CHAPTER 2
WHAT IS THE EVIDENCE FOR EVOLUTION ON EARTH?
The pre-Darwinian theories of evolution had little impact. Even though some evolutionary thinking was widespread among geologists, biologists, and even among literary people and philosophers, the Biblical story of Creation, as told in the book of Genesis, Chapters 1 and 2, was virtually unanimously accepted not only by laypeople but also by scientists and philosophers. This changed overnight, so to speak, in 1859 with the publication of Charles Darwin’s On the Origin of Species. Even though some of Darwin’s explanatory theories of evolution continued to encounter much resistance for another 80 years, his conclusion that the world had evolved was widely accepted within a few years after 1859.
However, throughout the nineteenth century whenever people talked about evolution, they referred to it as a theory. To be sure, at first, the thought that life on Earth could have evolved was merely a speculation. Yet, beginning with Darwin in 1859, more and more facts were discovered that were compatible only with the concept of evolution. Eventually it was widely appreciated that the occurrence of evolution was supported by such an overwhelming amount of evidence that it could no longer be called a theory. Indeed, since it was as well supported by facts as was heliocentricity, evolution also had to be considered a fact, like heliocentricity. This chapter will be devoted to a presentation of the evidence that led to the adoption of the “evolution is a fact” conviction among scientists. It will also challenge those who are still not yet convinced of the occurrence of evolution.
Evolution is a historical process that cannot be proven by the same arguments and methods by which purely physical or functional phenomena can be documented. Evolution as a whole, and the explanation of particular evolutionary events, must be inferred from observations. Such inferences subsequently must be tested again and again against new observations, and the original inference is either falsified or considerably strengthened when confirmed by all of these tests. However, most inferences made by evolutionists have by now been tested successfully so often that they are accepted as certainties.
WHAT EVIDENCE DOES THE EVOLUTIONIST HAVE?
The evidence for evolution is now quite overwhelming. It is presented in great detail by Futuyma (1983, 1998), Ridley (1996), and Strickberger (1996), and also in the anticreationist volumes listed in Chapter 1. My own treatment focuses on the classes of evidence now available to document evolution. It shows how remarkably congruent are the conclusions drawn from the most diversified branches of biology, which all support evolution. Indeed, these findings would make no sense in any other explanation.
The Fossil Record
The most convincing evidence for the occurrence of evolution is the discovery of extinct organisms in older geological strata. Some of the remnants of the biota that lived at a given geological period in the past are embedded as fossils in the strata laid down at that period. Each earlier stratum contains the ancestors of biota fossilized in the succeeding stratum. The fossils found in the most recent strata are often very similar to still living species or, in some cases, even indistinguishable. The older the strata are in which a fossil is found—that is, the further back in time—the more different the fossil will be from living representatives. Darwin reasoned that this is to be expected if the fauna and flora of the earlier strata had gradually evolved into their descendants in the later, more recent strata.
Given the fact of evolution, one would expect the fossils to document a gradual steady change from ancestral forms to the descendants. But this is not what the paleontologist finds. Instead, he or she finds gaps in just about every phyletic series. New types often appear quite suddenly, and their immediate ancestors are absent in the earlier geological strata. The discovery of unbroken series of species changing gradually into descending species is very rare. Indeed the fossil record is one of discontinuities, seemingly documenting jumps (saltations ) from one type of organism to a different type. This raises a puzzling question: Why does the fossil record fail to reflect the gradual change one would expect from evolution?
All of his life Darwin insisted that this is simply due to the unimaginable incompleteness of the fossil record. Only an incredibly small fraction of organisms that had once lived are preserved as fossils. Often the fossil-bearing strata were on plates that were subsequently subducted and destroyed in the process of plate tectonics. Others were strongly folded, compressed, and metamorphosed, obliterating the fossils. Only a fraction of the fossil-bearing strata is presently exposed at the Earth’s surface. But it is even highly improbable that any organism ever becomes fossilized at all, since most dead animals and plants are either eaten by scavengers or decay. They become fossilized only when, immediately after death, they are buried by sediment or volcanic ash. Fortunately, occasionally a rare fossil is found that fills the gap between ancestors and modern descendants. Archaeopteryx, for instance, a primitive fossil bird of the upper Jurassic (145 million years ago), still had teeth, a long tail, and other characteristics of his reptilian ancestors. However, in other respects, for instance in its brain, large eyes, feathers, and wings, it is rather similar to living birds. Fossils that fill a large gap are referred to as missing links. The discovery of Archaeopteryx in 1861 was particularly gratifying because anatomists had already concluded that birds must have descended from reptilian ancestors. Archaeopteryx confirmed their prediction.
