The Organization of the Living
IN LIFE EVERY BEGINNING IS UNIQUE, but none is isolated and self-contained. We start out as single cells, formed from the union of two parent cells, an egg and a sperm. Our parent cells, too, are offspring, produced in the bodies of our mothers and fathers, who owe their origins to still earlier couplings of egg and sperm. Every beginning has a beginning before it and another one before that, leading back through the receding biological past to its time and place of origin, the beginning of life on Earth.
Around four billion years stand between our time and that distant aeon of life’s emergence. A number of that magnitude is hard for the human mind to comprehend. Its vastness seems to diminish the force of pointing to our common ancestry with all the living things on Earth. Closer to home are the one hundred billion nerve cells or neurons that make up the human brain. All are the progeny of a small fold of cells that emerged when we were embryos of about four weeks. Inside each one of them, in its protein and DNA, we can find a family resemblance to the genes and enzymes of all the other living cells on Earth.
We harbor the past everywhere within our bodies. To the cells inside us the chemical composition of the somatic environment plays a role reminiscent of the ocean environment where the earliest cells resided. Three billion years ago bacteria swam in the warm shallows of the Earth’s primeval seas. Among their descendants today are the bacteria dwelling within our bodies, without which we could not live, while other remnants of their progeny, such as mitochondria and mobile cilia, exist inside our modern cells (Margulis 1984, 1993).
The idea that all life evolved from a common ancestor and hence that there is an underlying unity to the widespread diversity of life is central to modern biology. We are taught in school that the unity of life is based on three things: (1) all living things are made of cells; (2) the life cycles of all cells—their formation, growth, development, reproduction, and so on—are based on chemical reactions among similar sorts of molecules; and (3) the way that amino acids are put together to form proteins is specified by DNA and RNA according to a precise and nearly universal scheme.
To these points we can add a fourth, following theorists who address the question “what is life?” by searching for principles of biological organization. There is a basic formal organization of life, and its paradigm and minimal case is to be found in the single cell. A single-cell organism is a self-making or self-producing being. Self-production is different from reproduction: In reproduction, a cell divides in two; in self-production, a cell continuously produces itself as a spatially bounded system, distinct from its medium or milieu. What is remarkable about self-production is that every molecular reaction in the system is generated by the very same system that those molecular reactions produce. Some years ago the neurobiologists Humberto Maturana and Francisco Varela drew attention to this circular, self-producing organization and called it autopoiesis (Maturana and Varela 1973, 1980; Varela, Maturana, and Uribe 1974). My purpose in this chapter is to present some key ideas of the autopoietic approach to life.
In 1858, one year before the publication of Charles Darwin’s Origin of Species, the German biologist Rudolf Virchow propounded what is now known as the cell theory: the basic unit of all life is the cell and all cells arise from preexisting cells. In his book Cell Pathology Virchow wrote:
Each day brings forth fresh discoveries but it also opens up fresh matters for discovery. Is anything positive in histology, we have to ask ourselves. What are the parts of the body whence commence the vital actions? Which are the active elements, and which the passive? These are queries which have given rise to great difficulties, dominating the field of physiology and pathology, and which I have solved by showing that “the cell constitutes the true organic unit,” that it is the ultimate irreducible form of every living element, and that from it emanate all the activities of life both in health and in sickness. (Virchow 1967, p. 23)
Virchow was not the first to observe the living cell. The earliest observations of cells date back to 1674 when Anton van Leeuwenhoek, the Dutch naturalist and lens-grinder, looked at pond water through a microscope and saw tiny creatures swimming there. In Virchow’s own century many naturalists observed that the tissues of living organisms are composed of cells. But many of them also believed in an idea known as “the free formation of cells”—that cells develop out of a previous noncellular material. Virchow rejected this idea about how cells come into being. He summed up his rival view in a famous Latin aphorism: “The existence of a cell presupposes the prior existence of some other cell—omnis cellula e cellula (every cell must come from some other cell)—just the same as a plant cannot occur except it be derived from some other plant, or an animal from some other animal” (Virchow 1967, p. 25).
It might seem that biology has moved beyond Virchow’s view of the cell as “the ultimate irreducible form of every living element.” Molecular biology has taken us deep inside the cell, into the microscopic universe of DNA, RNA, and amino acids. There can be no doubt that irreplaceable insight and discovery have been gained from the molecular perspective. Yet as a number of biologists have discussed in recent years, molecular biology can lose sight of the organism as a whole (Goodwin 1994; Rose 1997). Here, too, Virchow’s words are worth recalling: “There is no doubt that the molecular changes occurring inside the cells are referred to some part or other composing them; the final result is, however, due to the cell from which the vital action started and the living element is not active except in so far as it presents us with a complete whole enjoying its own separate existence” (Virchow 1967, p. 23).
The idea that the cell is a complete whole and that all cells arise from preexisting cells would seem to pose a dilemma: Either life has always existed or there must have been a first cell. But if all cells arise from preexisting cells, how could life ever come into being or get started in the first place? “There is only one way to form cells,” Virchow wrote, “that is by fissiparity; one element is divided after the other. Each new generation proceeds from some preceding generation” (Virchow 1967, p. 25). If life had an origin, a historical beginning, then there must have been a first generation of living cells. But how could a first generation come into being without some previous generation to rely on? How could life arise from nonlife?
According to present scientific understanding, our solar system and the Earth took form about five billion years ago, followed some billion years later by the origin of life during the Archean Aeon, with the subsequent expansion of life into the five kingdoms of living beings known today. Monera is the oldest kingdom; it comprises all the prokaryotes or cells having no nuclei (bacteria). The members belonging to the other four kingdoms are all eukaryotes or organisms made up of nucleated cells—Protoctists (e.g., amoebae and green algae), Fungi, Plants, and Animals (Margulis and Schwartz 1988).
The origin, expansion, and history of life are usually pictured as a tree with many branches, an image that provides a way of visualizing the unity of life due to evolution from a common ancestor. Living things arise from earlier living things, and thus there is an underlying continuity from the most ancient to the most modern forms of life. With the origin of life over 3.5 billion years ago, however, something novel happened, a new form or order of things emerged. Although life is composed of physical and chemical elements, living organisms are different from other sorts of physicochemical things, and the surface of a planet populated by living beings is strikingly different from a planet that has no life.
Given this naturalistic and evolutionary perspective, the only way out of the dilemma posed here is to suppose that there must have been simpler, precursor chemical systems, protocells or “Ur-cells,” that led eventually to the “ancestor cells” of the oldest bacterial organisms (Morowitz 1992; see also Margulis 1984). We know that all contemporary life must derive from a universal ancestor, which might have been a single cell or population of cells, but we do not know how many independent origins of protocells there were leading up to this ancestral organism. As Harold Morowitz observes in his book Beginnings of Cellular Life: “The problem is not simply the origin of life, it is the physical chemical formation of the Ur-organism and a subsequent evolutionary epoch giving rise to the universal ancestor” (Morowitz 1992, p. 88).
A number of different, but interrelated, questions arise from this perspective: What were the environmental conditions of the early Earth when life arose? When exactly did life arise? Did life arise more than once, at different times and places? How did complex bacterial cells arise from simpler protocellular systems? How did protocellular life arise from nonlife? What exactly do we mean by “life” in this minimal sense?
Many scientists prefer to avoid the last question altogether because they see it as philosophical rather than empirical. But the question will not go away that easily. The question of how and when life originated is inseparable from the question of what a living system is. If the aim is to determine how and when life arose, then one needs a clear way of characterizing what distinguishes living systems from nonliving ones. Such a characterization could also serve as a standard or criterion for recognizing life elsewhere on other planets, or for determining whether anything we might someday synthesize artificially would qualify as living.
In contemporary biology, there are three main approaches to characterizing life. One approach is to characterize life on the basis of genetics and reproductive populations. One generation of bacteria makes the next generation of bacteria; one generation of fruitflies produces the next generation; and so on, for all plants and animals. From this perspective, life depends on historical continuity and evolution, on the genetically based linkage of generations and the arising of novel variants within a population as a result of various evolutionary factors.
