On the previous evening my father and I, with two companions, had entered the Singer Preserve, near Tallulah. This area at the time was a great virgin bottomland forest. We were in the quest of America’s rarest bird, a species that few living ornithologists had ever seen except as a museum specimen... After several unsuccessful attempts to see this great woodpecker in the Singer Preserve in the summer of 1934, I was still trying on the Christmas day mentioned above. My companions and I were out at daybreak... A slow drizzling rain that began to fall did not seem to better our prospects, but suddenly, in the distance through the great wood, a telltale sound reached our ears. Approaching cautiously in the direction indicated by the calls, we soon beheld not one but four Ivory-bills feeding on a tall dead snag! There were two males and two females, which with their powerful bills were proceeding to demolish the bark on this dead tree, in search, no doubt, for flat-headed beetles, or “betsy-bugs”...
It is possible that no future generation of Americans will be able to spend a Christmas morning, watching four Ivory-billed Woodpeckers go about their daily routine amid huge redgums whose diameters are greater than the distance a man can stretch his arms. I wonder what natural beauties we shall have, aside from the mountains and the sky, a hundred years from now!—George H. Lowery, Jr. (Louisiana Birds, 1955)
Sadly, George Lowery’s concerns proved to be well founded. It is highly likely that the ivory-billed woodpecker (Campephillus principalis) no longer exists. It was a striking creature in the floodplain forests of the great southern rivers of the United States. An extremely large black-and-white woodpecker with a red crest and white bill, it was never common. Very tame, it made an easy target for commercial hunters, who shot the animals to sell as specimens for wealthy gentleman collectors. This practice certainly decimated the population, but the bird’s vulnerability ultimately stemmed from its need for extensive areas of mature floodplain forest.
The highest regional density ever recorded for ivory-billed woodpeckers was only about one pair per 36 square miles (50 km2).1 The reason for this low density appears to be the bird’s specialized feeding habits and the scarcity of its feeding stations. As Lowery and others observed, the bird got its food by pecking away the loose rotting bark from large standing dead trees (Figure 1). Once a woodpecker had removed all the bark from a dead tree, it needed another standing dead tree with loose bark and available insects as a replacement feeding location.
How many large dead snags does a floodplain forest contain? At what rate are these replaced, to provide an Ivory-billed woodpecker with a steady supply of places to feed?
Unfortunately, such snags appear rarely. A suitable dead tree occurs 200 to 400 years after the tree’s birth as a seedling, its growth to maturity, and its death. Only when a large, mature tree dies while still standing will the resultant snag be usable by a feeding specialist such as the ivory-billed woodpecker—and then only for a year or two.
This consumption of a rare element of the forest illustrates how the mature forest is a dynamically changing and continually regenerating mosaic. If there is enough of it, this forest mosaic can regenerate the feeding habitat of the ivory-billed woodpecker as a sustained and renewable resource. Because of the slow rate at which feeding sites are produced, a large area of land is needed for enough dead trees to allow a pair of birds to raise a brood of young. This requirement is at the heart of the woodpecker’s eventual demise. The bird’s rarity requires an understanding of forest dynamics.
Vegetation looks like broccoli when viewed from above at some level—it is clumped and patchy.2 A forest canopy is a mosaic of tree crowns, interspersed with holes in the canopy (Figure 2). The holes, which forest ecologists call gaps, result from the violent deaths of large trees. A rich body of theory about the ever-changing nature of landscape mosaics is based on these forest gaps and their changes over time. The central concept is that the elements constituting the forest mosaic change cyclically; a canopy gap is one phase of this cycle.
Figure 1. Three ivory-billed woodpeckers (Campephillus principalis) feeding on beetles by flaking the loose bark from a dead tree. From J. J. Audubon, The Birds of America, vol. 4 (Philadelphia: J. B. Chevalier, 1840).
Figure 2. The Australian subtropical rain forest in Lamington National Park (Queensland, Australia), showing the forest canopy as a mosaic of trees.
To visualize the forest cycle, imagine that you are standing in a mature forest viewing a small plot of land dominated by a single large tree. The large tree shades the ground, thereby reducing the survival of smaller trees and seedlings below. A few smaller trees of shade-tolerant species survive under the large tree, but they are spindly and their growth is suppressed. The canopy tree dominates the available resources (light, water, and nutrients).