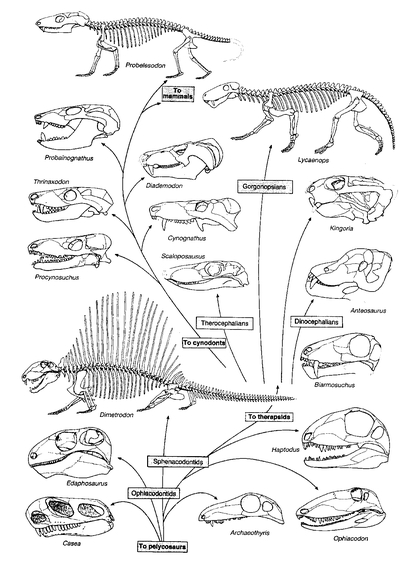
A few fossil lineages are remarkably complete. This is true, for instance, for the lineage that leads from the therapsid reptiles to the mammals (
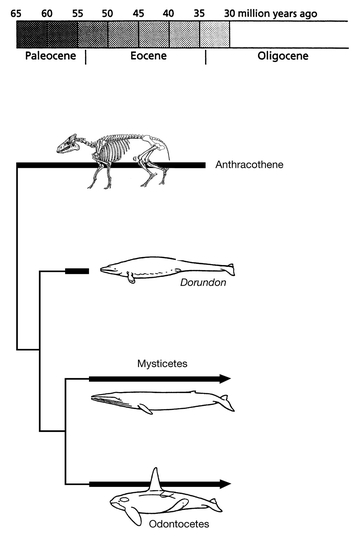
Fig. 2.1). Some of these fossils appear to be so intermediate between reptiles and mammals that it is almost arbitrary whether to call them reptiles or mammals. A remarkably complete set of transitions was also found between the land-living ancestors of the whales and their aquatic descendants. These fossils document that whales are derived from ungulates (mesonychid condylarthra) that increasingly became adapted to life in water (
Fig. 2.2). The
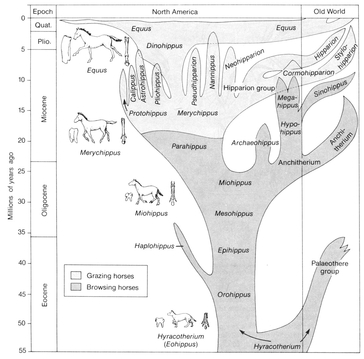
australopithecine ancestors of man also form a rather impressive transition from a chimpanzeelike anthropoid stage to that of modern man. The most complete transition between an early primitive type and its modern descendant that has been described is that between
Eohippus, the ancestral horse, and
Equus, the modern horse (
Fig. 2.3).
Evolution of the synapsid Reptilia, with the cynodonts forming a transition to the earliest mammals.
Source: Ridley, M. (1993). Evolution. Blackwell Scientific: Boston, p. 535. Reprinted by permission of Blackwell Science, Inc.
The study of phylogeny is really a study of homologous characters. Since all members of a taxon must consist of the descendants of the nearest common ancestor, this common descent can be inferred only by the study of their homologous character. But how do we determine whether or not the characters of two species or higher taxa are homologous? We say that they are if they conform to the definition of homologous: A feature in two or more taxa is homologous when it is derived from the same (or a corresponding) feature of their nearest common ancestor.
This definition applies equally to structural, physiological, molecular, and behavioral characteristics of organisms. But how are we to determine whether homology is substantiated in a particular case? Fortunately, there are numerous criteria (see Mayr and Ashlock 1991). For structures this includes the position in relation to neighboring structures or organs; by connecting two dissimilar features by intermediate stages in ancestors; by similarity in ontogeny; and by intermediate fossils. The best evidence for homology has been provided in recent years by molecular biology. Such research has provided reliable evidence on the relationship of nearly all higher taxa of animals, and rapid progress is now also being made in reconstructing the relationship of the higher taxa of plants. A taxon, delimited by the methods of Darwinian classification, and therefore consisting exclusively of descendants of the nearest common ancestor, is called monophyletic.
What is particularly convincing about fossil animal series is that each fossil type is found at the time level at which one ought to expect it. For instance, modern mammals began to evolve after the Alvarez extinction event at the beginning of the Paleocene (60 million years ago). No modern mammal, therefore, should be found in strata that are 100 or 200 million years old, and indeed none has ever been found. Or, to take another example, giraffes originated in mid-Tertiary times about 30 million years ago. It would upset all our beliefs and calculations if suddenly a fossil giraffe was found from the Paleocene 60 million years ago. But, of course, no such fossil has ever been found.
The descent of the whales from Eocene artiodactye ungulates is now reasonably well documented by transitional fossils.
Source: From various sources, particularly personal information from Prof. Philip D. Gingerich
Evolution of the horse family, from the Eocene Hyracotherium (Eohippus) to the modern horse (Equus). There was an extraordinary origin, flourishing, and extinction of types of horses in the Miocene.
Source: Strickberger, Monroe, W.,
Evolution, 1990, Jones and Bartlett, Publishers, Sudbury, MA.
www.jbpun.com. Reprinted with permission.
Formerly, the ages of these fossils were mere guesses. All one knew was that the lower strata were older than the higher strata. However, the clock provided by the constancy of radioactive decay now permits extremely precise age determinations of certain strata, particularly lavas and other volcanic deposits that appear between fossil deposits (see Box 2.1). Carbon dating can be used for the most recent past. The age of any fossil can now be determined with remarkable precision if one knows in what geological stratum it was found (
Fig. 2.4). At the turn of the twenty-first century, the sequence of accurately dated fossils has documented evolution in the most convincing manner (see page 37).
BRANCHING EVOLUTION AND COMMON DESCENT
The scala naturae was a linear progression from lower to higher, and in Lamarck’s presentation of evolution, each lineage originated with a (single-cell) infusorian believed to have originated by spontaneous generation. In the course of evolution its descendants became ever more complex and more perfect. Indeed, all pre-Darwinian evolutionary schemes postulated essentially straight phyletic lineages (see Chapter 4). One of Darwin’s major contributions was to have proposed the first consistent theory of branching evolution.
