CHAPTER 2
MEASUREMENT METHODS OF MUSCLE FATIGUE
Nicola A.Maffiuletti and David Bendahan
OBJECTIVES
The main objectives of this chapter are to describe:
• the different methods allowing quantification of muscle fatigue, which include the tools and the procedures for the assessment of force-time and power-time curves (part I);
• the techniques and the procedures for the assessment of neuromuscular mechanisms (part II); and
• the metabolic (part III) mechanisms underlying muscle fatigue. The advantages and drawbacks of the different tools and techniques will be briefly discussed.
INTRODUCTION
Muscle fatigue (hereafter referred to as fatigue) could be defined as an exercise-induced reduction in muscle performance. Two different measurement models are generally used to quantify such impairment. The first consists of quantifying the reduction of power output during real exercise such as cycling or running (generally at a maximal intensity), where dynamic performance decline could be attributed to reduced force and/or velocity. The second model is based on the assessment of maximal isometric muscle force-generating capacity before and immediately after real or simulated exercise, to describe the decline in static muscle performance. Therefore, measurement methods of fatigue mainly consist of maximal force or power recordings obtained before, during or after exercise.
Because fatigue is a decline in muscle force or power induced by exercise, all fatigue models (Cairns et al., 2005) entail the two following components: fatigue induction and fatigue quantification (Table 2.1). Accordingly, the two important factors that should be determined when designing a fatigue study are the type of exercise inducing fatigue and the measurement methods of fatigue, which include type and timing of the measurements. Depending on the study population and research question, investigation of the neuromuscular and metabolic mechanisms underlying fatigue (i.e. fatigue mechanisms) could also be considered (Table 2.1).
Fatigue is an important functional parameter for physical work and daily activities, and its assessment is relevant to practitioners in sport, occupational, geriatric and orthopaedic medicine. In all these settings, it is essential to respect the specificity between the activity of interest and the type of exercise inducing fatigue. The best option is represented by fatigue quantification during real exercise in specific conditions (e.g. cycling power loss for a road cyclist). However, it is quite challenging to assess accurately muscle force/power during the majority of sport, work and daily conditions, because of the acyclic nature of these activities. There is indeed the
necessity to select/conceive a fatiguing simulated exercise, which would mimic the type of muscle actions realised during real exercise (Table 2.2). It is worth noting that such specificity should also be respected when determining the characteristics of muscle actions for fatigue quantification (cf. next section).
In this chapter, we will enumerate the different methods allowing quantification of muscle fatigue, which mainly include the tools for the assessment of force-time and power-time curves (part I) and the techniques for the evaluation of neuromuscular (part II) and metabolic (part III) mechanisms of fatigue.
MEASUREMENT METHODS OF MUSCLE FATIGUE
Part I: tools and procedures for force/power measurement
In this section, we will present the methodology of performance decline assessment consecutive to real or simulated exercise, which includes the tools, test procedures, outcomes and some considerations (advantages, drawbacks) for different physical tasks. The choice of an appropriate fatigue outcome and of the timing of measurements should be made in relation to the research question. As previously outlined, two models are generally used to assess exercise-induced performance decline: mechanical power loss during dynamic exercise versus isometric force loss as a before-to-after exercise ratio. Therefore, these two modalities differ in the main outcome measure (power versus force) and in the number and timing of repeated measurements (several, during exercise versus two, before and after exercise).
Mechanical power decline can be quantified during whole-, lower- or upper-body short maximal exercise where dynamic contractions are performed by several muscles, mainly in closed-chain conditions. Maximal power is attained within a few seconds and a gradual fall is subsequently observed. These muscle actions are very similar to those realised during sport, work or daily activities. On the other hand, isometric open-chain testing (e.g. seated knee extension) has been, for technical reasons (standard conditions, high reliability), the most commonly used method to assess force decline (or torque, hereafter referred to as force) of single-muscle groups. However, isometric force measures underestimate significantly functional impairment, i.e. the decline in peak muscle power is higher than maximal isometric force, so that the former is considered to be more appropriate to assess performance in dynamic exercise. Power assessment may also give more information compared to isometric force because significant changes in the metabolic processes involved in energy release and utilisation are more easily detected by muscle shortening (Vøllestad, 1997).
Friction-braked (Monark), air-braked (Repco, Kingcycle), electromagnetically braked (Lode) and special constructed constant-velocity (McCartney et al., 1983) ergometers, as well as mobile systems (SRM, Powertap) mounted on the drive train of the cyclist’s own bike can be used to measure cycling power against a resistance. Systematic and random errors associated to power measurements provided by these different ergometers have been reviewed by Paton and Hopkins (2001). Typically, temporal changes in maximal or mean power output are examined through one or several repeated short-term all-out exercise bouts, where bout number and duration can be modulated according to anaerobic pathways contribution. The 30-s all-out Wingate test or repeated-sprint exercise (RSE) tests (e.g. five 6s maximal sprints every 30s, Bishop et al., 2001) are some examples of fatigue assessment during cycling exercise. Concerning the outcome measures, it has been demonstrated that (i) power output is more reliably assessed with measures of mean power as opposed to peak power during RSE (Glaister et al., 2004); and (ii) although fatigue measures have lower reliability than absolute power output, the per cent decrement recorded over each effort is the more reliable technique (Fitzsimons et al., 1993). Laboratory fatigue tests on cycle ergometers are largely used because of the possibility to combine cardiorespiratory, blood pressure or blood lactate assessments. Despite these advantages, cycle ergometry is not an exercise mode specific to sports involving repeated sprints.