If it were possible to speed up time, how would the forest change from this starting point? When the dominant tree dies, the forest floor (where previously a young tree had little chance of a survival) abruptly becomes a nursery for small seedlings and saplings. With adequate light and other resources, hundreds of seeds germinate. The resultant small seedlings survive and begin to grow toward the canopy. The small trees that struggled under the dominance of the now-dead canopy tree begin a race -with the seedlings. They compete -with one another to become the tallest. Some lose to more vigorously growing competitors. Eventually, one tree becomes the locally dominant canopy tree and begins to eliminate the others. The site returns to a state much like the starting point and closes the mosaic cycle. A large tree again dominates the site. When it dies, the cycle begins again.3
Forest ecologists have generated much of the theory about landscapes as dynamic mosaics. A. S. Watt, a forester, developed his doctoral research in 1925 on the pattern of vegetation in a mature European beech (Fagus sylvatica) forest in southern England.4 His surveys led him to develop an important concept in ecology: that uniform processes are responsible for the heterogeneous patterns seen in all vegetation. Over the next two decades, Watt perfected his ideas by studying a wide range of British vegetation types. In 1947, by then a recognized leader in plant ecology, he gave the presidential address before the British Ecological Society and extended these concepts to a range of other systems (bogs, sparse grasslands, alpine communities, heaths).5 Watt’s address, a classic paper in modern ecology, presented two sorts of examples to support his theory.
First, he developed cases in which he systematized a seemingly helter-skelter spatial pattern of vegetation mosaic to elucidate the underlying processes causing the pattern. Interpretation required the observer to deduce the appropriate ordering of the patterns in the vegetation. Once this botanical jigsaw puzzle had been solved and the correct order determined, the process causing the pattern was revealed.
For example, the heterogeneity of the mature European beech forest at the core of his doctoral work (Figure 3) was the product of underlying cyclic dynamics. The pattern of the forest resulted from the fact that different parts of the forest floor were in different parts of the underlying cycle. Watt documented several other such explanations of heterogeneity in very different British ecosystems: the patchy surface of a windswept sparse grassland on sandy soil in Breckland, near the center of Norfolk; clumps of different kinds of shrubs in British heath-lands; the hummocks and hollows of moors. In all of these examples, the size of the dominant plants and the cycle of the colonizing, growing, maturing, and senescing of individual plants created a mosaic pattern across the landscape.
Second, Watt discussed cases in which the cyclic processes seen in the examples above were naturally synchronized to some degree, usually by environmental conditions. No longer helter-skelter, the synchronization resulted in vegetation patterns that moved in a predictable way over the landscape. Watt’s examples were from vegetation in relatively adverse locations, notably from high in the Cairngorm Mountains in Scotland. They involved the same species—shrubs such as the common heather (Calluna vulgaris) and low-statured herbs and grasses. These cases are compelling because there is no need to infer the ordering of the patterns of the vegetation to understand the underlying processes that generate the patterns; the vegetation literally arranges itself.
Figure 3. Mosaic dynamics of mature European beech (Fagus sylvatica) forests in England. Above, Cross-section through a mature European beech forest. Notice the variation in the sizes and arrangement of the trees, as well as in the ground vegetation (Rubus, Oxalis). Below, Rearrangement of the variation in vegetation as different stages of the gap replacement process. From A. S. Watt. Pattern and process in the plant community, Journal of Ecology 35 (1941)1–22.
Since Watt’s 1947 paper, vegetation of larger stature has demonstrated this same tendency to be arranged in regular patterns. Two examples of these so-called self-organizing systems are “tiger bush” and “fir waves.” Both reinforce Watt’s interpretation of the cyclic processes of growth, death, and rebuilding as the generators of vegetation pattern.
