6
Strategies for Activating the Brain
Fundamental to fMRI is the collection of a time series of whole-brain snapshots, spaced on the order of one second apart. The hemodynamic response to brain activation, while sluggish, is extremely well behaved and consistent. This consistency over time allows for a wide array of possibilities in designing a brain activation experiment.
The goal of designing activation paradigms is to localize and extract behaviorally relevant brain activation as efficiently and cleanly as possible. Paradigm design has been an area of abundant innovation. Most fMRI neuroscientists become adept at designing paradigms and tailoring them to the limitations and advantages of specific acquisition and processing strategies. This chapter highlights the types of paradigms that have been most novel and promising. These include blocked design, event-related fMRI, phase-encoding, fMRI adaptation, resting state fMRI, naturalistic stimulus presentations, and real-time fMRI feedback. A background reference for this section is a paper by Amaro and Barker that summarizes study design and analysis in fMRI.1 Figure 14 lists and shows iconic representations of seven classes of brain activation strategies that can be used.
Fundamental to fMRI is the collection of a time series of whole-brain snapshots, one second apart. The hemodynamic response to brain activation, while sluggish, is extremely well behaved and consistent. This consistency allows for a wide array of possibilities in designing a brain activation experiment.
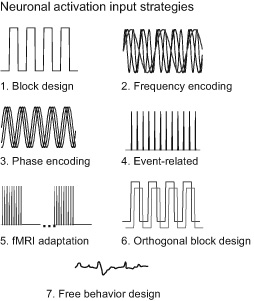
Figure 14 An iconic depiction of key brain activation strategies used in fMRI. The most esoteric and least used is frequency encoding, which applies the concept of “multiplexing” multiple activation on-off frequencies. The event-related brain activation strategy, depicted here using constant intervals, now primarily employs jittered intervals to allow for deconvolution. Free behavior design appears to be gaining the most popularity.
Blocked Design
Functional MRI began with the use of blocked designs where subjects alternated approximately 10–30 sec of performing a task or receiving stimuli with an equal or slightly longer period of rest. A longer period of rest allows for the post-stimulus undershoot to return to baseline. A complete return to baseline can take up to 40 sec. Multiple tasks can be inserted as blocks. Images collected during each condition can be averaged and compared. The on-off switching in the signal allows separation of brain activation-related changes from the slow drifts that are ubiquitous in fMRI time series due to scanner instabilities or slow subject motion. Therefore, in order to optimally minimize the effects of low-frequency drifts, it is generally desirable to turn the tasks on and off as rapidly as possible and as many times as possible, pushing the on-off frequency as high as possible—separating it from low-frequency artifactual drifts—without compromising the amplitude of the response. The highest on-off frequency at which this can be obtained is about 10 sec on and 10 sec off or 0.05 Hz. The highest on-off frequency that is able to induce any measurable fMRI signal change is 0.67 sec on and 0.67 sec off or 0.75 Hz.
One useful advancement in blocked design paradigms was demonstrated by Courtney et al.,2 in which six different time series related to working memory were efficiently incorporated into a single time series. The key aspect of this design was that the brain activation timing was such that the time series corresponding to each unique aspect of brain activity were all orthogonal to each other. By definition, all of the time series signals that are orthogonal to each other have a correlation of zero. These six regressors produced six different activation maps, all mathematically independent of each other. If paradigms are designed so that there is in fact some correlation between time series, time series can be mathematically “orthogonalized.” However, this results in a loss of statistical power.
Interestingly, the 10 sec on and 10 sec off timing produces an activation time series that is nearly completely orthogonal to stimulus-induced motion. Such timing can be useful when having the subject perform tasks in the scanner that involve brief motion, such speaking words.3
It should be noted that much of the brain is behaving in what is likely a more temporally rich manner than what is typically modelled in a blocked design paradigm. Time-locked signal changes have been shown to vary considerably relative to a well-behaved on-off regressor. These signals are time locked, so they represent neuronal activity associated with the task, but are often overlooked by a choice of only a few model functions. Care should be taken when investigating the data to determine if any other repeatable, non-canonic signal changes have been missed.
While blocked designs are statistically the most powerful of paradigms, many brain activation tasks do not lend themselves to a constant duration of activity for tens of seconds. Event-related fMRI helps address these issues.