Non-motorised treadmills (Woodway) instrumented with a force transducer (strain gauge load cell) have therefore been developed (Lakomy, 1984), which offer the possibility of testing RSE using running as the exercise model (ecological validity). A non-elastic belt is used to tether the wall-mounted height-adjustable strain gauge to the subject’s body via a harness. Treadmill speed is generally quantified using spring-loaded generators or reflective optical switches. Therefore, horizontal power can be calculated as the product of force and speed for average and instantaneous data. In these conditions, it has been demonstrated that measures of speed are more reliable than measures of power and force, and the use of average data further increases the reliability compared to instantaneous values (Tong et al., 2001). However, reliability of fatigue indices for RSE on non-motorised treadmills is still poor (Hughes et al., 2006).
Alternatively, the decline in mechanical power output with fatigue can also be quantified during daily activities, such as walking and stair climbing. Funato et al. (2001) recently described a self-driven instrumented treadmill which allows recording of both horizontal and vertical ground reaction forces, as well as horizontal pushing force on a handlebar during walking and running. Contrary to this high-tech approach, Margaria et al. (1966) proposed a formula to estimate mechanical power during stair climbing, which is simply based on the measurement of the time necessary to cover a given vertical distance at maximal velocity, two steps at a time, where:
 |
|
Unfortunately, there is a lack of information on the decline in walking and stair-climbing power in elderly and patient populations during daily life and as a result of a treatment or surgical intervention, which could be extremely relevant to geriatric, neurological and orthopaedic medicine.
Coming back to fatigue assessments for athletic populations, sport-specific ergometers such as wind-braked (Concept2), friction-braked (Gjessing) or air-braked (Rowperfect) rowing ergometers can be used, which generally provide online feedback about stroke parameters such as speed, pace and power. The most commonly adopted ergometer is probably the Concept2. Despite high test-retest reliability of rowing performance on this ergometer, power values provided by the machine are significantly underestimated (~7%) compared to the actual power produced by the rower (Boyas et al., 2006). However, rowing ergometers are rarely used to evaluate fatigue during very short all-out exercise, but rather, they are used to simulate rowing competitions (mainly 2,000m), where pacing strategy invalidates fatigue assessment.
Vertical jump power can be directly measured using fixed or portable force plates (Kistler, Bertec, AMTI), while it can also be estimated from flight time recordings (Bosco et al., 1983) provided by contact mats (Ergojump) or photoelectric cells (Optojump) on the field. Typically, mechanical power associated with single jumps is recorded during a series of consecutive counter-movement jumps (15–60s, at a frequency of about 1Hz), in an attempt to mimic sport-specific fatigue (e.g. alpine skiing). The power decline is subsequently expressed as a percentage of the higher of the first 3–5 jumps to the last 3–5 jumps. Another estimate of muscle fatigue is the number of jumps in which power declines to 25–50% of the initial power. The main limitations of these types of fatigue tests are the control of posture, particularly in the fatigued state, which is necessary for the validity of jumping power assessments, and the maximality of jumps throughout the entire test duration, since some subjects may pace themselves to some extent during the test. Portable accelerometry-based systems are now available (Myotest), which could provide jumping power under field conditions; however, their validity and reliability remain to be ascertained.
In addition to the valid measurement of muscle torque, work or power, commercially available isokinetic dynamometers (Biodex, Humac Norm, Con-Trex) offer the possibility to test muscle fatigue (or “endurance”, which is basically the opposite of fatigue), mainly in open-chain but also in closed-chain conditions. Isokinetic fatigue is often quantified as the percentage decline in gravity-corrected peak torque or average work (which additionally takes into account the range of motion) over a certain number of repetitions (usually 20–50), completed at a relatively fast concentric velocity (usually 180°·s−1). Thorstensson and Karlsson (1976) were the first to define a fatigue index as the torque of the three last contractions as a percentage of the initial three contractions out of 50 contractions. They also demonstrated a significant correlation between the fatigue index and the percentage of fast fibres in the vastus lateralis muscle of ten subjects. Recently, we (Maffiuletti et al., 2007) and others (Pincivero et al., 2001) have provided experimental evidence that the linear slope of the decline in torque or work output over the series of fatiguing contractions is a better indicator than the fatigue index in terms of test-retest reliability. However, reliability of isokinetic fatigue indexes is inferior to that of the absolute parameters (Maffiuletti et al., 2007), similar to cycling and running ergometry. Therefore we recommend interpretation of all these fatigue outcome measures with caution. It should also be remembered that the use of isokinetic actions for fatigue assessment is questionable because they do not exist in normal movements.
Finally, strength training machines equipped with linear encoders, accelerometers and force transducers allow force and power quantification during static or dynamic actions in both open-chain and closed-chain conditions. In isometric conditions, the force transducer (generally an S-shaped strain gauge load cell) should be fixed so as to record the direct line of force. In dynamic conditions, some commercially available machines (Technogym) are equipped with encoders measuring linear displacement, which in turn allows calculation of work and power (Bosco et al., 1995). Alternatively, linear encoder systems (MuscleLab) or accelerometers (Myotest) can be applied to standard machines and even to barbells or free loads. In isometric conditions, the decline in isometric MVC force is very often considered as the main measure of interest (see next section), while fatigue-induced alterations in rate of force development could also be examined. In dynamic conditions, the decline in concentric power during a typical strength training session, both within the same set and within different sets, has become a routine measure in the control of training for high-level and recreational sportsmen.
