12 Experiencing and Regulating Desire
In their everyday lives, people spend a lot of time experiencing and dealing with desires of all sorts. Examples include innate desires for food, drink, sleep, sex, and social connection as well as acquired desires such as for alcohol, cigarettes, and media. Whereas many desires are unproblematic and adaptive, sometimes desires can conflict with important self-regulatory or moral standards, goals, and values. Consider the alcoholic who is fully aware of the fact that giving in to the desire for another drink at the party he is attending will likely impair his ability to drive home safely. In addition to personal costs (e.g., receiving a ticket for drunk driving), the societal consequences of poor desire regulation can be enormous. According to some estimates (Schroeder, 2007), 40% of deaths in the United States each year can be attributed to behaviors that are at least partially due to the way people (mis)manage desires, including those for unhealthy foods, tobacco, alcohol, unprotected sex, aggressive urges, and illicit drugs. It is therefore important to gain a better understanding of the psychological mechanisms that contribute to successful and unsuccessful desire regulation. It is clear that some people manage the pitfalls of desire better than others. So which individual-difference factors set successful and unsuccessful individuals apart from each other?
The present chapter is centered on the concept of desire. We will begin with a brief definition of desire and draw important links between desire and emotion. Next, we sketch the dynamical nature of desire and the various routes by which it can impact behavior, integrating neuroscientific insights. We will then provide a tripartite taxonomy of different desire-regulation strategies, including (a) early-stage preventive strategies such as situation control, stimulus control, and early distraction; (b) middle-stage down-regulatory strategies such as cognitive reappraisal, suppression, and acceptance; and (c) late-stage inhibitory strategies and mechanisms such as inhibition, overriding, and moderation. This taxonomy is then used in mapping the potential effects of various individual difference variables regarding desire regulation. To approach these issues, we will integrate insights from the traditional self-control literature, cognitive psychology, especially the elaborated intrusion theory of desire (Kavanagh et al., 2005) and executive functioning research, the cognitive neuroscience of reward processing, and the field of emotion regulation (e.g., Gross, 1998) – all of which are valuable, in our view, in moving toward a balanced understanding of desire and desire regulation.
Defining Desire and Related Concepts
Because the term desire refers to all kinds of wishes and wants in everyday language, some clarifications are in order. When we speak of ‘desire’ here, we refer to the narrower sphere of appetitive desire, which can be defined as a ‘feeling of wanting that propels us to approach and consume objects or otherwise engage in activities that satisfy a need and, in doing so, yield a gain in immediate pleasure (or relief from discomfort)’ (Hofmann and Nordgren, 2015: 5). Appetitive desires are typically rooted in physiological need states, such as for food, alcohol, sex, rest, or acquired through a history of reinforcement learning as in the case of drugs, media addiction, spending urges, etc. Moreover, we use the term ‘craving’ to refer to desires across domains that are particularly high-intensity (e.g., drug craving, food craving). It is also important to note that ‘desire’ and ‘temptation’ are not synonymous terms (Hofmann and Kotabe, 2013; Hofmann and van Dillen, 2012). Rather, we refer to ‘temptation’ as a desire that stands in conflict with an important higher-order goal or standard. This is highly related to Mele's (2001) point that to say that somebody is ‘tempted’ by something implies that the person has a desire to do X on one hand and simultaneously has a good reason not to do X, because whether a person has good reason not to do X will typically be a function of the person's background set of endorsed self-regulatory goals, values, or otherwise activated competing motivations.
Moreover, in this chapter and elsewhere (Hofmann and Kotabe, 2013), we draw a strong conceptual analogy between desire and emotion: similar to emotions, desires are multi-faceted phenomena combining affective, motivational, and cognitive components. Regarding the affective component, desires consist of a phenomenological feeling of ‘wanting’ of varying intensity. The affective component has its primary basis in the in-the-moment computation of reward value that condenses past positive experiences into anticipated positive affect.1 Regarding the motivational component, desires prepare and motivate behavior. Desiring something means wanting to have, consume, or do something that is expected to yield pleasure (or reduce discomfort). Moreover, because desires are directed toward certain objects in the environment, like emotions, they share a sense of ‘aboutness’ (e.g., being happy, sad, angry, disgusted about something) that distinguishes them from more diffuse affective states such as mood (Higgins, 1998). Regarding the cognitive component, there is a close connection between desires and desire-related cognition (mirroring the affect-cognition interface of emotion). According to the elaboration-intrusion theory of desire (Kavanagh et al., 2005), desire is typically accompanied by intrusive thoughts (including fantasies) about the object of desire. Moreover, the interplay between the strength of a given desire and these cognitions is bi-directional and dynamical – as a person engages in more and more cognitive elaboration, the strength of the desire typically increases and more and more mental resources are allocated to it in working memory. This is why desires can sometimes escalate to the point where they fully ‘crowd out’ opposing mental representations such as those of self-regulatory goals (Hofmann et al., 2011; Hofmann and van Dillen, 2012; Kavanagh et al., 2005).
Desire Dynamics
What renders some things so attractive that they grab attention, occupy thoughts, and fuel desire? Theories of desire need to address the often complex interplay of stimulus features in the environment and people's inner predispositions (i.e., need states, learning history) that provide the background against which these external stimuli can have their powerful effects. Due to the evolutionary old nature of these systems, these models should be able to account for relatively automatic processes generated by evolutionarily old reward-processing regions in the brain (e.g., Heatherton and Wagner, 2011). At the same time, however, they should be able to integrate the previously mentioned idea that desire, once emerging in the global workspace of consciousness, may dynamically affect behavior in qualitatively different terms through escalatory mechanisms (Kavanagh et al., 2005).
We have recently attempted to integrate these various aspects into a dynamical model of desire (DMD; Hofmann and van Dillen, 2012), illustrated in Figure 12.1. According to the DMD, desire is assumed to emerge in a relatively automatic fashion as reward-processing centers evaluate external stimuli against the backdrop of internal need states and an individual's learning history (Hofmann et al., 2009b; Hofmann and van Dillen, 2012). The DMD thus predicts that strongest reward signals should emerge when (a) relevant stimuli are present and relevant stimulus features are salient, (b) the respective need state is high, and (c) the individual displays high reward sensitivity with regard to that stimulus – an individual predisposition that can be the result of either genetic factors or learning history (or their interplay). In a recent set of studies, we found good support for the predicted three-way interaction among stimulus exposure, need state, and learning history as a measure of predisposition (Ghoniem and Hofmann, 2016a).