A second way to characterize life is more ecological. From this perspective, individual organisms are seen not only as members of reproductively linked populations, but also as beings that interact constructively with their environments, and so change the world in which they and their descendants live. Organisms are “niche-constructing” beings (Odling-Smee 1988). A particularly large-scale example of this way of thinking can be found in the Gaia theory put forward by atmospheric chemist James Lovelock and microbiologist Lynn Margulis (Lovelock 1979, 1988; Lovelock and Margulis 1974; Margulis and Lovelock 1974). In their view, life is a phenomenon that can occur only on a planetary scale. Life’s persistence depends on integrated processes involving the biota (the sum total of living things) and the Earth’s material environment (atmosphere, rocks, and oceans). Lovelock in particular has argued that were it not for the global presence of living organisms, the Earth would be rendered uninhabitable for life.
Finally, there is a third way to characterize life, one in which the focus is on the single individual entity or organism, here and now (Luisi, Lazcano, and Varela 1996). Suppose you are looking at a single specimen. It could be a strain of bacteria, a newly discovered jellyfish, a synthetic supramolecular complex, a specimen of presumed life on a distant planet, or a new insect-like robot. You then wonder: Is this a living thing or not? This question targets an individual entity, and the evolutionary and ecological backgrounds are of less immediate importance (they could be unknown or difficult to establish). In this sort of local, here-and-now situation, the demand for a clear criterion to distinguish life from nonlife seems especially pressing.
These three ways of characterizing life are complementary, not mutually exclusive. In the case of life on our own planet—the only life we know at present—“life” in a full sense means reproductively linked populations of ecologically embedded and active individual organisms. Modern molecular biology, however, has neglected the individual characterization of life and adopted almost exclusively the genetic-population characterization. Yet the individual characterization has a certain logical priority over the reproductive one. The reason is that reproduction presupposes an individual and some process whereby that individual reproduces. Thus the individual logically and empirically precedes reproduction and the evolutionary process of selection (Fleischaker 1988; Maturana and Varela 1980, pp. 96–111; 1987, pp. 55–74). In other words, any given population or evolutionary series is secondary to the individuation of the members of that population or series. Hence a characterization of life that accounts for individuality has logical priority over one that does not.
In the history of biology, a variety of criteria for distinguishing between living and nonliving systems have been proposed—carbon-based chemical composition, nucleic acids, the capacity to move, the ability to reproduce, and so forth. A standard procedure in biology textbooks today is to list such characteristics in order to delineate the class of living things. Such lists usually include metabolism and self-maintenance, the genetic material of DNA and RNA, and evolution by natural selection. The problem with this approach is that it is descriptive, not explanatory. It takes for granted the distinction between living and nonliving, and then it lists some common characteristics of systems accepted as living. But how do we know which characteristics should be included on the list or when the list is complete? Lists describe things but do not explain them. To explain we need a theory.
According to the cell theory, all living things are made up of cells. Although this statement does provide a scientific criterion of life, it has shortcomings. It is based on the observation that no life on Earth has ever been found without cells. But in the absence of a clear characterization of what a living system is, we cannot simply assume either that noncellular life is impossible or that no such life has ever existed. Moreover, to define life in terms of cells is basically tautological: life is cellular because there is no life without the cell. To get out of the tautology we need to specify the basic properties of a cell without invoking the notion of life. In other words, we have to specify what it is about a cell that makes it living.
One strategy for meeting this demand would be to characterize a living system, such as a cell, in terms of its relational form or organization (Maturana and Varela 1980, 1987). A system’s organization consists of the relations that define the system as being a member of a specific class. For something to qualify as an automobile, for instance, its parts have to be arranged or related to one another in a certain way. In specifying a system’s organization, one abstracts a pattern or set of relations that defines what kind of system the system is. A system’s organization is thus not equivalent to its actual structural relations and components because the same organization can be structurally realized in different ways, and a system can undergo structural change without necessarily changing its organization. Thus the organization of an automobile can be realized in different physical materials and mechanisms, and these can change during the automobile’s lifetime. Similarly, a single cell, such as a bacterium, undergoes many structural changes during its life cycle without changing its organization as a unicellular organism. Is it possible to specify a definitive organization in the case of living systems? With regard to the cell theory, can one specify the organization of a cell independent of its structure?
The theory of autopoiesis, formulated by Maturana and Varela (1973, 1980, 1987), addresses precisely these questions about the organization of the living.1 Maturana and Varela focus on the single, biological individual, the living cell. A cell is a thermodynamically open system, continually exchanging matter and energy with its environment. Some molecules are imported through the membrane and participate in processes inside the cell, whereas other molecules are excreted as waste. Throughout this exchange, the cell produces a host of substances that both remain within the cell (thanks to its membrane) and participate in those very same production processes. In other words, a cell produces its own components, which in turn produce it, in an ongoing circular process. The word “autopoiesis” was coined to name this kind of continual self-production. A cell is a self-producing or autopoietic unity. Systems that do not produce themselves, but whose product is different from themselves, are said to be allopoietic. For example, a ribosome (a small spherical body within a living cell composed of RNA and protein, and the site of protein synthesis) is a crucial participant in the autopoiesis of a cell, but is produced by processes other than those that constitute its own operation (Varela, Maturana, and Uribe 1974, pp. 188–189). Maturana and Varela also distinguish autopoietic systems from heteropoietic ones, which are allopoietic systems that arise in the realm of human design, such as cars and digital computers. Maturana and Varela’s fundamental proposition is that living systems are autopoietic or have an autopoietic organization. They are organized in such a way that their constituent processes produce the components necessary for the continuance of those same processes.
The concept of the autopoietic organization arose from an attempt to abstract from the molecular processes of the cell the basic form or pattern that remains invariant through any kind of structural change, as long as the cell holds together as a distinct entity. To understand the autopoietic organization we therefore need to view it at a cellular level.
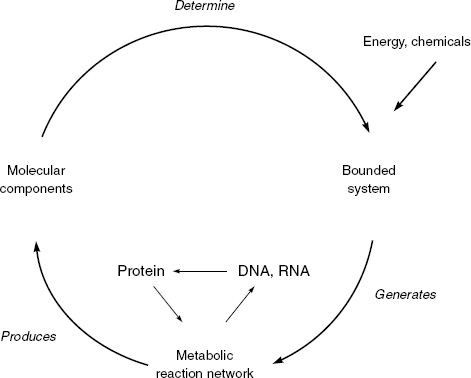
Figure 3.1, as we saw earlier, gives a schematic illustration of the basic pattern or organization of a minimal cell. By “minimal cell” I mean a cell with a minimal organization sufficient for it to be a distinct, independent entity. A cell is spatially formed by a semipermeable membrane, which establishes a boundary between the inside of the cell and the outside environment. The membrane serves as a barrier to free diffusion between the cell and the environment, but also permits the exchange of matter and energy across the boundary. Within this boundary, the cell comprises a metabolic network. Based in part on nutrients entering from outside, the cell sustains itself by a network of chemical transformations. But—and this is the first key point—the metabolic network is able to regenerate its own components, including the components that make up the membrane boundary. Furthermore—and this is the second key point—without the boundary containment provided by the membrane, the chemical network would be dispersed and drowned in the surrounding medium. Thus the cell embodies a circular process of self-generation: thanks to its metabolic network, it continually replaces the components that are being destroyed, including the membrane, and thus continually re-creates the difference between itself and everything else.
We can approach this idea in a slightly different way: a cell stands out of a molecular soup by creating the boundaries that set it apart from what it is not. Metabolic processes within the cell determine these boundaries, but the metabolic processes themselves are made possible by those very boundaries. In this way the cell emerges as a figure out of a chemical background. Should this process of self-production be interrupted, the cellular components no longer form a spatially individuated whole and they gradually diffuse back into a molecular soup.
The pattern presented in Figure 3.1 is what Maturana and Varela mean by the autopoietic organization. Anything that realizes this organization is an autopoietic system. In Maturana and Varela’s view, “the notion of autopoiesis is necessary and sufficient to characterize the organization of living systems” (1980, p. 82; emphasis omitted). They also state: “Autopoiesis in the physical space is necessary and sufficient to characterize a system as a living system” (p. 112). Some scientists have recently argued, however, that although autopoiesis is a necessary condition for life, it is not sufficient (Bitbol and Luisi 2005; Bourgine and Stewart 2004; Luisi 2003). I will take up this issue at the end of this chapter.
