6
Seeing movement
Detection of movement is essential for survival of all but the very simplest creatures. Moving objects are likely to be dangerous prey, or potential food, or a mate. They generally demand action of some kind, while stationary objects may be ignored with safety. Indeed, it is only eyes quite high up the evolutionary scale that produce signals in the absence of movement.
We can experience something of the long evolutionary development of vision, from the simplest creatures to ourselves, embalmed in the human eye.
The edges of our retinas are sensitive only to movement. You can see this by getting someone to wave an object around at the side of your visual field where only the edge of the retina is stimulated. Movement is seen, but it is impossible to identify the object, and there is no colour. When movement stops the object becomes invisible. This is as close as we can come to experiencing primitive vision. The extreme edge of the retina is even more primitive: when it is stimulated by movement we experience nothing; but a reflex is initiated, rotating the eye to bring the moving object into central vision, bringing our highly developed foveal region into play for identifying the object.
Eyes like ours which move in the head give information of movement into two distinct ways. When the eye remains stationary, the image of a moving object will run sequentially across many receptors, giving motion signals from the retinas. But when the eyes follow a moving object, the images remain essentially stationary on the retinas—so they cannot signal movement though we still see movement. This is so even when there is no background to sweep across the retina as the eyes move. This effect can be demonstrated with a simple experiment. Ask someone to wave a dim pencil torch slowly side to side in a dark room, and follow it with your eyes. You will see the movement of the light even though there is no image moving across your retinas. So evidently the rotation of the eyes in the head can give perception of object motion in the absence of movement signals from the retinas. This works because rotations of the eyes are signalled to the brain—in an unexpected way, as we will see.
There are, then, two essentially different visual systems for detecting movement: the image–retina system, and the eye–head system (Figure 6.1).
Image–retina movement
Recording electrical activity from the eyes reveals that there are various kinds of receptors, almost all signalling only changes of illumination; very few fire from continuous steady light. Some receptors signal when a light is switched on, others when it is switched off, while others again signal when it is switched on or off. These are named ‘on’, ‘off’, and ‘on–off’ receptors, respectively. It seems that those receptors responding only to changes of illumination are responsible for signalling movement, and that all eyes are primarily detectors of movement.
By placing electrodes in the retinas of frogs’ eyes, it has been found that analysis of the receptor activity for signalling movement takes place in the frog’s retina. Horace Barlow, at Cambridge, discovered retinal ‘bug detectors’ in the frog’s retina, which elicit reflex tongue fly-catching, faster than signals processed by the brain. Feature detectors were described in a paper charmingly called ‘What the frog’s eye tells the frog’s brain’, by J. Y. Lettvin and colleagues in 1959, at the Massachusetts Institute of Technology. They found:
(1) fibres responding only to sharply defined boundaries;
(2) fibres responding only to changes in the distribution of light;
(3) fibres responding only to a general dimming of illumination, such as might be caused by the shadow of a bird of prey.
The physiological discovery that movement is specially coded as neural signals in the retina, or in the visual projection areas of the brain, is important in many respects. In particular, it shows that speed is signalled without involving a clock, or estimate of time. It is, however, sometimes assumed that neural systems giving perception of velocity must refer to an internal biological clock of some kind. This assumption arises from the fact that velocity is defined in physics by the time taken for something to travel a given distance, (v = d/t). But the speedometer of a car has no clock associated with it. A clock is, indeed, needed for calibrating such an instrument in the first place, but once calibrated, it will give velocity measures without the use of a clock. The same is evidently true of the eyes. The image running across the retina sequentially fires receptors in its path, and the faster the image travels (up to a limit) the greater the velocity signal this will give—with no need for a clock. Although analogies with other velocity detectors, such as speedometers, show that velocity could be perceived without reference to a clock, they do not tell us precisely how the visual movement system works. A detailed model has been suggested for the compound eye of flies, and is the basis for a system in aircraft that detects drift due to wind blowing them off course. This movement detector was developed by biological evolution millions of years ago, and has now been discovered by applying electronics, to be useful for technology. This is a nice example of backwards and forwards bio-engineering. The human eye has different mechanisms for very slow and for fast movement, and different mechanisms for short-and long-range movements of textures, or random dots. These make engineering sense.