Figure 12.1A dynamical model of desire proposed by Hofmann and van Dillen (2012; adapted and reprinted with permission)
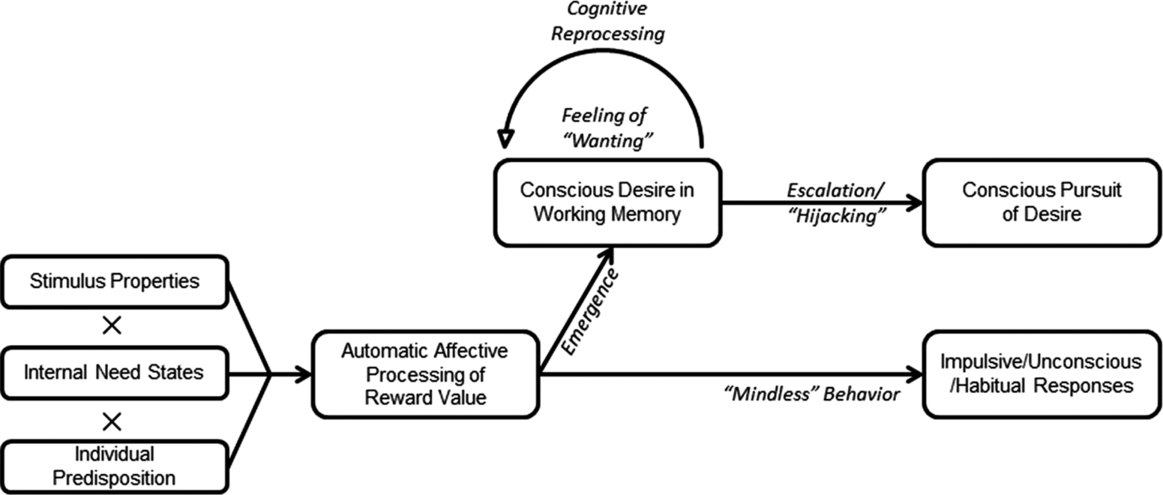
Early reward processing may have the potential to bias attention (e.g., Vollstädt-Klein et al., 2012) and trigger fast, impulsive, and habitual responses that may even happen outside of conscious awareness (e.g., Mogenson et al., 1980; Winkielman et al., 2005). Such impulsive responses may be most impactful in situations where the desired stimulus is already quite close in space and time (see Figure 12.1, lower pathway). However, desire may also gain access to consciousness and thereby deeply influence thinking and planning even when there is no direct opportunity for giving in to the desire. In case enough attention is allocated to it, desire may emerge into consciousness, thus occupying limited working memory resources. As a given desire attracts further attention, a ‘vicious cycle’ of reprocessing and rumination can result, during which people repeatedly imagine desire enactment and have recurring thoughts about the resulting pleasurable experience (Kavanagh et al., 2005). This is a vicious cycle inasmuch as cognitive elaborations can, in turn, increase the feeling of wanting and the motivational power of desire. As desire becomes more cognitively elaborated in working memory, so does its potential to instigate concrete action plans and behavioral intentions to consume the object of desire (see Figure 12.1, upper pathway). Crucially therefore, elaborated desires may predispose the organism toward (sometimes problematic) consumption via two important mechanisms. First, elaborated desires may crowd out (i.e., temporarily deactivate) other representations from working memory, leading to a preoccupation with the desire at the expense of everything else, including self-regulatory goals and values (Hofmann et al., 2011; Kemps et al., 2008a). Second, elaborated desires may instigate processes of motivated reasoning that license and justify indulgence (e.g., ‘I worked so hard today that I deserve a special treat'; ‘Others are having an ice cream, too'; ‘This is definitely going to be my last drink before I quit!') (De Witt Huberts et al., 2012, 2014; Hofmann and van Dillen, 2012). It is via these cognitive-motivational processes underlying desire escalation – thus ‘hijacking’ behavior through the same executive mechanisms that may otherwise support reasoned action – that desire may lead to passionate behavior that people later regret.
The introduced model places some emphasis on the emergence of desire, rather than just the regulation thereof. We have repeatedly argued that both motivational ‘forces’ – desire strength and the restraining force – are important in understanding the dynamics of desire regulation and self-control (e.g., Hofmann et al., 2009b; Kotabe and Hofmann, 2015). From this perspective, it is important to better understand individual-difference variables affecting the intensity of desire experiences. For instance, there is substantial evidence that individuals high in Behavioral Approach System (BAS) sensitivity experience stronger desires and cravings on average than those low in BAS. This effect on reward sensitivity was found both in cue-reactivity paradigms in the laboratory (Franken, 2002) and also when aggregating average self-reported desire strength across many desire domains (Hofmann, Baumeister et al., 2012). These findings clearly support Gray's (1982) Reinforcement Sensitivity Theory arguing that the BAS is responsible for the reward processing in appetitive motivation and suggest an important role for related constructs such as sensation seeking.
From a neuroscientific perspective, such differences in desire strength appear to be based on activity in largely subcortical neural regions that include the mesolimbic dopamine system and which are equally responsive to natural rewards like food and sex, cocaine, or gambling (Kelley et al., 2005). On the one hand, hypo-dopaminergic functioning has been thought to invoke deficiencies in reward responsiveness and to underlie a range of addictions including alcoholism, cocaine abuse, pathological gambling, and the development of obesity, as people seek out more extreme thrills to compensate (Bowirrat and Oscar-Berman, 2005; Wang et al., 2004). On the other hand, enhanced dopamine functioning has been found to elicit strong appetitive responses. For instance, amplification of the dopamine signal in human participants via a small dose of oral methylphenidate increased their desire to eat in response to a palatable food cue (Volkow et al., 2002). Whether linked to hypo- or hyper-dopaminergic functioning, it has, however, become clear that reward sensitivity deficiencies play an important role in the emergence of pathological desires that play a role in obesity, substance abuse, and gambling (for a more detailed discussion of hypo- versus hyper-responsivity in relation to obesity, see Stice et al., 2009).