Part II: techniques and procedures to evaluate neuromuscular mechanisms of fatigue
In this section we will present how surface EMG and muscle force recordings obtained before and immediately after exercise can be used to investigate central and peripheral mechanisms of fatigue. The classical approach consists of realising a series of voluntary and stimulated contractions of the agonist and antagonist muscles, with concomitant EMG and force recordings. It is essential that this series of measurements is completed immediately after (within 1–2 min) the end of the fatiguing exercise and its total duration does not exceed 5–10min. Measurements can also be performed during exercise using short interruptions to investigate the time course of alterations, and/or hours or days after the fatiguing bout, to examine the recovery process (particularly for muscle damage studies). Such neuromuscular assessments provide some evidence of the central and peripheral alterations induced by exercise, according to their localisation within the neuromuscular system. Central fatigue is defined as a progressive reduction of voluntary activation of muscle during exercise (activation failure) that could be due to supraspinal and/or spinal mechanisms (Gandevia, 2001). On the other hand, peripheral fatigue is produced by changes at or distal to the neuromuscular junction, and could include transmission and/or contractile failure (excitation-contraction coupling failure). It should, however, be remembered that contrary to animal studies, these in vivo assessments do not allow isolation of single processes of fatigue.
Force recordings
Isometric muscle force, as assessed using force transducers or isokinetic devices, is the most widely employed parameter for muscle fatigue studies. MVC force has been defined as the most direct assessment of fatigue and the first choice method, because the output is an integrated result of the total chain of neuromuscular events (Vøllestad, 1997). Very short (3–5s) actions are generally used to assess this variable, where MVC force is attained 400–600 ms after contraction onset and a plateau is seen in the subsequent period. Moreover, electrical or magnetic stimuli adequately superimposed to the MVC plateau allow investigation of the maximal evocable force (Table 2.3), and in turn of muscle activation failure (see below). It should, however, be pointed out that, although isometric MVC force assessment is highly reliable, easily administered, requires little skill involvement and is relatively inexpensive (Wilson and Murphy, 1996), several experimental recommendations should be observed to obtain real MVC and muscle activation scores, more particularly in the fatigued state. These criteria, which have been described in detail elsewhere (Wilson and Murphy, 1996; Gandevia, 2001) include: appropriate familiarisation and practice; visual feedback of performance with variations in gain; standardised verbal encouragement with clear instructions (hard versus hard and fast); several repeat trials; negligible pre-tension prior to the test; and eventually provision of rewards.
Muscle activation failure can be measured using twitch interpolation during an MVC, where one (or more) supramaximal stimulus is delivered to the motor axons innervating the muscle and voluntary versus evoked force outputs are compared. Supramaximality of stimulation should be ensured by using an intensity consistently higher (20–50%) than that eliciting the maximal M wave (i.e. compound muscle action potential) and peak twitch. Any additional force produced by the stimulation indicates incomplete motor unit recruitment and/or suboptimal firing frequency of active units. Two formulas are classically used to estimate the extent of activation: voluntary activation, where the size of interpolated twitch is divided by the
size of a control twitch produced by identical stimulation in a relaxed potentiated state (Thomas et al., 1989), i.e. voluntary activation (%)=[1−(interpolated twitch/control twitch)]×100, and central activation ratio, which consists of a simple MVC-to-maximal evocable force ratio (Kent-Braun and LeBlanc, 1996), i.e. central activation ratio=MVC/ (MVC+interpolated twitch). Importantly, voluntary activation has been shown to be a more sensitive (Bilodeau, 2006) and valid (Place et al., 2007) index of central fatigue than the central activation ratio. However, limitations in the twitch interpolation technique have been identified (Gandevia, 2001), which also include the contribution of intramuscular processes to superimposed force with fatigue (Place et al., 2008).
On the other hand, force evoked by single, paired or trains of stimuli under resting conditions can be used to examine contractile impairments (Table 2.3) and, together with the analysis of the M wave recorded by surface EMG (see below), allows excitation-contraction coupling failure to be approached in vivo (Desmedt and Hainaut, 1968). For example, M wave amplitude and twitch force both decrease with the occurrence of transmission failure. With contractile failure however, no changes are observed in M wave amplitude, while twitch force is significantly decreased, i.e. failure of excitation-contraction coupling. Additionally, fatigue-related changes in twitch-time course parameters such as increased time to peak twitch (contraction time) and half relaxation time (Duchateau and Hainaut, 1985) could be compared to M wave changes to gain insight into excitation-contraction coupling impairments.
Alternative approaches to the study of excitation-contraction coupling alterations with fatigue are represented by the analysis of low-to-high frequency force ratio (Edwards et al., 1977) (see the section below, “Electrical or magnetic stimulation?”) and muscle twitch post-activation potentiation (Miller et al., 1987). This latter refers to any enhanced contractile response which results from prior contractile activity and is generally computed as the ratio between the size of the twitch recorded before and immediately after an MVC. Since fatigue can coexist and therefore be masked by potentiation, we (Place et al., 2007) and others (Kufel et al., 2002) recently suggested that potentiated twitch or doublets are superior to nonpotentiated responses for detecting contractile fatigue. We would also like to remind that unpotentiated twitch force is an inappropriate measure of peripheral fatigue (Edwards et al., 1977; Millet and Lepers, 2004).
EMG recordings
Electromyography (EMG) is the recording of electrical signals that are sent from motoneurons to muscle fibres (action potentials) while they propagate along the sarcolemma, from the neuromuscular junction to the extremities of the muscle fibres (Enoka, 2002). The classical configuration for surface EMG of a whole muscle consists of using two small electrodes positioned between the innervation zone and the tendinous insertion (bipolar), while single electrode (monopolar) or arrays and even grids of electrodes (multipolar) can be used (see www.seniam.org for recommendations about electrode placement and location).
Surface EMG reflects both central and peripheral neuromuscular properties, since its main characteristics, such as amplitude and power spectrum, depend on muscle fibre membrane properties and on the timing of motor unit action potentials (Farina et al., 2004). Researchers should be aware of the physiological and nonphysiological factors influencing surface EMG (Farina et al., 2004), as well as of the real information extracted from this signal. For example, global surface EMG underestimates motor unit activity due to overlapping of positive and negative phases of motor unit potentials (i.e. amplitude cancellation) (Keenan et al., 2005). It should also be considered that the difficulties in interpretation of isometric EMG amplitude are further amplified in the case of dynamic contractions (Farina, 2006) and spectral estimations such as mean and median frequency.