Maturana and Varela give several precise definitions of autopoiesis in their writings. Their original and canonical definition is of an “autopoietic machine:”
An autopoietic machine is a machine organized (defined as a unity) as a network of processes of production (transformation and destruction) of components that produces the components which:
(i) through their interactions and transformations continuously regenerate and realize the network of processes (relations) that produced them; and
(ii) constitute it (the machine) as a concrete unity in the space in which they (the components) exist by specifying the topological domain of its realization as a network. (Maturana and Varela 1980, pp. 78–79)
By “machine” Maturana and Varela mean a system that is defined by its organization and that hence can be explained in terms of the relations constituting that organization, rather than in terms of the structural components realizing that organization in a particular concrete system. “Components” and “processes of production” are abstract ways of characterizing what in the molecular domain are molecules and chemical reactions. Thus, roughly speaking, a molecular autopoietic system is one in which chemical reactions produce molecules that (i) both participate in and catalyze those reactions and (ii) spatially individuate the system by producing a membrane that houses those reactions.
A slightly different definition of the autopoietic organization is given by Varela, Maturana, and Uribe (1974) and then illustrated with reference to a cell:
The autopoietic organization is defined as a unity by a network of productions of components which
(i) participate recursively in the same network of productions of components which produced these components, and
(ii) realize the network of productions as a unity in the space in which the components exist.
Consider for example the case of a cell: it is a network of chemical reactions which produce molecules such that
(i) through their interactions generate and participate recursively in the same network of reactions which produced them, and
(ii) realize the cell as a material unity.
Thus the cell as a physical unity, topographically and operationally separable from the background, remains as such only insofar as this organization is continuously realized under permanent turnover of matter, regardless of its changes in form and specificity of its constitutive chemical reactions. (Maturana, Varela, and Uribe 1974, p. 188)
One might wonder what the phrase “generate and participate recursively” means in the cellular illustration of the first condition. It seems plausible to interpret “participating” in a reaction as meaning to be a reactant, and “generating” a reaction as meaning to catalyze that reaction (McMullin 1999, p. 2). Thus the definition’s first condition “is that the reaction network which characterizes the organization of the system must produce all the species of molecular component which are considered to materially constitute the system, and these components must themselves generate the reaction network, in the sense of catalysing some (or all?) of the reactions (which would otherwise occur at negligible rate)” (McMullin 1999, p. 3). The second condition is that the reaction network must also establish the system as a “unity in space,” that is, demaracte the system by establishing a boundary between it and the external environment. This condition is met in a cell by the production of a semipermeable membrane.2
In later writings, Varela (2000a) proposed the following simplified definition of autopoiesis. For a system to be autopoietic, (i) the system must have a semipermeable boundary; (ii) the boundary must be produced by a network of reactions that takes place within the boundary; and (iii) the network of reactions must include reactions that regenerate the components of the system.
In summary, the form or pattern of the autopoietic organization is that of a peculiar circular interdependency between an interconnected web of self-regenerating processes and the self-production of a boundary, such that the whole system persists in continuous self-production as a spatially distinct individual.
The autopoietic organization captures the minimal organization of a cell without invoking the notion of life or defining life in terms of the cell. Indeed, the autopoietic organization could be realized in many different kinds of molecular system, not just in biological cells as we know them. For example, chemical reactions in living cells involve nucleic acids (RNA and DNA) and the corresponding enzymes. The autopoietic characterization of minimal life, however, is not dependent on this particular structural arrangement of nucleic acids and proteins. The autopoietic characterization is more general and so could hold for an “uncoded life”—for instance, an early terrestrial protocell (Fleischaker 1990a) or a synthesized molecular system lacking nucleic acids (Bachman, Luisi, and Lang 1992). In other words, the autopoietic characterization of minimal life, though based on the living cell, sets out the general pattern of which DNA-based life is one possible expression.
Figure 5.1 shows the autopoietic organization in relation to the DNA-based version of life found in the structure of living cells on Earth. As we have seen, the autopoietic organization is a circular one: molecular components produced by an internal reaction network assemble into a semipermeable boundary, which contains the internal microenvironment and reaction network, which again produces the molecular components, and so on continually as long as the cell holds together. Whereas Figure 3.1 depicts the autopoietic organization without specifying the type of reactions and the chemical nature of the components, Figure 5.1 explicitly shows the role played by DNA and RNA in living cells.
One advantage of characterizing life in terms of the minimal organization or pattern of autopoiesis is that we can list clear criteria for determining whether anything satisfies that organization:
1. Semipermeable Boundary: Check whether the system is defined by a semipermeable boundary made up of molecular components. Does the boundary enable you to discriminate between the inside and outside of the system in relation to its relevant components? If yes, proceed to 2.
2. Reaction Network: Check whether the components are being produced by a network of reactions that take place within the boundary. If yes, proceed to 3.
3. Interdependency: Check whether 1 and 2 are interdependent: are the boundary components being produced by the internal network of reactions, and is that network regenerated by conditions due to the boundary itself? If yes, the system is autopoietic.
By applying these criteria, one can determine whether any given system is autopoietic, even at a level where commonsense conceptions of life are of little help. Consider the items in Table 5.1, each of which is analyzed according to the three criteria above.
A bacterium (a prokaryote) and an amoeba (a eukaryote) are autopoietic because they satisfy all three criteria. A crystal, however, is not autopoietic because its components are not generated from within itself. Replicative molecules, such as DNA and RNA, are also not autopoietic because they have neither a semipermeable membrane nor an internal reaction network, and thus they do not instantiate a circular, self-producing organization. Viruses are often described as living, but they do not satisfy the autopoietic criteria. A virus is a bounded structure with a protein coat, thereby satisfying the first criterion. But the second (and hence the third) is not satisfied because the molecular components of a virus (nucleic acids) are not generated inside the virus, but outside in the host cell. A virus has no metabolism of its own and thus is not self-maintaining in the autopoietic sense. Outside of a host cell, in the environment, a virus can persist, but it does not exchange matter with its environment in a continual self-producing way.
Table 5.1 Determining whether an entity is autopoietic

Another interesting case is autocatalytic sets of molecules. A catalyst is a molecular substance whose presence is needed for a particular chemical reaction to take place, or to speed up a reaction, but the catalyst is not changed by the reaction. Autocatalytic reactions are those in which the product of the reaction is a catalyst for that very reaction. An autocatalytic set or a collectively autocatalytic system is a chemical network made up of many self-perpetuating loops of molecular reactions. The system is composed of molecules that act as catalysts for the very reactions that produce those molecules. Such a network will be able constantly to re-create itself as long as there is a sufficient supply of food molecules. The key property of this sort of chemical network is catalytic closure. Except for the food molecules supplied from outside, every molecule in the system is produced by the autocatalytic reactions that make up the network.
Stuart Kauffman has suggested that catalytic closure is the basis of life: “Life, at its root, lies in the property of catalytic closure among a collection of molecular species. Alone, each molecule is dead. Jointly, once catalytic closure among them is achieved, the collective system of molecules is alive” (Kauffman 1995, p. 50). Kauffman sees catalytic closure as akin to autopoiesis, and he has suggested that his models of autocatalytic sets of proteins can serve as models for the molecular realization of autopoiesis (Brockman 1995, p. 217; Kauffman 1995, p. 274). But although there is a family resemblance between an autopoietic system and an autocatalytic set, the two are not equivalent (Maturana and Varela 1980, p. 94; McMullin 1999). The family resemblance is due to the second, reaction-network criterion. The difference is that in a collectively autocatalytic system there is no semipermeable boundary that spatially demarcates and contains the network (Criterion 1), and so the autocatalytic set cannot be regenerated by conditions due to such a boundary (Criterion 3). Hence an autocatalytic set does not qualify as a spatially distinct individual in the way an autopoietic system does.