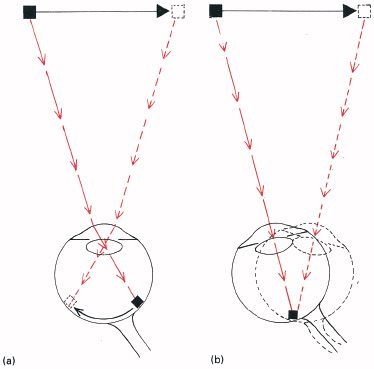
6.1 Two movement systems. (a) The image–retina system: the image of a moving object runs along the retina when the eyes are held still, giving information on movement through sequential firing of the receptors in its path. (b) The eye/head system: when the eye follows a moving object, the image remains stationary upon the retina, but we still see the movement. It is signalled from the commands to move the eyes. The two systems can sometimes disagree, to give paradoxical illusions of movement.
It is a strange thought that we are engineering devices; the deep question is whether this extends right into brain and mind. The Oxford philosopher, Gilbert Ryle, famously spoke of the ‘Ghost in the machine’ Should we turn this around—to question the machine in the ghost?
Eye–head movement
The neural system giving perception of movement by shifting of images across the retina must be very different from the system for detecting movement by rotation of the eyes in the head. Somehow, the fact that the eye is being moved is signalled to the brain and used to indicate movements of external objects. This is demonstrated with the moving torch experiment described above, for in that situation there is essentially no movement across the retina and yet the movement is seen (Figure 6.1(b)).
The most obvious kind of eye–head signal would be from the eye muscles; when they are stretched signals would be fed back to the brain, indicating movement of the eyes and so of objects followed by the eyes. This would be the engineer’s normal solution, but is it Nature’s? We find the answer when we look at what may seem a different question:
Why does the world remain stable when we move our eyes?
The retinal images run across the receptors whenever we move our eyes, and yet we do not experience movement: the world does not usually spin round when we move our eyes. This does happen when a cine or video camera is panned round.
We have seen that there are two neural systems for signalling movement: the image–retina and the eye–head systems. It seems that during normal eye movements these signals cancel each other out, to give stability to the visual world. This was discussed by the physiologist who did most to unravel spinal reflexes, Sir Charles Sherrington (1857–1952), and by Helmholtz (Figure 6.2); but they had different ideas as to how it comes about—especially about how the eye–head system conveys its information of movement. Sherrington’s theory is known as the inflow theory and Helmholtz’s the outflow theory (Figure 6.3). Sherrington thought that signals from the eye muscles were fed back into the brain when the eye moves, to cancel the movement signals from the retina. But neural signals from the eye muscles would take longer to arrive than signals from the retinas, so we might expect a sickening jolt, just after movements of the eyes, before the inflow signals reach the brain to cancel the image–retina signals.
Helmholtz had a very different idea. He thought that retinal movement signals are cancelled not by ‘inflow’ signals from the eye muscles, but by central ‘outflow’ signals from the brain, commanding the eyes to move.
The issue can be decided by very simple experiments, which you can try yourself. Try pushing one eye gently with a finger, having closed the other eye by holding a hand over it. When the eye is rotated passively in this way, the world will be seen to swing round, in the opposite direction to the movements of the eye. Evidently, stability does not hold for passive eye movements, though it does for the normal voluntary eye movements. Since the world swings round against the direction of the passive eye movements, it is evident that the image–retina system still works; it is the eye–head system which is not operating. Why should the eye–head system work only for voluntary, not for passive, eye movements? Sherrington thought that the eye muscles send signals from stretch receptors, in the same way as stretch receptors are known to give feedback signals from the muscles which move the limbs. But although the six eye muscles do have stretch receptors, the eye–head system does not work this way. As we have just said, the stretch receptors would deliver signals too late to cancel the image–retina signals, without a sickening delay, and they are not accurate.
We may stop all retinal movement signals and see what happens during passive or active movements of the eyes. This is easily done by staring at a bright light (or a photographic flash) to get an after-image—like a photograph stuck on the retina. This will move precisely with the eye, so it will not give any image–retina movement signals. If you observe the after-image in darkness (to avoid a background) you will find that, if the eyes are pushed with the finger to move passively, the after-image does not move. This is very strong evidence against the inflow theory. There will be signals from stretch receptors, but they do not produce sensations of motion.
6.2 Hermann von Helmholtz (1821–94)—physiologist, physicist, psychologist, philosopher—is the greatest figure in the experimental study of vision. His Physiological optics is the bedrock of the subject.
