Vancouver, B.C., September 8.—All trains held up on the main line of the Canadian Pacific by slides are now on their way east and west. The line was finally cleared at eleven o’clock this morning. Yesterday the population of Field, the town nearest the slide, which occurred between Pallises and Glenogle, 25 miles west of Field, was increased by the addition of 2,000 passengers who were held up by the delay.
The slide, which was 300 feet wide and 30 feet deep, was caused by the bursting of an old beaver dam high up on the mountains. District Supt. MacKay, at Revelstoke, says that the dam burst under the pressure of the heavy rain storms last week. The slide carried the track away completely and it went clear across the Kicking Horse River, damming that stream and endangering the track above the slide. The river was completely blocked up, and it was found necessary to blast a new channel for the stream to release the pent-up waters which threatened to cause a washout further east.
The Canadian Pacific Railway had two steam shovels and a hundred men at work. Huge trees were brought down with the slide and boulders nearly as big as a box car made the job of clearing the track a difficult one. Some of the trees that came down bore the marks of the little animals’ teeth, and the supports of the dam erected by the beavers were plainly marked as such by the bleaching of their upper ends and the lower points coated with mud and slime.—News article in the Gazette (Montreal), September 9, 1913
Beavers (Castor canadensis) are the largest North American rodents. They attain lengths up to 4 feet (1.3 m) and can weigh more than 75 pounds (30 kg). Compact, rather thickset animals, they have small eyes, short legs, and broad, scaly, flattened tails (Figure 27). Their paddle tails and their large webbed feet are adaptations to aquatic life. Other aquatic adaptations include valves in their ears and nostrils that close when they are submerged. A clear membrane, called the nictitating membrane, covers the eyes when beavers are underwater. Beavers have dense fur: a fine gray underfur overlaid with coarse, darker brown guard hairs. The animals feed on a wide range of plant material, mainly the bark of wetland trees and shrubs.1
One of the beaver’s outstanding attributes is its capacity to construct complex civil engineering projects. These modifications of small streams can involve dams of mud and sticks to heights of 6 feet (2 m) or more and lengths of up to a third of a mile (500 m). Beavers sometimes construct canal systems for transporting small trees and cuttings. In many instances they build “lodges” in the lakes formed by their dams. A typical beaver lodge is made up of sticks in the middle of a pond formed by a beaver dam, or on the side of a stream with an associated burrow. Beavers also construct nest burrows in the sides of river or creek banks. The entrance to the lodge or nest is always from below the water’s surface.
Figure 27. The American beaver, Castor canadensis. From J. J. Audubon and J. Bachman, The Quadrupeds of North America, vol. 1. (New York: V. G. Audubon, 1854).
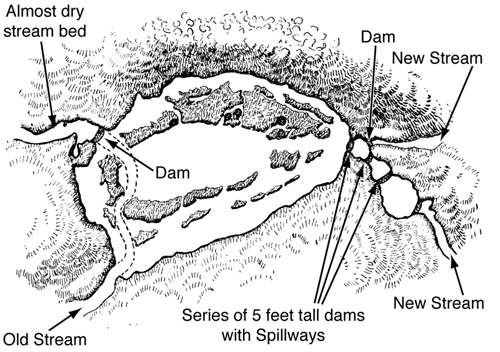
Figure 28. A beaver pond in North Ontario, Canada, with multiple dams and a stream rerouting. From A. R Dugmore, The Romance of the Beaver (Philadelphia: J. B. Lippincott, 1914).
Like the beaver dam that burst and stopped intercontinental train transport in the quotation from the Montreal Gazette, beaver projects can be impressive in their scale and complexity. One example (Figure 28) involved the complete rerouting of a stream, creating a pair of new dammed streams and a cascade of dams with spillways.
The beaver population of North America at the time of European arrival is estimated to have been somewhere between 60 million and 400 million animals.2 The species was found in virtually all aquatic habitats, particularly streams, from the north Mexican desert to the arctic tundra. The beavers’ geographic range encompassed about 6 million square miles (15 million km2).