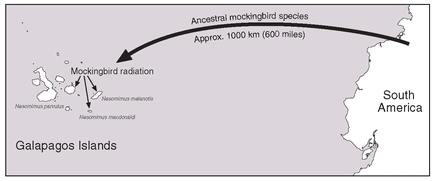
It was an observation he made on the birds of the Galapagos Islands that led him to the branching theory. The Galapagos Islands are actually peaks of submarine volcanoes that have never had a land connection with South America or any other continent. All of the Galapagos fauna and flora got there by over-water (distance) colonization. Darwin knew that there was only one species of mockingbird in South America, but he found a species of mockingbird on each of three islands in the Galapagos (
Fig. 2.5), with each species different from the others. He concluded quite rightly that a single colonization of the South American mockingbird had given rise, by branching descent, to three different species on three different islands in the Galapagos. Then, he further reasoned that probably all mockingbirds in the world had descended from a common ancestor, because they are basically so similar to each other. Mockingbirds and their relatives, like thrashers and catbirds, then presumably also had a common ancestor.
Box 2.1 Radioactive Clock
Certain rocks, mostly of volcanic origin (e.g., lava flows), contain radioactive minerals such as potassium, uranium, and thorium. Each of these minerals decays at a specific rate and physicists have determined their half-lives. Uranium 238, for instance, has a half-life of 4.5 billion years, producing lead 206 in the process. The age of a given rock can then be calculated from the ratio of uranium and lead. Sedimentary rocks, which do not contain radioactive minerals, are dated by their location relative to datable strata.
The geological timescale. The Precambrian ranges from the origin of life (ca. 3,800 million years ago) to the beginning of the Cambrian (ca. 543 million years ago). New fossil finds frequently require a correction of the date of the earliest occurrence of a higher taxon.
Source: Evolutionary Analysis 2nd ed. by Freeman/Herron, copyright © 1997. Reprinted by permission of Pearson Education, Inc., Upper Saddle River, NJ.
Colonization of the Galapagos Islands by an ancestral South American mockingbird species and its subsequent evolution into three local species.
This chain of inferences led Darwin to the ultimate conclusion that all organisms on Earth had common ancestors and that probably all life on Earth had started with a single origin of life. As Darwin wrote, “There is grandeur in this view of life, with its several powers, having been originally breathed into a few forms or into one; and that from so simple a beginning endless forms most beautiful and most wonderful have been, and are being, evolved” (1859: 490). As we shall presently show, numerous studies making use of different kinds of evidence have convincingly confirmed Darwin’s conjecture. It is now referred to as the theory of common descent.
Paleontologists, geneticists, and philosophers had long been puzzled over how and where the branching took place that leads to the phenomenon of common descent. This problem was solved by taxonomists, who showed that it is speciation, particularly often geographic speciation, that leads to branching (see Chapter 9).
The theory of common descent solved a long-standing puzzle of natural history. There seemed to be a basic conflict between the overwhelming diversity of life and the observation that certain groups of organisms often shared the same characteristics. Thus there were frogs, snakes, birds, and mammals, yet the basic anatomy of all of these so different appearing classes of vertebrates was very much the same, yet totally different from that of an insect. The theory of common descent provided the answer to this puzzling observation. When certain organisms share a series of joint characteristics, in spite of numerous other differences, it is due to the fact that they had descended from the same common ancestor. Their similarities were due to the heritage they had received from this ancestor, and the differences had been acquired since the ancestral lines had split.
How Well Is Common Descent Documented?
The fossil record provides abundant evidence for common descent. For instance, in mid-Tertiary strata we may find fossils that are the common ancestors of dogs and bears. In somewhat earlier strata we find common ancestors of dogs and cats. Indeed, paleontologists have succeeded in showing that all carnivores descended from the same common ancestral type. The same descent from the common ancestor is true for all rodents, all ungulates, and for all other orders of mammals. Indeed, this principle of common descent also holds true for birds, reptiles, fishes, insects, and all other groups of organisms.
Even before 1859, zoologists had been able to construct a rather detailed taxonomic hierarchy of animal taxa. What was still not yet understood was why there was such a hierarchy. It was Darwin who showed that it could be explained by the principle of common descent. All the species of a genus have a nearest common ancestor and so do all the species of a family or of any other higher category in the hierarchy. This joint ancestry is the reason why the members of a taxon are so similar to each other.
Morphological Similarity. Very suggestive evidence for common descent is also provided by the study of comparative anatomy. It was customary already in the eighteenth century to call certain organisms “related” when they were similar. At that time the French naturalist Comte Buffon described this for horses, donkeys, and zebras. The less similar that two kinds of organisms were, the less closely they were considered to be “related.” The systematists, the students of classification, used the degree of similarity to establish a hierarchy of taxonomic categories. The most similar organisms were placed in the same species. Similar species were placed in the same genus, similar genera in the same family, and thus all the way up to the taxa of the highest category.
The Linnaean hierarchy. Each category is nested within the next higher category, such as the species in the genus.
