TWENTY-TWO

Basal Ceratopsia
Ceratopsia consists of Psittacosauridae and Neoceratopsia, the latter formed by numerous basal taxa and Ceratopsidae. Consequently, this chapter on basal ceratopsians includes psittacosaurids and nonceratopsid neoceratopsians. Psittacosauridae is a monogeneric (Psittacosaurus) clade consisting of 10 species, while basal Neoceratopsia is formed by 11 genera, with 12 species of basal Neoceratopsia being recognized (table 22.1). Psittacosaurids are known from the Early Cretaceous of Asia, whereas basal neoceratopsians come from the latest Jurassic (Chaoyangsaurus youngi, Zhao et al. 1999; Swisher et al. 2002) to the latest Cretaceous in Asia and North America. Basal ceratopsians are small (1–3 m long), bipedal or quadrupedal herbivores (figs. 22.1, 22.2). Several taxa are extremely abundant and are represented by growth series from hatchlings to adults. Sexual dimorphism in Protoceratops is well supported (Dodson 1976; Lambert et al. 2001; Tereshchenko 2001). Basal neoceratopsians evolved larger skulls relative to their postcranial skeletons and more developed frills than psittacosaurids.
Definition and Diagnosis
Ceratopsia is defined as all Marginocephalia closer to Triceratops than to Pachycephalosaurus. Autapomorphies of this clade include a high external naris separated from the ventral border of the premaxilla by a flat area, a rostral bone, an enlarged premaxilla, well-developed lateral flaring of the jugal, wide dorsoventral length of the infraorbital ramus of the jugal, and contact of palatal extensions of the maxillae rostral to the choana.
Anatomy
The description below is based mainly on Psittacosaurus mongoliensis (Sereno 1990b), Archaeoceratops oshimai (Dong and Azuma 1997; You 2002; You and Dodson 2003), Protoceratops andrewsi (Brown and Schlaikjer 1940b; Dodson and Currie 1990), and Leptoceratops gracilis (Sternberg 1951; Dodson and Currie 1990), as they include the best preserved and described specimens representing the major subgroups among basal ceratopsians. Additional comments on other taxa are also included and are discussed further in the “Systematics and Evolution” section below.
Skull and Mandible
The skull of basal ceratopsians (figs. 22.3–22.5) is pentangular in dorsal view, with a narrow beak, a strong laterally flaring jugal, and a caudally extended frill. The beak is round in psittacosaurids and pointed in basal neoceratopsians. The jugal horn is more pronounced in psittacosaurids than in basal neoceratopsians. The frill is incipient in psittacosaurids and small but variably developed in basal neoceratopsians. The preorbital portion of the skull is dorsoventrally deep, especially in psittacosaurids, and rostrocaudally short in both psittacosaurids and the most basal members of neoceratopsians such as Archaeoceratops.
The external naris is highly positioned, especially in psittacosaurids, bounded by the premaxilla ventrally and the nasal dorsally. Its shape is subrounded in psittacosaurids and Leptoceratops, but elliptical in Archaeoceratops and Protoceratops. The antorbital fossa or fenestra is not present in psittacosaurids, but a small opening is enclosed between the premaxilla and the lacrimal. In basal neoceratopsians, the shape of the antorbital fossa varies from subtriangular (Archaeoceratops) to elliptical (Protoceratops and Leptoceratops), usually with a small fenestra on the caudodorsal part of the wall. An additional antorbital fenestra is present in Bagaceratops between the premaxilla and the maxilla. The orbit is smaller than the infratemporal fenestra in psittacosaurids and the two are subequal in size in most neoceratopsians, but the orbit is larger than the infratemporal fenestra in Archaeoceratops.
The rostral in psittacosaurids is thin in sagittal section and forms a convex shield that caps a triangular surface on the conjoined premaxillae. Its rostroventral end is round in dorsal view and does not curve ventrally to form a pointed beak. The rostral also contacts the slender rostroventral processes of the nasals. In basal neoceratopsians, the rostral is strongly compressed transversely and extends rostrally beyond the rostral tip of the lower jaw. A sutural contact between the rostral and nasal is absent. In Archaeoceratops and Protoceratops, the ventral edge of the rostral curves strongly rostroventrally to a point, while in Leptoceratops, the ventral edge is horizontal with a well-developed caudolateral process as long as it is high.
The premaxilla is a characteristic element in psittacosaurids. The tall, parrotlike face of psittacosaurids is formed almost entirely from the expansive caudolateral process of the premaxilla, which contacts the lacrimal and the prefrontal caudally. In palatal view, the palatal process of the premaxilla arches from the lateral margin of the beak to the midline. The caudal extension of the palatal process of the premaxilla does not reach the rostral margin of the choana and is separated from the vomer by the palatal extension of the maxillae in between. In basal neoceratopsians, the premaxilla is not as large as in psittacosaurids and is bounded by the rostral rostrally, the nasal dorsally, and the maxilla caudally. In Archaeoceratops, the premaxilla is nearly square in lateral view, but higher than long in Protoceratops and longer than high in Leptoceratops. The caudolateral process of the premaxilla is not developed in Archaeoceratops, but is prominent in both Protoceratops and Leptoceratops. Unlike the flat ventral edge of Archaeoceratops and Protoceratops, the premaxilla is ventrally convex in Leptoceratops. In Bagaceratops, the caudal edge of the premaxilla surrounds an additional antorbital fenestra together with the maxilla caudally.
TABLE 22.1
Basal Ceratopsia

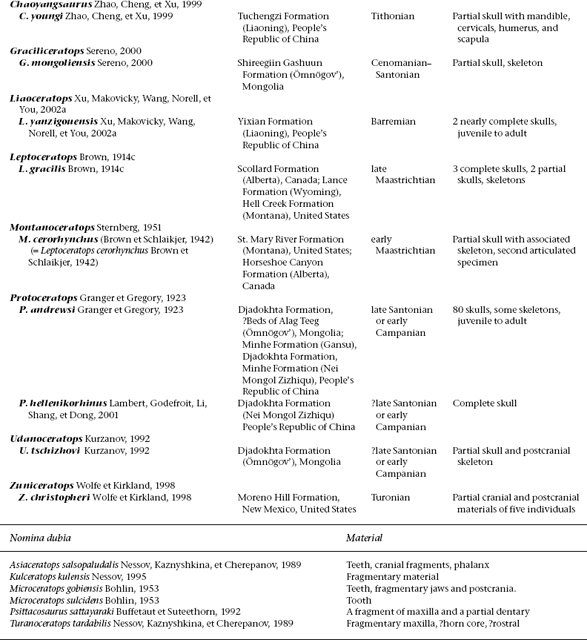
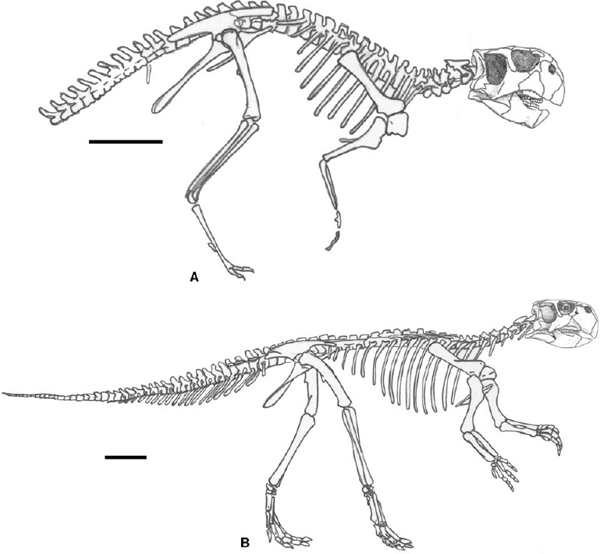
FIGURE 22.1. Psittacosaurus skeletons in right lateral view: A, Psittacosaurus neimongoliensis; B, Psittacosaurus mongoliensis. Scale = 10 cm. (A after Russell and Zhao 1996; B after Osborn 1924c.)
