
We’re Not Alone: Forest Die-Off Risks Around the Globe Craig D. Allen, PhD
Craig Allen conducts research on the ecology and environmental history of Western landscapes. He oversees the USGS Jemez Mountains Field Station, based at Bandelier National Monument in New Mexico. He was the lead author of the 2010 paper, “A Global Overview of Drought and Heat-induced Tree Mortality Reveals Emerging Climate Change Risks for Forests”, published in the journal “Forest Ecology and Management.”
By providing a global overview of this topic, I’m going to talk about why we are not alone here in Colorado in terms of forest die-off. The two main things this talk will do are illustrate worldwide patterns of drought and heat-induced tree mortality and discuss some of the scientific uncertainties that currently inhibit us from being able to predict future risks of climate-induced forest die-off.
Certainly hot and dry conditions in the past have been observed to drive episodes of forest health problems and tree mortality, illustrating that the causal linkages between severe drought and tree mortality are not unusual. They are really nothing new or surprising; but, at the same time, there are some places on this planet where trees are growing better recently than we’ve ever measured before. So it is certainly reasonable to ask: Is there is any reason for concern about the broad-scale forest die-off events we see today? Although in earlier talks today we’ve heard about particular examples of extensive climate-related tree mortality here in western North America, perhaps, at the global scale, this is being balanced out by more productive growth elsewhere.
Well, there is one particular climate trend occurring lately that is different than any other in the past thousand years: a significant warming of the world. An immense amount of research has been done on the topic of climate change, and a substantial scientific consensus exists that the planet has been warming rapidly in recent decades. This warming trend is projected to continue, along with a greater range in extreme climatic events such as severe droughts (IPCC 2007). Such climate changes could be driving increased tree water stress in many areas and thereby increasing risks of stress-related tree mortality. If so, we might expect to see patterns of greater tree mortality and forest die-off emerging in at least some forests.
In recent years, I have been working collaboratively with colleagues in the U.S. and abroad to document examples of drought-driven tree mortality on all forested continents and to assess whether any new trends were beginning to emerge. One result of these efforts is a paper, including twenty co-authors from around the world, published a year ago (Allen, et al. 2010). It is the first recent global overview of this phenomenon. One of our core findings is that there is no major forest type on the planet that is immune to the effects of drought and heat-induced tree mortality, with significant forest die-off observed from tropical rainforests of the Amazon Basin to boreal forests in Siberia and everything in between.
The map in Figure 1 shows the main climatic limitations to vegetation primary productivity. The bluish colors are where it’s limited by temperature. The reddish color is where it’s limited by water availability (note the deserts of the world). The greenish areas generally represent the tropics, where water and growing season length are usually adequate, but sunlight is limiting. So a broad range of different climate conditions limit vegetation productivity across the Earth. The superimposed dots indicate places we documented where there have been significant drought-related forest die-off events in the last 25 years. A key point is that drought and heat-driven tree mortality occurs widely across this whole environmental spectrum, and it can happen even in very wet places like Borneo and the heart of the Amazon Basin.
The 2010 global overview paper has satellite image maps of each of the six forested continents with numbers marking documented drought-induced mortality areas. Here, for example, are the maps for North America and Europe (see Figures 2, 3). The map numbers tie to continent-by-continent tables that list details and references for each mapped mortality event. The tables give specifics of what happened at any of the numbered map locations (e.g., type of forest and tree species involved, dates of tree mortality, size of affected area, etc.).

Figure 1: White dots indicate documented localities with forest mortality related to climatic stress from drought and high temperatures. Background map shows potential limits to vegetation net primary production (Boisvenue and Running 2006). Only the general areas documented in the tables are shown - many additional localities are mapped on the continental-scale maps. Drought and heat-driven forest mortality often is documented in relatively dry regions (~red/orange/pink), but also occurs outside these regions. From Allen et al. 2010.
