1 Biological Salmon
BECOMING SALMON
‘Of all the families of fishes’, insisted David Starr Jordan, the leading nineteenth-century American ichthyologist, ‘the one most interesting from almost every point of view’ was Salmonidae (Salmonid).1 His British counterparts agreed. Salmonidae’s members, remarked Francis Day, a former Inspector-General of Fisheries in India, ‘are universally admitted to rank second to none in value, whether regarded as food, as affording sport, or interesting objects for study’.2 From the angler’s standpoint, Salmonidae are the most interesting of all fish families because more game fishes belong to it than to any other – its ranks include trout, grayling and char as well as salmon.3 Particularly intriguing from the biologist’s viewpoint is that a number of family members (genera) are anadromous (Greek for ‘running upward’). Like the alewife, striped bass, sturgeon and shad, salmon are conceived and born in freshwater but spend most of their adult lives in the ocean, eventually returning to rivers to spawn. As such, they get the best of both worlds. ‘Like some persons of honour and riches’, observed Izaak Walton, ‘which have both their winter and summer houses’.4 This arrangement reflects the family’s residence within a belt spanning the northern hemisphere, where oceans are generally more productive than freshwater habitat (the reverse applies to southern latitudes). On the Atlantic side, Salmonidae’s historic range lies between the Arctic and 40 degrees, dropping to the 30th parallel in the Pacific.
The oldest-known salmon fossils: 1. Juvenile specimen of Eosalmo driftwoodensis from Driftwood Canyon, British Columbia. |
|

Salmonidae’s fossil record is scanty. The oldest known specimen (just 15 cm long) dates back 50 million years. The bones of Eosalmo driftwoodensis were found in the Eocene sediments of British Columbia’s Driftwood Creek canyon in the 1930s, but a paleoichthyologist (Mark Wilson) first identified it in 1970 (formally naming it in 1977). This particular salmonid, which lived in the lakes of western Canada, resembled today’s grayling and belonged to the class of bony, ray-finned fishes known as teolosts, which also include perch, pike and catfish. The discovery of the fossils of juvenile as well as adult fish suggests that, at this stage, Salmonidae were probably non-migratory.5 Fossilized skulls from an epoch between 5 and 24 million years ago – when Europe, Greenland and North America were connected – indicate that Salmonidae’s various genera were already established. The 12-million-year-old sabre-toothed salmon (Smilodonichthys rastrosus), a leviathan ten feet long and weighing 180 kilograms (the first fossils were found in Oregon in the 1960s), may be a direct ancestor of two of today’s Pacific species, the sockeye and chinook.

2. Adult specimen of Eosalmo drift woodensis from a site in southern British Columbia.
The north Pacific and the north Atlantic house discrete species of salmon, the two genera to which they belong corresponding, in turn, to the respective oceans: Salmo and Oncorhynchus. Of the five species comprising the Atlantic’s Salmo, only one is a salmon (Salmo salar); the rest are trout. The southernmost limit of the historic range of Salmo salar on the North American side is probably Massachusetts. On the European side, Salmo salar frequents the North Sea, Baltic Sea, Barents Sea and White Sea and roams as far south as the rivers of northern Spain that empty into the Bay of Biscay. Despite its distribution from the Gulf of Finland to the Gulf of St Lawrence, North American and European stocks of Salmo salar lack the differences that would divide them into sub-species.

‘Salmo a Salmon’ and ‘Salmulus’ (a Smelt), from Francis Willughby, De Historia Piscium Libri Quatuor, edited by John Ray (1686).
Pliny the Elder bestowed the Atlantic salmon’s formal name in the first century – perhaps inspired by reports from Julius Caesar’s legions, which, marching through Gaul in 56 BC, had noticed the riveting spectacle of a new variety of fish leaping up the Garonne (in Aquitaine, Pliny noted, salmon was the preferred fish).6 Salmo (indicating the genus) probably has its origins in Salmona, the name of a river in east-central Germany (‘Salm’ in German) that is a tributary of the Moselle (a usage that appears in Ausonius of Bordeaux’s fourth-century poem, ‘Mosella’). Salar (denoting the species) may derive from salire (‘to leap’).7
Ten species belong to Oncorhynchus, which ranges from southern Korea to southern California. Seven are salmon, the remainder are trout.8 Oncorhynchus, derived from two latinized Greek words: ‘hooked’ and ‘snout’, was coined in 1861 by George Suckley, a former assistant surgeon and naturalist with the US government’s Pacific Railroad Survey that explored the vast territory between Minnesota and Puget Sound (1853–6). Because the only Pacific salmon he observed were spawning males, whose most striking feature is a hooked lower jaw (kype), Suckley failed to appreciate that male (cock) and female (hen) salmon are sexually dimorphic (physical characteristics vary at different stages of their life cycle) and relegated females and immature males to a different species. Notwithstanding Suckley’s basic error, Oncorhynchus stuck.
The scientific nomenclature for the seven Pacific species is an adaptation of the common names applied by the Koryak peoples of the Kamchatka peninsula in eastern Siberia. These names first appeared in Europe, translated phonetically into German, in Georg Wilhelm Steller’s account (1743/4), posthumously published, of Kamchatka’s natural and human history.9 Johann Julius Walbaum, a physician-naturalist from SchleswigHolstein, first recorded them as official scientific names when he observed the various species in the Kamchatka River (1792).
Oncorhynchus tschawytscha (pronounced cha-vee-cha) is variously known on the North American Pacific coast as King, Spring, Sacramento, Quinnat and Tyee, but most commonly as chinook (after the dominant tribe of the Columbia River basin). Historically, it was distributed from Point Hope in the Alaskan Arctic to southern California’s Ventura River. On the other side of the Pacific, its range extends from the Japanese island of Hokkaido northward into the Bering Sea and round Siberia’s westernmost tip into the Arctic Ocean. Chinooks are now also found in the southern Pacific, having been transplanted to New Zealand’s South Island in the early 1900s and, later, to Chilean waters for aquaculture (escapees have started to colonize rivers descending from the Andes).