In psittacosaurids, the maxilla is triangular in lateral view and situated with its caudal half underneath the orbit. It is largely bounded by the premaxilla rostrally and the jugal caudally, has a small contact with the lacrimal dorsally, and does not reach the nasal. In basal neoceratopsians, the maxilla is tall and forms about two-thirds of the height of the face. It sutures with the premaxilla rostrally and the lacrimal and the jugal caudally, and it has a small contact with the nasal dorsally. The premaxilla-maxilla suture is vertical in Archaeoceratops, but inclined in both Protoceratops and Leptoceratops. A prominent elliptical antorbital fossa with a small antorbital fenestra exists in most basal neoceratopsians, except in Archaeoceratops, in which the antorbital fossa is triangular and the antorbital fenestra is not evident. In Protoceratops and Bagaceratops, there is a prominent maxillary sinus that communicates with the antorbital fossa but not the nasal cavity (Osmólska 1986).
The nasal of psittacosaurids is unusual in that a slender rostral process extends ventral to the external naris and reaches the rostral. In basal neoceratopsians, the nasal is long and narrow, but never extends beyond the rostral end of the external naris to contact the rostral bone. No horn-core is present in Archaeoceratops and Leptoceratops (You, pers. obs.). An incipient nasal horn core is evident in Protoceratops, Bagaceratops (Marya ska and Osmólska 1975), and Udanoceratops (Kurzanov 1992), which is located caudodorsal to the external naris.
ska and Osmólska 1975), and Udanoceratops (Kurzanov 1992), which is located caudodorsal to the external naris.
The lacrimal of psittacosaurids is bounded by the prefrontal dorsally, the premaxilla rostrally, the maxilla ventrally, and the jugal caudoventrally. The lateral wall of the lacrimal canal remains only partially ossified, and the canal opens externally in a small pore about halfway along its passage from the margin of the orbit to the nasal cavity, which is bounded rostrally by the premaxilla. In basal neoceratopsians, the lacrimal is largely bounded by the prefrontal dorsally and the maxilla rostroventrally, and has a small contact with the nasal rostrally and the jugal caudally. Unlike in psittacosaurids, premaxilla-lacrimal contact is prevented because the expanded maxilla contacts the nasal dorsally. The rostroventral corner of the lacrimal usually contributes a small portion to the caudodorsal wall of the antorbital fossa. The lacrimal of Leptoceratops is larger than those of other basal neoceratopsians.
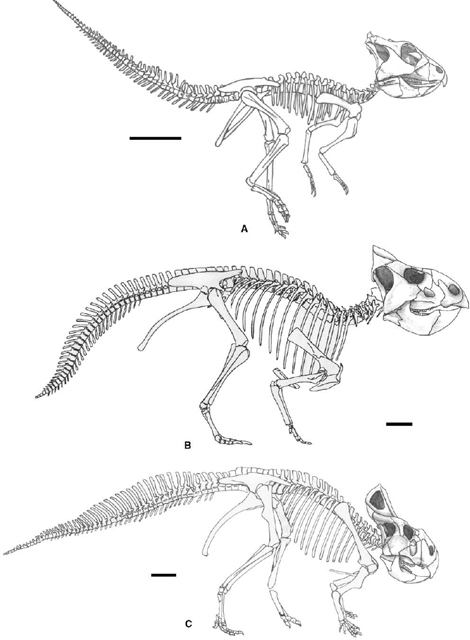
FIGURE 22.2. Skeletons of basal Neoceratopsia in right lateral view: A, Archaeoceratops oshimai; B, Leptoceratops gracilis; C, Protoceratops andrewsi. Scale = 10 cm. (A after Dong and Azuma 1997; B after Russell 1970; C after Granger and Gregory 1923.)
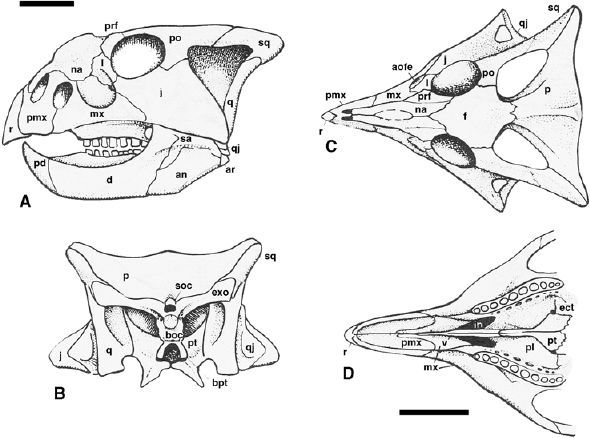
FIGURE 22.3. Skull of Bagaceratops rozhdestvenskyi: A, lateral; B, occipital; C, dorsal; and D, palatal views. Scale = 5 cm. (After Maryaska and Osmólska 1975.)
The prefrontal borders the rostrodorsal rim of the orbit in both psittacosaurids and basal neoceratopsians. It is surrounded by the frontal, the nasal, the premaxilla, and the lacrimal in psittacosaurids, whereas the premaxilla is excluded in basal neoceratopsians.
The palpebral usually attaches to the caudal rim of the prefrontal. In psittacosaurids, it is a short, dorsally arched rod associated solely with the prefrontal. In Archaeoceratops, the prominent palpebral is triangular with a caudally pointed end, and it articulates with both the prefrontal and the lacrimal.
In both psittacosaurids and basal neoceratopsians, the dorsoventral length of the jugal below the orbit is at least as long as that underneath the infratemporal fenestra. Although the flaring of the jugal characterizes both psittacosaurids and basal neoceratopsians, it shows different configurations in these two groups. In psittacosaurids, the breadth of the skull across the flaring jugal horns can be as long as or longer than the skull length, and the flaring projects from the midsections of the jugals; in basal neoceratopsians, the width of the skull across the flaring jugal horns never exceeds the basal skull length, and the flaring is usually directed caudolaterally from the caudal end of the jugal. The postorbital process of the jugal is short in psittacosaurids, but long and stout in basal neoceratopsians. In Protoceratops and Leptoceratops, there is an incipient jugal-squamosal contact around the rostrodorsal rim of the infratemporal fenestra. The caudolateral end of the jugal is often thickened in basal neoceratopsians, and this thickening is usually accentuated by an epidermal ossification, the epijugal, in Protoceratops and Leptoceratops, but not in Archaeoceratops.
The large quadratojugal of psittacosaurids is located on the caudoventral corner of the skull. The rostral portion passes medial to the jugal, and the caudal portion covers the ventral half of the quadrate shaft in lateral view. In basal neoceratopsians, the quadratojugal is largely excluded from lateral view by the caudal extension of the jugal. It is a transversely thin element inserted between the jugal laterally and the quadrate medially. In Archaeoceratops, a trace of the quadratojugal is still visible laterally at the caudoventral end of the skull.
The postorbital in psittacosaurids is a restricted element, with two rodlike, elongated processes, the jugal process and the squamosal process. In basal neoceratopsians, the jugal process is shortened and the squamosal process is stout. The enlargement of the jugal and the reduction of the infratemporal fenestra exclude contact between the postorbital and the infratemporal fenestra in Protoceratops and Leptoceratops.
In basal ceratopsians, the paired frontals form a major portion of the cranium, border the orbit laterally, and constitute the rostral limit of the supratemporal fenestrae. In Psittacosaurus, the dorsal surface of the frontal is restricted to the flat interorbital portion of the skull roof. In Protoceratops and Leptoceratops, a pair of modest frontoparietal depressions in adult specimens is associated with the rostral borders of the supratemporal fenestrae, reflecting expansion of the attachments of the jaw adductor musculature.
In psittacosaurids, the parietal roofs the braincase and forms the medial border of the supratemporal fenestra. It extends caudally over the occiput as a transversely broad shelf. In basal neoceratopsians, an incipient parietosquamosal frill extends behind the skull, which is simple and lacks the various decorations seen in ceratopsids. In Archaeoceratops, the frill is short, as indicated by the short squamosal, while in Leptoceratops it is short and unfenestrated. The frill of Protoceratops is moderately developed and fan-shaped, tilting caudodorsally with a pair of parietal fenestrae near the caudal end. A median keel develops on the dorsal surface in both Protoceratops and Leptoceratops.