A minimal autopoietic system is equivalent not simply to an autocatalytic network, but to an autocatalytic network housed within and interdependently linked to a semipermeable membrane boundary. The crucial property is that the membrane is not a mere containment device for the reaction network; rather, it is produced and maintained as a product of that network. This property is decisive for characterizing an autopoietic system as an autonomous individual (Varela 1979). A collectively autocatalytic system, because it lacks this property, does not qualify as a proper autonomous agent (pace Kauffman 2000; see Mc-Mullin 2001).
The three autopoietic criteria are clearly based on single-cell organisms. What about multicellular organisms? Are these also autopoietic systems?
In discussing this issue, Maturana and Varela (1987, pp. 87–89) distinguish between first-order and second-order autopoietic systems. Living cells are first-order autopoietic systems, whereas systems that include individual cells as structural components are second-order autopoietic systems. Maturana and Varela call such systems “metacellulars” and list as examples multicellular organisms, colonies, and societies. Given their definition of a metacellular as any unity in whose structure we can distinguish cell aggregates in close coupling, one could also include organs, such as the heart or liver. The crucial issue is whether any second-order autopoietic systems are also first-order autopoietic systems.
To answer this question it is not enough to point out that a second-order autopoietic system contains cells as elements of its structure, for the issue is whether the system’s proper components (whatever they are) realize the autopoietic organization. For instance, one might think that a multicellular organism such as an insect is a first-order autopoietic unity because its components, including those that make up its boundary, are being constantly replaced from within itself, subject to conditions imposed by the boundary. At the same time, an insect colony might be considered merely an aggregate of autopoietic systems, but not an autopoietic unity in its own right. The thought would be that because an aggregate of human bodies in a society is not a body at a higher level, an aggregate of autopoietic systems is not itself an autopoietic system. The problem is that for this intuitive differentiation to be rigorous, it must be grounded on an explicit account of the kinds of organization proper to metacelluar systems:
We know in great detail how a cell comes about as a molecular autopoietic unity, but how can we possibly describe in an organism the components and relations that make it a molecular autopoietic system? In the case of metacellulars, we are still ignorant of the molecular processes that would constitute those metacellulars as autopoietic unities comparable to cells . . . we shall leave open the question of whether or not metacellular systems are first-order autopoietic systems. What we can say is that they have operational closure in their organization: their identity is specified by a network of dynamic processes whose effects do not leave that network . . . whatever the organization of metacellulars may be, they are made up of first-order autopoietic systems and form lineages by reproducing through cells. These two conditions are sufficient to assure us that whatever happens in them, as autonomous unities, happens with conservation of the autopoiesis of their component cells, as also with conservation of their own organization. (Maturana and Varela 1987, pp. 88–89)
Given the knowledge we now have of molecular processes during the development of metazoan organisms (see Chapter 7), it seems reasonable to claim that metazoans are first-order autopoietic systems in addition to second-order ones. Nevertheless, difficulties remain, for in determining whether a given system qualifies as autopoietic, much depends on how we interpret “boundary” and “internal reaction network” in the criteria for autopoiesis. On the one hand, the theory of autopoiesis was formulated to apply to production processes in the molecular domain, and so allowing the boundary and network to be nonmolecular raises problems.3 Here Varela’s (1979) distinction between autonomy (organizational closure) and autopoiesis is pertinent. An autopoietic system is a specific kind of autonomous system—one having an organizational closure of production processes in the molecular domain—but there can be autonomous systems that are not autopoietic if their constituent processes exhibit organizational closure in their domain of operation. For example, an insect colony or animal group might qualify as autonomous in this sense. On the other hand, taking “boundary” to mean only a unicellular semipermeable membrane or even a multicellular epidermal layer seems too restrictive (plants and insects do not have a skin). Rather, the crucial matter is that the system produce and regulate its own internal topology and functional boundary, not the particular physical structure that realizes this boundary (Bourgine and Stewart 2004; Zaretzky and Letelier 2002).
According to the autopoietic criteria, the simplest living systems found on Earth are prokaryotic or bacterial cells. Unlike the structurally more complex eukaryotic cell, which contains a number of internal compartments, including a nucleus housing the cell’s chromosomes and DNA, the bacterial cell has no compartments, and its coiled DNA floats within the cell. Nonetheless, a bacterial cell is already a highly complex biological entity, in which some two thousand to five thousand genes and a similar number of proteins mutually produce each other within the cell membrane they fabricate together (Margulis and Sagan 1995, p. 50).
Could one model a more streamlined molecular system that meets the three autopoietic criteria? This question is relevant not only to modeling minimal autopoiesis, but also to research on the origins of life, for one central goal of this research is to explain how protocells could have arisen and eventually led to the complex living cells we see today (Morowitz 1992; Szostak, Bartel, and Luisi 2001).
One way to approach the topic of minimal autopoiesis is through computer models, an approach central to the field known as “Artificial Life” or “Alife.” One of the first computer models of minimal life was devised by Varela, Maturana, and Uribe (1974) some years before the field of Alife defined itself in the late 1980s (Langton 1989). This model has been revised and reimplemented by McMullin and Varela (1997).4 The qualitative chemistry of the model is inspired by and represents a simplification of the kind of chemical reactions found in living cells. The chemical domain of the model occurs in a discrete, two-dimensional space. This type of model is known as a tesselation automaton or two-dimensional cellular automaton. Each position in the space is either empty or occupied by a single particle. Particles generally move in random walks in the space. There are three distinct types of particles, and they engage in three distinct reactions (see Plate I):
• Production: Two substrate (S) particles may react in the presence of a catalyst (K) to form a link (L) particle.
• Bonding: L particles may bond to other L particles. Each L particle can form (at most) two bonds, thus allowing the formation of indefinitely long chains, which may close to form membranes. Bonded L particles become immobile.
• Disintegration: An L particle may spontaneously disintegrate, yielding two S particles. When this disintegration occurs, any bonds associated with the L particle are also destroyed.
Chains of L particles are permeable to S particles but impermeable to K and L particles. Hence a closed chain or a membrane that encloses K or L particles effectively traps them.
The basic autopoietic phenomenon predicted for this artificial chemistry is the possiblity of realizing dynamic cell-like structures that continually produce the conditions for their own maintenance. Such a system would consist of a closed chain or membrane of L particles that encloses one or more K particles. There can be continual production of L particles because they are formed by catalyzed reactions of the S particles that permeate through the membrane. Because they cannot escape through the membrane, L particles in relatively high concentration will build up inside the system. From time to time, however, the membrane will rupture as a result of the disintegration of its component L particles. Given the high concentration of L particles inside the membrane, the probability is high that one of these particles will drift to the rupture site and effect a repair, before the K particle escapes. In this way the conditions that allow for the buildup of a high concentration of L particles will be reestablished precisely.
A secondary phenomenon that might occur is the spontaneous establishment of a self-sustaining system from a randomized initial arrangement of the particles. Such spontaneous formation was reported in the original 1974 study by Varela, Maturana, and Uribe but was not studied by McMullin and Varela (1997). Instead, they artificially introduced a putative autopoietic entity into the system and investigated whether this entity could succeed in realizing the self-sustaining reaction network just described over a large number of time steps (the basic autopoietic phenomenon predicted for this artificial chemistry) .5
The qualitative chemistry presented in this section corresponds to the description originally given by Varela, Maturana, and Uribe (1974). Subsequent attempts to reimplement their work, however, led to the discovery of an additional interaction, present in their original program code but not documented in the description (McMullin 1997a, 1997b). This additional interaction prevents the spontaneous and premature bonding of L particles produced within the membrane. Such bonding makes the L particles immobile and unavailable to effect a repair to the membrane. McMullin and Varela (1997) call this additional interaction “chain-based bond inhibition” and describe it as follows: “This is an interaction whereby bonding is inhibited to any free L particle which is in the immediate vicinity of another L particle which is doubly bonded. In effect then, a free L particle cannot form a bond as long as it is alongside (as opposed to at the end of) an existing chain of L particles; but it can form bond(s) when it is at the end of a chain; and, especially, when it is positioned at a site where a chain has broken (i.e. a rupture site)” (McMullin and Varela 1997, p. 6). McMullin and Varela (1997) show that this interaction is crucial to realizing the basic autopoietic phenomenon in the model.