Of particular interest, the nucleus accumbens (NAcc) in the ventral striatum seems to be implicated in the generation of wanting experiences (Berridge et al., 2009; Peciña and Berridge, 2005). For instance, one study connecting laboratory brain data and everyday life desire experiences, showed that individual differences in NAcc activity during exposure to food cues in the scanner were reliably related to the average strength of food desires in people's everyday environments, as measured via experience-sampling (Lopez et al., 2014). Likewise, another study demonstrated that individual differences in NAcc responsivity to food and sexual pictures predicted subsequent weight gain and sexual activity six months later (Demos et al., 2012). Neural responses to oral calories in reward-related brain areas have also been found to relate positively to individual differences in reward sensitivity as measured by the BAS scale (van Rijn et al., 2015), and individual differences in food reward sensitivity have been found to predict enhanced NAcc activity in response to high-calorie compared to low-calorie food images (van Dillen and van Steenbergen, 2018).
One possible answer for such individual differences in general or domain-specific differences in reward sensitivity may be found in genetic differences. The presence of the A1 allele of the Taq1A gene has, for example, been linked to lower density of D2 dopamine receptors, which is thought to underlie hypo-responsiveness to reward as observed in the abuse of cocaine (Goldstein et al., 2007) and alcohol (Martinez et al., 2005). In one neuroimaging study, moreover, presence of the A1 allele moderated the relation between abnormal dorsal striatum activation in response to food receipt and weight gain over the subsequent one-year period (Stice et al., 2009). Another answer for differences in reward sensitivity may be found in different learning histories through which people acquire learned expectancies that a given stimulus will provide pleasurable, rewarding experiences or relief from discomfort (as is often the case in addiction). Similar to the presence of the A1 allele for example, animal studies have shown that repeated intake of sweet and fatty foods reduces the density and sensitivity of post-synaptic D2 receptors (Warwick and Synowski, 1999).
Why Regulate Desires?
On the one hand, desires are key motivators, often rooted in evolutionary old systems that secure one's own survival as well as the survival of the species. On the other hand, there is a multitude of good reasons for why people are often motivated to regulate their desires. At a general level, the unfettered expression of desire can either be self-harming or interfere with the well-being of fellow citizens (or both). For instance, excessive alcohol consumption may not only harm one's own body, but also have a disintegrating effect on one's family members (e.g., impulsive acts of family violence, failing to maintain a job). The capacity to regulate desire is a prerequisite for participating in society, and those who chronically fail to regulate their desires such as aggressive and sexual urges often find themselves removed from free society. Not surprisingly, legal, religious, and educational systems are all involved in the regulation of problematic desires. The ideal goal of such regulative attempts is to establish a consensual balance between individual liberty and collective interests, including public safety, order, and health.
But even beyond those institutionalized normative influences, there are often additional moral, health-related, or financial considerations that keep desires at bay. According to moral foundations theory (Graham et al., 2013), for instance, morality is based on several core moral principles: Care (i.e., ‘don't harm other people'), Fairness (i.e., ‘don't pursue your own advantage in disproportionate ways'), Loyalty (i.e., ‘don't betray your in-group'); Authority (i.e., ‘don't disrespect laws, rules, and authority figures'), and Sanctity (i.e., ‘don't do something ‘impure’ or ‘indecent'). Another, separable core moral principle may be Honesty (i.e., ‘don't manipulate the truth'; Hofmann et al., 2014). Each of these principles can serve as a moral reason of desire regulation, and, in fact, self-control goals often appear to be moralized (e.g., Haidt and Hersh, 2001; Rozin and Singh, 1999). An intriguing question in this context is whether self-regulatory goals that are moralized may be more effective than self-regulatory goals that do not have that additional moral ingredient, and some initial evidence seems to support this (Hosey, 2014; Rozin et al., 1997). Furthermore, are some of these moral reasons/dimensions more effective than others?
Clearly, individuals differ in the extent to which they endorse self-regulatory goals (such as health goals), as well as the different dimensions of morality that may support moralization of a given desire-related behavior (Graham et al., 2011). Of further relevance are traits/values relating to the degree to which people may see a need to regulate desires that other people may regard as problematic. Such different ‘thresholds’ for the problematic aspects of desire enactment may, for instance, be related to the endorsement of hedonism-related values (Ghoniem and Hofmann, 2016b), or to a pronounced sense of entitlement to rewards and other things (Hofmann, Baumeister et al., 2012).
Mechanisms of Desire Regulation
Given that desire is sometimes problematic, how can it be effectively self-regulated in accordance with self-regulatory goals and values? We propose that there are roughly three main types of desire-regulation mechanisms (see also Hofmann and Vohs, 2016): (1) those that constrain the emergence of desire, (2) those that help down-regulate desire experience (i.e., desire intensity), and (3) those that enable inhibition or overriding of the action tendency implied by the desire.
Constraining Desire
The first class of desire-regulation strategies encompasses early-stage strategies that may prevent an individual from experiencing an unwelcome desire. This class represents the fact that people can play an active part with regard to the types of situations and stimuli they encounter in their day-to-day lives. If successful, these strategies typically render middle- and late-stage desire regulation unnecessary. It should be noted, however, that these preventive strategies may require considerable foresight, understanding, and experience concerning which situations and stimuli are likely to trigger problematic desire. As people extract lessons from their past behavior (Baumeister et al., 2007), they may become increasingly better at avoiding problematic desire.
Situation selection and stimulus control
The most effective strategy to prevent desire is to avoid exposure to tempting situations or stimuli altogether through strategies of situation selection and stimulus control (Mahoney and Thoresen, 1972). Based on the assumption that external stimuli play a seminal role in the generation of desire (Hofmann and van Dillen, 2012), such strategies can greatly alter the odds that people will experience temptation in the first place (Fujita, 2011; Hofmann and Kotabe, 2012). Situation and stimulus control techniques may not only be applied by the self-regulator directly (e.g., keeping one's refrigerator free of alcohol) but also be imposed through so-called ‘nudges’ (Thaler and Sunstein, 2009), which have become a major area of interest at the intersection of public policy and health. The use of situation and stimulus control can be seen, for example, in ‘no smoking’ policies at restaurants and at school cafeterias that primarily offer healthy options (e.g., Hanks et al., 2012). Because situation and stimulus control are not always feasible, such as when one cannot escape a temptation-rich environment, they cannot be the only means of effective desire regulation.
The Everyday Temptations Study (Hofmann, Baumeister et al., 2012) showed that individuals high in trait self-control (TSC) reported lower average desire strength (as well as lower average conflict and less use of active resistance to control desire). This pattern of findings can be interpreted as high TSC individuals making more use of preventive situation and stimulus control strategies, thus avoiding tempting desires more often than their low TSC counterparts (and hence reducing the need for effortful control). In support, independent raters rated the desires reported by high TSC participants as less problematic for the ‘average person’ than the desires reported by low TSC participants (Hofmann, Baumeister et al., 2012). These findings are intriguing as they suggest that TSC may not so much be about how well temptations are resisted (the type of ‘late-stage’ self-control typically studied in the laboratory situations), but also about how well temptations are being avoided or appraised at early stages in the self-control process.