In psittacosaurids, the squamosal forms a simple bar with the postorbital that separates the infra- and supratemporal fenestrae and provides a cotylus for the head of the quadrate. A postquadrate extension of the squamosal developed in basal neoceratopsians. In Protoceratops, it runs caudodorsally along the ventral margin of the parietal, with which it forms the lateral edge of the frill. In Leptoceratops, the postquadrate extension is not well developed, but extends ventrally to hook the head of the quadrate caudally.

FIGURE 22.4. Skulls of Psittacosaurus. A, B, Psittacosaurus meileyingensis in lateral, A, and dorsal, B, views; C, D, Psittacosaurus mongoliensis in C, lateral, and D, dorsal views; E, F, Psittacosaurus neimongoliensis in E, lateral, and F, dorsal views; G, H, Psittacosaurus sinensis in G, lateral and H, dorsal views. Scale = 2 cm (A–F), 3 cm (G, H). (A–D, G, H after Sereno 1990b; E, F after Russell and Zhao 1996.)
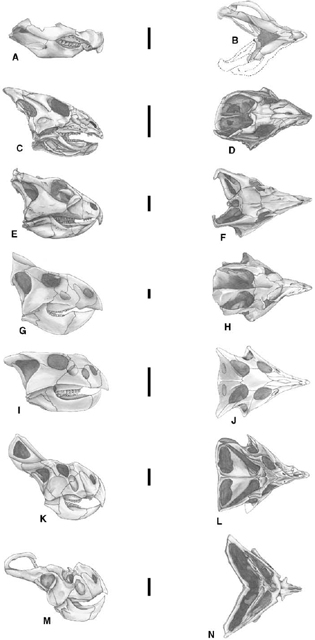
FIGURE 22.5. Skulls of basal Neoceratopsia. A, B, Chaoyangsaurus youngi in A, lateral, and B, dorsal views; C, D, Liaoceratops yanzigouensis in C, lateral, and D, dorsal views; E, F, Archaeoceratops oshimai in E, lateral, and F, dorsal views; G, H, Leptoceratops gracilis in G, G, lateral, and H, dorsal views; I, J, Bagaceratops rozhdestvenskyi in I, lateral, and J, dorsal views; K, L, Protoceratops andrewsi in K, lateral, and L, dorsal views; M, N, Protoceratops hellenikorhinus in M, lateral, and N, dorsal views. Scale = 3 cm (A–D), 4 cm (E, F), 5 cm (G, H), 8 cm (I, J), 10 cm (K-N) (A, B after Zhao et al. 1999; C, D after Xu et al. 2002a; E, F after Dong and Azuma 1997; G, H after Sternberg 1951; I, J, after Maryaska and Osmólska 1975; K, L after Brown and Schlaikjer 1940b; M, N, after Lambert et al. 2001.)
In psittacosaurids, the ventral half of the quadrate is erect and largely covered by the quadratojugal laterally. The dorsal half bends caudodorsally to contact the squamosal. In basal neoceratopsians, there is a progressive reorientation of the cheek. The ventral end of the quadrate is rotated forward, the infratemporal fenestra is compressed, and the jugal, quadratojugal, and ventral quadrate are telescoped to lie side by side rather than in series rostral to caudal (Dodson 1993).
In ventral view of the skull of psittacosaurids, a transversely arched secondary palate is present rostrally, formed principally by the premaxillae. Caudal to the choana, the remainder of the palate is composed of the palatine, pterygoid, and ectopterygoid. The vomers, which fuse rostrally, arch in the midline from the secondary palate rostrally to the palatine and pterygoid caudally. The suborbital opening persists as a foramen between the palatal bones and the maxilla. An elongate flange of the pterygoid, the mandibular ramus, is directed caudoventrally toward the adductor fossa of the lower jaw.
The palate of basal neoceratopsians is strongly vaulted both transversely and, in Bagaceratops at least, longitudinally as well. The choana is positioned far forward and oblique to the palatal plane due to the narrowness and vaulting of the snout (Osmólska 1986). The secondary palate is short. The vomer is a straight median bar running between the pterygoids caudally and the palatal processes of the maxillae rostrally. It rises steeply caudodorsally to meet the rostrodorsally inclined longitudinal process of the palatine. This orientation divides the nasal cavity into narrow, paired channels (Osmólska 1986). The intervomerine suture remains prominent in Protoceratops and Bagaceratops. In Bagaceratops and Leptoceratops, there is a palatine foramen situated between the pterygoid, ectopterygoid, and palatine. Expansion of the palatine is accompanied by restriction of the ectopterygoid to a small, flattened bone on the dorsum of the caudal process of the maxilla. Contact of the ectopterygoid with the palatine has been lost in Leptoceratops. The pterygoid is a complex bone that retains its primitive role as a link between the quadrate and palate, with a brace on the braincase. In Protoceratops, a concavity passes across the ventral surface of the pterygoid adjacent to the basipterygoid articulation and onto the quadrate ramus.
In psittacosaurids, the exoccipital and opisthotic are completely fused. The combined element borders the foramen magnum and sends a narrow paroccipital process laterally and slightly caudally. The foramen magnum is bounded by the supraoccipital dorsally and the basioccipital ventrally. The plane of the occiput is nearly vertical.
In basal neoceratopsians, the occiput is broad, and the exoccipital forms a long, slender, straplike process that extends laterally to reach the squamosal, usually contributing to the support of the dorsal end of the quadrate. The exoccipitals do not roof the foramen magnum and make only a slight to moderate contribution to the occipital condyle. The supraoccipital forms the dorsal border of the foramen magnum, with a rostral extension that contacts the prootic and the laterosphenoid. The basioccipital is excluded from the foramen magnum by the exoccipitals. The large size and spherical form of the condyle with its distinct neck is diagnostic of neoceratopsians. The caudal face of the prominent basal tubera is formed by the basioccipital, and there is a simple butt joint with the basisphenoid rostrally. The opisthotic, prootic, and laterosphenoid retain their usual morphology and contact with adjacent bones. The orbitosphenoid is ossified in Bagaceratops, although it is rarely separate from the laterosphenoid due to early fusion of these elements. A presphenoid is probably present as well. The parasphenoid is indistinguishable from the basisphenoid. The basicranial complex is pierced on the midline by the auditory opening in neoceratopsians, except in Bagaceratops, in which the opening passes between the basioccipital and basisphenoid.
The lower jaw of psittacosaurids is deep in lateral view. The predentary is subtriangular in lateral and ventral views, with short, weak lateral processes and a broad ventromedian process. The dentary extends caudally from the symphysis to the top of a moderately sized coronoid process. In Psittacosaurus mongoliensis and P. meileyingensis, a low flange hangs ventrolaterally from the ventral margin of the dentary. The external mandibular fenestra is open, except in P. sinensis, and enclosed by the dentary, angular, and surangular. The articular surface on the lower jaw for the mandibular condyle of the quadrate is flat. The flat ventral surface of the jaw articulation may have facilitated rostrocaudal sliding of the quadrate condyles during mastication.
In basal neoceratopsians, the predentary is short and terminates sharply, tilting rostrodorsally. The dentary is deep and the ventral border is curved in Protoceratops, Leptoceratops, and Udanoceratops, but is straight in Archaeoceratops and Bagaceratops. A strong lateral ridge runs the length of the dentary and is confluent with the coronoid process; thus, the tooth row is strongly inset and passes medial to the coronoid process. The dentary is thickest at the base of the coronoid process, where it surrounds the mandibular fossa in which M. intramandibularis is situated. The fossa is covered ventrally by an extensive splenial. The surangular is tall, contributing half to the coronoid. The angular is larger in Archaeoceratops than in both Protoceratops and Leptoceratops.