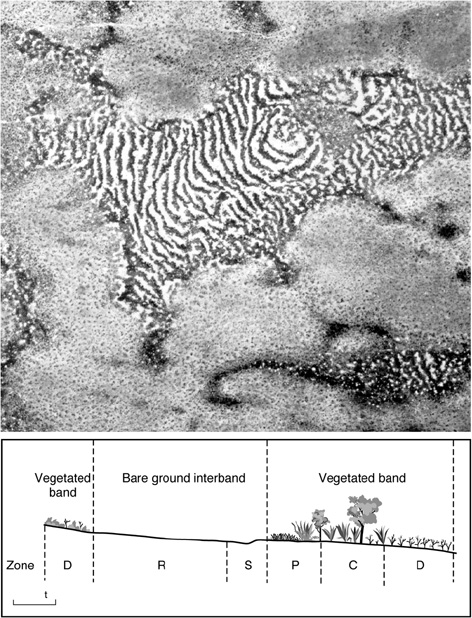
Tiger bush occurs in regions of arid or semiarid climates with infrequent but highly intense rains. In mapping surveys of the Sahara Desert after World War II, extensive aerial observations revealed parallel stripes of alternating vegetated and nonvegetated zones.6 These patterns resemble the stripes of a tiger; hence the English name tiger bush, or the French brousse tigrée. “ This strange phenomenon (Figure 4) occurs in Africa, North America, and Australia.7
Tiger bush is found on flat areas with gentle, regular slopes where the soils are not very permeable. When it rains, water does not penetrate the soils and often produces runoff that flows over the surface in sheets. Where there is vegetation, the soil structure is altered by the presence of roots and leaf litter and allows rainfall and runoff to soak into the soil. This moisture irrigates the vegetated band that forms the tiger “stripe.” The plants at the front of the tiger stripe have first access to water, then the stripes migrate slowly up the slope.
For example, in the Mapimi Biosphere Reserve in the Chihuahuan Desert of Mexico, the stripes have widths between 65 and 230 feet (20–70 m) and lengths of 300 to 1,000 feet (100–300 m). The areas covered by such patterns can be extensive. For example, tiger bush covers 32 percent of a 664-square-mile area (172,000 ha) mapped in Mapimi.8 Within each stripe are five relatively distinctive zones. The major woody species forming the vegetation stripes (in Mexico, Prosopis glandulosa and Flourensia cernua) regenerate better on the “first-to-drink” or upslope side of the stripe. Plants colonize this side during years of relatively high rainfall. Plants die on the “last-to-drink” down-slope side of the stripe in relatively dry years.9 Thus, the movement pattern of the stripes varies, but generally it reflects the regional climate variability.
Basically, the tiger stripes arise from a feedback interaction of the vegetation, the soil, and the climate. The presence of vegetation increases the amount of dead plant material in the soil, which in turn significantly increases the rate at which water penetrates the soil in the zone above the vegetation stripe. The soil area ahead of the vegetation gets more water and becomes more hospitable to plant growth. As a result, the vegetation crawls slowly up the gentle slope of the desert, propelled by colonization of the now-moister zone.
Our immediate suspicion is that some organizing force (such as hedgerows, tree plantations, or stripes of willows along a river) produces regular patterns of vegetation. But tiger stripe vegetation surprises intuition: brousse tigrée in Niger and tiger bush in Mexico organize themselves into patterns, just as Watt’s examples from the Cairngorm Mountains do.
Figure 4. Above, Tiger bush, or brousse tigrée, photographed from the air in Niger. The dark stripes are trees and shrubs. The photograph covers approximately 2 miles (3 km) on each side. Below, Cross-section of a brousse tigrée showing different vegetation zones. The soil in zone R (the run-off zone) has a surface layer that is difficult for the water to penetrate. The runoff water penetrates the soil when it reaches zone S (sedimentation zone) because the organic matter produced by the vegetated band loosens the texture of the soil. This penetrating water irrigates the vegetated band. Zone P (pioneer front zone) is the youngest vegetation, zone C (central zone) has the tallest vegetation, and in zone D (degraded zone) the vegetation is in a state of decline. The vegetation moves from right to left in the diagram because the upslope areas have access to rainwater before the downslope parts. From J. M. Thiery, J.-M. D’Herbès, and C. Valentin, A model simulating the genesis of banded vegetation patterns in Niger, Journal of Ecology 83 (1995):491–501.