Event-Related fMRI
The idea of presenting and averaging multiple brief stimuli drew upon the vast EEG and MEG literature. In 1992, Blamire et al. first demonstrated event-related fMRI with a 3 sec visual stimulation.4 Since they did not coin a term in that paper to describe this new paradigm, it is often overlooked. In 1996, Buckner et al. and McCarthy et al. first demonstrated event-related fMRI for cognitive tasks.5 Other researchers have since demonstrated that there is practically no limit to how brief a brain activation duration is detectable with fMRI. The limitation was simply in how brief a brain response could be elicited. Stimuli as brief as 16 ms have elicited robust responses.
In the early days of event-related fMRI paradigm implementation, a brief task was given, and then there was a waiting period typically longer than 15 sec to allow the hemodynamic response to return to baseline.6 This approach not only was statistically inefficient, it also was extremely boring for the subject and often induced undesirable “anticipation”-related activation as the subject awaited the next stimulus. During the mid- to late 1990s, an innovation—drawing on mathematics of linear systems—occurred in event-related fMRI. Researchers realized that the hemodynamic response could be modeled as a linear system. Neuronal inputs that are spaced such that their subsequent hemodynamic responses overlapped can be separated using simple deconvolution as long as the interstimulus intervals were varied or “jittered.” Rapid event-related fMRI can typically accommodate an average stimulus interval of about 4 sec and stimulus duration of 0.5–3 sec, allowing a wide range and number of stimuli to be presented in a single time series. Studies followed that characterized the optimal average and distribution of interstimulus intervals, suggesting that, with event-related designs, the necessary timing for optimal creation of activation maps is very different than for characterizing the shape of the hemodynamic response from already known areas.7
Both blocked designs and event-related designs can be parametrically modulated, meaning that the task or stimulus intensity can be changed in a systematic manner such that it is reflected in the amplitude of the fMRI response. Varying an aspect of the task and then comparing the relative change in fMRI amplitude allows the precise determination of how much each region contributes to the corresponding processing. Some regions may not show a signal intensity modulation with task modulation but instead show a constant amplitude at all task intensities, while other regions may show a linear relationship. Others might show more complicated relationships. In general, parametric designs are common in fMRI as they powerfully differentiate the functional relationship between specific regions and task or stimulus quality or intensity.
Phase Encoding
Vision neuroscientists were among the first to embrace and advance the paradigm design methodology in fMRI. In 1994, Engel et al. were the first to demonstrate a continuous activation design that “phase-encoded” visual stimuli.8 were the first to demonstrate a continuous activation design that “phase-encoded” visual stimuli. Here, the term “phase-encoding” means that the temporal “phase” corresponds to a specific visual-field location of the stimulus as well as a corresponding location in the brain in the case of retinotopy. The stimulus is never alternated with an “off” period, but rather, some aspect is varied continuously. In their study, a stimulus consisted of a flashing checkerboard ring that slowly increased in radius until it extended out to the periphery and then, once reaching the periphery, the ring repeated again, starting at the fovea. This cycle repeatedly activated a continuously varying ring of cortex. The second stimulus was a rotating wedge about 20 degrees in width and continuously rotated. The cycle time or the time for both approaches was about one minute, allowing for sufficient time for the hemodynamic response to keep up. A typical time series would be about five to ten minutes in duration, having five to ten phase-encoding cycles. Interestingly, this paradigm was able to delineate separate low-level visual areas as it is known that they are organized in a mirror-image fashion, such that the direction of activation associated with the moving wedge would cause opposite direction activation for each area; the borders of these areas can be determined if one pays attention to where the direction of activation changes.
In general, parametric designs are common in fMRI as they powerfully differentiate the functional relationship between specific regions and task or stimulus quality or intensity.