Although less used than the twitch interpolation, EMG amplitude obtained during an MVC (Table 2.4) is another technique to detect central alterations due to fatigue, which, contrary to twitch interpolation, allows changes in descending drive between the synergists of a muscle group to be distinguished (e.g. elbow flexors, knee extensors). In order to enable peripheral changes to be excluded from the interpretation of the data and therefore to provide an index of central activation failure, average rectified or root mean square EMG could be expressed relative to the size or surface of the M wave obtained with supramaximal nerve stimulation. However, due to the limitations outlined above, caution should be taken in using this latter technique for central fatigue assessment. The gold standard for activation failure is probably the comparison of the decline in MVC force with
the decline in maximal tetanic force (Merton, 1954; Bigland-Ritchie et al., 1978), even if it is not feasible in the large majority of anaesthetised human subjects.
On the other hand, compound action potentials can be evoked using mechanical, electrical or magnetic stimuli (Table 2.4) and their main characteristics (peak-to-peak amplitude and duration, area, latency) can be compared before and after fatigue. The maximal M wave is evoked by the recruitment of all motor axons and therefore provides an estimate of the response given by the whole motoneuron pool. Its amplitude is a measure of transmission across the neuromuscular junction and muscle membrane excitability. Maximal M wave amplitude is also used as a normalisation standard for all evoked potentials, provided the former is obtained in the same conditions (muscle action and muscle length) as the latter. Stimulated potentials include motor-evoked potentials and Hoffmann (H) reflexes. Fatigue-induced changes in motor-evoked potential amplitude and silent period duration, which are obtained using transcranial magnetic stimulation (see below), allow investigation of corticospinal excitability alterations. H reflex, evoked by selective (submaximal) stimulation of la afferents contained in the corresponding mixed nerve, can be potentiated (Trimble and Harp, 1998) or depressed (Racinais et al., 2007) following exercise, indicating altered motoneuron excitability and/or presynaptic inhibition of la afferents. Finally, spinal reflexes can also be evoked as a result of transient stretches, which can be produced in a reliable way using electromagnetic hammers or special ergometers (Avela et al., 1999) to obtain, respectively, the tendon (T) reflex and the stretch reflex. Compared to the already challenging H reflex, the interpretation of these reflex responses is further complicated by the influence of γ fusimotor drive, which controls the sensitivity to stretch of muscle spindle primary endings (Pierrot-Deseilligny and Mazevet, 2000).
Electric or magnetic stimulation?
As discussed above, non-invasive electrical and magnetic stimuli can be applied at different levels of the neuraxis (from motor cortex to peripheral nerve trunk) or over the muscle, both at rest and during voluntary contractions (see Table 2.1), to investigate central and/or peripheral mechanisms of fatigue through the analysis of evoked EMG and force signals. Electrical stimulation, in which current is transmitted to a body organ through surface electrodes of different sizes and shapes, has been used in clinical and research settings for several decades. This modality is very effective for stimulation of superficial peripheral nerves. However, deep nerves cannot be easily excited because of the high resistance of some tissues (e.g. bones) and because of the discomfort associated with strong stimulations. Magnetic stimulators were therefore developed in the 1980s (Barker et al., 1985) for painless cortical stimulation, which is in contrast to the sensations felt using electrical stimulation with electrodes positioned on the scalp. Since its introduction, transcranial magnetic nerve stimulation has been in widespread use for the investigation of motor-evoked potential characteristics and silent period duration, and more recently, also for the assessment of voluntary activation of upper limb muscles (Todd et al., 2003). Interestingly, magnetic stimulation of peripheral nerves (e.g. phrenic, femoral) has also been shown to provide valid and reliable assessment of muscle contractile function, including fatigue (Polkey et al., 1996; Vergès et al., 2009) and voluntary activation level (O’Brien et al., 2008), compared to classical electrical stimulation. Although magnetic stimulation presents some advantages with respect to electrical stimulation (e.g. lower discomfort, possibility to excite deep nerves), it should be remembered that magnetic stimulators are quite expensive and cannot deliver trains of stimuli at high intensity and high frequency, such as those generated by commercially available electrical stimulators.
Supramaximal trains are indeed required for investigating exercise-induced changes in the force-frequency relation, more particularly in the low-to-high frequency-force ratio (generally 10–20 Hz to 50–100 Hz) (Edwards et al., 1977). Any increase in this ratio following exercise is commonly associated with long-lasting, low-frequency fatigue, which may be caused by excitation-contraction coupling impairment. On the other hand, a decrease of the low-to-high frequency ratio is associated with high-frequency fatigue, which is thought to reflect impairment of muscle excitation at the level of the neuromuscular junction. However, tetanic electrical stimulation of intact muscles is painful and unsuitable in clinical practice. Therefore, we recently proposed to use force traces of paired stimuli at both 10Hz and 100Hz as a surrogate for stimulation trains for the assessment of low- and high-frequency fatigue (Vergès et al., 2009).