The species was exploited for its fur from the early seventeenth century until the latter part of the nineteenth century. As beavers fell victim to fashion in the East and were eliminated, explorers—followed by rough-and-tumble frontiersmen, mountain men, and voyageurs— pushed into the western -wilderness to find more beaver skins.3 By 1900 the beaver was almost extinct in North America.4 Today a managed population continues to recover from historic overexploitation for fur, but the species probably still numbers less than 5 percent of its former density.5
Trapping North American beavers for their fur began with the Italian explorer Giovanni Caboto (John Cabot), sailing for the Merchants of Bristol in 1497. Among other findings he discovered the Grand Banks, an extensive set of shallows off the coast of Newfoundland, Canada, where codfish were so numerous that they could be brought up with baskets. Caboto’s discovery made relatively little impression on his employers in beef- and mutton-eating Protestant England, but Catholic Europeans found it to be of considerable interest. Six hundred ships were working the Grand Banks by 1603 to produce salted and dried fish for shipment to Europe. Negotiations with several of the tribes of Amerindians on shore involved gifts to ensure that the fish drying racks (an obvious source of easy-to-get firewood) would be left for the next fishing season. These initial commercial contacts eventually propelled an expanding trade in furs, including beaver pelts. Subsequently, Europeans formalized trading relations with a diversity of native people, based on exchange of furs for a wide range of goods. A trade network ultimately stretched across the North American continent.6
New England is a microcosm of the expansion of fur trading in North America. Prior to European settlement, beavers were found in almost every body of water.7 Between 1620 and 1630, an average of 10,000 beaver pelts per year were taken in the fur trade in Connecticut and Massachusetts.8 Indeed, between 1631 and 1636, returns from the trade in beaver pelts amounting to £10,000 paid the debt of the fledgling Plymouth Colony. In New Netherlands (the New Amsterdam colony and its outreaches), the Dutch West India Company’s exports progressed from 400 beaver skins in 1624 to almost 15,000 beaver pelts annually by 1637.9 A century and a half later, by 1764, very few beavers were being taken in Massachusetts (or elsewhere in New England).10 As the species was locally wiped out by overtrapping, the westward expansion of trade and trapping commenced.
In its heyday as a hat-making material, beaver fur had been converted to a fine gray felt. The demand was fueled by European male fashion. The exploitation of beaver for hat-felt spread across North America at an ever-escalating pace. Then, in 1840, a hatter’s apprentice in London discovered that chopped silk fibers could be used to produce felt hats with a greater luster than those made of beaver felt.11 The silk top hat became the mark of a well-dressed gentleman, and the price of beaver fur collapsed by 30 percent or 40 percent over the next fifty years.12
Even with the decrease in the demand for beaver pelts caused by the switch in European hat styles, beaver continued to be exploited as a fur animal. The Hudson Bay Company sold 3 million beaver skins from 1853 to 1877.13 By the middle of the nineteenth century, the combination of hunting, trapping, and conversion of wetland habitat had taken the beaver populations down to a small percentage of their former abundance. The species seemed destined for extinction.
Today, many of the natural predators of the beaver (notably the gray wolf, Canis lupus) have been reduced or eliminated, and laws regulate the trapping of the species. Beaver populations are expanding and the population of North America is thought to be on the order of 6 million to 12 million individuals, still a small fraction of the former abundance.14 The loss of such a marvelous creature as a consequence of what in retrospect seems an almost trivial fashion has a darkly ironic aspect.
In North America the local demise of the beaver was followed by the clearing of land and agriculture settlement. Exploration and subsequent exploitation of natural resources often tended to move upriver. Tributaries of a river could feature the elimination of beaver on the upper reaches of a drainage system, while clearing and farming were altering the surrounds of the lower reaches.
In historical accounts, eastern North American waters are quite different from their contemporary condition. Great rivers were clear, with numerous woody snags, complex backwaters, and forested flood-plains.15 The bottom of coastal bays and lagoons was visible. For example, the Chesapeake Bay was transparent and full of eelgrass beds, along with extensive shell reefs with oysters of remarkable size. Today the eelgrass is gone and the muddy bay has a silt bottom that is a difficult substrate upon which to establish oysters.16 These changes appear to be strongly related to the increased sediment that was fed into the bay from poor agricultural practice and subsequent soil erosion.
Sedimentation rates in the Chesapeake before European settlement were on the order of 1 inch every thirty years (0.09 cm/yr). This rate doubled when the surrounding land was about 20 percent cleared and doubled again when the land clearing reached 50 percent.17 But was the siltation due to other causes, perhaps overexploitation of the Chesapeake Bay’s extensive oyster reefs that filtered and refiltered the Chesapeake’s water?18 Could it have been caused by the lack of clean source waters resulting from the loss of beavers and their dams? Perhaps all of these questions can be answered in the affirmative. The crux is really the degree of influence of each of these factors.