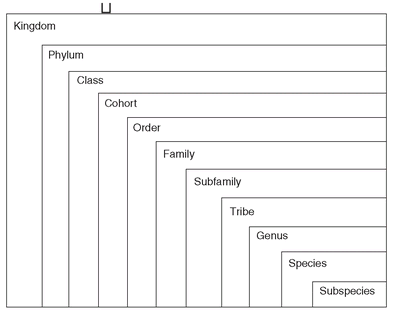
This arrangement of organisms by the degree of their similarity and relationship is called the
Linnaean hierarchy (
Fig. 2.6), after the Swedish botanist Carolus Linnaeus, who developed the system of binomial classification. Such a classification groups organisms into larger and larger taxa, finally comprising all the animals and all the plants. Beginning with a particular species, let us say the cat, one was able to construct this hierarchy. It was known that there were other species of cats rather similar to the house cat, which Linnaeus also placed in the genus
Felis. This group of cats could be combined with the lion, the cheetah, and other genera of cats into the family Felidae. This family of catlike mammals could then be combined with other predatory mammals such as the Canidae (doglike), Ursidae (bears), Mustelidae (weasels), Viverridae (civets), and related groups into the order of Carnivora.
In a similar manner, other mammals could be combined into the orders of Artiodactyla (deer and relatives), Perissodactyla (horses, etc.), Rodentia (rodents, etc.), and those of whales, bats, primates, marsupials, and so on to form the class Mammalia (mammals). A similar hierarchy exists for all other kinds of animals, such as birds and insects, and for plants. The nature and causation of this grouping, unless ascribed to creation, was a complete riddle until Darwin showed that it was evidently due to “common descent.” Each taxon (group of organisms), Darwin demonstrated, could be explained as consisting of the descendants from the nearest common ancestor, and such descent required evolution. The observed facts fit Darwin’s theory of evolution so perfectly that his theory of “common descent by modification” was accepted almost immediately after 1859. Classification, a most active occupation of so many nineteenth-century zoologists and botanists, now had an explanation. The most frequently used evidence, on the basis of which relationship and common descent was inferred, was morphological and embryological similarity, and the search for such similarity led in the second half of the nineteenth century to a great flowering of comparative morphology and embryology (Bowler 1996).
Phylogeny, a special branch of biology, deals with the pattern and history of the descent of organisms. The pattern of descent is often presented as a phylogenetic tree (dendrogram) or in a certain school of taxonomists as a cladogram. Inspired by Ernst Haeckel, a German zoologist and contemporary of Darwin, zoologists and botanists have devoted much time and effort to clarify the actual phylogeny of organisms (see Chapter 3).
The Explanation of Morphological Types A second, related branch of biology likewise found its explanation through common descent. The comparative anatomists, led by Georges Cuvier, had recognized a limited number of types of organisms that agreed with each other in their basic structure (archetype). Cuvier (1812) distinguished four major phyla (embranchements), all members of which, he thought, had the same Bauplan (body plan). The existence of these very distinct types, not connected by any intermediates or transitions, decisively refuted the validity of the scala naturae. Cuvier called these types Vertebrates, Mollusks, Articulates, and Radiates. This was a first step, but it was soon shown that three of his types were composite, while the vertebrates were ultimately classed as a subdivision of the Chordates. At the present time, about 30 phyla of animals are recognized, and in most of them several minor types are distinguished, for example, in the vertebrates there are fishes, amphibians, reptiles, birds, and mammals. Again, the existence of these morphological types made sense as soon as one recognized that each consisted of the descendants of a common ancestor who shared its basic body plan.
The preevolutionary morphologists, like Cuvier, were typologists (essentialists) in their thinking. They were followers of Plato. Each type (phylum) was considered to be completely separated from the others, it was defined by its essence, and it was constant. Even though the philosophical basis of this so-called idealistic morphology was quite wrong, its emphasis on the study of morphology led to numerous discoveries of great value for the reconstruction of phylogeny and, more broadly, for the understanding of evolution.
Homology It is quite remarkable how successful comparative morphology can be in the reconstruction of missing steps in an evolutionary sequence. T. H. Huxley, for instance, when reconstructing the nonflying ancestor of birds, concluded that it was an archosaurian reptile. Archaeopteryx, a remarkable bridge between birds and the archosaurians, was discovered only a few years later, in 1861. Evolutionary entomologists postulated that ants had evolved from wasplike ancestors and inferred what characters the earliest ants must have had. When a fossil ant was then discovered in mid-Cretaceous amber, it largely confirmed the inferred reconstruction. These are not isolated cases, for whenever a missing ancestor was reconstructed, it agreed remarkably well with the real ancestor subsequently discovered as fossil.
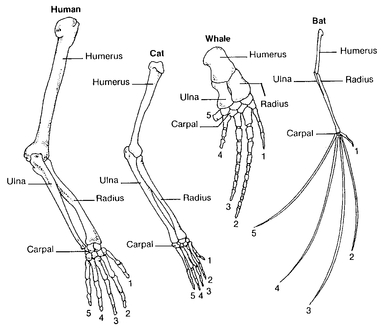
During evolution any characteristic of an organism may be modified. Yet, even in preevolutionary days, some comparative anatomists realized which modified structures were equivalent, such as the wings of birds and the anterior extremities of mammals. Richard Owen, a typological morphologist, said such structures were “homologous” and defined them as “the same organ in different animals under every variety of form and function.” This, of course, left it wide open how to decide when two organs were “the same organ.” This problem was solved by Darwin, who said that certain characteristics of two species were homologous if they were derived by evolution from an equivalent characteristic in the nearest common ancestor of the two species. The anterior extremity of a walking mammal, let us say a dog, was appropriately modified by evolution for such different functions as digging (mole), climbing (monkey), swimming (whale), and flying (bat) (
Fig. 2.7). Furthermore, this mammalian structure is homologous with the pectoral fin of certain fishes.