Surface electrical stimulation can be differentiated in nerve versus muscle stimulation according to the position and size of the stimulating electrodes. In the first case, small electrodes are positioned over the peripheral nerve trunk, at a site where it runs close to the skin (e.g. poplitea fossa for the tibial nerve, femoral triangle for the femoral nerve). Over-the-muscle electrical stimulation is performed with large electrodes placed in proximity to the muscle motor point, which activates intramuscular nerve branches and not the muscle fibres directly (Hultman et al., 1983). Even if this latter modality is largely adopted for central fatigue (twitch interpolation) and contractile fatigue assessment, three important limitations have to be acknowledged: muscle activation is incomplete and superficial; M waves cannot be easily recorded; central contributions from the recruitment of spinal motoneurons could overestimate considerably the recorded force (Collins, 2007). Therefore, nerve and muscle electrical stimulation should be considered as two distinct techniques of stimulation, which cannot be used interchangeably for fatigue studies. Nerve stimulation should always be used, whenever possible, to assess neuromuscular function.
Part III: techniques and procedures to evaluate metabolic mechanisms of fatigue
Thirty years ago, Hoult et al. reported for the first time that phosphorus metabolites could be observed in vivo using 31P Magnetic Resonance Spectroscopy (MRS), opening promising opportunities of understanding muscle energetics in vivo under strictly non-invasive conditions (Hoult et al., 1974). From that time, MRS technology has rapidly evolved with the development of RF-surface coils in 1980 (Ackerman et al., 1980), the availability of high-field, wide-bore superconducting magnets and methodological developments (dedicated pulse sequences, spatial localisation of NMR signal), making MRS a tool of choice for investigating muscle energetics non-invasively in animals and humans. So far, a large number of publications have been devoted to the investigation of muscle energetics in a variety of conditions ranging from diseases (Chance et al., 1980; Cozzone et al., 1996; Argov et al., 2000) to training regimens (McCully et al., 1988; Kent-Braun et al., 1990; Hug et al., 2004), as compared to normal conditions. Such studies have provided interesting information regarding not only pathologies and metabolic changes associated with training, but also about what normally occurs throughout muscle contraction in terms of balance between energy production and consumption. In this review, we intend to provide key information to the non-specialist in order to understand what 31P MRS can tell us about muscle energetics. After a brief introduction regarding the MR techniques and the corresponding information obtained in exercising muscle, we will present the different ergometers used in order to measure the mechanical performance throughout MR investigation of muscle fatigue both in humans and in animals. The MR results related to muscle fatigue are beyond the scope of the present chapter.
Informational content of a 31P MR spectrum
Measurement of phosphorylated compound concentrations in living cells is not easy. Traditional methods (such as percutaneous needle biopsy and freeze clamping) exhibit limitations, especially related to alteration of anatomic integrity and partial degradation of phosphorylated metabolites during extraction and analysis. In addition, repeated measurements cannot be performed on the same muscle, making the achievement of high time-resolution kinetics impossible. Compared to analytical methods, 31P MRS offers the opportunity of measuring non-invasively and continuously with high time-resolution, the concentration of phosphorylated compounds involved in muscle energetics. In addition, comparison of direct biochemical measurements with 31P MRS findings suggests that the two methods give comparable results (Tarnopolsky and Parise, 1999).
MR spectra are actually generated by placing samples in a powerful magnetic field and then exciting them with a radio-frequency energy. A typical 31P MRS spectrum exhibits 6–7 peaks corresponding to phosphocreatine (PCr), inorganic phosphate (Pi), the three phosphate groups of ATP (in position α, β and γ) and phosphomonoesters (PME) (Figures 2.1 and 2.2). The Pi signal of ATP displays occasionally an up-field shoulder corresponding to NAD+/NADH. Between the PCr and Pi signals, the phosphodiester signal is sometimes observed. This signal is usually assigned to glycerophosphorylcholine and glycerophosphoryethanolamine, which can be detected as a small peak in normal muscle spectra (mainly from the lower limb) and as a larger peak in patients with muscle dystrophy (Lodi et al., 1997), indicating membrane breakdown. Given the low sensitivity of the technique, the free metabolically active ADP concentration, which is only a tiny fraction of its total intracellular concentration, cannot be measured. It can, however, be calculated using the creatine kinase equilibrium, where the total creatine content is taken as either 42.5mM or considering that phosphocreatine represents 85% of the total creatine content (Kemp et al., 2001). Similarly, AMP concentration can be calculated using the adenylate kinase equilibrium (Kemp et al., 2001). In the absence of biochemical data, ATP is often assumed to be normal and is used as the equivalent of an internal standard in order to calculate the concentrations of other metabolites.

Figure 2.1 Typical series of 31P MRS recorded in human forearm flexor muscles. MRS have been recorded at 4.7T (Biospec 47/30 Bruker) during a standardised rest-exercise-recovery protocol with a time resolution of 15s. A single spectrum with the corresponding assignments is represented on the higher panel. Ref: reference compound (phenyl phosphonic acid), PME (phosphomonoesters), Pi (inorganic phosphate), PDE (phosphodiesters), PCr (phosphocreatine), phosphate groups of ATP in position γ, α and β

Figure 2.2 Typical PCr (A) and pH (B) time-dependent changes recorded throughout a standardised rest-exercise (shaded area)-recovery protocol in a group of 18 subjects. Measurements (black and white symbols) have been recorded seven days apart. Results are presented as means ±SD, and one can observe the very good reproducibility of measurements
Apart from the dynamic measurements of high-energy phosphate compounds, 31P MRS offers the only non-invasive way of assessing intracellular pH. Indeed, under conditions of physiological pH, and considering one of the pKa, i.e. 6.75, two forms of Pi coexist (H2PO4− and HPO42−). These two forms are exchanging so fast that only a single Pi signal is detected. However, the chemical shift of this single signal is a weighted average of both monobasic and dibasic forms. Due to this sensitivity of the Pi chemical shift to pH, it is possible, with appropriate calibration curves, to translate any Pi shift in terms of intracellular acidosis or alkalosis.