Premaxillary teeth do not occur in psittacosaurids. Denticulate maxillary and dentary crowns are positioned along the tooth rows with a small amount of overlap along the crown edges. Both upper and lower tooth rows are inset from the lateral surface of the skull by a buccal emargination. Maxillary and dentary teeth are approximately equal in size and number in any single individual. Tooth count increases during growth from 5 to 12 in Psittacosaurus mongoliensis. Adult tooth count varies from 8 to 12 among psittacosaurid species. In all species of Psittacosaurus, the buccal surface of the maxillary crown is flatter than the lingual surface. The buccal surface of dentary teeth is ornamented by a weak primary ridge that terminates at the apex of the crown and is flanked by a few weak secondary ridges. The lingual surface of the dentary crowns, in contrast, is dominated by a prominent median primary ridge. The primary ridge is bulbous near the crown base but tapers toward the apex of the crown and is flanked on either side by flat crescentic surfaces with weak secondary ridges. All crown surfaces are covered with enamel, which is several times thicker on the buccal surface of the maxillary crowns and the lingual surface of the dentary crowns than on the opposing sides. The psittacosaurid dentition is characterized by broad planar wear surfaces with self-sharpening cutting edges. The wear surfaces of a single tooth row lie in approximately the same oblique plane (dorsomedial-ventrolateral) but do not form a continuous wear surface along the tooth row. Some crowns are truncated by a single wear surface, while others are truncated by two contiguous wear facets. A precise pattern of occlusion between the upper and lower tooth rows, such as alternate or tooth-to-tooth alignment, does not occur in psittacosaurids.
Premaxillary teeth are present in Archaeoceratops, Protoceratops, and Bagaceratops in the form of simple cylindrical pegs. The crowns of both maxillary and dentary teeth of basal neoceratopsians are leaf-shaped, enameled on one side (buccally and lingually, respectively), and bear a strong median ridge, usually but not always asymmetrically placed (distal and mesial on maxillary and dentary teeth, respectively). Secondary ridges are prominent. The crown is set at a high angle to the root, resulting in an occlusal plane that is steep (greater than 60°). In Leptoceratops, the dentition uniquely combines vertical shear with a horizontal shelf on the dentary teeth, such that the functions of shear and crushing are combined (Sternberg 1951; Ostrom 1966). Tooth replacement is primitive; there is a functional tooth with a single replacement tooth at each position. The number of maxillary tooth positions increases ontogenetically from 8 to 15 in Protoceratops. The total number of tooth positions ranges from 10 in Bagaceratops, to 15 in Protoceratops, to 17 in Leptoceratops.
The slender stapes is preserved in two individuals of psittacosaurids. The length of the bony stapes equals the distance from the fenestra ovalis to the otic notch, which is formed rostrally by the quadrate shaft and caudally by the paroccipital process and caudal process of the squamosal. Gently curving, rod-shaped first ceratobranchials are preserved in Psittacosaurus mongoliensis and P. sinensis.
Hyoid bones have been described for Protoceratops andrewsi (Colbert 1945), and Leptoceratops gracilis (Sternberg 1951). In P. andrewsi, they have a flattened, bean shape; while in L. gracilis, they take the form of simple slender rods.
Sclerotic rings are found in psittacosaurids, but rarely preserved in basal neoceratopsians. A partial ring in Protoceratops, which contains 12 complete and 3 partial overlapping plates, was described by Brown and Schlaikjer (1940b).
Postcranial Skeleton
AXIAL SKELETON
The principal divisions of the axial skeleton (fig. 22.1) appear to be uniform across all species of Psittacosaurus (Sereno 1990b). There are 21 presacrals, 6 sacrals, and approximately 45 caudals. In the presacral division, there may be as many as 8 or 9 cervical vertebrae in P. mongoliensis and P. sinensis. Distinction between cervical and dorsal vertebrae, in this case, is based on the position of the parapophysis. In dorsal vertebrae, the parapophysis is located above the neurocentral suture.
Few neoceratopsians are known from complete vertebral columns. Leptoceratops is so known, while Protoceratops, known only to the level of caudal 32, has been estimated as having a complete caudal series of greater than 40 (fig. 22.2; Brown and Schlaikjer 1940b). There is more certainty about the presacral and sacral portions of the column. The basal neoceratopsian vertebral formula usually consists of 10 cervicals, 12 dorsals, and 6–8 sacrals.
A proatlas has not been observed in Psittacosaurus. The atlas is composed of four discrete elements: a lozenge-shaped centrum, a U-shaped intercentrum, and two neural arches. The axial centrum, longest in the cervical series, is fused rostrally to a small, wedge-shaped axial intercentrum in the adult. The axial neural spine is well developed, extending caudodorsally at approximately 45°. The centra of the postaxial cervicals are transversely pinched and bear low ventral keels. The change in shape of the postaxial centra from a trapezoid caudally to a parallelogram cranially indicates that the neck assumed a gentle S-shaped curve in natural articulation.
Coalescence of the centra and neural arches of the first cervicals to form a syncervical (Ostrom and Wellnhofer 1986) or cervical bar (Langston 1975) characterizes all neoceratopsians (fig. 22.6A). However, in Archaeoceratops the condition is not clear, and in Leptoceratops, the atlas is small, bearing no neural spine, and the first three cervicals co-ossify only when mature adult size is achieved. The neoceratopsian syncervical generally incorporates a fused atlas-axis complex as well as cervicals 3 and 4. Cervicals 5 to 10 in basal neoceratopsians are subequal in length, and the neural spines are reduced. Adventitious fusion of cervicals 5 and 6 is reported in Protoceratops. Double-headed ribs are borne on all centra beginning with cervical 3. The tuberculum is prominent, and the shaft is short and straight. Elongation of the ribs usually begins with the sixth.
As in the cervical centra, the articular surfaces in the dorsal centra of psittacosaurids are flat cranially and gently concave caudally. The dorsolateral angle of the transverse processes increases from approximately 35° in the cranial dorsals to approximately 40° in the mid-dorsals, before decreasing to the horizontal in the caudal dorsals. Freely articulating ribs occur in the presacral series. The long atlantal rib, like the two or three caudalmost dorsal ribs, is single-headed. The double-headed ribs of the axis and several of the cervicals that follow are short, extending only to the middle of the next centrum. The ribs of the caudal cervicals and cranial dorsals increase rapidly in length to a maximum between the fourth and seventh dorsals. Passing caudally from the first dorsal rib, the capitulum decreases in length as the parapophysis climbs the cranial edge of the transverse process. The last few dorsal ribs are short and rod-shaped.
There are usually 12 long, low dorsals in basal neoceratopsians. The centra faces are either round or pear-shaped. The neural canal decreases in diameter caudally down the dorsal series. Transverse processes are strongly elevated by dorsal 3 but decrease in prominence in the caudal half of the series, the region in which the zygapophyses increase in prominence. Neural spines are robust and generally show little variation in height. The tallest dorsal spine varies in position from dorsal 3 (Leptoceratops), to dorsal 9 (Protoceratops), and back to dorsal 12 (Montanoceratops, Brown and Schlaikjer 1942). The cranial dorsal ribs are comparatively straight with a prominent tuberculum widely separated from the capitulum. In the mid-dorsal region, the ribs are more curved and the tuberculum less prominent. Caudally, the tuberculum is weak and closer to the rib head. The second to sixth dorsal ribs are subequal in length, after which there is a variable shortening caudally. The twelfth rib may be weak. Bundles of ossified epaxial tendons have been noted at the bases of the neural arches in the dorsal and sacral regions of Leptoceratops.
There are six sacrals in psittacosaurids. The spool-shaped sacral centra are significantly longer than the adjacent dorsal and caudal centra. In adults, the sacral centra co-ossify but remain separate from the last dorsal and first caudal. The first sacral centrum is particularly robust. Sacral neural arches are characterized by nearly vertically oriented zygapophyses and neural spines that remain separate in the adult. Ossified epaxial tendons are present along nearly the entire dorsal and sacral column in Psittacosaurus mongoliensis and P. xinjiangensis (Brinkman et al. 2001). In the latter species, the tendons extend along at least the proximal half of the tail. In P. sinensis, in contrast, ossified tendons appear to be absent. In P. mongoliensis, individual tendons span approximately four or five vertebrae, intertwining without apparent order. The slender first sacral rib, fused proximally to the transverse process of the first sacral, attaches distally to the midsection of the preacetabular process of the ilium. The second to the sixth sacral ribs fuse to the sides of the sacral centra. Each rib is offset slightly cranially from its respective vertebra, such that the corner of each rib fuses to the caudal corner of the next sacral centrum.