The chinook salmon (Oncorhynchus tschawytscha).
The largest salmon by far, the chinook can weigh as much as 45 kilograms. (The heaviest catch on record is 57 kilograms.) Its spawning runs are the earliest of the Pacific species, beginning in spring, and among the longest. Before being stymied by dams in the 1930s and 1940s, chinooks migrated 1,600 kilometres up the Columbia, and still travel between 2,400 and 3,200 kilometres up the (dam-less) Yukon. Oncorhynchus nerka, though some-times known as red due to the colour of its flesh (especially in the canning industry), is most often referred to as sockeye (the anglicization of the Chinook word Sukkegh). Sockeye are more abundant today than chinook – though much smaller at between two and five kilograms – and the species most highly prized by aboriginal and commercial fishers alike. After chum (see below), Nerka travels the greatest distances upriver and spans the Pacific from Kamchatka to British Columbia.
Oncorhynchus gorbuscha is commonly known as pink. Spawning males develop a pronounced hump between head and dorsal fin – hence the other vernacular name of humpback (gorbuscha in Russian). The smallest of the Pacific species at one to five kilograms – and the most abundant – it follows a very regular, two-year spawning cycle, attaining sexual maturity earliest and venturing least far upstream. Transplanted to the Barents Sea and Newfoundland in the late 1950s, a few specimens have been logged in British waters. Oncorhynchus kisutsh (also known as coho, medium red and silver) spawns latest, in autumn, often in smallish streams with short runs, and does not venture far into the sea during its six-year life. After the chinook – the species to which it is the most closely related though considerably smaller at 3 to 6 kilos – the coho is the least plentiful. Cherished especially by anglers, the acrobatic coho was released in the Great Lakes in the 1960s to revive moribund sport fisheries.

The sockeye salmon (Oncorhynchus nerka).

The pink salmon (Oncorhynchus gorbuscha).


The chum salmon (Oncorhynchus keta).
Chum and dog are the most popular North American vernacular names for Oncorhynchus keta (keta means ‘the fish’ in the language of the Nanai peoples who inhabit the Amur River region where China and the Russian Federation meet). Chum probably stems from the Chinook word for striped – tzum/sum – inspired by the flamboyant streaks and blotches that decorate the fish’s body in spawning mode. Dog, the preferred Native Alaskan term, refers partly to its big head but mainly denotes the enlargement of the teeth during spawning, especially those at the tip of the lower jaw, which, Steller observed, assume canine fang shape and may measure half an inch. (That Native Alaskans routinely feed their sled dogs chum salmon adds extra currency.) The chum and its closest relative, the pink, are the most broadly distributed of the Paciic species, extending as far south on the American side as Monterey Bay, California, yet also frequenting the Arctic Ocean from Siberia’s Lena River to Canada’s Mackenzie. On the Asian side, chum even frequent the southernmost islands of Japan. Like the pink, it spends very little time in freshwater, heading seaward when just an inch long. This gives chums and pinks more time to feed and grow in the sea, explaining why they are the most numerous of Pacific salmon (they also undertake the longest migrations).
A further two species are found only in the Asian Pacific: Oncorhynchus masou and Oncorhynchus rhodurus. Oncorhynchus masou, known as the masu or ocean-running yamame (the latter a Japanese-Kanji name meaning ‘mountain woman fish’) is an anadromous fish similar to the coho. In Japan, masu (sakura masu) is popularly known as the cherry salmon because the appearance of spawning adults coincides with cherry blossom season.10 The masu (whose leitmotif is bold transversal stripes) is the most southerly of the Pacific salmon in distribution, with a large run on North Korea’s Tumen River and a presence in Taiwan’s Tachia River. O. rhodorus comes in two forms, the lake-living biwamasu (var. rhodorus) and the stream-dwelling amago (var. macrostomus), whose vermilion spots set it apart from the more widely distributed yamame. Yet identifying the various Pacific species and distinguishing them from their Atlantic counterpart often requires close examination of gum colour as well and the number of teeth and anal fin rays. Most readily grasped, though, is the difference in shape between the salmon of the two oceans: Salmo salar is sleeker – more hydrodynamic.