Adaptive modifications of mammalian forelimbs. The homologous bone elements of human, cat, whale, and bat have been modified by selection to serve their species-specific functions.
Source: Strickberger, Monroe, W.,
Evolution, 1990, Jones and Bartlett, Publishers, Sudbury, MA.
www.jbpub.com. Reprinted with permission.
The claim that certain characteristics in rather distantly related taxa are homologous is at first merely a conjecture. The validity of such an inference must be tested by a series of criteria (Mayr and Ashlock 1991), such as position in relation to neighboring organs, the presence of intermediate stages in related taxa, similarity of ontogeny, existence of intermediate conditions in fossil ancestors, and agreement with evidence provided by other homologies. Homology cannot be proven; it is always inferred.
Homology is due to the partial inheritance of the same genotype from the common ancestor. This is the reason why homology exists not only for structural characters, but for any inheritable feature, such as behavior. Characters that have independently arisen by parallelophyly are nevertheless homologous, because they were produced by the genotype of the common ancestor. Homologous structures may differ considerably in their development. For a review of the different ways in which the term homology has been used, see Butler and Saidel (2000).
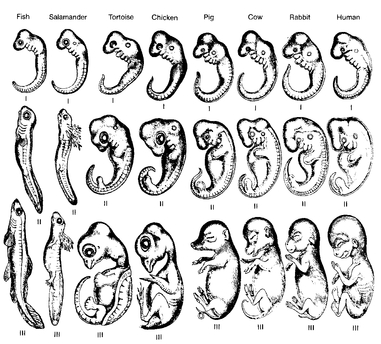
Embryology. Perceptive anatomists observed in the eighteenth century that the embryos of related kinds of animals are often far more similar to each other than are the adult forms. An early human embryo, for instance, is very similar not only to embryos of other mammals (dog, cow, mouse), but in its early stages even to those of reptiles, amphibians, and fishes (
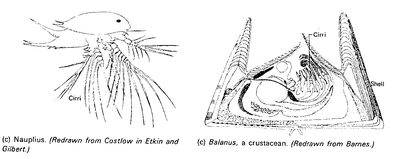
Fig. 2.8). The older the embryo, the more it shows the special characters of the higher taxon to which it belongs. When the adults are highly specialized (for instance, the sessile barnacles among the crustaceans) their free-swimming larvae are still very similar to those of other crustaceans (
Fig. 2.9). Some of Darwin’s opponents asserted that such larval similarities would prove nothing. All development by necessity moves from simple to complex, they said, and the early developmental stages, being simpler, are thus more similar than the later, more complex ones. This is in part true, but embryos and larvae always have some characteristics peculiar to the phyletic lineage to which they belong, and thus reveal their relationship. Furthermore, study of the embryonic stages very often shows how a common ancestral stage gradually diverges in different branches of the ancestral tree. This leads to a far better understanding of the evolutionary pathways.
Haeckel’s figure of 1870 showing the similarity of the development of human embryos to three comparable stages in 7 other kinds of vertebrates. Haekel had fraudulently substituted dog embryos for the human ones, but they were so similar to humans that these (if available) would have made the same point.
Source: Strickberger, Monroe, W.,
Evolution, 1990, Jones and Bartlett, Publishers, Sudbury, MA.
www.jbpub.com. Reprinted with permission.
Recapitulation The term “recapitulation” refers to the appearance and subsequent loss of structures in ontogeny, which in related taxa are retained in the adults. Thus it refers to the loss of an ancestral character in later embryonic stages in one phyletic lineage, but the retention of this character in living species of other lineages derived from the same common ancestor. For instance, embryos of the baleen whales still develop teeth at certain embryonic stages, but these are later reabsorbed and disappear. This appearance and subsequent loss of ancestral characters in succeeding embryonic stages is so striking a phenomenon that it led to a special theory, that of recapitulation. Two drastically different interpretations of these observations were offered by the embryologists.
The free-swimming larval stages of barnacles (cirripedia) are like those of other crustaceans, but the sessile adult stages are so different that the early zoologists considered them to be molluscs.
Source: Kelly, Mahlon G. and McGrath, John C. (1975). Biology: Evolution and Adaptation to the Environment. Houghton Mifflin.
According to Karl Ernst von Baer’s theory, embryos of different organisms in their earliest embryonic stages are so similar to each other that they can not be correctly identified unless one knows their origin. But during development they gradually become more and more similar to their adult stage and, thus, diverge increasingly from the developmental lineage of other organisms. Von Baer summarized this view in his well-known statement: “There is gradually taking place a transition from something homogeneous and general to something heterogeneous and special.” His explanation was widely accepted. However, this claim was clearly in conflict with certain facts of ontogeny. For instance, why should the embryos of birds and mammals develop gill slits, like fish embryos? Gill slits are not a more general condition of the neck region of a terrestrial vertebrate (see
Fig. 2.8). These embryonic gill slits had been discovered in the 1790s, that is, 70 years before the publication of the
Origin. At that time, the only available explanation was the Great Chain of Being, the
scala naturae, in which all organisms were arranged in a series of ever greater “perfection” from the lowest organism to fish, reptiles, and eventually man. This led to the suggestion that the embryo of a higher organism “recapitulated” the ontogeny of organisms lower on the
scala naturae. When evolution was accepted, a new definition offered by Haeckel (1866), “Ontogeny is the recapitulation of phylogeny,” obviously went too far, because at no stage of its development does a mammalian embryo look like an adult fish. Yet, in certain features, as in the gill pouches, the mammalian embryo does indeed recapitulate the ancestral condition. And such cases of recapitulation are by no means rare. The larvae of barnacles are very similar to those of other crustaceans (
Fig. 2.9), and embryonic structures are found in thousands of cases to be indicative of their ancestry, but these same structures are absent in the adult life-forms.