Also included here are a few photos illustrating examples of drought-induced tree mortality from around the world. The dead trees in Figure 4 are Atlas cedar (Cedrus atlantica) in Belezma National Park, Algeria, exhibiting wholesale die-off from severe drought between 2000 and 2008. Essentially the Sahara is moving north toward the Mediterranean Sea, and most of what is still green here is an evergreen oak (Quercus ilex) that remained alive after the taller cedar had died.
Note that in the global overview paper we only included recent examples that are securely documented in the (mostly English language) scientific literature, although apparently there are many other examples of drought-induced tree mortality known to local foresters and researchers around the world that currently are not reported in that particular way. For example, in Australia we were able to document recent drought-induced tree mortality only from Queensland in the northeast, where eucalyptus and acacia trees died across many millions of acres (see Figure 5); however, through interactions with various colleagues, I know of examples from southern and western Australia that haven’t been written up yet in the scientific literature. The pictured eucalyptus and acacia species in Figure 5 are evergreen, so when they look leafless in the photo, they’re dead.
In Asia, drought-related tree mortality has been occurring along the forest-steppe ecotone of inner Asia, and millions of acres in southern Siberia are being affected by various kinds of drought and heat-related tree mortality. Other Asian examples of forest die-off extend from Turkey in the west across to China and Korea in the east, and southward down into India and the tropics of Borneo. Whereas El Niño climate events tend to bring wet winter weather conditions to the southwestern U.S., El Niño conditions often bring drought in the tropics. The strong El Niño in 1998 left a signature of tropical tree mortality from Southeast Asia to the Americas. In South America, the 1998 El Niño even caused significant mortality of southern beech trees (Nothofagus dombeyi) in the temperate forests of Patagonia in northern Argentina, while severe droughts in 2005 and 2010 have driven substantial tree mortality in the Amazon Basin.

Figure 2: Satellite map of North America, with documented drought-induced mortality localities indicated with numbers, tied to Table A.5 references. Top photo: Aerial view showing severe mortality of aspen (Populus tremuloides) in the parkland zone of Alberta, Canada; 2004, by Michael Michaelian. Lower photo: Pinus ponderosa die-off, Jemez Mountains, New Mexico, USA; 2006, by Craig D. Allen. From Allen et al. 2010.
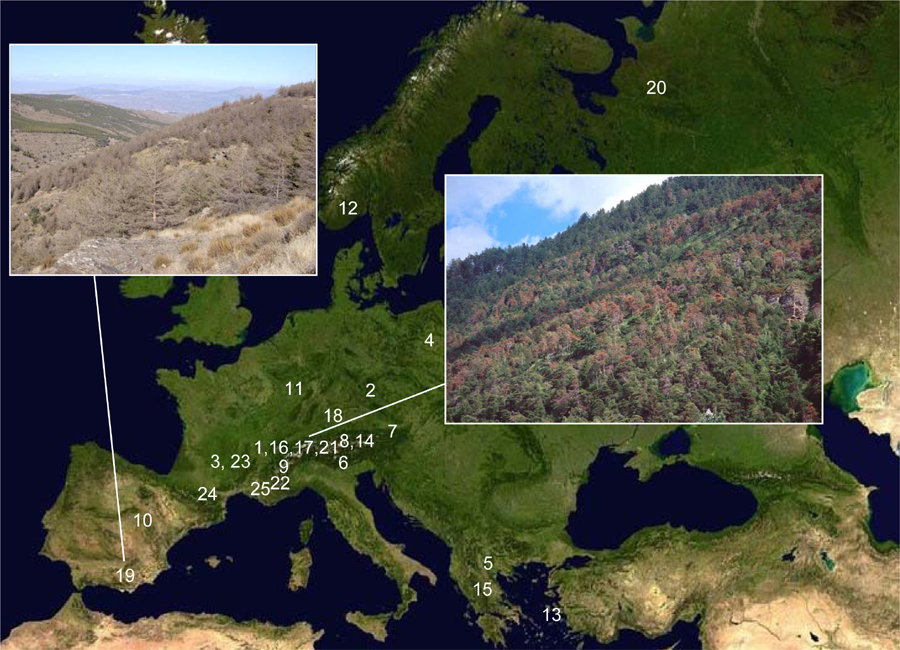
Figure 3: Satellite map of Europe, with documented drought-induced mortality areas indicated with numbers, tied to Table A.4 references. R photo: Pinus sylvestris mortality, Valais, Switzerland; 1999, by Beat Wermelinger. L photo: Pinus sylvestris die-off, Sierra de los Filabres, Spain; 2006, by Rafael Navarro-Cerrillo. From Allen et al. 2010.