The quantitative measurements of high-energy phosphate compounds and pH allow computation of a number of derived metabolic parameters, such as the oxidative phosphorylation potential and the free energy of ATP hydrolysis in vivo. All those parameters reflect the regulation and control of energy metabolism, ion transport and muscular contraction.
Also, magnesium concentration has been calculated from changes in ATP chemical shift on the basis that the chemical shift of the resonance corresponding to the beta group is sensitive to the intracellular magnesium concentration (Gupta et al., 1983; Ward et al., 1996; Iotti et al., 2000). Fat, fibrous tissue blood and extracellular fluid contribute no significant signal and mitochondrial metabolites are too tightly bound to interfere.
Given the low magnetic resonance sensitivity of 31P (6% of proton) and the low tissue concentrations of some of the relevant metabolites, MR signals are time-averaged over a period ranging from a few seconds to several minutes, depending on the required signal-to-noise ratio and the desired time-resolution. In addition, the MR signal is detected with a surface coil over a relatively large muscle volume proportional to the surface coil radius, making this signal a weighted average of the muscle fibres existing within the sampling volume. This has to be taken as a comparative item with histological and biochemical analyses, which are often carried out on very small samples of tissues, which may not give a representative picture of the biochemical state of the muscle. However, care has to be taken not to turn this advantage into a drawback while sampling exercising and non-exercising muscles at the same time. MRS can be combined with MRI in order to achieve a proper localisation of the coil.
Technical considerations
The requirement for magnetic field homogeneity generally dictates that the muscle examined be positioned at magnet centre, and remains in a fixed position during data collection. In that respect, dedicated ergometers have been designed in each laboratory in order to investigate exercising muscles within superconducting magnets. So far, adductor pollicis (Miller et al., 1987), forearm and wrist flexor muscles (Chance et al., 1980; Arnold et al., 1984; Bendahan et al., 1990), calf (Ryschom et al., 1995; Lodi et al., 2002) and thigh muscles (Bernus et al., 1993; Rodenburg et al., 1994; Whipp et al., 1999; Francescato and Cettolo, 2001) have been investigated using 31P MRS. These ergometers are intended to allow muscle exercise within a superconducting magnet and should provide accurate measurements of the corresponding mechanical performance so that metabolic changes can be properly analysed with respect to the mechanical measurements. Initially, basic ergometers have been designed for isometric contractions, but due to the absence of movement, mechanical work and power output could not be quantified (Miller et al., 1988; Bangsbo et al., 1993). Other ergometers, including bulbs (Quistorff et al., 1990; Saab et al., 2000), elastic bands (Francescato and Cettolo, 2001) or cable-and-pulley systems (Bendahan et al., 1990; Marsh et al., 1991; McCreary et al., 1996; Nygren and Kaijser, 2002; Raymer et al., 2004) have been designed for dynamic contractions of wrist and plantar flexor muscles. For the fewer ergometers developed for quadriceps, exercise was either limited to a single leg (Park et al., 1990; Weidman et al., 1991; Smith et al., 1998; Barker et al., 2006) or work-rate was not accurately determined (Gonzalez et al., 1993; Whipp et al., 1999). Indeed, no quantitative measurements of weight displacement, contraction frequency or displacement speed was performed, so the resulting power output was just roughly estimated (Whipp et al., 1999). Using a more elaborate ergometer, Rodenburg et al. reported both force and displacement speed values, but mentioned workload reduction due to temperature changes during a prolonged exercise (Rodenburgh et al., 1994). More recently, a new ergometer allowing dynamic and isometric knee extension exercise within a whole body 1.5 T Siemens Vision Plus magnet has been proposed. As reported by Layec et al. (2008), mechanical measurements obtained with this ergometer were highly reliable and accurate without any significant effect of magnetic field. In addition, it was possible to assess energy metabolism over a broad range of exercise intensities and for subjects with different heights. This ergometer allows dynamic (unilateral or bilateral) and isometric knee extension exercises using both legs. The isometric facility was particularly important, given that it allows a reliable standardisation of exercise on the basis of MVC measurements.
In addition to experiments conducted in humans, animal experiments can also be conducted as long as mechanical measurements can also be recorded and, more importantly, muscle can be exercised. Paradoxically, in animal research the benefit of MR being non-invasive is lost, because the experimental set-ups usually incorporate invasive systems to induce muscle contractions by direct nerve stimulation or to measure mechanical performance with a force transducer attached to the muscle tendon (Foley and Meyer, 1993; Hogan et al., 1998; Cieslar and Dobson, 2001). Surgical intervention is required to position the stimulation electrode on the nerve and to introduce the force transducer. In addition to the risk of disturbing muscle physiology, in particular the neurovascular supply, these interventions necessitate sacrificing the animal after the experiment and thus prohibit repetitive explorations in the same animal. Interestingly, Giannesini et al. (2005) have designed and constructed a new apparatus integrating two non-invasive systems for transcutaneous muscle stimulation and force measurement respectively (Figure 2.3). Briefly, this system integrates four distinct components allowing prolonged anaesthesia with control of the animal’s body temperature, transcutaneous electrical stimulation of the gastrocnemius muscle, force output measurement and multimodal MR acquisition. Prolonged anaesthesia is maintained by gas inhalation via a custom-built face-mask connected to an open-circuit gas anaesthetic machine. In order to maintain the rat at physiological temperature during anaesthesia, the cradle integrates an electric heating blanket in a feedback loop with a temperature control unit connected to a rectal probe. Transcutaneous stimulation of the gastrocnemius muscle is performed using two surface electrodes connected to an electrical stimulator. Electrodes are integrated in the cradle so that when the rat is placed in the cradle, one electrode is located above the knee, and the other at the heel. Force output is measured with a custom-built ergometer consisting of a foot pedal (20×42mm) directly connected to a hydraulic piston. The foot pedal rotates freely on a nylon axis, which is situated at the level of the ankle joint. A hydraulic circuit filled with water connects the piston to a pressure transducer, which is placed outside the magnet. The pedal position can be adjusted such as to modify the angle between foot and lower hind limb. This adjustment enables the gastrocnemius muscle to be stretched passively in order to obtain maximum force production in response to electrical stimulation. MR acquisition is performed using a custom-built spectroscopy/imaging probe consisting of an elliptic 31P MRS surface coil (10×16mm) geometrically decoupled inside a 30mm diameter Helmholtz imaging coil. Using this system, the failure of muscle performance has been compared among different stimulation protocols.