Early-stage distraction
Another, more proximal, mechanism than situation and stimulus control that appears to constrain the potential for desire experience is early-stage distraction. The underlying idea is that, sometimes, people may be so focused on a given current goal or activity (e.g., reading an engaging novel) that tempting stimuli in their environment do not capture enough attention anymore that would otherwise lead to the conscious representation (and reprocessing) of desire in working memory. Indeed, recent research has shown that cognitively demanding tasks (unrelated to the temptation at hand) can reduce or eliminate the experience of desire (Kemps et al., 2008b), even for people easily tempted by (food) rewards in their environment (van Dillen et al., 2013). Accordingly, more applied work has shown that distraction may therefore have high potential for craving interventions (Florsheim et al., 2008; Skorka-Brown et al., 2014; van Dillen and Andrade, 2016).
However, this strategy may only work to the extent that such powerful distractors can be easily found and are applied before strong cravings can be enacted, as some research suggests possible detrimental effects of distraction at later stages, especially when actual behavioral control is required (Friese et al., 2008; Shiv and Fedorikhin, 1999). However, this does not mean that attentional distraction is no longer effective once a desirable stimulus has been encountered and evaluated as such. For example, even when strong (food) cravings have already been induced, cognitive distractions can still downplay these cravings and subsequent impulsive choices (van Dillen and Andrade, 2016). We argue, however, that with ongoing cognitive elaborations in working memory, desires gain more and more potential to trigger actual behaviors, which should make attentional distraction – and arguably most other strategies aimed at reducing desire strength – less effective with time proceeding.
Individuals also differ considerably in their command of top-down attention. In the cognitive literature on executive functioning, top-down attention has been strongly linked with the concept of working memory capacity (WMC; Kane et al., 2001). WMC represents the ability to maintain and update relevant information in working memory and shield it from interfering processing or distraction (Engle, 2002; Kane et al., 2001). WMC may help people disengage faster from attention-grabbing desire-related cues, thus being less influenced by those cues. Eye-tracking studies, for instance, have shown that people low in WMC are prone to bias eye direction toward tempting cues, with the degree of bias correlating with their automatic affective reactions toward those cues (Friese et al., 2010; Friese and Hofmann, 2012). Those high in WMC, in contrast, were much less influenced by their automatic affective reactions, suggesting that they were faster at disengaging attention from salient cues.
In a related-vein, theorists have pointed out the importance of the involvement of prefrontal cortical regions in self-control (Heatherton and Wagner, 2011). These models have been supported by neuroimaging studies showing that participants instructed to inhibit cravings for drugs (Volkow et al., 2010) or cigarettes (Brody et al., 2007) display increased activity in the prefrontal cortex (PFC), and that the extent of this increase is associated with self-control at the behavioral level and reduced cue-reactivity in regions associated with reward processing such as the ventral striatum. An important question is to what extent these fronto-striatal interactions reflect actual down-regulation, or the absence of up-regulation of NAcc by the dorsolateral PFC (DLPFC). Whereas the DLPFC has been argued to play an important role in self-control processes (Hare et al., 2009; Wagner et al., 2013), studies have also pointed to its involvement in the further elaboration of motivationally relevant cues (Erk et al., 2007; van Dillen et al., 2009; for a review, see Goldstein and Volkow, 2011). Combining transcranial magnetic stimulation and functional magnetic resonance imaging, Hayashi et al. (2013) have demonstrated that the DLPFC modulates craving in response to changes in intertemporal availability. Subjective craving was greater when cigarettes were immediately available, and this effect was eliminated by transiently inactivating the DLPFC with transcranial magnetic stimulation. Inactivation of the DLPFC also reduced craving-related signals in the anterior cingulate and ventral striatum – areas implicated in transforming value signals into action (Walton et al., 2004). These findings indicate that the DLPFC is involved in the build-up of value signals based on contextual knowledge, as much as it down-regulates value signals in the service of overarching goals.
Down-Regulation of Desire Experience
The second class of desire-regulation strategies encompasses those that may down-regulate desire. In this case, desire is experienced and a prepotent action tendency may be activated, but the focus of regulation is on the desire experience rather than on the prepotent action tendency. The idea is that certain regulatory strategies may be more effective than others at reducing the intensity of the experienced desire. Just as a growing fire can be reduced in its power through the right strategy (e.g., repeatedly pouring buckets of water over it), and eventually be extinguished, desire may be reduced below a critical level and eventually fade as the individual employs the right mental strategy at the right moment. This analogy implies that each regulatory strategy may have its advantages and disadvantages, and that aspects of the stimulus, context, and perceiver all determine which strategy is most useful at some point.
Cognitive reappraisal
One way that desire down-regulation can be brought about is through strategies that modify how a tempting stimulus is appraised. Reappraisal is the term used to indicate mental strategies that alter how people perceive an event, idea, or feeling (see also Gross, 1998). Walter Mischel's groundbreaking work on delay of gratification has shown that young children are better able to resist immediate rewards if they learn to reappraise these rewards in non-consummatory ways (Mischel and Baker, 1975). Presumably, such strategies can reduce the immediate appeal of tempting stimuli. Recent research applying this idea to adults has demonstrated that cognitive reappraisal can have a profound impact on affective reactions to tempting stimuli. For instance, having people imagine tempting stimuli in non-consummatory ways appears to reduce people's automatic evaluation of these stimuli (Hofmann et al., 2010). Research using various reappraisal techniques, such as thinking about the negative consequences of enacting desire (Hollmann et al., 2012; Kober et al., 2010), or a combination of reappraisal techniques (Giuliani et al., 2014) provides converging evidence that cognitive reappraisal is generally effective in down-regulating self-reported desires and cravings.
Individuals differ in their use of emotion-regulation strategies such as cognitive reappraisal (Gross and John, 2003), and the effective use of this strategy has been linked to individual differences in WMC (Hofmann, Schmeichel et al., 2012; McRae et al., 2012; Schmeichel et al., 2008). It thus seems straightforward to assume that people high in chronic reappraisal strategy use should have an easier time down-regulating problematic desire than those low in this tendency, even though more direct individual-differences research linking the two appears missing so far.