Figure 4: Atlas cedar mortality, with surviving understory of Quercus ilex, in Belezma National Park (Aurès Région), Algeria. Photo by Cherif Ghazi in 2007, courtesy of Haroun Chenchouni.

Figure 5: Satellite map of Australasia, with documented drought-induced mortality areas indicated with numbers, tied to Table A.3 references. R photo: Die-off of mulga, Acacia aneura, the dominant tree across large areas of semi-arid Australia; 2007, by Rod Fensham. L photo: Eucalyptus xanthoclada mortality in Queensland, northeastern Australia; 1996, by Rod Fensham. From Allen et al. 2010.
In Europe, a lot of documentation exists from some regions, particularly around the Mediterranean where much good work is being done by strong research groups in Spain, France and Switzerland. Drought-related mortality of Scots pine (Pinus sylvestris) is observed in all of these countries (Figure 3), from the Pyrenees and Provence to the Alps, in both native forests and plantations. Spain alone has more than ten million acres of reforestations with various pine species, which are emblematic of similar landscapes all around the arc of the Mediterranean, where, in the past century, pine plantations have been established across extensive areas that have been desertified by intensive land uses over the past three thousand years. In the past two decades, we see signs of significant water stress, growth declines, and tree mortality emerging in these often-dense pine plantations, particularly associated with drought events and warmer temperatures.

Figure 6: In summer 2002, pinyon (Pinus edulis) began dying en masse from drought stress and an associated bark beetle outbreak. Jemez Mts. near Los Alamos, October 2002. Photo: Craig D. Allen, USGS
Finally, in preceding talks, we’ve heard about examples of recent big forest die-offs in western North America, including sudden aspen decline here in Colorado and the huge lodgepole pine (Pinus contorta) mortality event in British Columbia. In addition, one of Werner Kurz’s colleagues just published documentation of aspen dieback and mortality affecting more than two million acres in Alberta and Saskatchewan, associated with warm drought in the 2000’s (Michaelian et al. 2010). Similarly, severe and warm drought affected the southwestern U.S. from 2000 to 2004, driving significantly higher levels of mortality regionally for most tree species. For some species, like ponderosa pine (Pinus ponderosa) and Colorado piñon (Pinus edulis), substantial mortality and even wholesale die-off was mapped across millions of acres (Breshears, et al. 2005).

Figure 7: After the needles drop, grey skeletons of pinyon remain, with surviving Juniperus monosperma still green. Jemez Mts., May 2004. Photo: Craig D. Allen, USGS.
For example, Figure 6 shows the Jemez Mountains near Los Alamos and Santa Fe, New Mexico in the fall of 2002 —all of those orange-colored trees are dying piñon. A repeat photograph of that exact same view 19 months later (see Figure 7) shows the skeletons of the dead piñon trees as numerous grey stems; the small green trees still present are the more drought-tolerant one-seed junipers (Juniperus monosperma). This die-off of the dominant piñon trees has caused a major alteration of the ecology of this landscape. Although a cohort of young piñon seedlings and saplings survived and continue to grow on this site, it will be literally decades before they will be mature enough to become seed sources for future piñon regeneration.