Plates II–V show one of the runs of the experiment, during which an autopoietic reaction network is established and a succession of successful membrane-repair episodes occurs. The morphology that is established seems particularly robust, persisting for approximately one thousand time-steps of the model. The initial configuration, illustrated in Plate II, is a single, artificially constructed, cell-like entity—a closed membrane of L particles housing a single K particle. It is embedded in a toroidal space. Between time 0 and time 226 the initial membrane suffers two ruptures, which are repaired with no change of membrane morphology. Between time 227 and 444 there are four rupture and repair episodes, yielding the new membrane morphology shown in Plate III. This new morphology appears to be relatively robust. It persists up to time 1250, while undergoing 5 more rupture and successful repair episodes. Between time 1250 and time 1310 there are two rupture and repair episodes, which produce the new morphology shown in Plate IV. The entity persists in this morphology through two more rupture and repair episodes, until time 1741. Then there are two ruptures in quick succession, at times 1742 and 1745. At time 1746 the membrane fragments, and partially spirals into the cavity, as shown in Plate V. It is then no longer possible to recover the closed membrane through any simple process of self-repair.
Does this self-sustaining system meet all the requirements for autopoiesis? This question is actually somewhat tricky. Varela, Maturana, and Uribe (1974, p. 191) state explicitly, “Within this universe [of the model] these systems satisfy the autopoietic organization” (see also Varela 1979, p. 20). But there is one component in the model—the K particle or catalyst—that is not itself produced by any reaction in the network. Bourgine and Stewart (2004) take this feature of the model to imply that the system is not fully autopoietic. McMullin (1999), however, citing Varela, Maturana, and Uribe’s (1974) criteria for determining whether a specific system is autopoietic, suggests that it may be acceptable for some of the components not to be produced by reactions in the system, as long as these components play a necessary and permanent role in the production of other components. For instance, substrate particles are not necessarily produced by the reaction network because they can traverse the membrane and enter the system from outside. Furthermore, in the case of the spontaneous generation of a self-sustaining network, the preexisting catalyst (K) would interact with the preexisting substrate (S), thereby forming a chain of bonded links (L particles). Eventually the chain would enclose the catalyst, and as a result the L particles produced within the enclosure would replace the decaying L particles of the boundary. Such a self-sustaining network would appear to satisfy the three autopoietic criteria.6
In a recent paper, Bourgine and Stewart (2004) expand the tesselation-automaton model from two to three dimensions and give a mathematical treatment of the model as a random dynamical system (see Figure 5.2). This approach enables them to model temporary holes in the membrane. In a two-dimensional model with a one-dimensional membrane, two holes would immediately disrupt the membrane, but in a three-dimensional model with a two-dimensional membrane, the membrane retains a topological continuity despite a substantial fraction of holes in the membrane. Their system is made up of three types of components: A-components or substrate molecules; B-components formed by a reaction between two substrate molecules A (A + A → B); and C-components belonging to the membrane, which has the form of a two-dimensional sheet and whose inside surface catalyzes the reaction A+A → B. Within the membrane, B-components diffuse freely. If two single B-components collide, they do not combine, but if a free B-component collides with the edge of a hole in the membrane, it attaches to the surface and repairs the hole (either partially or completely depending on the hole’s size). (This feature is analogous to chain-based bond inhibition.) A B-component that integrates the membrane in this way thereby becomes a C-component. C-components (at the membrane’s inside surface) in turn catalyze the production of B-components. In this way, the system avoids the need for a catalyst not produced by the system itself. (C-components are formed by B-components, and C catalyzes the production of B.) Finally, each of the C-components can disintegrate, and the end-product D cannot integrate the membrane, but escapes into the outside environment, leaving a hole in the membrane (or enlarging a preexisting one).
Figure 5.2. A schematic illustration of the 3D tesselation automaton model of autopoiesis. From Bourgine and Stewart (2004). The enlarged inset shows the processes occurring in a thin volume just under the membrane (catalytic production of B, and B-components entering the membrane to become C-components). The reaction C → D represents the disintegration of a C-component, leaving a hole in the membrane. B-components are normally confined by the membrane but can be lost through holes. a (a1-am) gives the net flux of A across the membrane. Reprinted with permission from Paul Bourgine and John Stewart, “Autopoiesis and Cognition,” Artificial Life 10:3 (Summer 2004), 330. © 2004 by the Massachusetts Institute of Technology.
In their mathematical treatment of this system as a random dynamical system, Bourgine and Stewart show that the crucial variable for understanding the structural dynamics of the system is the proportion of the total surface area occupied by C-components. When the value of this variable is above 50 percent, the system maintains itself dynamically, such that the rate of disintegration of the membrane (C → D) is balanced by the repair process (B → C). Below a critical point, the repair dynamics can no longer balance the formation of holes. The holes increase in size and number, there is vicious positive feedback, and the system collapses. The phase space of the system’s operation thus has two qualitatively different regimes and a critical point of bifurcation. As Bourgine and Stewart note: “Like a candle-flame, the system is either ‘alive’ or ‘dead.’ If it is alive, it may waver at times (several holes may appear), but it can recover and go on as though nothing has happened. If it is dead, nothing can resuscitate it; the system collapses and disintegrates entirely” (Bourgine and Stewart 2004, p. 332).
The models we have been examining indicate that it should be possible for the three autopoietic criteria to be realized in comparatively simple chemical systems. In other words, it should be possible to realize a self-sustaining reaction network, whose boundary is produced by the network and in turn encloses the network, in a chemical system much simpler than a bacterial cell. Yet these models are not real physical systems but either computationally simulated ones (McMullin and Varela 1997; Varela, Maturana, and Uribe 1974) or abstract systems described in the mathematical framework of dynamic systems theory (Bourgine and Stewart 2004). Although the three autopoietic criteria may be necessary and perhaps sufficient to specify the organization of minimal life, for some entity to count as a living system, embodiment is required. That is, the system has to satisfy the three autopoietic criteria in the physical space (Maturana and Varela 1980, pp. 84, 112; see also Boden 2000; Fleischaker 1988). This point raises the following question: what about minimal autopoiesis in a real chemical medium?
The creation of chemical models of autopoiesis has been the research goal of chemist Pier Luigi Luisi and his colleagues since the early 1990s.7 Working with micelles and vesicles, they claim to have succeeded in creating minimal autopoietic systems in the laboratory (Bachman, Luisi, and Lang 1992; Luisi 1993). These systems may shed light on various chemical processes of molecular self-assembly involved in the origins of life (Szostak, Bartel, and Luisi 2001).
Micelles and vesicles are three-dimensionally closed, molecular structures capable of spontaneous formation or self-assembly. Certain types of molecule, when dispersed in aqueous solutions, form clusters or aggregates in a predictable way. A micelle is a tiny droplet whose boundary is formed by such clustering. The molecules, known as surfactants (surface active agents), aggregate in the form of a sphere, thereby producing a bounded microenvironment (the interior of the droplet). In addition, certain molecules can self-assemble into double-layer aggregates called bilayers and thereby form vesicles, which are small, fluid-filled sacs with a bilayer boundary like a cell membrane. The molecular self-assembly of a bilayer membrane and its closure into a vesicle to form a “minimal cell” have served as the foundation for several scenarios for the origins of life (Fleischaker 1990a; Morowitz 1992; Morowitz, Heinz, and Deamer 1998; Szostak, Bartel, and Luisi 2001), in line with the tradition going back to the Russian scientist Alexander Oparin (Oparin 1938).
