organ, particularly during bilateral use of the hands. The main variation in the structure of the prosauropod manus is that metacarpal I is more slender in Thecodontosaurus and metacarpals II–IV are more slender in Thecodontosaurus and Anchisaurus, in which the metatarsals are also more slender. Most prosauropods were large, and the more massive nature of the metacarpals was a weight-supporting adaptation that was unnecessary in the smaller forms. The basic adaptations of the first digit are present in the habitual biped Thecodontosaurus (figs. 12.1A, 12.8O), a basal prosauropod, so they were present prior to the group's becoming habitual quadrupeds. The retention of the same basic plan for the manus for at least 30 million years shows that it met the basic functions of weight support while walking quadrupedally and defense or offense while bipedal, with or without the propping action of the tail.