Consideration of a surprising geometric regularity in the vegetation of the Sahara Desert seems a leap from understanding the habitat of a large woodpecker in the floodplains of the southern United States. However, the brousse tigrée is a clear illustration that vegetation at fine spatial scales changes cyclically. The bizarre geometry of the tiger stripes reveals the underlying dynamic cycle of the vegetation. A next example moves closer to the forests of the ivory-billed woodpecker.
The second instance of self-organizing patterns is a naturally forming landscape geometry described as “fir waves” or “wave regeneration.”10 Crescent-shaped bands (or, in some cases, parallel stripes) of dead trees are arrayed regularly across mountainsides. Their appearance in the high-altitude forest of Japan has been described thus: “In the dark green of the gentle southwest slope of Mt. Shimagaree with subalpine coniferous forest, several whitish stripes horizontally running in parallel with each other can be seen in a distant view so distinctly that the mountain has been named ‘mountain with dead trees stripes.’”11 Fir waves have been observed in high-altitude conifer forests at several locations around the world.
In New England, the stripes of dead trees represent a slowly moving wave of synchronized mortality of balsam fir (Abies balsamea) trees. This wave of death is followed by a trailing wave of regenerating and growing trees, which in turn is followed by another wave of death. The waves travel as if propelled by the prevailing winds. The time needed for a wave to travel through an entire cycle is on the order of fifty-five years, which means that the movements of the waves are from 3 to 10 feet (1–3 m) a year.12 Faster-moving waves are found either on level ground or moving downhill.
Fir waves strongly resemble Watt’s inferred pattern of gap regeneration. His diagram of forests generating and repairing canopy gaps caused by the death of large trees (Figure 3) looks exactly like a cross-section through a fir wave found in the balsam fir forest in Maine (Figure 5). Internal ecological processes interact with environmental factors, giving fir waves a capacity for self-organization. These are special cases in which the underlying interactions discussed by Watt for forest are directly observable on the landscape.
Figure 5. Balsam fir (Abies balsamea) waves in the mountains of Maine. Above, The white stripes are the exposed trunks of dead trees (photo by Meg Ounsworth). Below, A cross-section through a fir wave. From D. G. Sprugel, Dynamic structure of wave-generated Abies balsamea forests in the northeastern United States, Journal of Ecology 64 (1976):889–911.
Because Watt so strongly joined “processes” (death of trees, growth of trees, germination of seedlings) to “patterns” (tiger stripes or fir waves), his ideas are often captured in the phrase “pattern and process.” That processes can cause patterns is particularly obvious in self-organizing vegetation, such as fir wave and tiger bush. The pattern of vegetation and its changes also can cause processes to change. Pattern and process are the yin and yang of vegetation dynamics; each interlocks with and changes the other.
The overhead architecture of forest canopies has strong implications for the processes operating inside forests over time. Trees can grow large enough to alter their own local environment and that of any subordinate trees. How strongly this effect occurs depends on the species, shapes, and sizes of trees involved. The environmental condition on the forest floor profoundly influences the regeneration success of different species of trees.13
Feedback from the canopy tree to the local microenvironment and subsequently to seedling and sapling regeneration influences which tree will become the next canopy tree that dominates the site.14 These interactions drive the working of a forest and determine its nature when mature.
The environmental alteration of the forest floor caused by a canopy tree is most easily observed in terms of light.15 Walk through a forest on a sunny day. Shadows and sun flecks cover the forest floor; the light is not homogeneous owing to the plants overhead. Along with light, other potentially important tree-environment interactions take place. Plants too can alter their local environment with respect to soil moisture, nutrient availability, soil temperature, and the amount of rainfall coming through the forest canopy. One of the most readily observed effects on the recovery of the canopy gap is that caused by the death of a tree.
When small trees die, the surrounding trees grow lateral branches and the site is relatively unchanged. With the death of a somewhat larger tree, the saplings growing underneath the tree may increase in size, and one of them may increase enough to begin to dominate the site. Small-tree gaps may not allow enough sunlight to reach the forest floor to trigger germination of the seeds on the shaded forest floor. Depending on a variety of factors (the latitude, which determines the average overhead angle of the sun; the height of the canopy; and the size of the gap), some size of tree is sufficiently large that, when it dies, light and other environmental factors on the forest floor change radically. Such tree falls are gaps in the usual sense and promote a competition among seedlings and existing small trees to control the location. Very large tree falls (or multiple tree falls) often favor the germination success of trees that disperse well, need high levels of sunlight, and grow rapidly. Certain existing tree species require large and highly disturbed sites. Some of the more important commercial tree species, notably various species of pines (Pinus spp.), are in this category. The standard treatment for promoting the next generation is clearing large areas of land (“clear-cutting”) and sloughing or ripping the earth to expose mineral soil (called site preparation by foresters).