Other applications of this type of paradigm have included: in the visual cortex, mapping spatial frequency selectivity; in the somatosensory cortex, mapping somatotopy; and in the auditory cortex, mapping tonotopy. In animal models, exquisite maps of cortical and subcortical auditory-frequency selectivity have been mapped using a paradigm in which the carrier frequency of a sound was varied slowly and continuously.
fMRI Adaptation
Grill-Spector and Malach originated “fMRI adaptation” paradigms where the amount of signal adaption with rapid sequentially presented stimuli is modulated by the degree to which neurons in each voxel consider the successive stimuli to be similar—either in spatial frequency or in semantic content.9 Here, a stimulus or task is briefly given and then, at short intervals, either a similar or different stimulus or task is presented in a repetitive manner. If the same or similar stimulus is presented repetitively, then the underlying neuronal activity will quickly habituate and result in a blunted fMRI response. If different stimuli are presented, then a different pool of fresh neurons will respond and therefore the overall signal will not “adapt.” This signal will have no subsequent adaptation with repeated stimuli. The assumption is that within each voxel there are different pools of neurons that respond differently to specific stimuli. These pools can be separated by their habituation to successive stimuli. This starts getting interesting when the stimuli are different yet share similar features. If neurons are sensitive to specific features of the stimuli only, then they will adapt when those features keep showing up even if the stimuli are all quite different.
Naturalistic Stimuli
A paradigm design trend that has been picking up momentum in the past decade presents naturalistic stimuli consisting of either movies, audio oration, or free self-paced multi-option behavior in the scanner. This approach then determines the precise correspondence of the time course of the fMRI signal change magnitude or connectivity with specific aspects of these stimuli or tasks as they are closely tracked and deconstructed.
Several analysis approaches have been used. The first is to create regressors corresponding to a specific aspect of the stimuli. In the case of watching a movie, separate regressors may show responses only when faces appear, or when people are talking. The regressors that are used could also be more specific, such as when subjects say a certain class of words, or gaze at the camera. After convolution with the hemodynamic transfer function, the calculations of the correlations between the regressors and the time series are carried out. Regions that show high correlation are inferred to be those activated by the specific aspect of the stimuli. The idea of creating regressors for as much from the stimuli as possible and then mapping the weightings of each of these onto the brain was coined “encoding” by Gallant who has implemented this approach in the context of visual, auditory, and semantic processing. The innovative processing approach that Gallant lab uses will be discussed more in the data processing section. The concept of “encoding” is related to “decoding” in that, for encoding, complex stimuli are broken down and the processing of the elemental parts are mapped onto the brain. For decoding, brain activation maps created by encoding are used as “training sets” that then can be used as templates to decode the pattern of brain activity with entirely novel stimuli—inferring what the subject was seeing, hearing, or doing based on the similarity of the pattern of activation.10
In the mid 2000s, the University of Pittsburgh, through the Organization for Human Brain Mapping (OHBM) and sponsored by Defense Advanced Research Projects Agency (DARPA), put on a competition in which entrants were challenged to deduce, based on brain-activation time courses, specifically what a subject was viewing, using training data sets in which the brain activation corresponding to a known movie stimulus was known. The winner was able to “decode” a movie, based on a previous training set of known movies, with extremely high accuracy from the brain activity alone.11
Hasson et al. advanced a different approach to analyzing naturalistic stimuli paradigms.12 Their approach uses rich and varied yet identical stimuli and/or task timings across separate runs in a single subject or even across subjects such that the time course from one run can be used as the reference function for another run—assuming that the stimuli for all subjects are precisely time-locked with each other. This approach addressees a central problem prevalent in fMRI: “catching” all the interesting signals that arise in the data that may not resemble a canonical regressor shape. Any fluctuations in the signal that occur for the separate runs will be highly correlated, and therefore likely resulting from what was presented rather than from random fluctuations. Taken further, any differences in subjects’ responses with identical time-series stimuli would imply a difference in how they process information.
Performing cross-subject correlation also helps eliminate spurious, non-task related temporal variations in the stimuli given that no two subjects are likely to have the same artifactual signal timings. Such a method has promise for effectively identifying salient differences and similarities across large numbers of subjects, and thus may be used to identify clinically relevant “biomarkers” corresponding to pathological conditions.
Resting State fMRI
Resting state fMRI is one of the most significant innovations since the inception of fMRI and is based on this observation: when the brain is not performing any overt task or receiving any stimulus, it is spontaneously active such that regions that are functionally related show correlation in their low-frequency (about 0.1 Hz) fluctuations—presumably driven by temporally correlated synchronous and spontaneous activity. The term “resting state fMRI” is a misnomer given the assumption that during “rest” the brain is not at all quiescent but rather shows constant churning activity that results in constant generation of fMRI signals. The temporal correlation of these signals from separate regions in the brain implies that they are “connected.” Another name for resting state fMRI is functional connectivity fMRI. This name is problematic because many other variables than spontaneous activation can influence the degree of correlation in these time series signals.