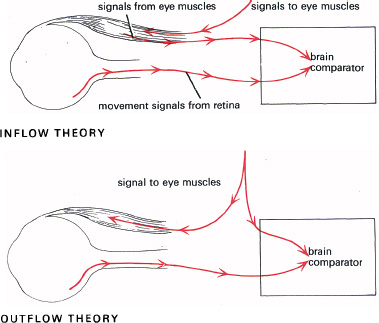
6.3 Why does the world remain stable when we move our eyes? The inflow theory suggests that movement signals from the retinas (image–retina system) are cancelled by afferent signals from stretch receptors of the eye muscles. The outflow theory suggests that the retinal movement signals are cancelled by the efferent command signals to move the eyes (eye–head system), through an internal monitoring loop. The evidence favours the outflow theory.
However, if you move your eyes voluntarily, with the after-image viewed in darkness, you will see the after-image moving with the eyes. Helmholtz explained this by supposing that it is not signals from the eye muscles which are involved, but commands to move the eyes. This suggests that the command signals to move the eyes are monitored by an internal loop (Figure 6.3), to cancel the image–retina signals. When these are absent (as when an after-image is viewed in darkness) the world swings round with the eyes because the command signals are not cancelled by retinal motion signals. Passive movements of the eye give no movement of the after-image because in these circumstances neither system gives a movement signal.
In clinical cases, where something is wrong with patients eye muscles or their nerve supply, the world swings round for them when they try to move their eyes—moving in the direction their eyes should have moved. This also occurs if the muscles are prevented from functioning by curare, the South American arrow poison which blocks neural signals to muscles. The Austrian physicist Ernst Mach (1838–1916), bunged up his eyes with putty, so they could not move, to get the same result. (Not to be repeated!)
So the eye–head system does not work by detecting actual movement of the eyes, but from commands to move them. It works even when the eyes do not obey the commands. It is surprising that command signals can give rise to perception of movement; we usually think of movement perception as always coming from the eyes, not from a source deep in the brain controlling them.
Why should such a peculiar system have evolved? It is even more surprising when we find that there are in fact stretch receptors in the eye muscles. It seems that an inflow or feedback system would be too slow: by the time a feedback signal got back to the brain to cancel the retinal movement signal, it would be too late. In fact, because the signals from the retina have further to travel to reach the brain, the command signal could arrive too soon. Evidently it is delayed to match the small inevitable retinal delay (the retinal action time) so it does not arrive before the motion signal from the retina. We can literally see this in the movement of the after-image with voluntary eye movements. Whenever the eye is moved, the after-image takes a little time to catch up. Can one imagine a more beautiful system?
The case of the wandering light
You might like to try the following experiment. The apparatus is a small dim light (such as an LED) placed at the far end of a completely dark room. If you look at the light for more than a few seconds, it will wander around, in a curious erratic manner, sometimes swooping in one direction, and sometimes oscillating gently to and fro. Its movement may seem paradoxical: it may appear to move, yet not change its position.
The wandering light, is known as the autokinetic phenomenon. It has been the subject of a great deal of discussion and experimental work. A dozen theories have been advanced to explain it, and it has been used as an index of suggestibility and group interaction by social psychologists. The explanations are extraordinarily diverse.
One theory is that small particles drifting about in the aqueous humour, in the front chamber of the eye, may be dimly seen under these conditions. This theory then proposes that the spot of light, rather than the particles, seems to move just as the moon seems to scud through the night sky when clouds are driven by the wind. This ‘induced movement’ will be discussed later (pages 113–14). There is, however, plenty of evidence that this is not responsible for the auto-kinetic effect. The movements occur in directions unrelated to the drift of the eye’s particles (when these are made clearly visible, with oblique lighting) and in any case the particles are not generally visible.
A very different theory, held by some ophthalmologists, is that the eyes cannot maintain their fixation accurately on a spot of light viewed in darkness, and the drifting of the eyes causes the image of the spot of light to wander over the retina, so that the light seems to move. This theory was all but disproved in 1928 by Guilford and Dallenbach, who photographed the eyes while subjects observed a spot of light and reported movements they saw. When the reported movements of the spot were compared with the photographic records of the eye movements, no relationship was found between them. In addition, the eye movements under these conditions were extremely small, while the autokinetic movements can be huge. This experiment seems to have gone largely unnoticed.
Attempts to explain the wandering of the light in the dark generally suppose that something is actually moving—particles in the aqueous humour, the eyes, or some sort of reference frame in the brain. This last suggestion formed an important part of the Gestalt psychologists’ theory of perception, and they attached great weight to the wandering-light effect. Kurt Koffka, in his celebrated Principles of Gestalt psychology of 1935, says of it:
The autokinetic movements are the most impressive demonstration of the existence and functional effectiveness of the general spatial framework, but the operation of this framework pervades our whole experience.