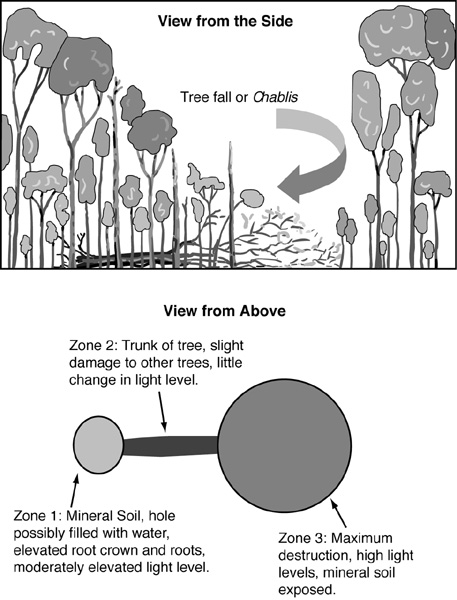
The fall of a tree and its attendant destruction have also been called the chablis, in part to sharpen the focus on the patterns of heterogeneity produced.16 “Chablis” actually comes from an old French word that means an opening in a forest. The variety of grape used to produce chablis wine may derive from the gap-colonizing habit of the original vines.17 The chablis has several important parts. The area beneath the crown of a fallen large tree undergoes a major change in light level at the forest floor. If the tree is uprooted, bare soil may also be exposed. The interwoven roots rip up a heap of soil as they are levered from the ground. A pit, which may be filled with standing water, replaces the space once filled by the heaved-over root ball (Figure 6).
In the area immediately adjacent to this root zone, the fall of the tree can have relatively little effect. Unless the fallen tree happens to strike another relatively large tree and topple it as well, the light levels at this adjacent site are relatively unchanged. However, the fallen tree lies on the ground and creates a microsite that is different from the rest of the forest floor. Some species of plants require elevated sites to regenerate. Farther away from the point of tree fall, in the area where the crown of the tree has landed, considerable destruction of the other trees living in that location may take place, as well as exposure of mineral soil and increased levels of sunlight. In this location, species that demand high light levels have an opportunity to grow into the canopy in the otherwise shady forest.
The death of a single tree kaleidoscopically produces a micromosaic on the forest floor, with the different pieces favorable to the regeneration of different species. Tropical rain forests, for example, may thereby maintain high species diversity. Long after a chablis event in rain forests (and in other forests as well), one can see the evidence of its occurrence in the patterns of the trees. Sometimes a straight row of trees of the same species, all about the same size, will be found. These are trees that regenerated on the trunk of a long-downed tree. Often they will be on “stilt” roots that grow on top of the downed tree trunk and send roots to the ground. When the tree trunk decomposes, the new trees are perched on the stilts that once wrapped around the downed log. Giant trees that are fast-growing, light-demanding species may emerge from the shady forest floor to occupy the canopy; these individuals are likely to have originated in the zone of destruction caused by the fall of a massive tree crown.
Just as the biology of the species determines regeneration success in gaps of various sizes and in different parts of the chablis, species attributes also determine the size and nature of gaps created by canopy trees. For example, the yellow poplar or tulip tree (Liriodendron tulip-ifera) of eastern North America almost always dies violently. One often finds blown-over trees with their leaves still fresh and green; almost never are dead Liriodendron trees found standing. Violent death arises from stress and old age weakening a tall tree’s root system so that it topples with a strong wind. Other species grow differently and respond to stress differently; they die slowly and leave the standing dead snags with flaking bark that provide food for the ivory-billed woodpecker.
Figure 6. A tree fall, or chablis. In a mature forest it produces a complex pattern of altered microenvironments. Thus, the death of a tree of one species leads to regeneration opportunities for several species with very different regeneration needs. From F. Hallé, R. A. A. Oldeman, and P. B. Tomlinson, Tropical Trees and Forests (Heidelberg: Springer-Verlag, 1978); and R. A. A. Oldeman, Forests: Elements of Silvology (Berlin: Springer-Verlag, 1991).