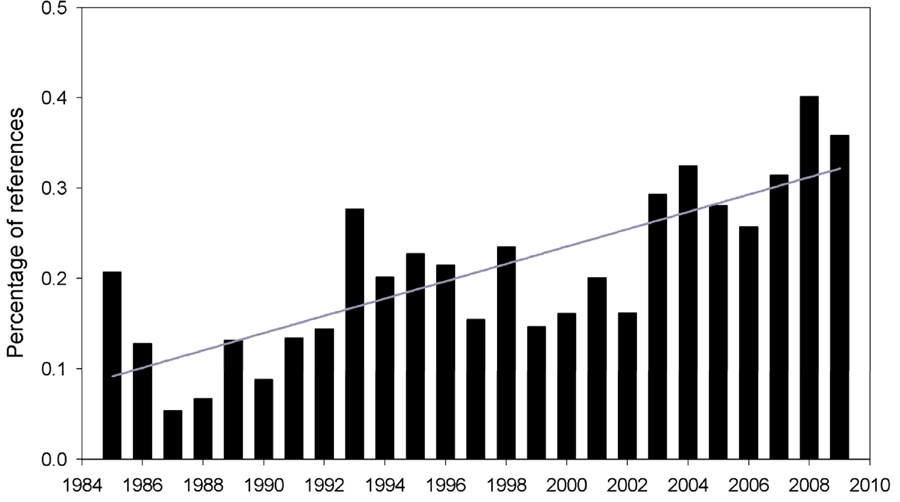
Figure 8: ISI Web of Science search of the trend in published reports of climate-related forest mortality in scientific literature, the scientific for the literature, 1985-2009. Plotted bars show the percent of references using the topic words “forest AND mortality AND drought”, relative to all “forest” references. Line represents the linear regression model fitted to the data (R2 = 0.61; F = 35.73; p < 0.001).
Now I’d like to briefly discuss some of the scientific uncertainties associated with climate-induced tree mortality. Three big uncertainties exist. First, we see that there are examples of significant of forest mortality being reported from many parts of the world recently, but maybe this is nothing unusual. Are new trends emerging globally? In the past two years there has been new research published showing increasing drought and heat-related forest stress and mortality in portions of southern Europe (Carnicer et al. 2010) and the Amazon Basis (Phillips et al. 2009, Lewis et al. 2010). And certainly in western North America, background tree mortality rates have increased recently (van Mantgem et al. 2009), and the magnitude and huge scale of climate-stress and bark beetle-related episodes of extensive forest die-off in western North America over the past two decades are apparently unprecedented in the historic record of the past 100-plus years (Raffa et al. 2008).
But what about at the global scale? Figure 8 displays patterns in the scientific literature on this topic from 1984 to the present, as a percentage of all forest related scientific references. We see that there is a very strong trend of an increasing percentage of references that are looking at forest mortality associated with drought. While this does not prove that more forest mortality is occurring on the ground, it does mean that more scientific interest and attention exists on this topic.
For example, a paper came out last summer that used satellite imagery across the planet to look at terrestrial vegetation net primary productivity (NPP), which is basically land plant growth averaged for the past 10 years across the Earth’s land surface (Zhao et al. 2010). The authors found that overall NPP had declined globally over that 10-year period because of drought stress across large parts of the earth, which was contrary to many projections that, with increasing CO2, global NPP would increase. While there are hints that worrisome trends in global forest health may be emerging, we cannot yet determine definitively if worldwide tree mortality rates and forest die-off events are rising, because we lack long-term measurements of forest health trends at a planetary scale. Accurate documentation of global forest mortality patterns and trends requires the establishment of a worldwide monitoring program that will likely combine satellite remote-sensing approaches and ground-based measurements in a coordinated and consistent manner.
A second big uncertainty comes from the difficulties in determining the physiological stress thresholds of tree mortality well enough to rigorously predict mortality in computer models that link climate to tree growth and death. Basically, as ecologists, we know how to grow trees fairly well in computer models, but we don’t know yet how to kill them reliably in quantitative, process-based ways (McDowell et al. 2011). Trees have a variety of ways to buffer themselves from stress, and they commonly die gradually or incrementally, challenging our ability to precisely determine how or even when they died.