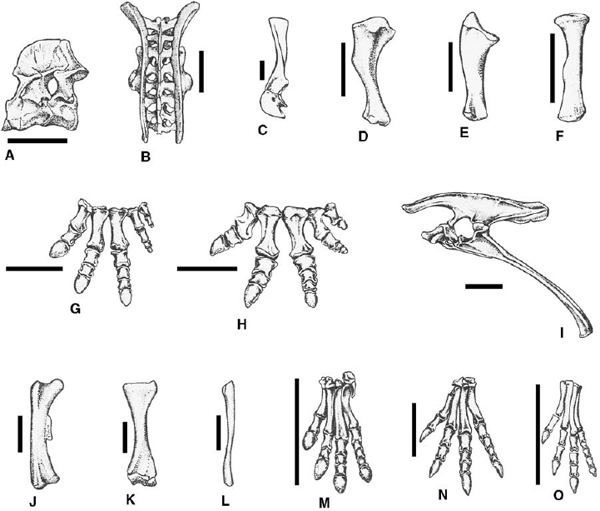
FIGURE 22.6. Postcranial skeletal anatomy of basal ceratopsians: A, syncervical of Protoceratops andrewsi (left lateral view); B, sacrum and ilia of Montanoceratops cerorhynchus (dorsal view); C, left scapulocoracoid of Leptoceratops gracilis (lateral view); D, left humerus of Leptoceratops gracilis (caudal view); E, left ulna of Leptoceratops gracilis (medial view); F, left radius of Leptoceratops gracilis (cranial view); G, left manus of Protoceratops andrewsi (dorsal view); H, left manus of Leptoceratops gracilis (dorsal view); I, pelvis of Protoceratops andrewsi (left lateral view); J, left femur of Montanoceratops cerorhynchus (caudal view); K, left tibia of Montanoceratops cerorhynchus (cranial view); L, left fibula of Montanoceratops cerorhynchus (lateral view); M, left pes of Protoceratops andrewsi (dorsal view); N, left pes of Montanoceratops cerorhynchus (dorsal view); O, left pes of Graciliceratops mongoliensis (dorsal view). Scale = 10 cm. (A, G, I, M after Brown and Schlaikjer 1940b; B, J–L, N after Brown and Schlaikjer 1942; C–F, H, after Brown 1914b; O after Maryaska and Osmólska 1975.)
The number of sacrals in basal neoceratopsians varies from six to eight. There are six sacrals in Archaeoceratops and Leptoceratops, seven in Graciliceratops (Maryaska and Osmólska 1975; Sereno 2000), and eight in Protoceratops and Montanoceratops (fig. 22.6B; Brown and Schlaikjer 1942). Sacral fusion is less derived in basal neoceratopsians than in ceratopsids, but an acetabular bar, formed by the fusion of the distal ends of the sacral ribs, is present. Neural spines are tall and always separate from each other. The sacrum is not arched, and the ventral surface is not excavated.
In psittacosaurids, the caudal centra exhibit a regular decrease in length and height along the tail. The decrease in height is greater than the decrease in length, resulting in elongate cylindrical centra at the distal end of the tail. The fused caudal ribs decrease rapidly in length from the first caudal distally, disappearing at midlength along the tail in Psittacosaurus mongoliensis and P. sinensis. The neural spines and zygapophyses decrease in height more gradually, the latter recognizable until the last few vertebrae. The first chevron articulates between the second and third caudal centra. The chevrons, which are always longer than the respective neural spines, decrease in length distally, disappearing in P. mongoliensis near the end of the tail.
Complete caudal series are rarely found in basal neoceratopsians. One individual of Leptoceratops has 38, and another, 20% larger, has 48, suggesting an ontogenetic increase in caudal count. The caudals are simple, the size of all components decreasing steadily toward the end of the tail. Transverse processes terminate about halfway down the tail (caudal 13 in Protoceratops and 18 in Leptoceratops). Chevrons are about as long as their corresponding neural spines but they incline backward more strongly and terminate about five segments from the end of the tail. A unique feature of Protoceratops and Montanoceratops is elongation of the proximal neural spines. In Protoceratops, the neural spines increase in height until caudal 14, accounting for two-thirds of the length of the tail, and then decrease to the end of the tail.
APPENDICULAR SKELETON
The pectoral girdle and forelimb of psittacosaurids show some important characters that differ from those of basal neoceratopsians (fig. 22.6C–O), while the pelvic girdle and hind limb of both psittacosaurids and basal neoceratopsians are similar.
In psittacosaurids, the scapula is long and has a narrow blade with a prominent acromial process. The scapula shares equally in the formation of the glenoid with the coracoid, a subquadrate bone with a well-developed cranioventral process. Psittacosaurus mongoliensis has an ossified clavicle. The supposed clavicle in P. sinensis (Young 1958a), however, appears to represent a displaced first ceratobranchial (Sereno 1987). In P. mongoliensis, the short, strap-shaped clavicle lies along the cranial margin of the coracoid and would not have reached the midline. A pair of platelike crescentic sternals is centered on the midline with a section of their thin medial edges in contact. The cranial end of each sternal is rounded and apparently nonarticular. The rectangular caudal end is scalloped for attachment to cartilaginous extensions of the cranial dorsal ribs.
In basal neoceratopsians, a scapular spine is incipient, usually crossing from the craniodorsal blade to the caudal supraglenoid ridge. The coracoid is moderately prominent, with a protracted caudal process that effectively limited the range of humeral extension. The coracoid fuses to the scapula in adults. Ossified clavicles are known in Protoceratops and Leptoceratops. Paired, flattened, bean-shaped sternals are also known. There is always a strong disparity in length of the forelimbs and hindlimbs, forelimbs being only about 70% of the length of the hindlimbs in forms like Bagaceratops, Protoceratops, and Graciliceratops, compared to 58% in Psittacosaurus.
In psittacosaurids, the humerus is shorter than the scapula, and the broadly expanded deltopectoral crest of the humerus forms a rectangular sheet of bone, which projects at an angle to the shaft. The poorly defined head is directed craniodorsally toward the glenoid cavity. In contrast, in basal neoceratopsians, the head of the humerus is hemispherical and eccentric, extending onto the proximocaudal surface of humerus. The radius and ulna are stoutly constructed with moderately expanded proximal and distal ends. The ulna, more robust than the radius, terminates proximally in a blunt olecranon process.
The carpus and manus are known primarily in P. mongoliensis. The carpus is composed of four carpals: the radiale, intermedium, ulnare, and distal carpal 3, which are not fitted tightly to one another. The manus is asymmetrical, showing strong reduction of the fourth digit and loss of the fifth. The inner three digits are robust and terminate in broad unguals. The phalangeal formula is 2-3-4-1-0. The first metacarpal and first digit are stout. The medial distal condyle of the first metacarpal is more proximal in position, which results in medial offset of the subsequent phalanges. During flexion, the phalanges of the first digit converge toward the center of the palm. The second metacarpal is longer than the first, and the phalanges of the second digit curve gently medially. Metacarpal III is the longest metacarpal and digit III is the longest digit. Metacarpal IV, in contrast to the inner three metacarpals, has a cylindrical shaft, which terminates distally without the development of paired condyles. Digit IV is reduced with only a small terminal phalanx.
In basal neoceratopsians, the manus is always smaller than the pes. Five carpals are ossified in Protoceratops. The manus is comparatively broad, with a phalangeal formula of 2-3-4-3-1 or 2. The unguals on the first three digits are blunt, and digits IV and V terminate in small nubbins of bone. In Leptoceratops, all phalanges, except for the unguals, are wider than they are long.