Is the argument sound? I believe it contains an important fallacy. What is true for the observed world of objects, does not necessarily hold for errors of observation, or illusions. It is important to appreciate the difference. Any sense organ can give false information: pressure on the eye makes us see light in darkness; electrical stimulation of any sensory nerve endings will produce the experience normally given by that sense. (See page 97).
If movement is falsely signalled by some kind of neural disturbance, we should expect to experience movement though nothing is actually moving. This is familiar from man-made instruments: the speedometer of a car may become stuck at a reading of say 50 m per hour and will indicate this speed though the car is stationary, locked up in its garage. Similarly, pressure on the eyes makes us see light when there is no light.
It is true that all real movement of objects in the world is relative, and we can only speak of or measure movement of objects by reference to other objects. This, indeed, is the basis of Einstein’s theory of special relativity. This point was made two hundred years earlier, by Bishop George Berkeley, when he challenged a point in Newton’s Principia, of 1687:
If every place is relative, then every motion is relative … Motion cannot be understood without a determination of its direction which in its turn cannot be understood except in relation to our or some other body. Up, Down, Right, Left all directions and places are based on some relation and it is necessary to suppose another body distinct from the moving one … so that motion is relative in its nature.
Therefore, if we suppose that everything is annihilated except one globe, it would be impossible to imagine any movement of that globe.
It has been assumed by many writers on perception, that if nothing is moving—not the eyes, particles in the eyes, nor anything else—it would be impossible to experience even illusions of movement, for example of the spot of light in darkness. The wandering light has been taken to represent the same situation as Berkeley’s globe, when everything except it is annihilated; but it is actually very different.
The error lies in supposing that false estimates of movement—illusions of movement—require something moving relative to something else. But they can result simply from a disturbance or a loss of calibration of the measuring instrument, whether it be a speedometer or the eye. We may now seek the kind of disturbance or loss of calibration of the visual system responsible for the wandering light. To do this, we will produce large systematic illusory movements of the spot of light by deliberately upsetting the eye–head system.
If a person looks hard in any direction for several seconds, and then returns the eyes to their normal central position, while a small dim light is viewed in darkness, the light will usually be seen to speed in the direction in which the eyes were held. It sometimes moves in the opposite direction, but seldom in any other. The illusory movement may continue for several minutes, when the eye muscles are asymmetrically fatigued (Figure 6.4). Now fatigue of the eye muscles requires abnormal command signals to hold the eyes’ fixation on the stationary light; but these are the same as the signals which normally move the eyes, when they follow a moving object. We thus see movement when some of the muscles are fatigued, although neither the eyes nor the image on the retinas are moving. The usual wandering movements of the autokinetic effect seem to be due to command signals maintaining fixation in spite of slight spontaneous fluctuations in the efficiency of the muscles, which tend to make the eyes wander. It is not the eyes moving, but the correcting signals applied to prevent them moving which cause the light apparently to move in the dark.
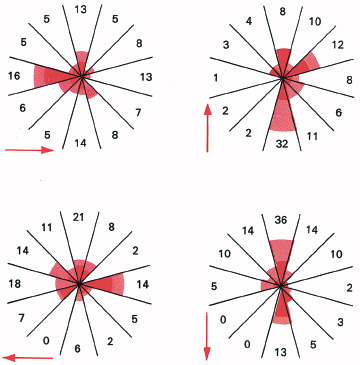
6.4 These ‘clock histograms’ show how a small dim light viewed in darkness appears to move after straining the eyes in one of four different directions for 30 seconds each time. The arrows show the direction of strain; the dark tinted areas show illusory movement during the next 30 seconds, while the light tinted areas show the following 30 seconds. The numbers give the duration in seconds of illusory movement in each direction during the first two minutes after strain.
We may now ask: if the correction signals move the spot of light in the dark, why don’t they cause instability of the whole visual world, in normal conditions? Why is the world generally stable? There is no certain answer to this question. It may be that in the presence of large fields of view, the aberrant signals are ignored, because the brain assumes that large objects are stable, unless there is clear evidence to the contrary. This is borne out by the effect of ‘induced movement’ (page 113). But first we should recognize that sometimes the world does swing round:
The case of the wandering world
The world swings around when we are very fatigued, or suffering the less pleasant effects of alcohol. This was described by the Irish wit Richard Brinsley Sheridan. After a party, friends led him to the front door of his house in Berkeley Square and left him. Looking back, they saw him still standing in the same position. ‘Why don’t you go in?’ they shouted. ‘I’m waiting until my door goes by again—then I’ll jump through!’ replied Sheridan.