A specific set of tools helped lay the groundwork for the discovery of the resting state fMRI innovation. In the software program AFNI, as well as its predecessors, users are able to choose a time course signal from the data—either from a voxel or from the average of multiple voxels—and then immediately use it as a reference function for comparison with all the individual voxel time courses in the brain. This tool set was extremely useful for exploring activation data in a rapid preliminary manner.
It was only a matter of time before someone would apply this tool to resting data where there was no obvious task. It took the curiosity of Bharat Biswal to use this tool in this manner.13 His initial observation that the left and right motor cortex showed a high level of correlation even when the subject was not performing any overt task did not catch on immediately. His manuscript was published in 1995, yet the number of studies performing resting state fMRI were a mere trickle—accumulating no more than about five a year—until about 2005, when the field decided that resting state fMRI was real and useful for more than just mapping connectivity in the primary motor, visual, and auditory cortex. The rapid advance after this incubation period may also have been due to the fact that acquisition and processing methods had reached a state of sophistication such that detection of these resting state correlations was easy enough for a larger fraction of users to undertake. After about a decade researchers started to consider resting state studies that were complementary to their own activation studies, and then the floodgates opened as processing methods became more sophisticated. More interesting and encouraging results came out as more resting state networks were discovered. The number of networks or correlated nodes is not known, having increased in number—due to better processing methods and more sensitivity—from 5 or so upward to 350.14 It’s likely that the number of well-characterized nodes will continue to grow—perhaps until columnar organization is reached—as our ability to delineate fine detail increases.
Methods for de-noising, clustering, displaying, and comparing resting state data advanced after 2006. The field of resting state fMRI has helped spawn new journals such as Brain Connectivity and has become perhaps the dominant topic of study at the annual Organization for Human Brain Mapping Meeting. A large amount of federal funding has been redirected into understanding the human “Connectome” with resting state fMRI a major paradigm in this research.
Early on, and to some degree, still, it was thought that this signal was either an artifact of imaging, vigilance, or “vasomotion”—implying that the signal, while related to an underlying physiologic process, was not directly related to functional connectivity and therefore could not be reliably used as a measure of internal brain connectivity. The technique met skepticism as the fundamental observation of time series correlation between disparate and spatially independent signals in the brain had not been made in the context of other imaging modalities. It was almost too good to be true; however, as we are finding, it is true and works well.
Functional connectivity appears to be constantly changing over time during time series scans, suggesting a dynamic reconfiguration of networks or “brain state” changes. The ability to observe dynamic brain state changes has been brought about by a method by which the correlations of every region with every other region over a window of perhaps minute or so in duration is carried out for each time point that is then “slid” along in time. This is known as the sliding window approach. In the presence of networks changing over time, it is a challenge even to really know what a network is. Some nodes jump from being correlated with one network to being correlated with another network. Which network does it belong to? A subfield of fMRI that focuses on dynamic connectivity changes has emerged, as it has been suggested that these dynamic states carry information about populations or individuals.
The field of resting state fMRI has helped spawn new journals such as Brain Connectivity and has become perhaps the dominant topic of study at the annual Organization for Human Brain Mapping Meeting. Federal funding has been redirected into understanding the human “Connectome” with resting state fMRI a major paradigm in this research.
A central problem in all fMRI is how to cleanly separate meaningful fMRI signal from noise and artifact. This problem is much more difficult when studying resting state fMRI as there is no ground truth as to when the brain is active or not. Simultaneous external measures of neuronal activity, breathing, and cardiac function have can be used to help sort the fMRI signal but they are far from optimal. Currently, about half of the overall signal power that remains in the fMRI time series is not related to neuronal activity.15
Many open questions exist regarding resting state fMRI. Among the most pressing are the following:
- 1. What is the “purpose” of resting state fluctuations? What evolutionary need do they satisfy? What is the purpose of so much low frequency energy?
- 2. What temporal characteristics define each resting state network? Can they be defined are characterized in this way at all?
- 3. What about latencies? Accounting for hemodynamic latency variations across the brain may increase detectability and interpretability of resting state fMRI data. Do latencies contain more biologic meaning than simply downstream “draining vein” effects?
- 4. How many brain “states” exist? What are the time constants or sequence of these states? Do these measured states correspond to actual states of mind? Are they sensitive to pathology?