In psittacosaurids, the ilium is low with a straplike preacetabular process. The postacetabular process, subequal in length to the preacetabular process, is proportionately deeper. The acetabulum is completely open without any development of a descending iliac flange. The narrow pubic peduncle projects cranioventrally. The ischial peduncle, in contrast, is robust and projects ventrolaterally. A distinct, ovate surface on the acetabular margin of the ischial peduncle articulates against the broad proximal end of the femur. The two transversely compressed peduncles of the ischium join caudal to the acetabulum and pass distally as a flattened blade, angling ventromedially toward the midline at approximately 30° above the horizontal. Contact between right and left blades, which constitutes the ischial symphysis, is restricted to a small facet on the medial corner of the distal margin. The pubis is small relative to the ilium and ischium but remains an integral part of the margin and articular surface of the acetabulum. The acetabular surface is located on the body of the pubis and faces caudolaterally and slightly dorsally. A short prepubic process projects craniolaterally, terminating before the tip of the preacetabular process of the ilium. The slender pubic shaft, preserved only in Psittacosaurus mongoliensis and P. sinensis, projects caudomedially and passes along the ventral margin of the ischium. It does not join its opposite to form a pubic symphysis.
The pelvic girdle of basal neoceratopsians is generally similar to that of psittacosaurids. The ilium of Leptoceratops is low and shows no eversion of the dorsal margin. In Protoceratops, eversion of the preacetabular process is incipient. The pubis is a small, short element. The ischium is long, slender, and straight or gently decurved and lacks an obturator process.
In psittacosaurids, the femur is gently bowed in lateral view. The head, which arches medially from the shaft, shares the formation of a convex proximal articular surface with the greater trochanter. In lateral view, the width of the latter is several times the craniocaudal width of the fingerlike cranial trochanter, which projects dorsally from the craniolateral side of the proximal shaft. Just above midshaft, a pendent fourth trochanter projects from the caudomedial side and is flanked medially by an oval depression. The distal condyles are separated by a shallow depression cranially and by a much deeper flexor groove caudally. The laterally curving cnemial crest of the tibia diminishes distally along the shaft. The compressed distal end of the tibia is rotated approximately 70° from the transversely compressed proximal end. The narrow distal articular surface is divided into a small lateral condyle, which extends behind the calcaneum, and a larger medial condyle, which is capped ventrally by the astragalus. The fibula is modestly expanded at both ends, which exhibit rotation to the degree observed in the tibia. The distal end lies against the flat cranial surface of the tibia and butts distally against the calcaneum. The femur of basal neoceratopsians is similar to that of Psittacosaurus, shorter than the tibia, with a pendent fourth trochanter.
The tarsus of psittacosaurids, known from articulated specimens in several species, consists of the astragalus, calcaneum, and lateral and medial distal tarsals. The cup-shaped astragalus tightly caps the medial distal condyle of the tibia. The short ascending process of the astragalus usually establishes a minor contact with the distal end of the fibula. The small wedge-shaped calcaneum is positioned on the cranial side of the tibia, articulating proximally with the fibula and distally with the lateral distal tarsal. The medial and lateral distal tarsals are tabular and positioned over the third and fourth metatarsals, respectively. The compact digitigrade pes, known largely in Psittacosaurus mongoliensis and P. sinensis, does not depart significantly from the primitive ornithischian condition. The first digit, for example, is significantly shorter than the second, third, and fourth digits, and the fifth digit is reduced to a metatarsal splint. The short fifth metatarsal appears to articulate proximally against the lateral side of the lateral distal tarsal. The phalangeal formula is 2-3-4-5-0. As in the manus, the unguals are broad and all preceding phalanges have well-developed distal condyles.
The foot is also compact in basal neoceratopsians. In Archaeoceratops and Protoceratops, the foot is decidedly elongate. There are four functional metatarsals and frequently a small splint representing metatarsal V. The phalangeal formula in all neoceratopsians is 2-3-4-5-0. Unguals range from acute claws in Archaeoceratops and Leptoceratops to blunt tapered in Protoceratops.
Systematics and Evolution
Monophyly of Ceratopsia, Psittacosauridae, and Neoceratopsia is well established (Sereno 1986, 1997, 1999a, 2000; Dodson and Currie 1990; Makovicky 2001; Xu et al. 2002a; You 2002; You and Dodson 2003). However, the internal topology of these clades has been unstable. Consequently, we performed a cladistic analysis based on a data matrix of 11 taxa and 148 characters. Hypsilophodon foxii and Stegoceras validum were selected as outgroups, representing Ornithopoda and Pachycephalosauria, respectively. Ingroups included Psittacosaurus mongoliensis, Chaoyangsaurus youngi, five well-represented nonceratopsid neoceratopsians (Archaeoceratops oshimai, Leptoceratops gracilis, Bagaceratops rozhdestvenskyi, Protoceratops andrewsi, and Montanoceratops cerorhynchus), and two ceratopsids (Centrosaurus apertus and Triceratops horridus).
The 148 characters come from Sereno (2000) and Makovicky (2001), with an additional 10. Sereno (2000) and Makovicky (2001) used 72 and 98 characters, respectively; among them, 32 are equivalent, although not expressed the same way. The 148 characters include 77 from the skull, 20 from the lower jaw, 20 from the dentition, and 31 from the postcranial skeleton.
A PAUP*4.08b analysis was performed, with all characters unordered and equally weighted. Delayed transformation is chosen for character-state optimization. Branch-and-bound search found a single most parsimonious tree: L = 232 steps, CI = 0.74, and RI = 0.79 (fig. 22.7).
Psittacosaurus is the sister group to Neoceratopsia. Chaoyangsaurus is identified as the most basal Neoceratopsia (see also Sereno 2000; contra Makovicky 2001). Archaeoceratops is the sister taxon to Coronosauria, which includes Protoceratops, Triceratops, their most recent common ancestor, and all descendants (Sereno 1998). Protoceratops and Bagaceratops are closely related and are members of Protoceratopsidae. Leptoceratops is the sister taxon to Montanoceratops, and together they form Leptoceratopsidae, which is a member of Ceratopsoidea and the sister group to Ceratopsidae. Psittacosauridae includes a single genus, Psittacosaurus, and seven possible valid species, P. mongoliensis, P. sinensis, P. xinjiangensis, P. meileyingensis, P. neimongoliensis, P. ordosensis, and P. mazongshanensis (Osborn 1923a; Young 1958a; Sereno and Chao 1988; Sereno et al. 1988; Russell and Zhao 1996; Xu 1997; Xu and Zhao 1999; Sereno 2000). Psittacosauridae is characterized by the following features: short preorbital portion (less than 40% of the basal skull length); elevated external naris; lack of antorbital fossa and fenestra; extremely broad caudolateral premaxillary process; long rostral process of the nasal extending below the external naris; convergence of premaxilla, maxilla, lacrimal, and jugal sutures to a point on the snout; eminence on the rim of the buccal emargination of the maxilla near the junction with the jugal; an unossified gap in the wall of the lacrimal canal; a well-developed jugal process from the midsection of the jugal; elongate jugal and squamosal processes of the postorbital; a dentary crown with a bulbous primary ridge; manual digit IV with only one simplified phalanx; manual digit V absent.
Neoceratopsia, the stem-based clade defined as all ceratopsians more closely related to Triceratops than to Psittacosaurus, shares the following characters: an enlarged head relative to the body; a keeled rostral end of the rostral bone; a short jugal process of the postorbital; a much reduced quadratojugal; the basioccipital excluded from the formation of the foramen magnum; the coronoid process covering the caudal tooth row in lateral view; a primary ridge on the maxillary teeth; a caudal process on the coracoid; the development of the humeral head; and a gently decurved ischium.
Chaoyangsaurus is represented by partial cranial and postcranial material from the Tuchengzi Formation in Liaoning Province, China (Zhao et al. 1999). The age of the upper part of the Tuchengzi Formation could reach the earliest Cretaceous (Swisher et al. 2002). Chaoyangsaurus is the most basal member of Neoceratopsia and differs from Psittacosaurus in possessing a large skull, a keeled predentary with a narrow caudoventral process, a reduced retroarticular process, and straight and sub-cylindrical premaxillary teeth, as in later neoceratopsians.