Just how this ties up with the wandering light is not entirely clear. It may be that the eye movement command system is upset; or it may be that alcohol serves to reduce the significance of the external world, so that error signals normally disregarded are accepted. Just as we can become possessed by fantasies and irrational fears when tired or drunk, so might we become dominated by small errors in the nervous system, which are generally rejected as insignificant. If this is so, one might expect schizophrenics to suffer from instability of their visual world, but I know of no evidence for this.
The waterfall effect
We have found that the illusory movements of the spot of light viewed in darkness are due to small disturbances of the eye–head system. We might now expect to find illusions of movement due to disturbances of the image–retina system—and indeed we do. Various regions of the visual field may appear to move in different directions, and at different rates. The illusory effects are bizarre, and sometimes paradoxical, for it is possible to see movement without change of position.
After-effects from a turntable
A rotary after-effect is produced by looking steadily for say half a minute at the central pivot of a rotating record player. If the turntable is stopped suddenly it will seem to continue rotating—but in the opposite direction.
Rotating a spiral (Figure 6.5) produces illusory expansion or contraction. This transfers to another object, such as someone’s face. As different regions are affected differently (and can be rotations, and changes of size) the effect must be retinal—in the image–retina system.

6.5 When the spiral is slowly rotated it is seen to shrink, or expand, depending on the direction of rotation. When stopped it seems to do the opposite of the inducing stimulus. This illusory shrinking or expanding is paradoxical, as the spiral does not appear to change in size.
This best known image–retina disturbance was described by Aristotle. Just after watching a waterfall, he saw the bank of the river apparently moving. So it is known as the waterfall effect though it is produced by any continuous motion, including expansion or contraction. The after-effect is always in the opposite direction to the ‘inducing stimulus.
This can also be shown by following a moving belt of stripes with the eyes, returning the eyes rapidly to the beginning (with the lights switched off), following the movement again, and repeating this several times (Figure 6.6). In this way movement in one direction is experienced using the eye–head, but not the image–retina system. When the belt is stopped after prolonged viewing with the following eye movements, there is no noticeable after-effect. So we may attribute the waterfall effect, at least almost entirely, to adaptation of the image–retina system. Only this could give illusory expansion or contraction.
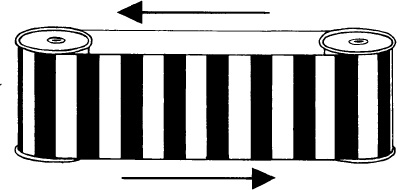
6.6 The waterfall effect. The stripes can be seen moving either by holding the eyes still (with a stationary fixation point) or by following the movement with the eyes. When the eyes are stationary, the image–retina system is stimulated. When they are following, the eye–head system signals movement—but not from the retina. Is there an after-effect from the eye–head system? It is certainly much less—probably absent.
Just why adaptation to retinal motion occurs is an interesting question. We have seen from the work of Hubel and Wiesel that movement is represented in separate neural channels, and that different channels indicate different directions of movement (Figure 4.8). It is reasonable to assume that some of these channels become ‘fatigued’ with prolonged stimulation, and this unbalances the system, giving illusory movement in the opposite direction. But it may have a use—for re-calibrating the system.
It remains a problem as to whether the adaptation takes place in the retina or in the brain. The retina may seem too simple for such a complex effect, but it is very difficult to rule out retinal adaptation as at least a part-cause. One might think (and several experimentalists who ought to have known better have thought) that the issue could be decided by looking at the moving stimulus with one eye, while holding the other closed, then noting whether the after-effect occurs when viewing a stationary object with the previously unstimulated eye. The answer is that it does occur, at about half strength. But this does not show conclusively that the adaptation took place in the brain—for it is possible that the previously stimulated eye goes on sending up movement signals after it is shut, and that these are ‘projected’ into the field of the unstimulated eye. This is perfectly possible, as one does not know which eye is active, although one tends to think, often wrongly, that whichever eye is open is doing the seeing. (To avoid this problem ‘pressure blindness’ has to be employed to block signals from the stimulated eye—but pressing the eye to impede blood flow to the retina is extremely dangerous; definitively not to be tried).