If mode of regeneration and mode of death are under biological control, then evolution might favor joining these features in a way that would promote species survival. An extreme example of such a conjunction is seen in the so-called suicide tree.18 Tachigalia versicolor is a very large, highly branched canopy tree species that is found in evergreen and partially deciduous lowland forest in Panama, parts of Costa Rica, and Colombia. A tree blooms only once in its lifetime and then dies within a year after releasing a wind-dispersed fruit. Several, but usually not all, of the trees in a given area bloom, pollinate one another’s flowers, and die at the same time. The Tachigalia seeds require a gap opening in the forest canopy for successful regeneration and subsequent growth and maturation. The death and fall of the parent produces a gap that greatly increases the opportunity for seedlings to grow to adulthood. Saplings of this species are often found growing in openings created by the fall of the dead adult Tachigalia that likely produced them.
What one sees in these suicide trees is a successful battle tactic of coupling the mode-of-birth and mode-of-death attributes of a tree. The violent death of the mother tree creates regeneration sites for its seedlings. The species creates in death the conditions needed for successful growth of the next generation. Tachagalia is an extreme case, but certainly other trees promote in the manner of their deaths the conditions needed for regeneration of their species.
Plants battling for ownership of space in the forest mosaic are locked in a complex battle full of enemies, allies, and intrigue. Thomas Hardy insightfully captured this dynamism in the following verse, excerpted from “In a Wood”:19
Heart-halt and spirit lame,
City-opprest,
Unto this wood I came as to a nest;
Dreaming that sylvan peace
offered the harrowed ease—
Nature a soft release from men’s unrest.
Great growths and small
Show them to be men akin—
Combatants all!
Sycamore shoulders oak,
Bines the slim sapling yoke,
Ivy-spun altars choke
elms stout and tall.
How do trees promote themselves and suppress their enemies? What are the broad strategies and the tactics of close infighting that are used by combatants in the war of trees?
A simple classification of the strategies can be developed from two questions: Does a tree species require a gap to regenerate? and Does a tree species generate a gap when it dies? The combinations of answers identify four broad strategies in a tree’s battle to dominate the forest canopy. For Tachigalia, the answers to both questions are affirmative, and the species is what is known as a Role-1 species of tree. The four roles are as follows:

These strategic roles of trees are intentionally simple; one could easily make the categorization more complicated. However, even these roles can help us to gain insight into the potential complexities of interactions in a forest.20
Successful species in different categories influence the other species on a small patch in different ways. When a large, mature, Role 1 tree dies, it would be expected to create a large gap that would encourage its own regeneration, as well as that of all the other Role 1 species. In addition, it would encourage the Role 3 species in the community that also need gaps to regenerate. Trees in the other categories play out their strategies to influence their own regeneration and that of other species. The interplay creates a complex web of species interactions (Figure 7) on the mosaic battlefields of the forest landscape. Even with just four simple strategic roles, a forest has a complexity of interactions comparable to the royal entourage in the court of a medieval Italian castle.
Figure 7. The complex relationships among trees of four simple ecological roles. Existing alongside the competition among individual trees are other interactions that could be classified as predation or mutualism.
The overall pattern of interactions among trees of different roles includes also a considerable mutualism (where two species reciprocally help each other) along with predatory interactions (where one species gains from another without reciprocation). For example, a Role 1 species creates gaps and has a positive effect on Role 3 species, which need these gaps. Role 3 species do not make gaps and thus have a negative effect on Role 1 species. This Role 1-Role 3 interaction is positive on the one side but negative on the other. In classic ecology, this asymmetry resembles the interaction between a prey and its predator, or a host and its parasite. The interactions among plants that involve withholding or losing space are as complex as the webs of interactions of competition and predation in animal communities.
Given this complexity, how should a forest change over time? It seems that this question has two answers, depending on whether we consider the changes on the scale of a gap or on the scale of the entire forest mosaic.