However, emotion regulation research has examined the conditions under which people choose to employ reappraisal (as compared to distraction). These findings suggest that although reappraisal tends to be more beneficial for long-term adaptation, people choose to reappraise especially in response to mildly intense emotional stimuli. In contrast, people prefer distraction in response to strongly intense emotional stimuli. This preference is likely explained by the finding that compared to reappraisal, distraction requires less cognitive effort (as evidenced by reduced skin conductance and effort-related brain potentials) and more effectively deals with the material's immediate impact, as evidenced by reduced self-reported negative emotions and neural indices of emotion-processing (Shafir et al., 2015; Sheppes et al., 2009; Sheppes and Meiran, 2008). Reappraisal, compared to distraction, however, has been shown to have fewer information-processing costs (e.g., reduced memory of the material; Kron et al., 2010; Sheppes and Meiran, 2008) and allows the person to make sense of the emotional event. Clearly, these different strategies have different implications depending on the situation, and individuals incorporate these in their regulation choices. Whereas no research has yet looked into the choices people make to down-regulate desires of varying intensities, a similar logic might apply.
Suppression
In contrast to the reappraisal literature, the distinct strategy of trying to simply suppress emotion and accompanying thoughts does not seem to fare as well. There is a large body of general emotion regulation research showing that suppression is generally ineffective, if not maladaptive (Gross and Levenson, 1993; Webb et al., 2012). The related literature on appetitive thought suppression comes to the same conclusion (Barnes and Tantleff-Dunn, 2010; Erskine, 2008; Johnston et al., 1999; Mann and Ward, 2001). Thought suppression seems to heighten levels of craving, and binge behavior. Somewhat ironically, thought suppression may backfire (Wegner, 1994). According to Wegner's theory, when trying to actively suppress something, attention may be repeatedly redirected toward the mental content one tries to avoid. This feature of suppression may contribute to keeping the desire-processing loop alive, or even leading to the hyper-accessibility of desire-related thoughts. In one study that addressed the effect of spontaneous instances of suppression on later craving rebounds, nicotine-deprived heavy smoking participants who displayed suppression-related facial expressions during cue exposure (lighting and holding the lit cigarette for thirty seconds) subsequently valued smoking more than those who did not show these expressions (Sayers and Sayette, 2013).
Individuals differ with regard to the propensity to use suppression as a regulatory strategy (Gross and John, 2003). On average, those reverting to this strategy more often than others may be more likely to fail in down-regulating their desires and cravings. For instance, (dysfunctional) perfectionists appear to focus too much on suppression as a regulatory strategy (Bergman et al., 2007). They may thus become overly obsessed with their (problematic) desires. This speculative idea is supported by the finding that people high in dysfunctional perfectionism reported stronger desire intensity in the Everyday Temptations Study (Hofmann, Baumeister, et al., 2012). Nonetheless, it is important to note that some emotion suppressors may be more effective than others because some research suggests that individuals high in WMC seem to be (relatively) better at suppressing unwanted thoughts, as indicated by fewer thought intrusions (Brewin and Smart, 2005; Kane et al., 2007).
Inhibition/Overriding
The third class of desire-regulation strategies encompasses those abilities and strategies that enable the effective inhibition or overriding of the actual desire-related behavior. In this case, a problematic desire is experienced, a prepotent action tendency has already been activated, and the situation allows for the enactment of the behavior. The focus of this strategy is on preventing or limiting the impact of the action tendency on actual behavior (rather than on down-regulating the desire). The basic assumption underlying inhibition/overriding is that either through automatic activation or hijacked executive mechanisms (typically used for self-regulation; see above), desire may activate prepotent action tendencies. Unless inhibited or overridden, these tendencies may be expressed in overt behavior once a certain threshold of activation is reached (Norman and Shallice, 1986; Strack and Deutsch, 2004). Thus, inhibition/overriding requires that the individual manages to keep that prepotent action tendency from influencing behavior for as long as the tempting episode lasts. For successful inhibition/overriding to take place, the individual therefore needs to be both motivated enough to recruit the required executive functions and possess enough capacity to successfully keep the activation of desire-related action tendencies below a critical threshold (Kotabe and Hofmann, 2015). The fact that both motivation and ability need to be present in order for this strategy to be effective means that it is often a difficult endeavor.
A wealth of cognitive experimental research has linked the capacity aspect of behavioral inhibition and response overriding to individual differences in the inhibitory control facet of executive functioning (for a review, see Hofmann, Schmeichel et al., 2012; Miyake et al., 2000). Poor inhibitory control has been implicated in a large number of impulse-control problems including dietary failures (Hofmann et al., 2013; Nederkoorn et al., 2010), drug use and abuse (Berkman et al., 2011; Nigg et al., 2006), inappropriate social responding (von Hippel and Gonsalkorale, 2005), and sexual cheating in romantic relationships (Pronk et al., 2011). Moreover, many studies across these diverse domains have demonstrated that people low in behavioral inhibition are more strongly influenced by prepotent action tendencies than those high in inhibition (e.g., Hofmann et al., 2009a; Houben and Wiers, 2009; Nederkoorn et al., 2010; Payne, 2005), providing more evidence for the underlying mechanism. But inhibitory control skills may also protect individuals who show greater than average reward sensitivity. In one study, for example, higher food reward sensitivity predicted greater palatable food intake at low levels of inhibitory control, but was not associated with intake at high levels of inhibitory control (Appelhans et al., 2011).
From a neuroscientific perspective, recent advances in methodology have enabled researchers to not only look at activity in certain areas of the brain, but, rather, at the degree of coupling or functional connectivity among different brain regions, allowing for a more comprehensive understanding of control dynamics. Of particular interest for desire regulation is the connection between mesolimibic structures, such as the NAcc and prefrontal regions of the brain, assumed to subserve the control of action (Heatherton and Wagner, 2011), and already discussed in relation to desire strength. Interestingly, some first findings suggest that individual differences in impulsivity, as measured via trait impulsivity self-report measures or through behavioral choice tasks that require balancing desire and reason, may be related to the degree of functional connectivity between NAcc and PFC regions (Diekhof and Gruber, 2010). Specifically, individuals high in impulsivity appear to show a stronger coupling between reward-processing centers and PFC regions, and increased responsivity in NAcc to potential rewards, possibly suggesting more bottom-up impact of desire on conscious reward representations and action control. Connectivity between these regions was also inversely related to behavioral success (rejecting the reward). In line with this, those low in impulsivity show a stronger disconnect between ‘wanting’ and ‘reasoned action', as well as reduced NAcc responsivity to reward, which can be interpreted as greater behavioral flexibility, including the increased ability to not act on immediate desire (Diekhof and Gruber, 2010).