As we have said, if the after-effect from the rotating spiral is examined carefully, it is seen that the illusory movement is paradoxical—for example, the spiral expands without getting larger (Figure 6.5). So it is changing yet not changing. This is impossible for real objects, but we must remember that what is true for real objects may not hold for perception. We can even experience things that are logically impossible. In this case of the after-effect of movement, of expansion with no change in size, we may suppose that this comes about because velocity and positions are signalled by separate neural channels. When one of these is upset, it will disagree with signals from the other channel. So the brain is in the position of a trial judge getting conflicting evidence from witnesses. When incompatible stories are accepted, the judge is stuck with a paradox. As the perceptual system has many channels and many sources of information, the brain must serve as judge—sometimes rejecting channels, or sources of information, sometimes accepting incompatible signals, or conflicting information. Then we experience a paradox. We should not be too surprised that illusions and hallucinations can be impossible to describe in terms of normal experience.
Relativity of movement
So far, we have considered the basic mechanisms for perceiving movement, either by stimulation of the retina by moving images or by the eye following moving objects. There is, however, far more to the perception of movement. Whenever there is motion the brain has to decide what is moving and what is stationary. Although, as we have seen, it is fallacious to suppose that illusory movement necessarily involves any actual movement, it remains true that all real movement is relative, and so a decision is always required about what is moving. An obvious example occurs whenever we change our position by walking, driving, or flying. We generally know whether the movement is due to our own motion among surrounding objects, rather than due to their movement; but this involves a decision. Sometimes the decision is made wrongly, giving errors which can be particularly dangerous, because perception of movement is of such prime importance for survival. This is as true in the case of humans living in an advanced civilization, as ever it was in the primitive state in which eye and brain evolved.
Most perceptual research has been undertaken with the observer stationary, often looking into apparatus showing flashing lights or pictures. But real-life perception occurs during free movement of the observer, in a world where some of the surrounding objects are also in motion, such as when driving a car in traffic. There are severe technical problems for investigating such real-life situations but the attempt is worth while, as the results are important for driving, flying, and also space flight. Astronauts are trained on highly realistic simulators to make judgements of speed, and size, and distance. The same is true for pilots. Visual simulators are important for flying and space training, and the simpler aspects of driving; but simulators never copy reality quite perfectly, so skills learned from them are seldom quite appropriate for the real thing. Discrepancies can show up at awkward moments as bad habits—‘negative transfer’, from the simulator.
As we have seen, there is always a decision involved as to what is moving. If the observer is walking, or running, there is seldom a problem; for he or she has a lot of information from the limbs, signalling movement in relation to the ground. But when carried along in a car, or an aircraft, the situation is very different—the feet are off the ground, and the only source of information is the eyes. This is so except during acceleration (or deceleration, or moving in a curved path) when the balance organs of the middle ear give some, though often misleading information.
Induced movement
The phenomenon known as induced movement was investigated by the Gestalt psychologist Karl Duncker. He devised demonstrations to show that, when there is relative motion, we generally see the largest object as stationary, the smaller as moving. Thus we tend to see the small moon as rushing across the sky, though it is the large clouds that are moving past. This is not easy to demonstrate in a room, but is just possible by projecting a small patch of light on a large movable screen (Figure 6.7). When the screen is moving, what is seen as moving is the small stationary patch of light.
It may be noted that actually there is some information available on what is moving—whether the eyes rotate. But the eye–head signal is not always sufficient to decide the issue. This is relevant to the curious question of why the visual world does not swing round with eye movements, though it does when a cine camera is panned around a scene (page 101).
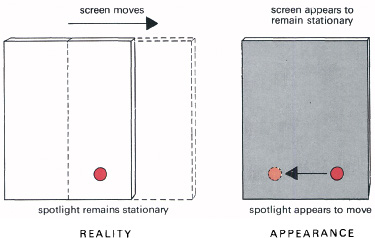
6.7 Induced movement. A small patch of light is projected on a large screen, which is moved. This makes the small stationary light appear to move in the opposite direction.
It seems clear that the brain bets on small objects moving, as generally large things (trees and houses) are stationary. When the bet is wrong, it can be disturbing and dangerous, as when driving a car: ‘Is it my hand-brake that is off—or is that idiot over there running backwards?’ The answer is important!