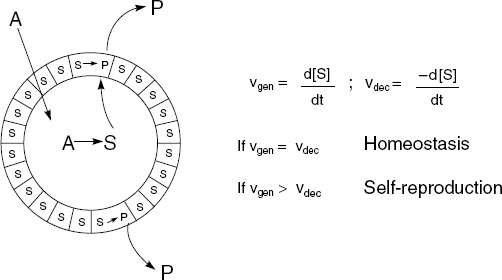
The basic idea behind using micelles and vesicles to create a minimal autopoietic system is to synthesize a bounded structure that hosts in its aqueous interior a chemical reaction that leads to the production of the surfactant, which in turn aggregates to form a boundary for the reaction. A schema for such a system is presented in Figure 5.3 (compare to Figure 5.2). One begins with a semipermeable-membrane boundary consisting of only one type of component S (Figure 5.3). A single type of nutrient A can permeate the membrane. Inside the system A is transformed into S. The system is characterized by two competitive interactions, “generation” and “decay.” Generation yields S from A, and decay breaks down S into a product P that leaves the boundary and enters the environment. When the velocities of these two generation and decay interactions are numerically equal, then the system is in a self-perpetuating steady state or homeostasis. When the velocity of generation is greater than that of decay, then the system can grow and eventually reproduce by splitting into two smaller systems. Finally, when the velocity of decay is greater than that of generation, then the system will become impoverished in the S-component and eventually implode.
Figure 5.3. The minimal autopoietic system. Two competitive reactions—one that builds the component of the boundary and another one that destroys it—characterize the system. According to the relative value of these two velocity constants, the system can be in homeostasis, or grow, or die. From Pier Luigi Luisi, “Autopoiesis: A Review and Reappraisal,” Naturwissenschaften 90 (2003): 53, fig. 4, © 2003 Springer-Verlag. Reprinted with kind permission of Springer Science and Business Media.
An example of the second, reproductive mode, implemented in vesicles, is shown in Figure 5.4. It begins (top left) with a relatively static aqueous vesicle formed by the surfactant S. Then a highly lipophilic precursor of S, indicated as S-S, binds to the boundary of the vesicle, where it is hydrolyzed, yielding the same surfactant S. The vesicle grows and eventually divides into two or more thermodynamically stabler and smaller vesicles. The whole process is autocatalytic: the more vesicles are formed, the more S-S is bound and the more vesicles are formed.
Figure 5.4. Autopoiesis and self-reproducing vesicles. Experimental procedure to obtain self-reproducing vesicles (liposomes). S-S represents the water-insoluble precursor of the surfactant, S, and S_S is hydrolyzed on the membrane of the micelles or of the vesicles. From Pier Luigi Luisi, “Autopoiesis: A Review and Reappraisal,” Naturwissenschaften 90 (2003): 56, fig. 7, © 2003 Springer-Verlag. Reprinted with kind permission of Springer Science and Business Media.
Do such micelles and vesicles qualify as minimal autopoietic systems? The answer is not entirely clear. On the one hand, these systems clearly meet Criteria 1 and 2: there is a semipermeable boundary, and the boundary is produced by the system itself. There is a clear analogy between the circular organization of these systems and that of an autopoietic minimal cell. On the other hand, it is unclear whether these systems fully meet Criterion 3—the presence of an internal reaction network that regenerates itself and the boundary. As Luisi observes: “Reaction takes place on the boundary, which is part of the system, but a restricted one: the internal aqueous core is not part of the reaction system, and furthermore we are dealing with only one reaction instead of an internalized reaction network” (Luisi 2003, p. 56). (The same point can also be raised for Bourgine and Stewart’s three-dimensional tesselation automaton.) Luisi maintains that these systems are the simplest possible case of experimental autopoiesis, albeit a limiting case (Bitbol and Luisi 2005). One could argue, however, that they are better described as proto-autopoietic than minimally autopoietic. Which description one chooses depends on how one evaluates these systems with respect to the third criterion. As we will see in the last section of this chapter, this issue also bears on whether autopoiesis is necessary and sufficient, or merely necessary, for minimal life.
To close this section, let me briefly connect the idea of minimal autopoiesis to research on the origins of life on Earth. In 1985, Freeman Dyson in his book Origins of Life (1985) pointed out that life as we know it requires both metabolic self-maintenance and the replication of nucleic acids. He proposed that living cells arose from the “symbiotic” merger of two independently evolved, prior systems—one a self-replicating system composed of nucleic acids and the other a self-maintaining, autocatalytic system of proteins. From an autopoietic perspective, however, the crucial step required for biological individuality—for an individual living system, here and now, or there and then—is that such subsystems be enclosed within a membrane that they produce together. As Margulis and Sagan observe, prior to this step neither subsystem is alive, and after this step it is the whole system that is living (and hence the word “symbiosis” to describe their merger is not quite right) (Margulis and Sagan 1995, p. 60).
In Beginnings of Cellular Life: Metabolism Recapitulates Biogenesis, Morowitz (1992) proposes that many features of nucleic acids and proteins evolved only after these molecules came to be enclosed within membrane-bounded units or protocells. Consistent with an autopoietic perspective, he emphasizes the importance of the membrane for establishing and stabilizing life: “It is the closure of an amphiphilic bilayer membrane into a vesicle that represents a discrete transition from nonlife to life” (Morowitz 1992, p. 9). He distinguishes between “Ur-cells” (or protocells) and ancestor cells. Vesicle formation is the key event in the transition from nonlife to life at the protocell stage. We do not know how many times this happened—how many independent origins of protocellular life there might have been. We do know, however, that all contemporary life derives from a universal ancestor, which could have been a single cell or population of cells. Thus the problem of the origins of life comprises both the chemical formation of the Ur-cells and the subsequent evolutionary sequence that gave rise to the universal ancestor (Morowitz 1992, p. 88). In Morowitz’s scenario, protocells in the form of vesicles originated first, a simple form of metabolism or internal reaction network came next, then proteins developed, and genes arrived last.
The first step was the formation of vesicles in the ocean at the water-atmosphere interface some 4 to 3.8 billion years ago. This event created a three-phase system of an interior, boundary, and exterior. In the autopoietic viewpoint, this step corresponds to Criterion 1, the establishment of a semipermeable boundary separating inside from outside. The second step was the development of a simple metabolic reaction network inside the vesicles. This network could synthesize the molecules needed for self-maintenance, including the molecules of the membrane boundary. This second step corresponds to Criteria 2 and 3 and thus to an autopoietic protocell. Morowitz hypothesizes that the metabolism of these early cells was not protein-based. The protocells were also potentially self-reproducing and hence would have proliferated. What then ensued was the first evolutionary radiation leading from the protocells to the universal ancestor. This transition involved both the formation of amino acids, as a result of the protocellular metabolism becoming more complex, and the origin of nucleic acids and the so-called genetic code.
Morowitz’s scenario is clearly congenial to the autopoietic view of life. The details, of course, require experimental verification. But what matters to me here are not so much the details of any single origins-of-life scenario, but rather the overall theoretical framework. In an autopoietic framework, minimal life is not identified with any particular molecular structure, such as RNA/DNA, but rather with a bounded, self-producing concatenation of processes, which can in principle be structurally realized in different ways.
Earlier in this chapter I mentioned three different ways of characterizing life—the evolutionary, ecological, and individual characterizations. The focus of this chapter is the individual characterization. The autopoietic characterization of life highlights the organization of an individual living system. Models of minimal autopoiesis try to shed light on how such a system could emerge from a prebiotic chemical environment. We should not lose sight, however, of the ecological context of autopoiesis. As Morowitz writes:
When we speak of the origin of life, we are referring generally to the rise of organisms . . . It is well to remember, however, that this can take place only within a global ecological system. In a certain sense, the origin of life refers to a transformation of the planetary surface from an inorganic state to one dominated by the activities of plants, animals, and microbes. (Morowitz 1992, p. 6)
Traditional biology has tended to concentrate attention on individual organisms rather than on the biological continuum. The origin of life is thus looked for as a unique event in which an organism arises from the surrounding milieu. A more ecologically balanced point of view would examine the protoecological cycles and subsequent chemical systems that must have developed and flourished while objects resembling organisms appeared. (Morowitz 1992, p. 54)
Two points are important here. First, autopoiesis always has to be ecologically embedded. “Self-producing” refers to the kind of circular organization that makes the cell an individual; it does not mean that the cell makes itself apart from its environment. Second, although life may have begun in small, local pockets, it quickly proliferated, evolved, and transformed the entire surface of the planet. Life as we know it is a phenomenon of evolutionary and planetary scale. Take, for example, the prokaryotic bacteria, which according to the autopoietic criteria are the simplest living beings on Earth. Bacteria are hardly isolated self-makers. On the contrary, they never live as isolated individuals in nature; rather, they form microbial communities or associations (Margulis and Sagan 1986). Moreover, they can pass their genes among each other without reproducing, in effect sharing a common potential genome. For these reasons, some biologists have argued that all bacteria form one organism, or superorganism:
[B]acteria, in addition to carrying out their individual and localized team activities, together form a planetary entity of communicating and cooperating microbes, an entity that, we think, is both genetically and functionally a true superorganism. It is at the social level—the level of their associations—that bacteria manifest their exceptional capacities and play a major role in nature. (Sonia and Panisset 1983, p. 8)
The idea that life is a planetary phenomenon is central to the Gaia theory advanced by chemist James Lovelock and microbiologist Lynn Margulis (Lovelock 1979, 1988; Margulis 1998). Their theory states that the biota (totality of living organisms), atmosphere, oceans, rocks, and soils make up a single planetary entity that is self-regulating and self-maintaining. They call the entity Gaia, which Lovelock often describes as a planetary organism or superorganism. The theory is controversial, in part because of the issues about life discussed in this chapter. For this reason, it is worth considering the theory here.