Ocean-run Salmo salar, from A History of the Fishes of the British Isles, 4 vols (1860–65), by Jonathan Couch, a physician from the Cornish fishing village of Polperro. His skill in the dissection and depiction of fish won him a fellowship of the Linnaean Society.
THE SALMON’S LIFE CYCLE
Other, more important differences stem from their life cycles. Whereas Pacific salmon without exception die after spawning, up to 6 per cent of their Atlantic relatives survive and return to the sea. Another feature setting them apart is the portion of their lives spent at sea. The Atlantic salmon typically resides in fresh water for two to three years before going to sea. In northern Norway and Labrador, it can remain there for eight years – much longer than any Pacific salmon species. Atlantic salmon also spend more time in freshwater on their return to their spawning grounds. The time spent in fresh and saltwater is partly determined by the relative availability of food in these two mediums.
After migrating downstream as young fish, salmon roam the oceans for between one and six years, often travelling thousands of miles. They mingle with others besides fish from their home river. Fish from both sides of the Atlantic share a common feeding ground off western Greenland. Pacific species also become jumbled up. Then, as they mature sexually, salmon perform an astonishing feat of biological reorganization, separating into distinct groups of compatriots that head for their respective home streams (examples of straying to the other side of the Atlantic are extremely rare). Salmon re-enter their rivers year round, but some only run in late summer or autumn.
A profound physiological transformation during a sojourn in brackish water equips the salmon for re-entry into freshwater. Because the salt concentrates in their bodies are usually higher than in the surrounding freshwater, they do not need to drink while in rivers. Water flows into their tissues toward these salt concentrations. In the ocean, however, the salt concentration in the water is much greater than in a fish’s body, so the water flow is reversed. To rectify this water loss, the marine salmon must reverse the operation of its salt pumps to absorb water, expelling excess salt through its gills.
Many Atlantic salmon face relatively short upstream journeys compared to their Pacific cousins. European salmon rivers are often less than a hundred miles long, allowing for a fairly leisurely pace. On Scotland’s Dee and Spey, daily rates of between 0.18 and 0.74 kilometres have been logged. By contrast, some Pacific populations face staggering journeys. Averaging between 16 and 32 kilometres daily, they may clock up a much higher rate if their spawning grounds lie far upstream. At Dawson City, 2,415 kilometres up the Yukon, chum were registered travelling a 80-kilometre daily average in the 1890s. Salmon also need tremendous energy reserves because of the potential delays they face in the shape of floods, low water and obstructions; they may wait weeks below a waterfall until conditions are right for an ascent. Laboratory experiments in which a sockeye swam at a rate of 2.9 kilometres an hour against a stiff current for nearly 13 days without pause (equivalent to 1062 kilometres) suggest the fish’s amazing strength. Another experiment involved observing sockeye on a fish-way’s treadmill; they climbed continuously for five days – the equivalent of 6,648 metres. Eluding predators and surmounting obstacles also demands bursts of speed. And salmon can make faster and more abrupt turns than any other fish.
King salmon jump the flume of a mining company on Copper Creek, Alaska, c. 1907. |
|

On leaving the sea, salmon stop eating. A century ago, many fish biologists and anglers found it barely credible that a fish could go for so long – sometimes up to a year – without eating. ‘It would be inexplicable, indeed’, remarked the American sportsman Charles Hallock, ‘if salmon alone of all creatures, were not required by nature to fortify and strengthen themselves for the supremest act of physical existence’.11 Thomas Huxley, the British scientific all-rounder, was less impressed by their ability to cope on an empty stomach, pointing out (1860) that they had gorged at sea while swimming ‘in a species of animal soup’. 12 Peter Duncan Malloch, a self-taught Scottish authority, conducted some unusual early twentieth-century experiments to confirm the fasting theory. He dropped prawns and sweets off a bridge over the Tay, commenting that ‘they were readily taken, but after a few nibbles the fish allowed them to fall to the bottom’.13 As the salmon lives off the fat stored in its tissues during its ocean pasturage, its stomach shrinks to the size of a finger to make room for the eggs or milt and gonads that eventually occupy most of the body cavity. At the point of spawning, the ovaries may constitute over 15 per cent of the hen’s live body weight, while the cock’s testes are five times heavier than when he quit the sea.
On reaching the coast, salmon run the gauntlet of a predatory horde. If they survive seals, otters, porpoises and herons, they face human fishermen armed with nets, traps, spears and rods. In addition to dams and weirs, other impediments may confront them. Construction of the Canadian Northern Pacific Railroad triggered a massive rockslide that plugged Hell’s Gate canyon, 240 kilometres up British Columbia’s Fraser River, effectively preventing further upstream migration of the major portion of the sockeye’s autumn run in 1913. This was the world’s greatest salmon run and spawning sockeye backed up for 16 kilometres below the blockage, choking the river with red bodies ‘packed together like seeds in a pomegranate’ (to quote a Canadian poet).14
A blocked Hell’s Gate presented too daunting a challenge. Nonetheless, as Walton remarked, salmon can surmount obstacles ‘even to a height beyond common belief’.15 For many (like Pliny) this became the fish’s defining activity (many pubs along British salmon rivers are called The Leaping Salmon’). Andrew Young of Invershin, Scotland, manager of the Duke of Sutherland’s nineteenth-century salmon fisheries, reckoned that an upward, slanting leap of 12 feet (3.6 metres) was within the capacity of most salmon, though few succeeded at the first attempt. Their indefatigability was legendary. In the 1920s, a British salmon biologist observed repeated efforts to scale a six-foot fall that lasted an hour.16