Motion and distance
When we move, say to the right, near objects move against our motion—to the left—and further objects seem to move with us, to the right. This is motion parallax, and it has a simple explanation. It is the same effect as the slightly different views of each eye giving stereoscopic vision, and indeed (as Brian Rogers at Oxford University has pointed out) stereo depth perception may have evolved from motion parallax—which itself is a valuable cue to depth. Although the geometry of dynamic parallax is simple, there are surprisingly subtle visual effects.
When we observe the moon while travelling in a car at night, we see it apparently moving along with us—rather slowly. At 50 km per hour, the moon may seem as moving at say 10 km per hour. We see it as moving more slowly than us yet keeping up—again a paradox.
The moon is so distant, we can regard it as at infinity. As the car moves along, the angle to the moon remains unchanged. So unlike near objects it does not change its position, though we are moving along under it. But perceptually it lies at a distance of only a few hundred metres. We know this from its apparent size. It subtends an angle of 0.5°, but looks the size of say an orange a few hundred metres away. The moon does not get left behind, because it is so distant that its angular position does not change—yet it appears to be not so far away. The only way the perceptual system can reconcile these, is to ‘assume’ that the moon is moving with the car. The apparent velocity of the moon is set by its apparent distance. If we change the apparent distance (by viewing it through converging prisms to change convergence of the eyes) then it seems to move at a different speed.
A particularly interesting, closely related effect, can be seen by projecting stereo pictures on a screen (with crossed polaroids, or red–green glasses). When the observer moves his or her head from side to side, the stereo scene seems to rotate, to follow the observer’s every movement although it is fixed on the screen. Thus, a stereo projection of a corridor swings round, so the apparent front moves with the observer. This is opposite to the motion of a true scene—such as the real corridor. There is more: if the stereo pictures are set to coincide far away, at the end of the corridor, it will not rotate, but will shift equally along its entire length. If now, the stereo pictures are made to coincide near the middle of the corridor (so there are opposite disparities at its far and near ends), the corridor appears to rotate around the point of zero disparity, still with the observer’s movements.
Stereo projection is revealing, because the scene lies flat on the screen, so there is no physical motion parallax even though it is seen in depth. Normally the world rotates around the point of fixation of the eyes, in the opposite direction to the observer’s motion. (This is clearly seen while travelling in a train). But observing flat pictures in stereo depth, the opposite happens; they appear to rotate with the observer’s movements, around the point of convergence of the eyes where disparity is zero—set by the separation of the stereo-pair on the screen. These effects are well worth trying out. As these illusory movements are in the opposite directions to normal motion parallax, evidently what we are seeing are compensations to normal motion parallax. It also works for ordinary pictures seen realistically in depth.
Movements of life
The Swedish psychologist Gunner Johansson has produced extremely striking demonstrations of how little information is needed for seeing moving humans, and animals. If small lights are placed at the joints of someone’s arms and legs, he or she is effectively invisible in a dark room—until he or she moves (Figure 6.8). Immediately the lights are seen as a human figure, and can be identified as male or female, from the slight differences of movements. This does not work well for unfamiliar objects, and works best with living beings, especially humans.
The ability to recognize friends and foes has always been very important for survival, especially perhaps at night, and where the person or animal is partly hidden. So very little information is needed, when there are movement signals. The more probable the object, the less information is needed. This is a wonderfully powerful demonstration of the important perceptual principle that we can see more than meets the eye.
Movement in cinema and television
All the sensory systems can be fooled. Perhaps the most persistent fooling is by cinema and television. They present a series of still pictures, but what we see is continuous action. This relies on two distinct (though often confused) visual principles: persistence of vision and the phi phenomenon.
Persistence of vision
This is simply the inability of the retina to signal rapidly changing intensities. A light flashing at a rate greater than about 50 flashes per second appears steady; for bright lights this critical fusion frequency (CFF) may reach 100 or more flashes per second. Cinema presents 24 frames per second; but a three-bladed shutter is used to raise the flicker rate to 72 flashes per second—three for each picture. Television (British standard) presents 25 fresh pictures per second, each given twice, to raise the flicker rate to 50 per second. Television flicker is reduced by ‘interlaced scanning’, in which horizontal sections of the picture are built up by scanning in bands of lines, rather than continuously down the screen. Nevertheless, television flicker can be annoying, and is potentially dangerous to sufferers from epilepsy. Indeed, flicker is used to evoke symptoms for diagnostic purposes. Flicker can be a hazard when driving along a row of trees with a low sun. When landing a helicopter in tropical conditions, the rotor blades produce a violently flickering light which can be dangerous. The ‘stroboscopes’ used in discotheques can be similarly disturbing, and should probably be avoided; as also, of course, should the high levels of sound, capable of producing permanent hearing loss. The senses are too precious to be abused and damaged unnecessarily.