- 5. How finely delineated are these networks?
- 6. How sensitive is the resting state signal to pathology and prediction of treatment?
- 7. How much of the resting state signal is hemodynamic (i.e., vasomotion) versus neuronal in origin?
- 8. To what predominant neuronal oscillation frequency does the resting state fMRI signal correspond?
- 9. How is connectivity truly represented through correlation? Also, changes in connectivity typically are inferred by changes in correlation. A change in correlation—in the presence of noise—could also be brought on by a simple change in amplitude of one signal or the other, or a simple change in SNR of one signal or the other.
Tenth, and last, a controversy that has been debated over the last few years concerns whether to regress out the global signal or not in order to increase sensitivity.16 The growing consensus is that we should not remove the global signal as it artificially induces negative correlations between previously uncorrelated regions—thus complicating interpretation. However, a recent study has shown that this global signal is highly sensitive to the state of vigilance and removing it may normalize for vigilance variations across subjects and over time.17 The question remains, what should we do about the global signal? It appears to depend on what we want to do with the time series. If we want to reduce vigilance-related variability across subjects, we might want to regress it out—but with caution. If we want to characterize differences in vigilance or vigilance-related processing across subjects, we might want to keep it in.
The growth of resting state processing methods is still accelerating. There will not be a single best way to analyze resting state data, but rather an array of approaches that give the researcher a tool set to address a particular question.
Real Time fMRI
Real time fMRI is the ability not only to collect and display the echo planar images during time series collection but also to continuously perform updated functional MRI analysis on the time series and to display this functional data to the investigator, or in some cases, back to the subject—allowing for image-based biofeedback. Real time fMRI was first developed in the mid-1990s by Cox, Jesmanowicz, and Hyde.18 At this time, it pushed the limits of the available computation speed. Since then, with the advancement of computation speed and memory, having the ability to reconstruct and perform data analysis in real time is more commonplace. At many centers, real time fMRI has served for fMRI demonstration purposes. Showing brain activation as it happens in individual subjects during a scan can convey a powerful message to nonexperts that fMRI is indeed real. It gives a profound sense of the method’s capacity to image activation as it is occurring.
However, real time fMRI has a much more important role. First, for fMRI to be used clinically, it is absolutely necessary that a technologist or physician be able to assess the data’s quality. Even more than conventional MRI, motion can completely corrupt a data set. Often, motion artifacts cannot be corrected in post-processing, so the only reasonable option is to rescan if a subject or patient moves significantly. No clinical procedure would be workable—especially something as expensive as fMRI—with even a 10% failure rate. Therefore, being able to see the data and then run a rescan during the same session is very important for clinical applications.
Second, real time fMRI has been used to communicate with or assess locked in or otherwise uncommunicative patients. Rainer Goebel’s group in Maastricht have developed methods for encoding letters in specific activation types.19 They have also developed, using real time fMRI with feedback to the subject, the ability for a subject to move a video game paddle with a specific set of thoughts. They have demonstrated that two individuals, each being scanned, can play “pong” with only their thoughts controlling the paddles. Owen has been able to demonstrate, using real time fMRI, conscious thought processes and memory recall of subjects that are locked in.20
Compelling recent work has demonstrated that fMRI has potential as a therapeutic tool. DeCharms has demonstrated innovative work involving reducing chronic pain in subjects.21 Here, the main regions involved with pain perception first were mapped, and then the amplitude of the signal in these regions was immediately displayed to the subject. The simple instruction given to the subject was to reduce the signal amplitude by whatever mental strategy possible. After several sessions, subjects began to experience some success. These feedback-based mental strategies were correspondingly successful in reducing the pain. In addition, the ability for the subject to remember the learned strategy for reducing the pain intensity continued for several months after these procedures.
Clinical trials that involve the use of real time fMRI feedback to help alleviate depression have begun.22 In these experiments the networks for emotion are mapped and the subject is then instructed to upregulate the fMRI signals associated with the emotional network. After several sessions, many subjects have figured out to upregulate this network, and consequently, feel less depressed.
So far, real time feedback fMRI is still in its infancy as we continue to learn the best feedback methods and timings, the optimal networks to modulate in order to modulate subsequent behavior, and the types of behavior that actually can be modulated. The potential therapeutic implications of resting state fMRI are substantial; however, much work needs to be done to introduce it into clinical practice.