This uncanny ability was once regarded as proof of supernatural powers. Another fanciful and surprisingly tenacious view was that the salmon held its tail in its mouth and bent itself taut like a bow before jerking itself upward by suddenly letting go. In his early seventeenth-century English epic poem, Poly-olbion, Michael Drayton described this accomplishment at the falls (‘Salmons leape’) that rear up as fish enter the Tivy in west Wales.
Heere, when the labouring fish doth at the foote arrive,
And finds that by his strength he does but vainly strive;
His taile takes in his teeth; and, bending like a bowe
That’s to the compasse drawn, aloft himself doth throwe,
Then springing at his height, as doth a little wand
That bended end to end, and started from man’s hand,
Far off itself doth cast; so does the Salmon vault:
And if, at first, he faile, his second Summersaut
Hee instantlie assaies; and, from his nimble Ring
Still yarking, never leaves untill himselfe he fling
Above the streame full top of the surrounded heape17
Others believed that airborne salmon propelled themselves higher by flapping their tails. In actuality, responding to sound vibrations, they negotiate falls by jumping from the spot where most water is falling, diving deep to reach the upward moving water that provides a vital added boost. So, a waterfall considerably lower than 3.6 metres may offer an insuperable challenge if the launch pool is too shallow.
The leaping salmon is undergoing a metamorphosis in appearance and physiology (particularly pronounced in the male) that is often described as a monstrous deformity. The fins thicken and the skin becomes spongy, mucous-covered and leather-like in texture, virtually absorbing the scales. Breeding teeth replace feeding teeth, which, especially in the male, are considerably larger and sharper. The head lengthens into a snout and the body becomes more compressed, developing a cartilaginous hump. Most mature males develop extended upper and lower jaws. The apex of the lower jaw’s dentary bone enlarges and curves upward to form the grotesque-looking kype that so impressed Suckley (Walton compared it to a hawk’s beak).18 Meanwhile, the upper jaw’s premaxillary bone grows and dips downward to accommodate the kype with an interlocking socket, though the mouth no longer closes properly.19
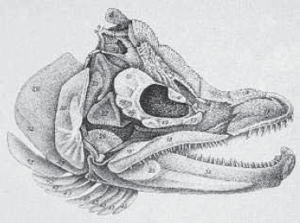

‘Salmo salar (Der Hakenlachs)’ from Ökonomische Naturgeschichte der Fische Deutsch-lands, vol. 3 (1785), by M. E. Bloch, a Berlin physician who supplied an accurate descriptive and visual account of all German species. The use of silver and gold accentuates the sheen of the salmon’s scales.

A cock salmon in spawning livery.
Dazzling transmogrifications in hue accompany this bodily reconfiguration. The silver-sided oceanic fish assumes a striking range of darker colours. Pinks turn slate grey on their backs and sides and creamy white on the ventral surface. Chum undergo the most stunning make-over. Within days of entering freshwater, their sides display jagged red and black (sometimes green and purple) bars, with patches of dappled grey and yellow (hence the archaic name, calico salmon). Chinook and coho assume the trademark dark red that lends spawning rivers a blood-streaked appearance. Sockeye go bright red and their heads olive green. Atlantic salmon also change colour, though the female generally becomes darker and duller than the male. So profound were these changes, remarked Steller, that unless one was familiar with their life cycle ‘they would never be taken for the same fish that enter the rivers’.20
‘Colour changes in spawning salmon’, from William Jardine, Illustrations of British Salmonidae with Descriptions by Sir William Jardine (1839–41). |
|

Sir William Jardine (1800–1874), seventh Baronet of Applegarth (Dumfriesshire, Scotland), drew an exquisite picture of a male weighing 7.25 kilograms taken from a pool in the River Annan, one December during the 1830s. Captured at the height of its spawning phase, the fish sported a brilliant scarlet and orange hue on his sides while his upper parts were brownish. Jardine, who later served as president (1860) of the Royal Commission of Inquiry into the English and Welsh salmon fisheries, supplied a vivid accompanying written description of this ephemeral coat of many colours: The colouring, when newly taken from the river, was beautiful beyond description, and is not to be conveyed by any representation: independent of the brighter tints and rich purplish-grey running along the lateral line, the whole body was, as it were, glazed over with a delicate crimson-pink, changing and fading with the life of the fish’.21
Changes in colour may well be connected to the switch to freshwater, a respiratory response to lower oxygen content. Since salt water contains more oxygen, salmon in the sea go through 40 breathing cycles per minute. In freshwater, they must compensate for their reduced rate of respiration and red pigments such as cartenoid are effective carriers of oxygen. Shedding oceanic colours also provides superior camouflage.22 Charles Darwin took a different line, though, contending that the male’s colours (and hump) were co-called secondary sexual characteristics, i.e. male attributes that females find attractive or that males think females will find attractive. He emphasized the extreme pugnacity of cock salmon, citing reports of a day-long struggle between two males and rivers littered with the bodies of dead combatants. Darwin also interpreted the kype as a defensive weapon against the ‘fangs’ of sexual competitors.23
‘Duelling males’. The first sketch represents a salmon charging a rival; the second, a male covered with honourable scars, after the wars are over. From Lewis Lloyd, Scandinavian Adventures (1854). |

According to some salmon biologists, ‘preliminary’ pairing begins at the spawning grounds.24 Others reckon that they team up much earlier, possibly on entering freshwater. Nobody has suggested, though, that salmon are monogamous. But they are very fussy about where they mate. Spawning grounds must supply exactly the right kind of water and gravel (meaning pebbles rather than small fragments of stone). The water must be clear, cool and fast flowing – sediment and vegetal growth associated with sluggish water inhibit hatching – and devoid of predators. Riffles (a patch of shallow but fast-moving water) are ideal for digging a redd (a Celtic term for spawning pit now in universal use). If the water is deep enough, the hen assesses the gravel’s quality by assuming a vertical position and fanning her tail furiously to dislodge it. She then plunges down to ‘feel’ with her anal fin; gravel must be sufficiently loose to allow circulating water to oxygenate the eggs yet not so loose that digging a trough would be like trying to excavate a hole in dry sand.