For only a few tree species do we know in a controlled laboratory situation how much water stress an individual tree can take before it dies. In uncontrolled real landscapes, many interacting factors further complicate understanding and modeling tree mortality responses to drought and heat stress, ranging from diversity in site conditions (slope, aspect, soils, variable temperature and precipitation patterns) to competition with other plants and feeding impacts from animals. Actually, in many cases we are unsure whether drought-stressed trees die from direct desiccation effects, indirect starvation as they can not photosynthesize carbohydrates adequately when severely water-stressed, from the primary or secondary effects of attacking insects and diseases (McDowell et al. 2011), or from some combination of these processes.
Scientists are now just beginning the experiments needed to better document these physiological responses and mortality processes to determine how even a few specific tree species respond to particular kinds of stress. Because we currently lack the knowledge needed to accurately predict major episodes of tree mortality, we continue to be surprised when a big forest die-off event occurs. Another reason why we’re taken by surprise when extensive tree mortality happens is because it commonly manifests as a non-linear threshold response to a variable climate. A drought-stressed forest may look relatively healthy for some extended period of time, until a tipping point of some sort is reached. Then trees can suddenly begin to die en masse.
Until we better understand the various non-linear threshold processes that underlie tree mortality, projections of future tree mortality risks in response to climate variability and change will remain quite uncertain. But given that the best current global climate models show that by 2100 many of the world’s climate patterns will shift somewhat, it is reasonable to expect these climate changes to drive substantial shifts in the distribution of most major ecosystems, including forests. And from the examples of drought and heat-induced forest die-off recently documented around the world, one manifestation of the expected ecosystem shifts is likely to be substantial climate-induced tree mortality in at least some regions.

Figure 9: Young Allen’s in an alligator-bark juniper tree, Bandelier Natonal Monument, New Mexico. Photo: Craig D. Allen.
Finally, a third major scientific uncertainty I want to mention is our limited knowledge about diverse feedbacks between forests and the overall Earth system (Lenton et al. 2008, Adams et al. 2010). Forests cover about 30 percent of the world’s land surface and provide many important ecosystem services, including capturing and sequestering about one-quarter of the excess carbon that humanity releases into the atmosphere every year through tree growth. But there are risks that this huge pool of forest-stored carbon might not be stable if growing climate stress drives widespread tree mortality, and global forests could release a lot of this stored carbon back to the atmosphere, potentially becoming a further positive feedback for greenhouse gas concentrations and associated climate warming.
Forests are important in modulating the water cycle at scales from local to global, and forests partly drive how much sunlight is absorbed at the Earth’s surface, and how much is reflected, affecting planetary heat fluxes. Any major changes in the world’s forests will have big feedbacks to the whole earth climate system. Another related topic area that we don’t know enough about includes the feedbacks and interactions among climate-induced forest stress and other tree-killing processes like fires and insect outbreaks. Given these various uncertainties, additional monitoring of global forest health and new research are needed to improve scientific certainty regarding risks of future climate-induced tree mortality.
In conclusion, let me say that forests are extremely important to people as well as to the planet. The earth is graced with a great diversity of forests that are essential to local and global ecologies and economies. Forests and even unique individual trees also are widely revered by societies for many less tangible reasons, including the timeless sense of continuity and enduring character they provide to particular regions and local landscapes. As mature trees typically can live much longer than any person, so forests often seem immutable, essentially permanent, with any natural changes so gradual as to be imperceptible. But we see that climate stress can cause big forest die-off events to emerge in just months or a few years. Because people care about forests at many levels for many reasons (see Figure 9), a focus on forests is one good approach to get people to pay attention, sooner rather than later, to the actions we all need to take, individually and collectively, to ensure the ongoing continuity of healthy forests.