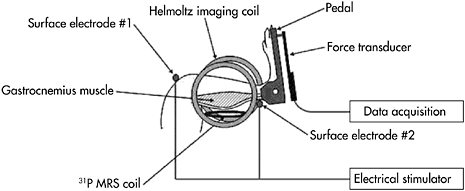
Figure 2.3 Schematic representation of the non-invasive setup allowing in vivo MR investigation of skeletal muscle function in rat gastrocnemius muscle
Reproducibility of measurements
For meaningful comparisons, data must be reproducible from study to study and it is important to assess repeatability in groups of subjects and patients. Foremost, it must be remembered that metabolic changes in a control population are highly heterogeneous (Miller et al., 1987; Bendahan et al., 1990; Miller et al., 1995; Kemp et al., 2001). The usual standardisation procedures, such as measurements of maximum voluntary contraction, do not reduce this between-subject variability (Bendahan et al., 1990; Miller et al., 1995; Kemp et al., 2001). In that respect, it is clear that a set of factors which could influence metabolic changes in exercising muscle has to be taken into account and ad hoc standardisation procedures that would reduce the inter-subject variability have to be used for proper comparisons among various groups of subjects.
Between-subject variability
In a limited number of subjects, it has been shown that neither fasting nor carbohydrate loading significantly modified the extent of metabolic changes in exercising muscle (Miller et al., 1995). Similarly, severe metabolic acidosis induced by ammonium chloride loading had no effect on changes recorded during exercise (Miller et al., 1995). Ageing is another potential factor that should be taken into account. The studies based on measurements of enzymatic activities have suggested a decreased oxidative capacity with age (Kohrt et al., 1991; Coggan et al., 1992). Results from MRS investigations differed in whether physical activity has been taken into account or not (Russ and Kent-Braun, 2004). Ageing has been related to both a 50% reduction in the PCr recovery rate and a reduced mitochondrial content, with no alteration regarding the metabolic changes in exercise (Conley et al., 2000). When physical activity has been taken into account, no deficit of aerobic capacity has been reported, whereas the PCr depletion was not modified and the pH was more alkalotic in older subjects (Kent-Braun and Ng, 2000), in agreement with previous studies conducted in sedentary and moderately active subjects (Chilibeck et al., 1998). These results would imply a primary role of disuse in the decline of oxidative capacity rather than an inherent age-related defect. In addition, the recent observation of increased maximum aerobic capacity in older subjects as a result of training would be in keeping with the hypothesis that the decline in oxidative capacity would mainly result from a reduction in habitual physical activity rather than of ageing per se (Jubrias et al., 2001). However, this conclusion must be moderated on the basis of two points. First, the oxidative capacity per mitochondrial volume measured in older subjects after a training period was still below the value reported in young adult muscle (Conley et al., 2000); and adaptive mechanisms were distinct from what has been reported so far in young adult muscle, especially regarding resistance training (Jubrias et al., 2001).
Another factor that should be taken into account regarding the between-subject variability is related to high body fat stores. Indeed, changes in fibre type proportions that could affect muscle energetics have been reported in obese subjects (Wade et al., 1990) and high-energy phosphate metabolism has been investigated using 31P MRS in order to test this hypothesis. Studies conducted in moderately obese women (BMI range: 27–30kg·m2) (Larson-Meyer et al., 2000b) and in prepubertal girls with a familial predisposition to obesity (Treuth et al., 2001) have clearly shown that muscle energetics was not altered. In addition, a weight-reduction programme had also no effect on mitochondrial function, thereby suggesting that a low substrate oxidative capacity of skeletal muscle is not involved in the pathogenesis of obesity on the contrary to what has been suggested earlier (Kriketos et al., 1996).
Within-subject variability
Beside the between-subject variability, the within-subject variability is also important to analyse. Several studies have clearly demonstrated very low within-subject variability, as illustrated in Figure 2.2, and indicated a very high reproducibility (Miller et al., 1995; Larson-Meyer et al., 2000a; Taylor, 2000; Bendahan et al., 2002). For instance, variation coefficients calculated from repeated experiments were 5–10% for metabolic indices at rest, such as ADP and PCr/Pi, and 3–15% for variables measured during exercise and in the recovery period (Larson-Meyer et al., 2000a). The reliability of MRS parameters has also been investigated in children in response to three repeated exhaustive ramp exercise tests (Barker et al., 2006). Indices recorded at rest, at the end of exercise and a threshold value (IT) related to the combined evolution of pH and Pi/PCr ratio were calculated. While the inter-observer variability was estimated as 5%, the coefficient of variations (CV) associated with the Pi/PCr ratios recorded at rest, at end of exercise and the IT values were 37%, 50% and 16%, respectively (Barker et al., 2006). This indicates an acceptable reproducibility for IT values, but a poor reliability for Pi/PCr values. Values related to pH were also highly reproducible, with CV corresponding to 0.6% and 0.9% for resting and end-of-exercise values.