The presence of beavers and their projects strongly affects the streams that feed rivers. First-order streams are the smallest ones in a drainage system. A second-order stream is formed when two first-order streams join, a third-order stream is created by the joining of two second-order streams, and so on. With occasional catastrophes such as the one described at the start of this chapter, beavers are able to construct dams on streams up to fourth order. Streams any larger have floodwaters strong enough to regularly wipe out the beavers’ construction projects.
The nature of smaller streams is substantially altered by the presence of beavers. The ponds caused by their damming of small streams act as settling basins for silt and sediment. The emergent vegetation in the shallows of the ponds ties up nutrients in the water and serves to reduce what in present times is called stream eutrophication—the overenrichment of the waters. The changes caused by beaver ponds are eventually transmitted downstream to the higher-order reaches of rivers. We have seen that beaver projects have the ability to modify water courses and water quality. The filling of beaver ponds builds rich moist meadows, which go through ecological succession to produce diverse plant and animal communities.19
Primarily, the beaver demonstrates the key role of animals in the functioning of landscapes. This short chapter provides a transition between the animal stories that have come before, which are largely involved with the ecological concepts in “natural systems,” and the stories that follow, which add the human component of landscape change.
The beaver is an animal agent of landscape change. Its story establishes the need for careful stewardship of the species and of similar creatures that have the capacity to alter the systems in which they occur. Indeed, this point has been elaborated with a rich panoply of studies of the role of individual animals in ecosystems around the world. Discussions of the effects of animals on ecological systems not only fill books, they fill library shelves and sections.
Animals pollinate, bioturbate, predate, and aerate; they ingest, digest, egest, and divest vegetation of seeds, buds, bark, and roots. They are a small part of the living biomass (surely for terrestrial ecosystems), but they can have profound effects on the structure and functioning of the ecosystems in which they occur. Charles Darwin, after traveling the world and developing the theory of evolution, settled into the important problem of understanding how the actions of earthworms modify the soil.20 Thousands of other biologists and ecologists have dedicated their lifetimes to deriving similar insight into the effects of other creatures on ecological systems.
What emerges from this rich body of work is a “law of unintended consequences.” We can find numerous examples of ecological systems that appear susceptible to change. The elimination (or conversely, the introduction) of what might seem a minor species can potentially produce profound and unexpected results in natural landscapes, sometimes over large areas. Some of these unintended consequences have had huge economic impact and have altered human history.
Some ecosystems are changed greatly by the elimination (or addition) of a single species. Other ecosystems are extremely robust with respect to some of their features, even in the face of remarkable levels of species extinction. The deciduous forests of the eastern United States, Europe, and eastern China are very similar in terms of their physical appearance and in the taxonomy of the component species. Further, the interaction between a forest ecosystem and the underlying geology produces soils with similar physical and chemical properties in all three continents. Yet the number of species and genera in Europe is considerably less than in North America, which is still less than in the comparable forests of China.
The European diversity, once much greater, has been regularly eroded by extinctions resulting from the periodic glaciations in the Pleistocene.21 Over the past 2 million years or so, as glaciers have proceeded and receded across Europe, many species and genera of major tree species have been lost. For the region of Europe that is now The Netherlands, the number of tree genera has dropped from about forty-two to around fourteen today. Some of these “lost” genera are not only gone from The Netherlands, but from all the rest of Europe as well. Species of these same genera are still important trees in North America and Asia. How has the loss of such a substantial section of tree diversity from the forest vegetation of Europe changed the way these ecosystems function? The answer is far from obvious. Yet European data on nutrient cycling, productivity, and water use by forests apply to forests in North America, and vice versa.
The differences in biotic diversity among the three major temperate deciduous forests (China, North America, and Europe) are substantial. Species or genera with seemingly unique and important ecological attributes may be missing in one of these deciduous forests but present in the others. A similar observation can be made for the tropical rain forests of Africa: they are floristically less diverse than those of South America, which in turn have lower diversity than the Southeast Asian rain forests.22 We are only beginning to understand the consequences of such differences. Perhaps because the period of scientific data collection on forests has been short relative to the slow response of forests, there is a great deal more that we need to know.
Certainly we have seen instances where the elimination of a key species, or the introduction of a new species, has resulted in large and in some cases surprising changes in natural ecosystems. We also know of instances where changes in ecosystems follow the loss of an important species or group of species that carry out some essential function in an ecological system. For purposes of illustration, here are a few examples.