Summary and Conclusion
Appetitive desires are common because people spend about half of their waking time desiring certain things (Hofmann, Baumeister et al., 2012). They are deeply connected to the needs that have secured the species’ survival over millennia. Yet, with desire comes the potential for mental conflict and the need to regulate in accordance with individual, social, and societal enactment constraints. Whatever the source of such inner conflict, humans are, by and large, remarkably effective desire regulators (Hofmann, Baumeister, et al., 2012). At the same time, there is room for improvement – as a quick look into contemporary societal problems such as overeating, over-drinking, smoking, or sexually transmitted diseases confirms.
We have argued that effective desire regulation, just as emotion regulation, can take many forms and unfold at various time points. Desire-regulation strategies can roughly be classified into early-stage strategies that proactively constrain desire experiences; middle-stage strategies that support the effective down-regulation of consciously-experienced desire; and late-stage strategies that involve the inhibition, overriding, or moderation of desire-related behavior. These distinctions are difficult to demarcate exactly, however, and what is the right strategy depends on the moment, the situation, and the individual.
We have also argued that individuals differ as to how intensely they experience desire and how they go about regulating it. To recap the most central insights: individuals with particularly sensitive reward centers appear to experience stronger desires and individuals high in TSC seem to make more use of early preventive strategies such as situation and stimulus control. Being dispositionally high in WMC may make it a lot easier to resist the attentional pull of tempting stimuli and to reappraise tempting situations. Being high in inhibitory control capacities aids people in keeping impulsive action tendencies in check. Taking a closer look at what strategies work for what types of people should help practitioners to devise more effective customized treatments for those who suffer from tempting desires and cravings. As technological possibilities mature, we expect to see fascinating new developments in the years to come with regard to one of the central challenges of what it means to be human: how to shape one's internal landscape of appetitive desires in a way that strikes a healthy balance between desire enactment and the occasional need to curb one's passion for what might be a pretty good reason.
As psychological and neurocognitive insights into desire and desire regulation mature, new treatment methods and technologies will surely emerge. Applied research is likely to profit from these insights and will make progress in helping those individuals deal with problematic desires who, according to our analysis, are in the strongest need for external support and decision aids: those with above-average desire experiences and those with particularly poor emotion regulation and behavioral inhibition abilities.
Note
1. However, additional (negative) evaluations may be added on top of this base affective component, leading to the well-known ambivalence of desire experiences (Kavanagh et al., 2005; van Harreveld et al., 2016). Furthermore, although enacting or thwarting desires may have certain self-conscious emotional consequences such as guilt, we view such emotional consequences as separable from the distinct, intrinsic phenomenology of desire, which is grounded in the anticipation of rewarding experiences.
References
, , , , , & (2011). Inhibiting food reward: delay discounting, food reward sensitivity, and palatable food intake in overweight and obese women. Obesity, 19, 2175–2182.
, & Tantleff-Dunn, S. (2010). Food for thought: examining the relationship between food thought suppression and weight-related outcomes. Eating Behaviors, 11, 175–179.
, , , & (2007). How emotion shapes behavior: feedback, anticipation, and reflection, rather than direct causation. Personality and Social Psychology Review, 11, 167–203.
, , & (2007). Correlates with perfectionism and the utility of a dual process model. Personality and Individual Differences, 43, 389–399.
, , & (2011). In the trenches of real-world self-control: neural correlates of breaking the link between craving and smoking. Psychological Science, 22, 498–506.
, , & (2009). Dissecting components of reward: ‘liking', ‘wanting', and learning. Current Opinion in Pharmacology, 9, 65–73.
, & Oscar-Berman, M. (2005). Relationship between dopaminergic neurotransmission, alcoholism, and reward deficiency syndrome. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 132B, 29–37.
, & (2005). Working memory capacity and suppression of intrusive thoughts. Journal of Behavior Therapy and Experimental Psychiatry, 36, 61–68.
, , , , , , … (2007). Neural substrates of resisting craving during cigarette cue exposure. Biological Psychiatry, 62, 642–651.
, , & De (2014). ‘Because I am worth it': a theoretical framework and empirical review of a justification-based account of self-regulation failure. Personality and Social Psychology Review, 18, 119–138.
, , & De (2012). License to sin: self-licensing as a mechanism underlying hedonic consumption. European Journal of Social Psychology, 42, 490–496.
, , & (2012). Individual differences in nucleus accumbens activity to food and sexual images predict weight gain and sexual behavior. Journal of Neuroscience, 32, 5549–5552.
, & (2010). When desire collides with reason: functional interactions between anteroventral prefrontal cortex and nucleus accumbens underlie the human ability to resist impulsive desires. Journal of Neuroscience, 30, 1488–1493.
(2002). Working memory capacity as executive attention. Current Directions in Psychological Science, 11, 19–23.
, , & (2007). Valence-specific regulation effects in a working memory task with emotional context. Neuroimage, 37, 623–632.
(2008). Resistance can be futile: investigating behavioural rebound. Appetite, 50, 415–421.
, , , , & (2008). An experimental test of a craving management technique for adolescents in substance-abuse treatment. Journal of Youth and Adolescence, 37, 1205–1215.
(2002). Behavioral approach system (BAS) sensitivity predicts alcohol craving. Personality and Individual Differences, 32, 349–355.
, & (2012). Just a little bit longer: viewing time of erotic material from a self-control perspective. Applied Cognitive Psychology, 26, 489–496.
, , , & (2010). Here's looking at you, Bud: alcohol-related memory structures predict eye movements for social drinkers with low executive control. Social Psychological and Personality Science, 1, 143–151.
, , & Wänke, M. (2008). When impulses take over: moderated predictive validity of implicit and explicit attitude measures in predicting food choice and consumption behaviour. British Journal of Social Psychology, 47, 397–419.
(2011). On conceptualizing self-control as more than the effortful inhibition of impulses. Personality and Social Psychology Review, 15, 352–366.
, & (2016a). Trio infernale and the emergence of desires: how stimulus properties, need states and learning history interact in elicting desire. Unpublished manuscript.
, & (2016b). When impulsive behaviors don't equal self-control failures: the role of valuation of temptation enactment. Unpublished mansucript.