The embryologist could not escape the question of why in these cases ontogeny followed such a roundabout way to reach the adult stage, instead of simply eliminating the embryonic structures that are no longer needed, just as many cave-dwelling species eliminate pigmentation and eyes. The reason was eventually discovered by experimental embryologists, who found that these ancestral structures serve as embryonic “organizers” in the ensuing steps of development. For instance, if one cuts the pronephric duct of an amphibian embryo, there will be no development of the mesonephros. In a similar manner, the removal of the midline stripe of the archenteron roof prevents the development of a notochord and a nervous system. Thus the “useless” pronephros and midline stripe are recapitulated because they have the vital function of being embryonic organizers of later developing structures. This is the same reason why all terrestrial vertebrates (tetrapods) develop gill arches at a certain stage in their ontogeny. These gill-like structures are never used for breathing, but instead are drastically restructured during the later ontogeny and give rise to many structures in the neck region of reptiles, birds, and mammals. The evident explanation is that the genetic developmental program has no way of eliminating the ancestral stages of development and is forced to modify them during the subsequent steps of development in order to make them suitable for the new life-form of the organism. The anlage of the ancestral organ now serves as a somatic program for the ensuing development of the restructured organ (Mayr 1994). What is recapitulated are always particular structures, but never the whole adult form of the ancestor.
Vestigial Structures. Many organisms have structures that are not fully functional or not functional at all. The human caecal appendix is an example, and so are the teeth in baleen whale embryos and the eyes in many cave animals. Such vestigial structures are the remnants of structures that had been fully functional in their ancestors but are now greatly reduced owing to a change in niche utilization. When these structures lose their function owing to a shift in lifestyle, they are no longer protected by natural selection and are gradually deconstructed. They are informative by showing the previous course of evolution.
These three phenomena—embryonic similarities, recapitulation, and vestigial structures—raise insurmountable difficulties for a creationist explanation, but are fully compatible with an evolutionary explanation based on common descent, variation, and selection.
Biogeography. Evolution also helped to explain another great puzzle of biology, namely, the reasons for the geographic distribution of animals and plants. Why are the faunas of Europe and North America on both sides of the North Atlantic so relatively similar, whereas those of Africa and South America on both sides of the South Atlantic are so very different? Why is the fauna of Australia so strikingly different from that of all other continents? Why are there normally no mammals on oceanic islands? Could these seemingly capricious patterns of distribution be explained as the product of creation? Not easily. Darwin, however, showed that the present distribution of animals and plants is due to the history of their dispersal from their original points of origin. The longer that two continents were isolated from each other, the more different their biota became.
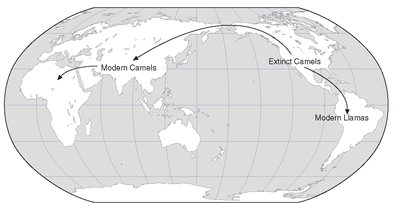
Many organisms have what is called discontinuous distributions. For example, camels and their relatives are found on two different continents: the true camels in Asia and Africa, and their close relatives the llamas in South America. If we believe in continuous evolution there should be a connection between the two now isolated areas; in other words, camels should occur in North America, but they are absent. This situation led to the inference that camels had indeed at one time existed in North America, serving as a connecting link between the Asian and South American camels, but then had become extinct. In due time, this conjecture was indeed confirmed by the discovery in North America of a large fossil fauna of Tertiary camels (
Fig. 2.10). Likewise, the reasons for the similarity of the fauna of Europe and of North America were not fully understood until it was discovered that in the early Tertiary (40 million years ago) there was a broad land connection across the North Atlantic between the two now-separated continents. This permitted an active faunal exchange. By contrast, Africa and South America were separated by continental drift 80 million years ago and their biota diverged greatly during their long isolation. Again and again, puzzling distribution patterns can be explained as the result of common descent and sometimes subsequent extinction. Thus evolution continues to provide the answer to many previously puzzling observations.
The ranges (Asia to South America) of the existing members of the camel family are widely separated. The discovery of a rich fossil fauna of camels in the Tertiary of North America showed that once there was a complete faunal continuity.