The answer is most straightforward on the gap scale. Considering the forest gap-replacement cycle, the total living mass of trees (biomass) at a location drops abruptly with the death of the dominant large tree. The biomass then slowly increases as the regenerating trees compete and the new winner emerges and fills the gap. Thus, a graph of biomass over the centuries for multiple cycles of gap generation and gap filling should have a distinctive “saw-toothed” shape. The teeth in this graph have sharp points at times when a large canopy tree has died. The distances between the teeth reflect how long trees live and how fast they grow to dominate a canopy gap. The saw-toothed curve is the pattern one would expect for the small area controlled by a canopy tree, an area perhaps 60 feet in diameter (about 1/40 ha).
What is the expected change over a large forest? A forest landscape’s biomass dynamic is the sum of the changes of all the pieces of the landscape mosaic. If a landscape were devegetated by a single event such as a clear-cutting or a wild fire, then all the small patches of the former forested landscape would be treeless. The living biomass of the entire landscape would be near zero. As trees began to grow on all the patches, the number of living plants on the landscape would increase. After the passage of time, all the patches of the landscape mosaic would be covered by large trees. With every patch of the forest occupied by a large tree, the living biomass would be at a maximum. However, this maximum would not be sustainable. Large mature trees eventually become senescent and die. With the death of large trees covering the landscape, the total amount of biomass on the landscape would drop. Of course, the geriatric trees would not all die during the same year. Growth in the canopy gaps would eventually average out the losses from death, and the amount of living biomass on the forest floor would be in a state of balance. The landscape would be covered with a mosaic of patches of all ages, representing all of the stages of the underlying gap dynamics cycle.
How does the mature forest (or “virgin forest” or “climax forest”) look? It should have patches with all stages of the gap-replacement cycle, and the proportions of each should reflect the duration of the different gap-replacement stages. The ecologists F. H. Bormann and G. E. Likens depicted a mature forest landscape as a dynamic mosaic of changing patches: “The structure of the ecosystem would range from openings to all degrees of stratification, with dead trees concentrated on the forest floor in areas of recent disturbance. The forest stand would be considered all-aged and would contain a representation of most species, including some early-successional species, on a continuing basis.”21 T. C. Whitmore expected this pattern and underlying process to work for all forests and asserted that the “forests of the world are fundamentally similar, despite great differences in structural complexity and floristic richness, because processes of forest succession and many of the autecological properties of tree species, worked out long ago in the north temperate region, are cosmopolitan. There is a basic similarity of patterns in space and time because the same processes are at work.”22
The processes to which Whitmore refers are gap-replacement processes—the processes that produce dynamic forest mosaics. Indeed, the occurrence of such patterns has been documented for several kinds of forests.23
There are some exceptions. The scale of the mosaics in many natural forests is somewhat larger than one would expect from gap filling of single-tree gaps, indicating the importance of other phenomena that cause multiple-tree replacements.24 In forests of very high latitudes, the sun is at an extremely low angle; a gap caused by a single tree is unlikely to create a canopy hole large enough to allow direct sunlight to hit the forest floor.25 Some of the relatively longer records of changes in forest structure and composition indicate that the forest composition fluctuates.26 Certain species seem to have periods of relatively weak recruitment of seedlings to replace large trees, but then undergo strong recruitment in other periods.27
The mature forests of the world are not collections of very large trees. Rather, they contain trees of all sizes in mosaic. They are not a primeval constant. They are dynamic, ever changing, and heterogeneous.
The sad demise of the ivory-billed woodpecker reveals much about the dynamics of natural vegetation. From the forest in which it occurred, the bird appears to have required a short-lived but slowly generated portion of the gap-replacement cycle. Its feeding habits required large, standing-dead trees with loose bark and insects underneath, a condition that is not always generated in the cyclic change in a forest; some trees, because of winds and other factors, fall over while still alive.28
The woodpecker “used up” its scant resource rapidly because of its habit of only removing the loose bark of the trees to search for insects. A large tract of mature forests would be required to supply enough forest mosaic elements to satisfy the bird’s needs. The clearing for agriculture of the floodplain forests of the U.S. South spelled trouble for a large animal that required a significant amount of mature forest to meet its highly specialized feeding requirements. And, certainly, the shooting of this rather tame bird by professional hunters was a contributing factor.