, , , & (2014). Neural systems underlying the reappraisal of personally craved foods. Journal of Cognitive Neuroscience, 26, 1390–1402.
, & (2011). Dysfunction of the prefrontal cortex in addiction: neuroimaging findings and clinical implications. Nature Reviews Neuroscience, 12, 652–669.
, , , , , , … (2007). Decreased prefrontal cortical sensitivity to monetary reward is associated with impaired motivation and self-control in cocaine addiction. The American Journal of Psychiatry, 164, 43–51.
, , , , , , & (2013). Moral foundations theory: the pragmatic validity of moral pluralism. Advances in Experimental Social Psychology, 47, 55–130.
, , , , , & (2011). Mapping the moral domain. Journal of Personality and Social Psychology, 101, 366–385.
(Ed.). (1982). The Neuropsychology of Anxiety: An Enquiry into the Functions of the Septo-Hippocampal System. New York, NY: Oxford University Press.
(1998). The emerging field of emotion regulation: an integrative review. Review of General Psychology, 2, 271–299.
, & (2003). Individual differences in two emotion regulation processes: implications for affect, relationships, and well-being. Journal of Personality and Social Psychology, 85, 348.
, & (1993). Emotional suppression: physiology, self-report, and expressive behavior. Journal of Personality and Social Psychology, 64, 970–986.
, & (2001). Sexual morality: the cultures and emotions of conservatives and liberals. Journal of Applied Social Psychology, 31, 191.
, , , & (2012). Healthy convenience: nudging students toward healthier choices in the lunchroom. Journal of Public Health, 34, 370–376.
, , & (2009). Self-control in decision-making involves modulation of the vmPFC valuation system. Science, 324, 646–648.
, , , & (2013). Dorsolateral prefrontal and orbitofrontal cortex interactions during self-control of cigarette craving. Proceedings of the National Academy of Sciences, 110, 4422–4427.
, & (2011). Cognitive neuroscience of self-regulation failure. Trends in Cognitive Sciences, 15, 132–139.
(1998). The aboutness principle: a pervasive influence on human inference. Social Cognition, 16, 173–198.
, & (2012). A general model of preventive and interventive self-control. Social and Personality Psychology Compass, 6, 707–722.
, & (2013). Desire and desire regulation: basic processes and individual differences. In J. J. Gross (Ed.), Handbook of Emotion Regulation (2nd edn, pp. 346–360). New York, NY: Guilford Press.
, & (2015). Introduction to the psychology of desire. In W. Hofmann & L. F. Nordgren (Eds), The Psychology of Desire (pp. 1–13). New York, NY: Guilford Press.
, & van (2012). Desire: the new hotspot in self-control research. Current Directions in Psychological Science, 21, 317–322.
, & D. (2016). Desire and Self-Regulation. In K. D. Vohs & R. F. Baumeister (Eds), The Handbook of Self-Regulation: Research, Theory, and Applications (Vol. 3, pp. 204–226). New York: Guilford Press.
, , , & (2013). Dieting and the self-control of eating in everyday environments: an experience sampling study. British Journal of Health Psychology, online advance publication.
, , Förster, G., & (2012). Everyday temptations: an experience sampling study of desire, conflict, and self-control. Journal of Personality and Social Psychology, 102, 1318–1335.
, , , & (2010). Cooling the heat of temptation: mental self-control and the automatic evaluation of tempting stimuli. European Journal of Social Psychology, 40, 17–25.
, , & (2009a). Three ways to resist temptation: the independent contributions of executive attention, inhibitory control, and affect regulation to the impulse control of eating behavior. Journal of Experimental Social Psychology, 45, 431–435.
, , & (2009b). Impulse and self-control from a dual-systems perspective. Perspectives on Psychological Science, 4, 162–176.
, , , & (2011). Working memory and self-regulation. In K. D. Vohs & R. F. Baumeister (Eds), The Handbook of Self-Regulation: Research, Theory, and Applications (Vol. 2, pp. 204–226). New York, NY: Guilford Press.
, , & (2012). Executive functions and self-regulation. Trends in Cognitive Sciences, 3, 174–180.
, , , & (2014). Morality in everyday life. Science, 345, 1340–1343.
, , , , , , … (2012). Neural correlates of the volitional regulation of the desire for food. International Journal of Obesity (London), 36, 648–655.
(2014). Moralizing Goals: Consequences for Goal Pursuit and Judgment. Chicago, IL: University of Chicago.
, & (2009). Response inhibition moderates the relationship between implicit associations and drinking behavior. Alcoholism: Clinical and Experimental Research, 33, 1–8.
, , & (1999). Suppressing thoughts about chocolate. International Journal of Eating Disorders, 26, 21–27.
, , , & (2001). A controlled-attention view of working-memory capacity. Journal of Experimental-Psychology: General, 130, 169–183.
, , , , , & (2007). For whom the mind wanders, and when: an experience-sampling study of working memory and executive control in daily life. Psychological Science, 18, 614–621.
, , & (2005). Imaginary relish and exquisite torture: the elaborated intrusion theory of desire. Psychological Review, 112, 446–467.
, , & (2005). Neural systems recruited by drug- and food-related cues: studies of gene activation in corticolimbic regions. Physiology and Behavior, 86, 11–14.
, , & (2008b). Concurrent visuo-spatial processing reduces food cravings in prescribed weight-loss dieters. Journal of Behavior Therapy and Experimental Psychiatry, 39, 177–186.
, , & (2008a). Food cravings consume limited cognitive resources. Journal of Experimental Psychology: Applied, 14, 247–254.
, , , , , , & (2010). Prefrontal-striatal pathway underlies cognitive regulation of craving. Proceedings of the National Academy of Sciences of the United States of America, 107, 14811–14816.
, & (2015). On integrating the components of self-control. Perspectives on Psychological Science, 10, 618–638.
, , , & (2010). Feelings don't come easy: studies on the effortful nature of feelings. Journal of Experimental Psychology: General, 139, 520–534.
, , , , & (2014). Neural predictors of giving in to temptation in daily life. Psychological Science, 25, 1337–1344.
, & (1972). Behavioral self-control: power to the person. Educational Researcher, 1, 5–7.
, & (2001). Forbidden fruit: does thinking about a prohibited food lead to its consumption? International Journal of Eating Disorders, 29, 319–327.