‘Female Salmon Cutting a Redd with her Tail’, from P. D. Malloch, Life-History and Habits of the Salmon, Sea-Trout, Trout, and Other Freshwater Fish (1912).
If satisfied with the gravel’s condition – and having identified a satisfactory mate – the hen ‘cuts’ a redd. Lying on her side, sometimes in water so shallow there is barely water to cover her back, she fans her tail to create upward suction. Assisted by the current, this action dislodges gravel and creates gaps, also serving to loosen and lift off any silt and sand that may jeopardize the survival of eggs and fry (ova suffocate without sufficient oxygen). Redds can be circular or trench and furrow-like, and are between 8 and 30 centimetres deep. According to some commentators, female salmon can shift gravel as large as a half brick. This sounds implausible. But heavy objects are much lighter in water and the current is a great help. Having deposited gravel the size of eggs in a trough of water, C. F. Walsh of Dundee recorded that ‘holding a dead fish by the head and on its side, I gently undulated it, and I found the stones were puffed away as if by a gentle breeze of wind’.25 She sometimes interrupts her cutting to drive off female intruders.
Wedded to conventional notions of the division of labour between male and female, many Victorian scientists insisted that the cock fish dug the redd. However, Walsh, who witnessed hundreds of nineteenth-century spawnings, reported that ‘I never saw the male fish take any part in the work; the fanning up of the gravel is all done by the female. I say fanning, because I never saw any boring of the head into the ground’.26 Dissenters from Victorian scientific orthodoxy believed that the spawning male’s head injuries were sustained fighting off rivals rather than from digging. Walsh’s views are no longer disputed.
Meanwhile, like a rutting stag, her mate stands guard to repel other males. These contests could be bloody and exhausting. One Victorian observer recorded that ‘there was one fellow who appeared to be king. Whenever [another male] attempted to come too near the female he made a rush at him, seized him and shook him as a retriever would shake a hare’.27 Other contemporaries disputed the gravity of these encounters, claiming that the kype’s function was precisely to prevent males from inflicting grievous harm, because it prevented them opening their mouths wide enough to grab their opponents with their savage teeth.
When done digging, still lying on her side, the hen sinks to the bottom of the trough. By flexing her tail, she extrudes thousands of eggs (900 for each 450 grams of body weight is the norm). The male then squirts a milky substance called milt (sperm) over them. Other Victorian accounts relate a more intimate act. The male moves alongside and vibrates his body vigorously against her for a few seconds (a muscular spasm preparing for milt delivery that has been described as ‘quivers just like the final shudder of a dog after it has been in the water’).28 Rubbing against each other, they eject ova and milt simultaneously in an act sometimes referred to as an orgasm (though a British angler living in Scandinavia, Alexander Keiller, was keen to reassure that ‘nothing that could be construed as sexual intercourse took place between them’).29 Another male may release his cloud of milt at the same time and dominant males may mate with other hens if there’s a surplus. The female fills in the redd with what remains of her battered tail.

A spawning pair of Atlantic salmon lie on their sides, virtually out of the water, the female’s excavating tail the only submerged body part; from Malloch’s Life-History and Habits of the Salmon . . . (1912).
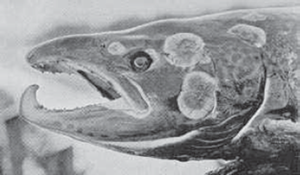
‘Head of a MaleGrilse (7 lb.) Afflicted by Fungus, December 1905’, from Malloch’s Life-History and Habits of the Salmon . . . (1912).

Two scraped sections reveal the thickness of the encrusted fungus, much of which developed after the fish had died; from Malloch’s Life-History and Habits of the Salmon . . . (1912).
The grim reality that spawning colours, however resplendent, cannot disguise, is that salmon bodies are steadily disintegrating. Worms infest the gills. Flesh becomes soft, watery and white from loss of oil. The fungus, Saprolegnia ferox, appears on the flanks and head – the external manifestation of the microbe Bacillus salmonis pestis (so-called salmon disease). This microbe enters a skin abrasion and spreads, attacking the tissue. Having established itself on dead tissue, the fungus invades living tissue and continues to spread after death. Having lost between a third and a half of its body weight, the spawned out salmon is emaciated and exhausted (digging a redd can take three days and nights). If a member of a Pacific species, it invariably dies within weeks – literally starving to death. Roderick Haig-Brown, an American angler, described the death of a female chinook (Spring’) in her spawning pool, standing guard over her eggs (though sitting like a bird on thousands of eggs is not an option).