The late Early Cretaceous Archaeoceratops from northwest Gansu Province, China (Dong and Azuma 1997; Tang et al. 2001b; You 2002; You and Dodson 2003) constitutes the sister taxon to all other neoceratopsians, a clade known as Coronosauria. Derived features that occur in Archaeoceratops focus on two bones: the rostral and the jugal. The rostral became keeled, pointing ventrally along its rostral margin, and developed a caudolateral process along its ventral edge. The lateral expansion of the jugal migrated from the midsection to the caudal end, and a prominent crest developed along its lateral surface. Changes also occurred in the dentition. The primary ridge of the maxillary tooth crown becomes prominent and is placed caudal to the midline. The last dentary tooth is situated beside or caudal to the apex of the pronounced coronoid process.
The skull of Coronosauria has an elongated preorbital portion, more than half of the basal skull length, with an oval, rather than a triangular, antorbital fossa. In dorsal view, the supratemporal fenestra is triangular and directed caudolaterally, probably as a result of the more developed frill. Another important change that occurred in Coronosauria is the development of the syncervical (unknown in Archaeoceratops), in which elements of the atlas, axis, and several proximal cervical vertebrae are fused together to support the enlarged head.
The clade of Asian Protoceratops and Bagaceratops, basal among coronosaurs, appears to have evolved slowly compared to North American Ceratopsoidea, but also achieved many changes. A small nasal horn is situated caudal to the external naris. The quadratojugal is triangular in coronal section with a slender rostral prong articulating with the jugal. The palatine has an elongate parasagittal process. The lower jaw is modified in several ways. For example, the rostral end of the predentary is rostrodorsally pointed, the surangular has a long ventral process that overlaps the angular, the surangular-dentary and surangular-angular sutures form an acute angle on the lateral face of the mandible, and the caudal end of the splenial is bifid and overlaps the angular.
Two species are known for Protoceratops: P. andrewsi and P. hellenikorhinus. The latter was discovered from the Upper Cretaceous Djadokhta Formation of Inner Mongolia (Lambert et al. 2001). It is the larger of the two and shows some distinct differences from the former, such as the absence of premaxillary teeth, two small separated nasal horns, a strong frontoparietal depression, and a forwardly recurved frill with reentrant of the parietals along the midline.
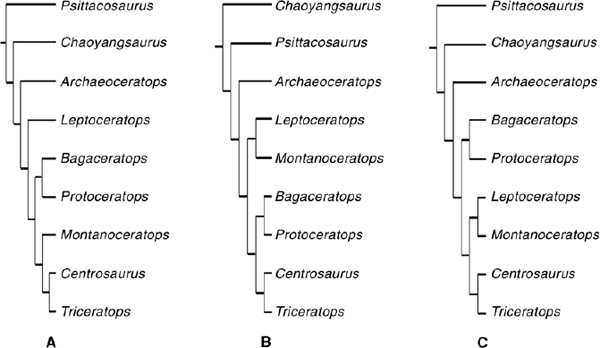
FIGURE 22.7. Three views on the phylogenetic relationships of basal Ceratopsia. (A after Sereno 2000; B after Makovicky 2001; C this chapter, a single most parsimonious tree with a L = 232 steps, CI = 0.74, and RI = 0.79.)
The clade of Leptoceratops and Montanoceratops marks another major step in the evolution of horned dinosaurs on the way to Ceratopsidae. A suite of features characterizes Ceratopsoidea, which is not seen in Protoceratopsidae. In the facial region, the external naris is round, the caudolateral process of the rostral is elongated, and the ventral margin of the premaxilla is convex. The caudal portion of the skull shows a tendency toward downward placement: the lateral expansion of the jugal projects more ventrally than laterally; the squamosal develops a postquadrate ventrally directed process, the exoccipital is in contact with the quadrate, and the supraoccipital is in the same plane as the caudal face of the basioccipital rather than inclining rostrally. The manus is stout, with the nonungual phalanges wider than long.
Several basal ceratopsians were deemed too incomplete for the phylogenetic analyses conducted here. Udanoceratops tschizhovi from the Late Cretaceous of Inner Mongolia (Kurzanov 1992) is probably closely related to Protoceratopsidae, as indicated by the elliptical external naris, the short ventral edge of the rostral, and the flat ventral margin of the maxilla. However, the ventrally curved ventral margin of the lower jaw is shared by Udanoceratops and Leptoceratops. Graciliceratops mongoliensis (formerly Microceratops gobiensis from the Late Cretaceous of Mongolia, Maryaska and Osmólska 1975) is characterized by the slender median and caudal parietal frill margins and high tibiofemoral ratio (1.2:1), and it is probably a member of Protoceratopsidae (Sereno 2000). Another Late Cretaceous Mongolian neoceratopsian, Breviceratops kozlowskii (Kurzanov 1990a, 1990b), is considered a juvenile specimen of Bagaceratops (Sereno 2000; Makovicky 2001).
Two recently described nonceratopsid ceratopsians could not be included in the cladistic analyses, although they are likely to have an important impact on the topology of ceratopsian phylogeny: Liaoceratops yanzigouensis and Zuniceratops christopheri. L. yanzigouensis is represented by two almost complete skulls (one adult, one juvenile) from the Lower Cretaceous Yixian Formation of Liaoning Province, China. Cladistic analysis (Xu et al. 2002a) indicates that Liaoceratops, not Chaoyangosaurus, is the most basal neoceratopsian, while the latter is repositioned as the most basal member of Ceratopsia. Derived characters shared by Liaoceratops and other neoceratopsians include the lateral processes of the rostral, an expanded frill with squamosal participation, a spherical occipital condyle, a deep temporal bar, a triangular postorbital and laterally convex surangular. Liaoceratops exhibits characters traditionally used to diagnose either psittacosaurids or more exclusive clades within Neoceratopsia, such as a weak ventral flange on the dentary and a ventrally wide infratemporal fenestra. Liaoceratops retains a number of primitive ceratopsian characters, such as a flat rather than a transversely expanded quadratojugal, the unkeeled rostral, the absence of an epijugal, and weakly developed primary ridges and oblique occlusion angles of the maxillary teeth.
In contrast, Zuniceratops christopheri is considered the sister group to Ceratopsidae. From the Moreno Hill Formation (Turonian) of west-central New Mexico, Z. christopheri is represented by partial cranial and postcranial remains of as many as five individuals (Wolfe and Kirkland 2001) but has not yet been fully described. Its considerable age compared to well-known North American ceratopsians and its suite of characters make this a taxon of unusual interest. Large paired postorbital horns would otherwise be attributed to a member of Ceratopsidae, but teeth with simple (undivided) roots preclude such membership. A mosaic of both generalized and derived features is seen throughout the skull and skeleton. The external naris is enlarged and ovate, differing from both centrosaurines and chasmosaurines (Wolfe 2000). There is no nasal horn core. Several partial squamosals suggest a well-developed but thin frill, but details are unknown. The occlusal plane of the teeth is vertical as in ceratopsids but there is only a single functional tooth per tooth row; teeth of the largest individual give evidence of incipient division of the roots. Postcranial skeletal elements are slender, but the ischium is strongly decurved. Wolfe and Kirkland (1998) considered Zuniceratops a close relative of Ceratopsidae based on the presence of orbital horns in both.
Biostratigraphy, Taphonomy, Paleoecology, and Biogeography
Psittacosaurids are broadly distributed in central Asia, ranging from eastern and western China and Mongolia to southern Siberia. We regard the report of a specimen from Thailand (Buffetaut and Suteethorn 1992) as unsubstantiated. Psittacosaurids are among the most abundant of all dinosaurs (Dodson 1990b) and occur over a range of environmental conditions. Association with sediments indicating semiarid environments and eolian sands is well known (e.g., Russell and Zhao 1996), but in Inner Mongolia and Liaoning specimens are found in fluviolacustrine deposits containing aquatic invertebrates and vertebrates (Eberth et al. 1993; Swisher et al. 1999).