, , , Hwang, D.-R., , , … Abi-Dargham, A. (2005). Alcohol dependence is associated with blunted dopamine transmission in the ventral striatum. Biological Psychiatry, 58, 779–786.
, , , , & (2012). Individual differences in reappraisal ability: links to reappraisal frequency, well-being, and cognitive control. Journal of Research in Personality, 46, 2–7.
(2001). Autonomous Agents. From Self-Control to Autonomy. Oxford: Oxford University Press.
, & (1975). Cognitive appraisals and transformations in delay behavior. Journal of Personality and Social Psychology, 31, 254–261.
, , , , & (2000). The unity and diversity of executive functions and their contributions to complex ‘frontal lobe’ tasks: a latent variable analysis. Cognitive Psychology, 41, 49–100.
, , & (1980). From motivation to action – functional interface between the limbic system and the motor system. Progress in Neurobiology, 14, 69–97.
, , , , & (2010). Control yourself or just eat what you like? Weight gain over a year is predicted by an interactive effect of response inhibition and a preference for high fat foods. Health Psychology, 29, 389–393.
, , , , , , … (2006). Poor response inhibition as a predictor of problem drinking and illicit drug use in adolescents at risk for alcoholism and other substance use disorders. Journal of the American Academy of Child and Adolescent Psychiatry, 45, 468–475.
, & (1986). Attention to action. Willed and automatic control of behavior. In R. J. Davidson, G. E. Schwartz & D. Shapiro (Eds), Consciousness and Self Regulation: Advances in Research (pp. 1–18). New York, NY: Plenum Press.
(2005). Conceptualizing control in social cognition: how executive control modulates the expression of automatic stereotyping. Journal of Personality and Social Psychology, 89, 488–503.
, & (2005). Hedonic hot spot in nucleus accumbens shell: where do μ-opioids cause increased hedonic impact of sweetness? The Journal of Neuroscience, 25, 11777–11786.
, , & (2011). How can you resist? Executive control helps romantically involved individuals to stay faithful. Journal of Personality and Social Psychology, 100, 827–837.
, & (1999). The moralization of cigarette smoking in the United States. Journal of Consumer Psychology, 8, 321–337.
, , & (1997). Moralization and becoming a vegetarian: the transformation of preferences into values and the recruitment of disgust. Psychological Science, 8, 67–73.
, & (2013). Suppression on your own terms: internally generated displays of craving suppression predict rebound effects. Psychological Science, 24, 1740–1746.
, , & (2008). Working memory capacity and the self-regulation of emotional expression and experience. Journal of Personality and Social Psychology, 95, 1526–1540.
(2007). We can do better – improving the health of the American people. New England Journal of Medicine, 357, 1221–1228.
, , , & (2015). Emotional intensity influences pre-implementation and implementation of distraction and reappraisal. Social Cognitive and Affective Neuroscience, 10, 1329–1337.
, & (2008). Divergent cognitive costs for online forms of reappraisal and distraction. Emotion, 8, 870–874.
, , & (2009). Reappraisal (but not distraction) is going to make you sweat: physiological evidence for self-control effort. International Journal of Psychophysiology, 71, 91–96.
, & (1999). Heart and mind in conflict: the interplay of affect and cognition in consumer decision making. Journal of Consumer Research, 26, 278–292.
, , & (2014). Playing ‘Tetris’ reduces the strength, frequency and vividness of naturally occurring cravings. Appetite, 76, 161–165.
, , , & (2009). Relation of obesity to consummatory and anticipatory food reward. Physiology and Behavior, 97, 551–560.
, & (2004). Reflective and impulsive determinants of social behavior. Personality and Social Psychology Review, 8, 220–247.
, & (2009). Nudge: Improving Decisions about Health, Wealth, and Happiness. New York, NY: Penguin Books.
, & (2016). Derailing the streetcar named desire. Cognitive distractions reduce individual differences in cravings and unhealthy snacking in response to palatable food. Appetite, 96, 102–110.
, & van (2018). Tuning down the hedonic brain. Cognitive load reduces neural responses to high-calorie food pictures in the nucleus accumbens. Cognitive, Affective, and Behavioral Neuroscience Available online at https://link.springer.com/article/10.3758/s13415-018-0579-3.
, , & (2009). Tuning down the emotional brain: an fMRI study of the effects of cognitive load on the processing of affective images. Neuroimage, 45, 1212–1219.
, , & (2013). Turning a blind eye to temptation. How task load can facilitate self-regulation. Journal of Personality and Social Psychology, 3, 427–443.
, , & (2016). You shan't always get what you want: the consequences of ambivalence towards desires. In W. Hofmann & L. F. Nordgren (Eds), The psychology of desire (pp. 267–285). New York, NY: Guilford Press.
, , , & (2015). Neural processing of calories in brain reward areas can be modulated by reward sensitivity. Frontiers in Behavioral Neuroscience, 9, 371.
, , Wang, G.-J., , , , … (2010). Cognitive control of drug craving inhibits brain reward regions in cocaine abusers. Neuroimage, 49, 2536.
, , , , , , … (2002). ‘Nonhedonic’ food motivation in humans involves dopamine in the dorsal striatum and methylphenidate amplifies this effect. Synapse, 44, 175–180.
, , , , , von der , … (2012). Validating incentive salience with functional magnetic resonance imaging: association between mesolimbic cue reactivity and attentional bias in alcohol-dependent patients. Addiction Biology, 17, 807–816.
, & (2005). ‘That is bloody revolting!’ Inhibitory control of thoughts better left unsaid. Psychological Science, 16, 497–500.
, , , , & (2013). Self-regulatory depletion enhances neural responses to rewards and impairs top-down control. Psychological Science, 24, 2262–2271.
, , & (2004). Interactions between decision making and performance monitoring within prefrontal cortex. Nature Neuroscience, 7, 1259–1265.
, , , & (2004). Similarity between obesity and drug addiction as assessed by neurofunctional imaging. Journal of Addictive Diseases, 23, 39–53.
, & (1999). Effect of food deprivation and maintenance diet composition on fat preference and acceptance in rats. Physiology and Behavior, 68, 235–239.
, , & (2012). Dealing with feeling: a meta-analysis of the effectiveness of strategies derived from the process model of emotion regulation. Psychological Bulletin, 138, 775.
(1994). Ironic processes of mental control. Psychological Review, 101, 34–52.
, , & (2005). Unconscious affective reactions to masked happy versus angry faces influence consumption behavior and judgments of value. Personality and Social Psychology Bulletin, 31, 121–135.