6.8 When lights attached to the joints of arms and legs move, two people are immediately seen. (After Johansson 1975, © 1975 Scientific American, Inc., all rights reserved.)
Low-frequency flicker produces very odd effects on normal observers, as well as on those with a tendency to epilepsy. At flash rates of five to ten per second, brilliant colours and moving and stationary shapes may be seen and can be extremely vivid. Their origin is obscure, but they probably arise from direct disturbance of the visual system of the brain, the massive repeated bursts of retinal activity overloading the system. The patterns are so varied it is difficult to deduce anything about the brain systems which have been disturbed. This can be an unpleasant experience, leading to headache and nausea.
Phi movement
The other basic visual principle upon which cinema depends is the apparent movement known as the phi phenomenon. It is usually studied in the laboratory with a very simple display, just two lights, switched so that just after one light has gone off the other comes on, in sequence. What is seen—when the distance between the lights and the time intervals between their flashes is about right—is a single light, apparently moving between the flashing lights. It was argued by the Gestalt psychologists that this apparent movement across the gap between the lights is due to an electrical charge, sweeping across the brain, to give sensation of motion and fill the gap. As the Gestalt psychologists, early this century, thought that the phi phenomenon demonstrated such a basic process, it was studied intensively. Probably all authorities would now, however, consider the Gestalt view of the matter fundamentally mistaken.
The simple notion that the image–retina system is tolerant of gaps explains phi movement. For vision needs tolerance, to cope with all manner of inadequacies. (The situation is like a key and a lock: the key does not have to be exactly a certain shape; some tolerance is essential, or any slight change in the lock or the key would stop it working.) This use of tolerance is a basic engineering principle.
Moving objects can momentarily disappear, as when a running animal for a moment is hidden behind a nearby tree; but it is useful for observers to see this as a continuous movement of the same object. The image–retina system tolerates gaps, provided the jumps in space and time are not too large. As a fortunate pay-off, this tolerance in space and time allows cinema and television to be economically possible.
Seeing causes
The importance of regular lawful relations for perception was studied experimentally by Albert Michotte, at Louvain in Belgium. For many years he investigated visual perception of causality, moving patches of colour with various velocities and time delays, using the apparatus shown in Figure 6.9. He arranged for one coloured patch to move towards, and sometimes to touch another which then moves off, generally after a small delay. With some combinations of velocity and delay, there was an irresistible impression that the first patch had struck the second—pushing it causally, as objects such as billiard balls interact in the world of physics, though these were but coloured patches. Michotte thought that the seeing of causes is an innate response, but this is hard to prove, as it might be learned from years of seeing how objects interact.
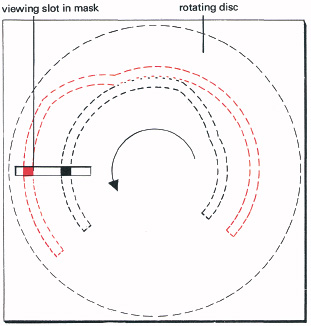
6.9 Michotte’s apparatus for finding rules of perceived causality. A coloured patch moves towards another, which after a controlled delay moves away. This appears causal when delays and velocities approximate interactions of typical objects. (This can now be done far more simply with computer graphics.)
Similarly, one sees causes in a cartoon film, even though the objects are abstract. This gets interesting when causal interactions are inappropriate to knowledge of the objects. Indeed, much of the humour of cartoons lies in disagreements between causal rules and object meanings. Here, again, we find a rich mixture of rules (syntax) and meaning (semantics) in perception. Cartoons are an ideal medium for playing upon and investigating primitives and sophistications of seeing.
Although Michotte was inclined to think that seeing causes is given innately because his observers had general agreement (though there were difficulties with differences in their verbal descriptions), we all encounter much the same kinds of objects, so shared experience might set up similar perceptions of cause. Observers’ agreement over common situations does not distinguish innateness from learning from experience; but this is always a very difficult issue to resolve. Experiments with unnatural objects in computer games and virtual reality might give new evidence for settling this old controversy.
However this may be, causal relations in science are often very different from appearances—as ‘arrows’ of causal direction must be established by conceptual models. Here is an important difference between perceptual and conceptual hypotheses. Night follows day, but neither causes the other. We need a mental model of the Earth spinning round to ‘see’ the cause conceptually.