Dead bodies of spawned pink salmon washed up in the shallows of the Cheakamus River, British Columbia, 1963.
The strong current caught Spring as she went down from the surface. It drew her to itself, rolled her over and swept her on and she no longer resisted. Her tail moved once or twice, feebly, but all the urgencies, all the desires that had driven life thru her were spent. So she lay quietly across the stream flow, drifting as no strong salmon does and the water opened her gill plates and forced under them and she died.30
These mass die-offs were not a pretty sight. The passage of the river is a sickly spectacle’, remarked Hallock, ‘maimed and decaying fish in myriads offending sight and smell, and befouling the entire length of the water course from the sea to its springheads.’31
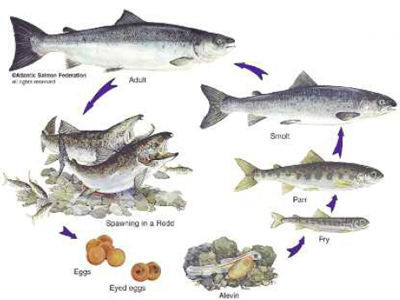
Walton’s reference to the spawning bed as a ‘grave’32 initially seems odd but is actually highly appropriate for a place of birth in view of this symbiotic relationship between death and new life. Once the ova hatches, the salmon’s life cycle goes through seven stages: alevin, fry, parr, smolt, grilse, mature adult and kelt. The time it takes to run its course varies from just over a year to a maximum of ten. The duration of each stage also varies enormously. The offspring of the spawning season of 1913 on the Wye returned to breed in phases over the next six years – a staggering mechanism to militate against fluctuating extremes of bumper and lean crops of young fish.
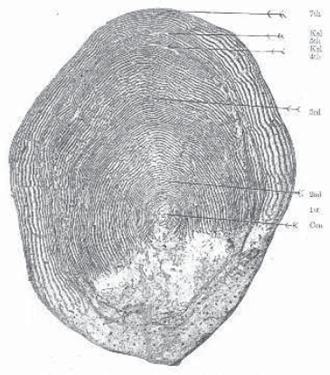
A salmon’s age and life history can be determined by scrutinizing the rings on its scales under a microscope (a variation on counting tree rings, though salmon produce several each year). As fish grow, the number of scales remains the same but each one gets bigger. The proximity of the rings – which resemble roughly circular concentric ridges – reveals a good deal. Periods of rapid summer growth are marked by pronounced, widely spaced rings. Colder periods with less abundant food are marked by fainter, more closely spaced rings, a wide band and a narrow band indicating a typical year.
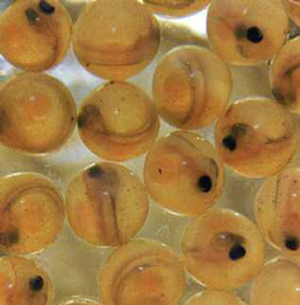
The first three stages of the salmon’s life are exclusive to freshwater habitat. The pink, pea-sized eggs remain buried a few months, depending on season and water temperature. For successful spawning, the water must be under 58°F The creature that hatches, the alevin, is effectively an orphan, a large-headed and bulging-eyed infant that must sustain itselfon the orange yolk in its umbilical sac during its first few months. The alevin then becomes a fry. Measuring about an inch and a quarter at first, its basic food is diatoms – tiny brown algae (phytoplankton) that thrive in the bright light of open-canopied stream banks.
When a fry is as long as a man’s middle finger – usually by the end of its first summer – it changes into a parr. The parr (or samlet) begins the journey downstream, floating tail-first so that water does not enter its gills. Though readily identifiable from the bluish-olive fingerprint-like marks that run down its sides (another reason for the alternative name, fingerling), young parr are easily confused with other juvenile Salmonidae. Sir Humphry Davy, the English chemist renowned as the inventor of the miner’s safety lamp, argued that the parr was the hybrid offspring (mule) of salmon and trout.33 But the first artificial raising of salmon eggs (1836–7) confirmed that parr were young salmon.
|
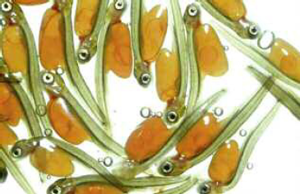
‘Young Salmon, About a Week Old’, from Frank Buckland, Manual of Salmon and Trout Hatching (1864). |
|
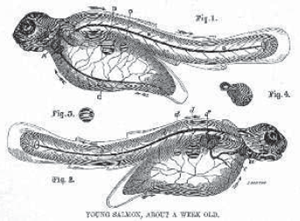

Feeding on freshly hatched flies, the parr measures four to ten inches by the time it reaches estuarine waters. Here, a profound and relatively rare kind of internal change converts it into a fish that can live in saltwater. Smolt also assume the livery (to use Victorian parlance) of the adult salmon, which is better suited to life in the sea. A blue back, white belly and silver sides provide perfect camouflage (the silvery, iridescent scales result from an increase in the amount of guanine – awhitish substance).
The grilse is an adolescent male that attains sexual maturity early and makes a precocious attempt to spawn after just a few months at sea (or a single winter). Like the parr, the grilse perplexed fish biologists for centuries, though no one still insists that they constitute a separate species. The grilse’s tail is more forked than a mature adult salmon’s and its wrist is thinner, lending it a ‘dainty and delicate appearance, quite different from the somewhat rotund outline of the sea-trout, and more graceful than the decidedly athletic proportions of the adult salmon’.34 The grilse probably quits the ocean so soon as part of an evolutionary insurance policy. The production of a reserve supply of willing and able males ensures that any potentially unfertilized eggs are not wasted.
A spawned salmon is called a kelt, or, sometimes, in the case of the male Atlantic, a kipper (‘black, cadaverous fish with heads seemingly enormous, and lank, slimy bodies, repulsive to themselves and abhorred by the anglers’).35 Spawned salmon lose control over their swim bladders and float belly up. They bounce against rocks and flap in the shallows. Those few kelt that survive (exclusively Salmo salar) begin to repair themselves as they head downstream, completing their restoration at sea. This includes shedding the kype, as it would be impossible to breathe and feed in the ocean if still encumbered with it. At this ‘well-mended’ stage, the characteristic silvery hue has returned, making them practically indistinguishable from ‘clean’ fish. Under 10 per cent of mended kelts return to spawn again. And very few are male. This seems odd, initially, for females appear to have expended far more energy digging and refilling redds. But fighting off competitor males is evidently more tiring.

‘Parr or Samlet’, from Sarah (Mrs T. Edward) Bowdich, The Fresh-Water Fishes of Great Britain (1828). Bowdich accepted the orthodox view that he parr (‘skegger’) was a distinct species rather than a young salmon.
The salmon’s life in freshwater is far better understood than its oceanic existence. It has become increasingly clear, though, that they don’t loiter in coastal waters. The recapture of tagged fish raised in hatcheries in Canada’s Maritime Provinces in the early 1900s indicated that Atlantic salmon travelled up to a thousand miles from their natal streams.36 The longest recorded distance is 5,630 kilometres: a chinook tagged in the central Aleutian islands was recaptured a year later in Idaho’s Salmon River. Salmon tend to congregate in areas of up-welling, where prevailing offshore winds drive surface waters away from land, allowing currents to rise vertically to the surface, bringing up nutrients from the bottom. The urge to migrate is evidently connected to the urge to mate, which presumably becomes more powerful than the desire to feed.
The rate of return, though, is extremely low. Salmon eggs are remarkably tough. To test their resilience, one researcher dropped an egg from waist height, recording that ‘it rebounds quite in the manner of a tennis-ball, and externally remains equally undamaged’.37 But their survival rate is nonetheless poor. Some are washed downstream and others are never fertilized. Many creatures prey on them, including their parents. The tini est alevin and the largest parr are vulnerable to trout and pike as well, also eagles, eels and otters. The fierce otter that Tom encounters in Charles Kingsley’s fantasy novel, The Water-Babies (1863), explains that salmon are