• Pollinators: The tremendous explosion in the diversity of flowering plant species some 100 million years ago was probably caused by the interactions between plants and pollinating animals.23 Some plants are highly dependent on specific pollinators and compete for access to them. The adaptations to secure pollinators with a high fidelity to the flowers of a particular species can be marvelously complex: differences in the shape, size, and position of the flowers; timing of the flowering; and a variety of chemical cues and attractants. One of the most surprising adaptations is the coevolution of biochemical poisons in the nectar of flowers that can only be tolerated by the specific pollinator of that plant.24 In the most highly developed plant-pollinator coadaptations, if one of the partners is lost, then so is the other. Similar adaptations associated with animals in the dispersal of seeds are also often highly co-evolved and represent an equivalent case of strong interdependency between species.
• Keystone species: The sensitivity of highly coevolved pollination or seed-dispersal systems to the extinction of one of the partners represents a particular case of a larger category called keystone species. By analogy to the keystone that holds together the stones in an arch, these are species that hold together the interactions among a group of species.
For example, in the central wheat belt region of Western Australia a collection of nonmigratory birds called honeyeaters (family Meliphagidae) live from flower nectar and pollinate an array of attractively flowered shrubs and small trees in the Proteaceae family. The honeyeaters are attractive, mostly small birds; the plants that they pollinate are to some degree adapted to them.
And many of the region’s plants are dependent on the honeyeaters for pollination.25 The different flowers are of sizes and shapes to accommodate honeyeaters of appropriate sizes and with particular bill lengths. The plants flower at different times, a phenomenon associated with timing reproductions so as not to compete with other flowers for the services of an appropriately-sized honeyeater. However, during one short time of year only one species of Banksia (B. prionotes) blooms, and it sustains all of the honeyeaters through a critical period. This Banksia species is widespread, but it is relatively uncommon and quite locally distributed in the central wheat belt region. If this one plant were somehow eliminated, the honeyeaters and all of the species they pollinate could disappear from the region.26 Thus, Banksia is a keystone species.
Or a keystone species could be a predator that controls a pest which might otherwise destroy the vegetation; it could be a pest or disease that keeps the density of a predator from becoming too great; it could be a necessary food plant. If keystone species are a common part of the fabric of ecological communities, then they are agents for surprise in management of ecological systems.
• Pests and diseases: Probably the most easily documented large changes in ecological systems from a single organism result from pests and diseases. Animal pests can be seen as exercising strong control over the success of plants and other animals; they can control the abundance or even the presence of dominant species of plants and animals and thereby change the nature of the ecosystems. A fungus parasite, the chestnut blight (Endothia parasitica), was introduced into New York on imported Asiatic chestnut nursery stock.27 Within thirty years of its discovery (in 1904), this fungus had destroyed virtually all of the American chestnut (Castanea dentata) timber in the eastern United States.
The paradox that ecosystems are observably resistant to elimination of species in some cases and highly sensitive to seemingly analogous species deletions in others is a consequence of internal feedbacks. Understanding the basis of animal interactions motivated Charles Elton to introduce an ecological niche concept that emphasizes the way species feed on one another. This concept increasingly stressed the competitive interaction of animals to generate patterns of abundance. Some ecological systems seem to be composed of highly interactive species populations whose interactions can be seen as generators of patterns. This feature seems not to occur in other ecosystems. The interactions among components produce a spectrum of regulatory internal feedbacks, which range from being system altering to being inconsequential. Sadly, we are not exceptionally successful at differentiating major from minor interactions a priori—at least not at present. We do understand ecosystem feedbacks in theory, and we are developing a portfolio of what has happened in the past when environmental change has yielded large ecosystem consequences.
We need to make significantly more discoveries before we can confidently predict the results of eliminating species from ecosystems. Nonetheless, it is clear that plants and animals are not passive players. Large animals can destroy parts of the systems in which they occur, and creatures such as the beaver construct visible changes to the landscape structure. It is certainly no less true of the smaller creatures that make subtle alterations in the fabric of ecosystem interactions. Change in the vectoring of disease, and in the activity of animals as parasites and pests, can profoundly alter the nature of ecosystems. The fungal and bacterial components of ecological systems dominate ecosystems as diseases, processors of material, and transformers of the physical and chemical structure of the soil. In a time of species extinction, the effect on ecosystem functioning of change in the biota must be better understood.