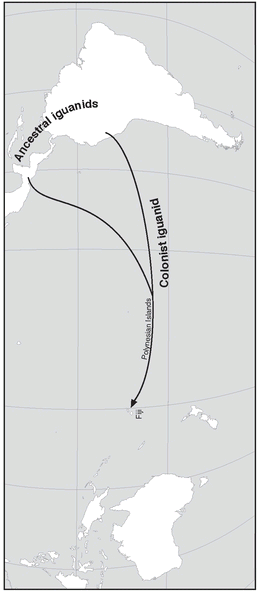
Dispersal Different species can have highly divergent dispersal abilities. More than 100 species of New Guinea birds are so averse to crossing water gaps that they are not found on a single island more than one mile distant from the mainland coast. On the other hand, some species have truly miraculous dispersal facilities. The lizard family Iguanidae is confined to the Americas, except for one genus (with two species) found in Fiji and Tonga (
Fig. 2.11). Since these are endemic species they could not have been brought there by humans. The only possible explanation is that a long time ago they floated there on logs and flotsam carried by ocean currents. It is indeed almost unbelievable that these colonists should have been able to survive such a trip of several thousand miles. Even if at first they had only reached eastern Polynesia, where they were since exterminated by the Polynesians, it still was an extraordinary achievement. How-ever, there is no other explanation, and other cases of long survival on rafts are documented.
Extreme achievement of dispersal. The reptilian family Iguanidae is found only in North and South America, except for the two species of the endemic genus Brachylophus found thousands of kilometers away, in western Polynesia (Fiji, Tonga). It could have reached the Polynesian islands from the Americas only by rafting.
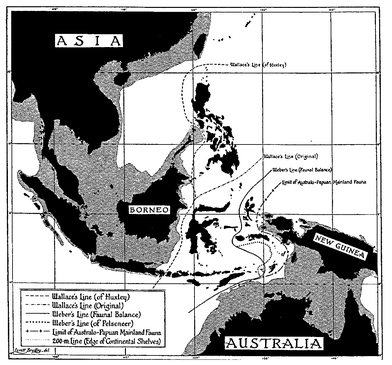
Differences in dispersal ability explain most of the apparent problems of distribution. Mammals (except bats) are notoriously ineffective in crossing water gaps, which is why they are usually absent from oceanic islands. This is also the reason why Wallace’s Line in the Malay archipelago, a line between the Greater Sunda Islands in the west and the Lesser Sunda Islands and Sulawesi in the east, is an important biogeographic border for mammals, but much less so for birds and plants (
Fig. 2.12). Actually this line separates the edge of the Sunda shelf from deep water to the east. Mammals are restricted to the land of the Sunda shelf, while many birds and plant seeds can cross water gaps with considerable ease.
Distributional Gaps The ranges of some taxa are broken by a gap in which the taxon does not occur. There are two different ways by which such gaps may originate. The North American gap in the range of the camel families, as we saw above, was caused by their extinction there. Originally they ranged continuously from Asia to South America. This is referred to as the vicariance hypothesis. Most discontinuities on continents seem to be such remnants of previously continuous ranges. Many arctic species, for instance, were able at the height of the Pleistocene glaciation to colonize the Alps and Rocky Mountains, but are now left after the retreat of the ice as montane relics, widely separated from the arctic populations of their species.
A second type of range discontinuity is primary. It originates when members of a species establish a founder population beyond the present species border after dispersing across unsuitable terrain (water, mountains, or an unsuitable vegetation area). Such dispersal discontinuities are particularly characteristic for areas with insular distributions. The taxa of the Galapagos Islands never had a continuous range with South America, their source area. All species of this insular biota reached the Galapagos Islands by crossing the 600-mile water gap between the two areas. For a creationist there is no rational explanation for distributional irregularities, but they are completely compatible with a historical evolutionary explanation.
Molecular Evidence. It was one of the unexpected happy discoveries of molecular biology that molecules evolve just the same as do so-matic structures. On the whole, the more closely related two organisms are, the more similar are their respective molecules. In many cases when there was considerable doubt as to the relationship of two organisms because the morphological evidence was ambiguous, a study of their molecules revealed the real relationship. As a result, molecular biology became one of the most important sources of information on phylogenetic relationships.
The contact zone between the Indo-Malayan and the Australo-Papuan faunas. The shaded area in the west is the Asian (Sunda) shelf, and in the east lies the Australian (Sahul) shelf. The area between the two shelves, never connected by a land bridge, is referred to as Wallacea. The real border (line of balance) between the Asian and the Australian faunas is Weber’s Line.
Source: Mayr, Ernst. (1944). Quarterly Review of Biology 19(1): 1–14.
Genes, or more precisely the structure of the molecules of which they consist, undergo an evolutionary change just as do macroscopic structures. By comparing homologous genes and other homologous molecules of different organisms, one can determine the degree of their similarity. However, different kinds of molecules have different rates of evolutionary change. Some change very rapidly, like the fib-rinopeptides , and others change very slowly, like the histones. Even though the lineages of man and chimpanzee separated at least 6 million years ago, the highly complex molecules of the hemoglobins of these two species are still virtually identical. What is gratifying is the fact that when a phylogeny based on morphological or behavioral characteristics is established, it is usually found to be essentially the same as a phylogeny based exclusively on molecular characteristics.
A comparison of the results of both sources of evidence is most helpful in all cases in which the analysis of morphology has led to ambiguous results. Such cases can now be tested against the molecular phylogeny of these taxa. Many different genes are available for such an analysis. In some cases it is the molecular evidence that reflects the phylogeny more correctly than the morphology. To mention just two cases from the recent literature, molecular analysis showed that the golden mole of South Africa and the tenrecs of Madagascar are quite unrelated to the Insectivora, among which these animals were traditionally classified on the basis of morphological evidence. Likewise, the Pogonophora and the Echiura, always considered independent phyla, were shown to be more closely related to certain families of polychaetes than these are to other polychaetes. The extremely close relationship of man to the chimpanzee and to the other anthropoid apes is as convincingly documented by molecular as by structural characters.