‘Severn Salmon’, from Sarah Bowdich’s Fresh-Water Fishes . . . (1828). Bowdich was convinced that the true colours of a fish could only be captured at the moment that it was removed from the water. Her plates are hand coloured and use gold and silver foil to recreate the metallic shine and shimmer of fish scales.

‘Atlantic Grilse’, from William Jardine, Illustrations of British Salmonidae with Descriptions (1839–41).
great fish, nice fish to eat. They are the lords of the fish, and we are the lords of the salmon . . . We hunt them up and down the pools, and drive them up into a corner . . . and we catch them, but we disdain to eat them all; we just bite out their soft throats and suck their sweet juice . . . and then throw them away, and go catch another.38
Young salmon that elude river-based predators and make it to the ocean are eaten by fish ranging from Greenland shark to cod. Figures for the sockeye indicate that, of approximately 3,600 eggs laid, 106 smolt enter the sea, of which 10 will return to their natal stream. Of these, eight will die during the upstream journey, leaving two to spawn – the minimum number required to sustain existing populations.
Hector Boethius (Boece), a Scottish ecclesiastic, provided the first more or less reliable account of this ‘strange’ fish’s ‘procreation and nature’ (1527).39 Boethius – doubtless familiar with salmon from the rivers Dee and Don, which empty into the North Sea at Aberdeen, where he served as the university’s first principal – explained that ‘salmond’ ‘with maist fervent desire and appetite returnis to the samyn placis, quhair thay wer generit’.40 In the fourth volume of his magisterial Historiae Animalium (1558) – a compendium of all existing classical and medieval knowledge of a zoological nature, upgraded with contemporary observations – Conrad Gesner (1516–65), a polymath scholar from Zurich, supported Boethius’ account of freshwater spawning. And Walton cited what may have been the first tagging experiments to support the belief that ‘every salmon usually returns to the same river in which it was bred’. Ribbon, he reported, was tied to the tails of juveniles caught in weirs as they swam seaward; about six months later, some were recaptured with ribbons intact at the same spot.41 By the nineteenth century, ribbons had been replaced with copper and brass wire that often inflicted serious cuts. Other researchers simply sliced off the small, fleshy fin in front of the tail (adipose fin).
The parent stream and the natal stream are usually but not necessarily the same. And if fertilized eggs are relocated, the salmon that hatch will return to the stream where they were born rather than to the stream where they were conceived. In 1880, Jordan referred to this ‘extraordinary instinct’ to spawn where they were born as ‘one of the most difficult problems in animal psychology’.42 Most nineteenth and early twentieth-century ichthyologists scorned the notion of a homing instinct, reasoning that fish did not possess large enough brains to navigate so precisely. Scientific orthodoxy held that salmon did not venture far out to sea, and had the choice of any number of rivers for spawning.43 Given, though, that they remained within the orbit of their natal stream, by default some would return there. As well as seeming impossible, there appeared to be no good motive for a return to the natal stream.
These scientists lacked an understanding of local adaptation and its ecological and evolutionary importance. For them, chinook that spawned in California were no different from chinook that spawned in Alaska. This failure to appreciate differences was not entirely unreasonable. After all, different species and populations intermingle in the ocean to an enormous degree. Spawning rivers are also shared: all five species in the North American Pacific use different parts of the same river or the same parts at different times. Believing that it made no odds if fertilized ova from Alaskan chinooks were deposited in a stream in California, the first artificial propagation programmes in the late nineteenth century assembled a melting pot of stock. This weakened the gene pool (there are no known cases of hybrids between species or within species in the wild).
Having finally embraced natal stream theory, it remained for ichthyologists to explain its mechanisms. Salmon lack the cerebrum and neo-cortex organs that permit higher intelligence. But they have excellent eyesight, even in turbid water, and can see colours. More significantly, their sense of smell is acute. Our ocularcentric modern western culture underestimates the power of smell. Yet smell is the biotic world’s oldest sense, perfected hundreds of millions of years ago by fish that developed the first noses. All but the most primitive fish forms have two external nostrils connected to a nose brain, one on each side of the head, and can smell in ‘stereo’.44 Smell’, remarked Jean Jacques Rousseau, ‘is the sense of memory and desire’, and one of the first to identify this as the vital tool was Frank Buckland, Victorian natural scientist extraordinaire. Using the expression ‘to follow his nose’, Buckland provided a charming, if erroneous image of a salmon glued to an odour trail like a hunting dog. Thus a salmon coming up from the sea into the Bristol Channel would get a smell of water meeting him. “I am a Wye salmon”, he would say to himself. “This is not the Wye water; it’s the wrong tap, it’s the Usk. I must go a few miles further on”, and he gets up steam again.’45
Though this line of inquiry was not pursued at the time, it was resumed in the 1950s, when Arthur Hasler confirmed the integral role of olfactory imprinting. The internationally renowned freshwater ecologist had his brainwave while walking along a mountain stream near his hometown in Utah. Stimulated by the scent of native wildflowers, he started thinking about the power of smell to evoke childhood memories. Freshwater minnows could be trained to distinguish between water from two rivers. But when their olfactory organs were damaged or destroyed, they proved unable to find their way home. On the other hand, blinded fish with smell organs intact returned successfully. Further experiments exposed hatchery-raised coho to odorant chemicals before release into Lake Michigan. Two years later, when they were ready to return to their native streams, Hasler’s team laced one stream near the original release site with morpholine and another with phenethyl alcohol. Of the returning fish that were monitored, over 90 per cent were found in the stream carrying the chemical to which they had been exposed as hatchlings.46 Hasler’s findings indicated that each stream has a unique chemical fingerprint derived from its peculiar mix of soil, flora and fauna, whose presence is unaffected by pollution and fluctuating water levels. During their return journey, salmon follow the sequence of smells in reverse order, progressively getting closer to the only right smell.
The odour hypothesis seems much more believable with regard to a salmon that spawns at no great distance up a big river than in the case of a salmon that faces an upstream journey of 1,600 kilometres, followed by an additional trip up a small tributary. How can the home stream’s signature smell remain discernible in an estuary into which a major river pours the waters of an enormous catchment area? Eel larvae manage to find their way back from the Sargasso Sea to their freshwater habitat in Europe across 8,000 kilometres of ocean. And they can detect unbelievably low concentrations of alcohol (comparable to one shot of vodka in Lake Erie – the world’s eleventh largest lake). So the salmon’s ability to discriminate between smells should not be underestimated. Hasler drew a homely analogy: ‘The first fragrance of the morning coffee is tantalizingly sharp, but as it permeates the air, one ceases to be aware of it: the olfactory organs have become fatigued to it. However, other scents – bacon and eggs, oatmeal, pancakes – can cut through the coffee odour and be perceived immediately despite the fatiguing or accommodation of the olfactory sense to the coffee.’47 Like the eel’s, the salmon’s sense of smell is so keen that it can detect the most dilute concentration of home stream odour amongst a myriad of other smells.
How, though, do salmon find their way from open sea to coast? And how do they know which direction to follow on reaching the shore? Can we accept that the odour trail begins thousands of kilometres out at sea? In 1960s experiments, gill nets with descending horizontal sections mostly captured salmon in their upper layers, suggesting that they navigate using a sun compass mechanism. Hasler’s experiments with white bass in a lake indicated that fish – identified with yellow floats – that were observed on cloudy days or that had opaque plastic eye caps attached had trouble orientating themselves. ‘That an animal is actually able innately to accomplish something for which men require instruments, charts, and tables’, remarked Hasler, is quite incredible.’48 That salmon (like migrating birds and turtles) employ celestial navigation methods is the most popular current explanation of their movement from ocean to coast. Then the distinctive odour of the home stream takes charge.
Salmo (salm) and Sälmling (Samlet), from Conrad Gesner, Tomus IV: Et Ultimus oder Fisch-Buch (1670). |