When Lovelock and Margulis first proposed their theory in the 1970s, they called it the Gaia hypothesis (Lovelock and Margulis 1974; Margulis and Lovelock 1974). The hypothesis was that the physical and chemical conditions of the Earth’s surface, atmosphere, and oceans are kept within limits favorable to life and that this homeostasis is accomplished automatically by the biota through negative feedback processes. Since this original formulation, Lovelock and Margulis have expanded the hypothesis into a theory. According to the Gaia theory, the biota and their material environment evolve together, as a single “tight-coupled system,” with the self-regulation of the planetary climate and atmosphere occurring as emergent phenomena. Not only is life a planetary phenomenon, but the material environment of life on Earth is in part a biological construction. As Lovelock puts it: “In no way do organisms just ‘adapt’ to a dead world determined by physics and chemistry alone. They live with a world that is the breath and bones of their ancestors and that they are now sustaining” (Lovelock 1991, p. 32). Lovelock and Margulis believe that this planetary system, Gaia, is the largest known biological individual:
Gaia is best thought of as a superorganism. These are bounded systems made up partly from living organisms and partly from nonliving structural material. A bee’s nest is a superorganism and like the superorganism, Gaia, it has the capacity to regulate its temperature. (Lovelock 1988, p. 15)
The assertion that Gaia is a planetary superorganism has not gone without criticism. Some of the main early criticisms came from the biologists W. Ford Doolittle and Richard Dawkins (Dawkins 1982, pp. 234–237; Doolittle 1981, 1987). They argued that Gaia should not be accorded the status of a living system because it is not a reproducing individual and hence has no hereditary lineage. They also maintained that it is unlikely that the planetary self-maintaining mechanisms that Lovelock and Margulis hypothesized could result from natural selection, and so if they exist they are “accidental.”
These criticisms bring us back to the issues about life. Doolittle and Dawkins were looking at life from the perspective of evolution, reproduction, and genetics. Is this the right perspective for answering the question of whether there is a planetary biological individual? I do not think so. The right perspective is that of the individual characterization of life, the one that figures in our examination of minimal life in this chapter. As we have seen, at this level reproduction does not enter as a defining feature of the organization of a living system. Reproduction requires an individual to be reproduced, and hence is logically and empirically secondary to the processes whereby an autonomous system constitutes itself as an individual. Therefore, a self-producing but non-reproducing planetary system could count as a genuine biological individual. Furthermore, its self-producing and self-maintaining mechanisms, though not the result of natural selection acting on a population of similar systems, would nevertheless hardly be “accidental.” Indeed, they would presumably reflect lawful principles of self-organization and emergence in complex systems.8
If Lovelock and Margulis are right, what sort of biological entity would Gaia be? Lovelock describes Gaia as a superorganism. Margulis describes Gaia as an autopoietic system like a cell:
The simplest, smallest known autopoietic entity is a single bacterial cell. The largest is probably Gaia. Cells and Gaia display general properties of autopoietic entities; as their surroundings change unpredictably, they maintain their structural integrity and internal organization, at the expense of solar energy, by remaking and interchanging their parts. Metabolism is the name given to this incessant activity. (Margulis 1991, p. 237)
Margulis’s words echo something Lewis Thomas wrote in his book The Lives of a Cell, around the same time that Lovelock first announced the Gaia hypothesis:
I have been trying to think of the earth as a kind of organism, but it is no go. I cannot think of it this way. It is too big, too complex, with too many working parts lacking visible connections. The other night, driving through a hilly, wooded part of southern New England, I wondered about this. If not like an organism, what is it like, what is it most like? Then, satisfactorily for that moment, it came to me: it is most like a single cell. (Thomas 1974, p. 5)
Should Gaia, the whole ecosphere of the planet Earth, be considered an autopoietic system? Margulis has been a long-standing advocate of autopoiesis as the most adequate characterization of life at the individual level. Her work on symbiosis, cell evolution, and the Gaia theory has greatly enriched the autopoietic perspective (Margulis 1993, 1998; Margulis and Sagan 1995, 2002).9 Yet whether we should count Gaia as an autopoietic system is a difficult question. The crucial issue is whether “boundary” and “internal reaction network” have a clear interpretation when we shift from the molecular to the planetary scale. We have already had occasion to notice that interpreting “boundary” to mean only semipermeable membrane or skin is too narrow. Rather, what is important is that the system produce and regulate its own internal topology and functional boundary, in whatever domain the system operates. Lovelock and Margulis make a strong case for considering the ecosphere to be a large-scale system that produces its own internal topology and functional boundary, actively regulates its boundary conditions, and continually produces the components that in turn produce it.
But to make this intuitively compelling picture precise would require both more detailed knowledge of the “geophysiological” processes that produce and realize Gaia, and more precise elaborations of the three autopoietic criteria in nonmolecular domains. As Bourgine and Stewart remark: “It must be an open question at present as to whether the terrestrial ecosphere actually is a bona fide autopoietic system; to answer this question, it will be necessary to put our mathematical definitions of autopoiesis in full working order, and then to carry out the considerable task of applying them to the case of the ecosphere. However, we can already say at this stage that we certainly do not want to definitely rule out this possibility, simply because the ecosphere does not have a single clearly reified membrane” (2004, p. 337).
In one of his articles Lovelock uses the term ecopoiesis to describe Gaia (Lovelock 1987). This term seems just right for conveying both the resemblance and difference between Gaia and the autopoietic cell. The resemblance is due to the ecosphere and the cell being autonomous systems, the difference to the scale and manner in which their autonomy takes form.
According to Maturana and Varela, “the notion of autopoiesis is necessary and sufficient to characterize the organization of living systems” (1980, p. 82; emphasis omitted). They also state: “Autopoiesis in the physical space is necessary and sufficient to characterize a system as a living system” (p. 112). My aim in the final section of this chapter is to evaluate these necessity and sufficiency claims.
The necessity claim can be expressed as the proposition that for an individual entity, here and now, to be characterized as a living system, it must realize the autopoietic organization, that is, satisfy the three autopoietic criteria. In other words, no system that deserves, here and now, to be called living can fail to be autopoietic. The way to test or challenge this claim is to look for a putative counterexample—that is, a system that deserves, here and now, to be called living but is not autopoietic. Given that the criteria for autopoiesis are based on living cells and that multicellular organisms and colonies are considered to be higher-order autopoietic systems, such a counterexample would have to come from outside the domain of cells. Two candidates that come to mind are viruses and replicative molecules such as RNA and DNA.
As we saw earlier in this chapter, a virus does not meet the autopoietic criteria. It does not produce from within itself its own protein coat or nucleic acids. Rather, these are produced by the host cell in which the virus takes up residence. Nonetheless, viruses are often described as living. Do they therefore pose a challenge to the necessity claim? It is important to be clear that this issue is not about the meaning of the everyday word “living.” The issue is whether the theoretical concept of autopoiesis succeeds in characterizing the organization of those systems that are the definitive objects of biological investigation. Viruses are clearly objects of microbiological research. Therefore the issue comes down to whether the theory of autopoiesis can provide a good reason not to characterize them as living systems.