The Importance of Molecular Analysis One of the most important contributions made by molecular biology to the understanding of evolution is the discovery that the basic molecular framework of all organisms is very old. The particular structures acquired by the phyla of animals, fungi, and plants that enable them to survive and prosper in the particular niche or adaptive zone that they occupy are, on the whole, considerably more recent. So we can use these adaptive structures to classify animals, fungi, and plants, but they tell us little about how the fungi are related to animals or plants. For instance, fungi traditionally were always considered to be related to plants and their study was the job of botany departments. To be sure, it was puzzling that their cell walls consisted of chitin, a substance supplying all the hard parts of insects but not found anywhere in plants. This was simply treated as one of the typical exceptions that are so common in biology. But molecular analysis finally showed that in much of their basic chemistry fungi are quite closely related to the Animalia.
The gradual straightening out of the chaos of the 50–80 phyla of “protists” is also a great achievement of molecular biology (and of the study of membranes and other fine structures), after a study of the traditional morphological characters had failed to produce clarity. The successful arrangement of the angiosperms into groups of related orders and families was likewise largely accomplished by the application of molecular methods. Perhaps the greatest virtue of the molecular approach is that there are so many potential characters to study. When one particular gene leads to ambiguous results, one can in principle shift to any of thousands of other genes to test a suspected connection.
The Molecular Clock In the absence of an adequate fossil record, for a long time it was essentially impossible to determine the geological age of many evolutionary lineages. However, Zuckerkandl and Pauling (1962) showed that many, perhaps most, molecules have a rather constant rate of change over time. Such molecules can serve as a molecular clock. Well-dated fossils with modern descendants provide us with a yardstick for calibrating a given molecular clock. It was by the molecular clock method that the branching point between chimpanzee and man was shown to be as recent as 5–8 million years ago, rather than 14–16 million years, as had been previously generally accepted.
However, the molecular clock method must be applied with caution because molecular clocks are not nearly as constant as often believed. Not only do different molecules have different rates of change, but a particular molecule may vary its rate over time. These represent cases of mosaic evolution. In cases of discrepancy it is always advisable to determine also the rate of change of a different molecule and to try to find another suitable fossil.
The Evolution of the Genotype as a Whole With the help of greatly improved methods it is now possible to determine the essentially complete DNA sequence of the entire genome of a whole organism. This was first done for several bacteria (eubacteria and archaebacteria), including
Escherichia coli, then for yeast
(Saccharomyces), a plant
(Arabidopsis), and some animals, such as the roundworm (nematode)
Caenorhabditis and the fruit fly
Drosophila (
Table 2.1). The completion of the essential sequencing of the human genome was celebrated in June 2000. The field dealing with the molecular structure of the genome is called
genomics.
SOURCE: From Maynard Smith and Szathmary (1995), p. 5.
| Organism | Genome Size (base pairs x 109) | Coding DNA |
|---|
| Bacterium (Escherichia coli) | 0.004 | 100 |
| Yeast (Saccharomyces) | 0.009 | 70 |
| Nematode (Caenorhabditis) | 0.09 | 25 |
| Fruit fly (Drosophila) | 0.18 | 33 |
| Newt (Triturus) | 19.0 | 1.5–4.5 |
| Human (Homo sapies) | 3.5 | 9–27 |
| Lungfish (Protopterus) | 140.0 | 0.4–1.2 |
| Flowering plant (Arabidopsis) | 0.2 | 31 |
| Flowering plant (Fritillaria) | 130.0 | 0.02 |
These sequences are now the material for the most fascinating comparative studies. Although genes (base pair sequences) evolve, the function of a gene sets severe limits on the amount of change. In other words, the basic structure of a gene is usually preserved over many millions of years and this permits the study of the phylogeny of each gene. The most astonishing result of these studies is that some basic genes of higher organisms can be traced all the way back to homologous genes in bacteria. Many genes in the yeast Saccharomyces, the worm Caenorhabditis, and the fly Drosophila can be traced back to the same ancestral gene. Such a gene may not have exactly the same function in all the organisms in which it occurs, but it will have a similar or equivalent function.
The Origin of New Genes Bacteria and even the oldest eukaryotes (protists) have a rather small genome (see Box 3.1). This raises the question: By what process is a new gene produced? This occurs, most frequently, by the doubling of an existing gene and its insertion in the chromosome in tandem next to the parental gene. In due time the new gene may adopt a new function and the ancestral gene with its traditional function will then be referred to as the orthologous gene. It is through orthologous genes that the phylogeny of genes is traced. The derived gene, coexisting with the ancestral gene, is called paralogous. Evolutionary diversification is, to a large extent, effected by the production of paralogous genes. The doubling sometimes affects not merely a single gene, but a whole chromosome set or even an entire genome.
CONCLUSIONS
As we have seen, whatever aspect of biology is studied, it provides irrefutable evidence in support of evolution. As the famous geneticist T. Dobzhansky has said so rightly, “Nothing in biology makes sense, except in the light of evolution.” Indeed, there is no other natural explanation than evolution for the facts presented in this chapter.
Perhaps nowhere has the evolutionary approach produced more clarity and understanding than in the ordering of the bewildering diversity of living organisms. As a result we can now describe in remarkable detail the gradual rise of higher organisms (plants and animals) from the simplest forms of life. The next chapter is devoted to a presentation of this ascent of life.