The evolutionary purpose of homing is tied to the fact that each river has its distinctive salmon stock’ or population’ (races’ and ‘tribes’ were the terms employed when this notion was first mooted in the 1920s). A population is a unit comprising the individual members of a species living in a specific locale that share a genetic ancestry and are much more likely to breed amongst themselves than with individuals from another comparable unit. There is only one species of Atlantic salmon, but those that spawn on the North American coast and their European counterparts are genetically different. The timing of spawning runs and smolt migrations are simple ways to identify different populations.
Physical and behavioural traits carefully honed over generations confer a survival advantage on a specific stream’s population. Yet home-stream fidelity also has its downside. When a river is lost to a log jam or dam, its stock is also lost. By the same token, salmon can fail to exploit fresh opportunities. Experiments in eastern Canada (1938–9) indicated that spawning salmon only used one of a river’s two branches. Though the second branch was available at that time, a barrier had effectively excluded all salmon from it until the late 1920s. So, even though this tributary’s potential spawning grounds had been open for a decade, salmon ignored them in deference to their time-honoured destination. In this sense, the umbilical link between population and stream can be self-destructive. To write off a perfectly suitable spawning river forever, however, makes no evolutionary sense. And so each salmon population generally includes a few enterprising individuals capable of colonizing new habitat.49 This explains why a small number (about 2 per cent) occasionally ‘stray’ elsewhere to spawn. This also permits eventual recolonization of a stream whose local population has been extinguished. Straying also facilitates the occupation of salmon habitat that humans create. Before the mid-nineteenth century, the Ballisodare River in County Mayo, Ireland, had never seen a salmon. Where it empties into the Atlantic, a series of natural falls and rapids rule out upward migration. By 1855, though, thanks to installation of fish passes, hatchery raised salmon could enter what has become one of Ireland’s most productive salmon rivers (while ova planted higher up could descend).
We tend to think of evolution as a process that takes thousands of years and transpires without human agency. Yet it can occur within a single human lifetime. Within 30 generations, distinctive life-history traits have evolved among specific ocean running populations of chinooks transplanted to New Zealand between 1901 and 1907. They also look different. Females undertaking the longer spawning runs tend to be bigger and to lose more body weight during their migrations. That neither physiological identity nor range are frozen in time will become clearer when this discussion of the wild fish is supplemented (and complicated) in the next two chapters by coverage of salmon raised in hatcheries and produced on fish farms. The meat and drink of the next chapter, though, is the salmon in its most familiar role. This initial examination of the biological salmon has only incidentally been concerned with food – dwelling mostly on why salmon don’t eat after they re-enter freshwater. Now the edible salmon swims into view as we explore its identity as sustenance for other creatures.