Such a reason is not hard to find. Viruses are not dissipative systems or metabolic entities. They do not exchange matter or energy with the environment. Outside of a host cell a virus is completely inert and is entirely subject to the vicissitudes of the environment. Inside a cell it makes use of the cell’s metabolism, but it has no metabolism of its own. A virus is thus a fundamentally different kind of physicochemical entity from both prokaryotic cells or bacteria and autocatalytic protocells. The theory of autopoiesis succeeds in capturing this difference at the organizational level. Hence viruses do not pose a compelling challenge to the claim that autopoiesis is necessary to characterize the organization of a living system.
What about replicative molecules? One might argue that minimal life can be characterized or defined in terms of RNA and DNA and the molecular structures they form. For instance, certain RNA strands known as ribozymes can act as catalysts for their own replication. Ribozymes are remarkable because they have both catalytic abilities like enzymes and the template specificity of nucleic acids. In a test-tube they rapidly evolve self-replicating patterns. If such molecules deserve to be described as living, then autopoiesis is not necessary to characterize a system as living.
This line of thought presents a problem, however: it shifts tacitly from the individual, here-and-now account of life to the population and genetic-evolutionary account. A single RNA molecule does not catalyze its own replication and evolve by natural selection. Autocatalytic replication requires a large family of molecules, and evolution by natural selection requires a reproductive population. Thus the molecular description derives from the evolutionary, population level and so has no force as a counterargument to the autopoietic account at the individual level. Furthermore, the population and evolutionary perspective presupposes the individual perspective: a population is by definition composed of individual members, and evolution requires reproductive individuals. The population perspective thus cannot characterize life at the level of single individuals, here and now, especially individuals that have no genetic heritage or lack genetic material, such as synthetic laboratory entities or early terrestrial protocells (Luisi 1998).
Let me turn now to the sufficiency claim, which states that if something displays autopoiesis, then it is living. This claim has recently been called into question by two groups (Bitbol and Luisi 2005; Bourgine and Stewart 2004). It is significant that both groups are important proponents of autopoiesis and that they offer essentially the same argument: all living systems are both autopoietic systems and cognitive systems, but an autopoietic system is not necessarily a cognitive system.
This argument raises the question of the relationship between autopoiesis and cognition. Maturana and Varela (1980) originally proposed that all living systems are autopoietic systems and that all autopoietic systems are cognitive systems. “Autopoiesis” pertains to the self-producing organization of a living system, whereas “cognition” pertains to the behavior or conduct of a system in relation to its environment. According to Maturana and Varela (1980), the relation between autopoiesis and cognition has two crucial features. First, the instantiation of the autopoietic organization in an actual, concrete system entails a cognitive relation between that system and its environment. Second, this cognitive relation reflects and is subordinated to the maintenance of autopoiesis. As Maturana put it in a groundbreaking early paper: “A cognitive system is a system whose organization defines a domain of interactions in which it can act with relevance to the maintenance of itself, and the process of cognition is the actual (inductive) acting or behaving in this domain. Living systems are cognitive systems, and living as a process is a process of cognition. This statement is valid for all organisms, with and without a nervous system” (Maturana 1970, p. 13; emphasis omitted).
Bourgine and Stewart (2004) and Bitbol and Luisi (2005) accept the proposition that all living systems are cognitive systems. What they challenge is that autopoiesis entails cognition. Bourgine and Stewart take their three-dimensional tesselation automaton to be a minimally autopoietic system but not a cognitive one. Bitbol and Luisi consider autocatalytic micelles and vesicles to be minimally autopoietic systems but not cognitive ones. In each case, the system autocatalytically produces its own boundary but does not actively relate to its environment (see also Ruiz-Mirazo and Moreno 2004, pp. 244–245). There is nothing comparable to the taxic responses of microorganisms, tropisms of plants, or sensorimotor activities of animals. Hence these systems need not be described as having what Maturana and Varela call a “cognitive domain”—a domain of interactions or niche, defined by the system’s organization, in which the system operates or behaves so as to maintain its viability, that is, its autopoiesis (Maturana 1970, pp. 10–11; Maturana and Varela 1980, p. 119).
For Bourgine and Stewart, cognitive interactions are those in which sensory responses guide action and action has consequences for subsequent sensory stimulation, subject to the constraint that the system maintain its viability. “Sensory response” and “action” are taken broadly to include, for example, a bacterium’s ability to sense the concentration of sucrose in its immediate environment and to move itself accordingly. For a living system, viability requires maintaining autopoiesis. In the case of the tesselation automaton, however, there is no behavior guided by sensation. The system, though minimally autopoietic, is not cognitive and therefore is not a living system.
For Bitbol and Luisi, “cognition is tantamount to metabolism.” They distinguish two steps or levels of cognition. The first corresponds to the normal metabolic assimilation of select compounds from the environment. The second corresponds to the incorporation of novel elements that effect a change in the structure of a living system—that is, to the existing metabolism but not to the system’s organization. They liken the first step to Piaget’s notion of assimilation, and the second to his notion of accommodation. In the autocatalytic micelles and vesicles, however, there is no metabolic network. Instead there is only a single type of autocatalytic reaction that takes place at the boundary of the entity, and the internal aqueous core is not part of this reaction system. These chemical systems, though autopoietic, do not arrive at even the first stage of cognition as metabolic assimilation and therefore are not living systems.
On the basis of these considerations, Bourgine and Stewart, as well as Bitbol and Luisi, conclude that autopoiesis is a necessary but not sufficient condition for a system to be a living system, whereas the union of autopoiesis and cognition is both necessary and sufficient.
The main question to raise about this line of thought is whether the tesselation automaton and the autocatalytic micelles and vesicles really qualify as bona fide minimal autopoietic systems rather than proto-autopoietic ones. (The tesselation automaton is of course an abstract model, not a real physicochemical system, but in its architecture and dynamics it is close to the micelles and vesicles.) One could argue that a minimal autopoietic system requires more than merely a closed and self-producing boundary; it also requires a distinct internal reaction network that regenerates both itself and the boundary, and regulates the system’s interactions with the outside environment by way of that boundary. This interpretation of the criteria for autopoiesis is probably closer to Maturana and Varela’s original formulation. Nevertheless, Varela maintained that the autocatalytic micelles and vesicles are minimal autopoietic systems. Also, he did not hesitate to call them living (Luisi 2003; Luisi and Varela 1989).
It would seem, therefore, that we are faced with the following choice. We can maintain that the tesselation automaton and the autocatalytic micelles and vesicles are models of minimal autopoiesis. We can also state that autopoiesis in the physical space is necessary, but not sufficient, to characterize a system as a living system. Or we can claim that these models are not autopoietic but only proto-autopoietic, for autopoiesis requires a distinct internal reaction network that regenerates both itself and the boundary, and regulates the system’s interactions with the outside environment by way of that boundary. On this option, autopoiesis in the physical space entails being a cognitive system, and is both necessary and sufficient to characterize a system as a living system.
Let us remind ourselves of what cognition means in this context. Cognition is behavior or conduct in relation to meaning and norms that the system itself enacts or brings forth on the basis of its autonomy (see Chapter 4).10 Another way to put the issue before us, therefore, is whether autopoiesis is sufficient for cognition. If autopoiesis is taken narrowly to mean internal self-production of the minimal sort that does not involve active interaction with the outside environment, then autopoiesis is not sufficient for cognition. But if autopoiesis is taken more widely to mean internal self-production sufficient for constructive and interactive processes in relation to the environment, then autopoiesis does entail cognition.
I will not presume to legislate this matter: I will leave it for the scientists to work out in whichever way is most productive for their research. For my purposes it is enough to maintain that any living system is both an autopoietic and a cognitive system, for this thesis is sufficient to establish a deep continuity of life and mind. The task before us now is to examine this deep continuity in greater detail.