Psittacosaurids are best known from the later Early Cretaceous (Barremian–Aptian or possibly Albian), but there are reports of Psittacosaurus as old as Berriasian–Valanginian from the Sangiin Dalai Nuur depression (Bakhurina 1983; Shuvalov 2000). The later date has been established by associated pollen and spores in Mongolia for Psittacosaurus mongoliensis (Bratzeva and Novodvorskaja 1975) and in the Gansu Province of China for P. mazongshanensis (Tang et al. 2001b); and by radiometric Ar/Ar dates from Liaoning for Psittacosaurus specimens from the Yixian Formation (Swisher et al. 1999, 2002). In Mongolia, an Early Cretaceous age is also supported by the associated invertebrate faunas (including ostracodes and mollusks) and the vertebrate faunas, including freshwater fish (Lycoptera), turtles (Hangiemys, Mongolemys), dinosaurs (Iguanodon, Shamosaurus), and mammals (Prokennalestes, Prozalambdalestes) (Cockerell 1924; Kalandadze and Kurzanov 1974; Shuvalov 1975; Tumanova 1981). Several species of Psittacosaurus are found in Inner Mongolia (Russell and Zhao 1996), and dates, including that from the widespread P. mongoliensis, fall into the range of Barremian to Aptian. At Tebch, fossil-bearing sediments are overlain by a basalt at 110.6 Ma, thus of mid-Aptian age. A rich palynoflora dominated by gymnosperms is found in lacustrine deposits with associated insects, conchostracans, ostracodes, fish, salamanders, frogs, turtles, lizards, pterosaurs, theropods, birds, and mammals (Swisher et al. 1999). In Shandong Province, psittacosaurid-bearing deposits (P. sinensis, P. youngi) are indisputably overlain by Upper Cretaceous sediments and faunas (e.g., the hadrosaurids Tsintaosaurus and Tanius; Young 1958a).
Psittacosaurus was a small-bodied herbivore. The slicing psittacosaurid dentition, with its self-sharpening cutting edges, is suited for mastication of plant material. Polished gastroliths, exceeding 50 in number, are associated with two psittacosaurid skeletons and must have played a significant role in the break-down of plant materials.
When limb length is estimated by the addition of respective propodials, epipodials, and third metapodials, the length of the forelimb in Psittacosaurus mongoliensis is approximately 58% that of the hindlimb. This percentage of hindlimb length is higher than in Hypsilophodon foxii, which has a forelimb/hindlimb ratio of 51%. Proportions within the hindlimb in Psittacosaurus are comparable to the presumed cursorial ornithopods Hypsilophodon, Dryosaurus, and Parksosaurus (Janensch 1955; Galton 1973e) and the neoceratopsian Graciliceratops (Maryaska and Osmólska 1975; Sereno 2000). The tibia is slightly longer than the femur, as in Dryosaurus and Graciliceratops, but proportionately less elongate than in Hypsilophodon and Parksosaurus. Given the proportions of the fore- and hindlimbs, it is reasonable to suppose that P. mongoliensis was facultatively bipedal; that is, a capable and perhaps habitual biped at most speeds. The length of the forelimb and the structure of the manus, however, do not preclude effective use of the forelimb in locomotion. The divergence of digit I suggests a limited grasping capability in the manus, which may have been used to procure vegetation.
A growth series of Psittacosaurus mongoliensis from Khamaryn-Us, Hühteeg Svita, Mongolia, was subjected to a histological study (Erickson and Tumanova 2000). The sample ranged from estimated body masses of 0.9 to 19.9 kg. It apparently took nine years to achieve the maximum body size observed. This translates to a growth rate of about 12.5 g/day, four times higher than unspecified reptiles and one-quarter higher than marsupials, but 3–20 times slower than eutherian mammals or birds.
The fossil record of basal neoceratopsians is generally good. About half of the genera are known from complete skulls, and many have multiple specimens and associated skeletal material (Chinnery and Trexler 1999). Juveniles (Dodson 1976) and embryos (Dong and Currie 1993; Weishampel et al. 2000) form an important part of the record. Asia preserves more members of basal neoceratopsians than North America does, and most of them are Late Cretaceous, especially late Late Cretaceous.
Basal neoceratopsian fossils are found in a wide range of sedimentary environments. Remains of Protoceratops andrewsi are found in areas of eolian dunes (Osmólska 1980; Jerzykiewicz et al. 1993; Eberth 1993; Fastovsky et al. 1997). The climate was hot, semiarid, and seasonal. It is significant that protoceratopsids are absent in the mesic Nemegt Formation of Mongolia, which overlies the Djadokhta and Barun Goyot formations. In North America, basal neoceratopsian remains are found in sediments laid down in upper plain regions (Horner 1984c) that appear to have been seasonally dry. There is little evidence of basal neoceratopsians in the rich fauna of the contemporary lower coastal plain deposits of Alberta and Montana (Ryan and Currie 1998). The coexistence of the basal Leptoceratops gracilis and the derived Triceratops horridus in terminal Cretaceous strata of Alberta is an interesting paradox that occurred in cooler, drier environments distant from the shores of a receding inland sea.
Protoceratops andrewsi and other basal neoceratopsians appear to have been gregarious animals that nested communally. Fifteen Protoceratops hatchlings were collected from a single nest from the eolian sandstones in the central Gobi region (Weishampel et al. 2000). They are oriented in a common direction to the southeast and faced away from the prevailing wind. No eggshell was found within this nest, the same condition also reported for the embryos from Bayan Mandahu of China (Dong and Currie 1993). The discovery of a skeleton of Protoceratops andrewsi entangled with that of the dromaeosaurid Velociraptor mongoliensis (Kielan-Jaworowoska and Barsbold 1972; Barsbold and Perle 1983) suggests that the latter preyed on the former. What once appeared to be the eggs of Protoceratops have turned out to be the eggs of the predators themselves, Oviraptor (Norell et al. 1995). A preliminary study has shown that the style of Protoceratops posture was neither mammalian nor reptilian (Matsumoto 1997).
Sexual dimorphism in Protoceratops andrewsi has been recognized for both the cranial (Kurzanov 1972; Dodson 1976) and the postcranial features (Tereshchenko 2001), and is further shown in the second species of Protoceratops, P. hellenikorhinus (Lambert et al. 2001). The Protoceratops frill is well developed, and it is clear that it was sexually dimorphic.
According our phylogenetic analysis, convergent evolution of horns and frills occurs in Protoceratopsidae and Ceratopsoidea. In Protoceratopsidae, the nasal horns are situated far caudal to the external naris and therefore closer to the orbit, while in Ceratopsoidea, the nasal horns are placed dorsal to the external naris. The frill of protoceratopsids is fenestrated and parietal dominated, with a partial contribution from the caudal extension of the squamosal, ending with a nondecorated and straight caudal edge (Dodson 1993). In contrast, in ceratopsoids the squamosal is expanded both caudally and caudoventrally, even in forms with a short frill, such as Leptoceratops (Sternberg 1951) and Avaceratops (Penkalski and Dodson 1999).
Ceratopsoids evolved a different feeding mechanism from protoceratopsids, as shown by changes in the rostral portion. In Ceratopsoidea, the rostral and premaxilla elongated rostrocaudally. The rostral developed a long caudolateral process covering the ventrally curved ventral edge of the premaxillary. The premaxillary teeth were lost. This configuration is probably related to a more efficient crushing function, associated with other modifications of the dentition, such as the lingual-buccally widened cheek teeth. In contrast, in Protoceratopsidae, the rostral extended further ventrally to accommodate the rostrodorsally pointed predentary, and the premaxillary teeth remained. This structure might be useful for selective cropping and feeding.
An Asian origin for Neoceratopsia is suggested by the basal neoceratopsian status of both the Late Jurassic Chaoyangsaurus (Zhao et al. 1999; Swisher et al. 2002) and the late Early Cretaceous Archaeoceratops. Recognition of separate clades in Asia and North America in the evolution of basal Ceratopsia brings a biogeographic coherence that was not apparent previously (You 2002; You and Dodson 2003). Divergence between North American Ceratopsoidea and Asian Protoceratopsidae probably occurred in the early Late Cretaceous. Ceratopsoidea could have originated either in Asia or in North America, and the further finds of basal Ceratopsoidea in Asia or Protoceratopsidae in North American will help to clarify this. The origin of Ceratopsidae was probably in North America, as the entire fossil record of this group and its sister group, Leptoceratopsidae, is currently known only in North America. However, it is possible that there were further exchanges of Protoceratopsidae and Ceratopsoidea between Asia and North America.