Standardisation procedures
This within-subject variability may also be used in order to understand muscle energetics and to initiate ad hoc standardisation procedures that could be used as reliable comparative methods between groups of subjects. It has been shown in normally active adults that an index of PCr consumption ((PCr+Pi)/PCr) (measured with 31P MRS) was highly correlated with power output normalised to the volume of muscle (measured with MRI) in the plantar flexor compartment, indicating that combined MRS and MRI measurements could offer a way of reliably comparing subjects with different body size (Fowler et al., 1997). Similarly, expressing metabolic changes such as PCr breakdown and intracellular acidosis in exercise muscle with respect to power output has been reported to offer an interesting standardisation procedure, independent of exercise protocols and anthropometric measurements (Mattei et al., 2002). In agreement with such an approach, it has been shown, in exercising muscle, that the extent of PCr breakdown
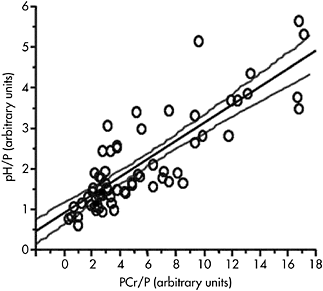
Figure 2.4 Linear relationship between PCr and pH values scaled to power output (P) and recorded in a group of subjects for different exercise intensities. Results are presented as arbitrary units. This linear relationship allows the standardisation of metabolic changes regardless of exercise intensity
was linearly linked to the intracellular acidosis, regardless of exercise protocols (Bendahan et al., 1990; Mattei et al., 2002), as illustrated in Figure 2.4. This suggests that various combinations of [PCr] and pH are multiple solutions for muscle fibres to reach a given level or range of ADP, which in turn will act as a regulator of energy production as previously described (Kemp et al., 1993).
Future investigations
Employing non-MR-based techniques simultaneously with MRS can aid in data interpretation and will certainly broaden the scope of muscle investigation. Near-infrared spectroscopy (NIRS) provides data on tissue oxygenation, but reliability has still to be proven, mainly on the basis of comparative and/or combined analyses using MRS and NIRS (Hamaoka et al., 1996). Electromyography is another technique of interest which can be used to study correlation between metabolic and electrical changes and provide interesting features regarding muscle fatigue (Bendahan et al., 1996). Functional MRI can also be of interest for understanding muscle activation during exercise. Based on T2 changes due to uptake or redistribution of fluid within the exercising muscle, functional MRI is considered as a semi-quantitative method of assessing muscle recruitment during exercise (Meyer et al., 2001).
CONCLUSION
It is concluded that maximal power (dynamic exercise) or MVC force (static exercise) can be used as a sort of gold standard to identify whether fatigue occurs or not. For fatigue studies based on power output assessment during dynamic exercise, mean power (cycling), torque slope (isokinetic) and speed (running) are better outcomes than peak power. The reliability of fatigue indexes obtained in these conditions should, however, be improved in the future. On the other hand, a series of isometric MVC and stimulated contractions (at rest and MVC-superimposed) completed before and immediately after exercise using concomitant force and EMG recordings can provide a valid and rapid assessment of central and peripheral fatigue induced by sport, work and daily activities.
MR spectroscopy and imaging can be used in order to investigate the metabolic and functional bases of muscle fatigue. The biological variability of results has to be acknowledged in order to determine invariant criteria which characterise “normality”. A multimodal MR approach including MRS and MRI, together with other non-invasive techniques, should be chosen in order to thoroughly identify the exact mechanisms leading to muscle fatigue.
FIVE KEY PAPERS THAT SHAPED THE TOPIC AREA
Study 1. Vøllestad, N.K. (1997). Measurement of human muscle fatigue. Journal of Neuroscience Methods, 74, 219–227.
A short review focusing on the measurement methods of human muscle fatigue. After a quick introductory section, and basic definitions of fatigue (MVC force, maximal evocable force, maximal power output, muscle fatigue and central fatigue), the author presents the possible sites of fatigue from CNS processes to force/power output, and kindly relates these sites to the methods described in the last part of the review. These methods include direct assessments such as MVC force generation, power output, tetanic force and low-frequency fatigue, which should be privileged in fatigue studies, but also indirect assessments (twitch interpolation, endurance time, EMG), which are criticised for their limited value in measurement of human muscle fatigue.
Study 2. Gandevia, S.C. (2001). Spinal and supraspinal factors in human muscle fatigue. Physiological Reviews, 81, 1725–1789.
This excellent review covers all there is to know about central fatigue. After presenting the historical aspects and definitions related to central fatigue (“a progressive reduction in voluntary activation of muscle during exercise”), and providing evidence about the notion of “submaximal” voluntary activation during “maximal” efforts, the author describes in detail the spinal (e.g. altered input from muscle spindle, tendon organ and group III and IV muscle afferents) and supraspinal (e.g. changes in cortical excitability and inhibitability) mechanisms of muscle fatigue, as inferred from electrophysiological recordings (voluntary activation, evoked reflexes, motor-evoked potentials, etc.).
Study 3. Cairns, S.P., Knicker, A.J., Thompson, M.W. and Sjøgaard, G. (2005). Evaluation of models used to study neuromuscular fatigue. Exercise Sport Sciences Reviews, 33, 9–16.
This original review describes and appraises the different approaches (models) used to study neuromuscular fatigue in human subjects and muscle preparations. The authors suggest that the cause of fatigue has to be regarded as multi-factorial, and that the mechanisms of neuromuscular fatigue relate to the entire approach of how fatigue is studied (i.e. the fatigue model involved). They also propose that the diversity of fatigue mechanisms can be explained, in part, by the use of different fatigue measures (muscle force, displacement, work or power) and the timing of their measurement. Several methodological considerations are proposed, in line with the idea that fatigue assessment should depend on the specific question to be addressed. At the end of the paper, specific recommendations for models to study neuromuscular fatigue are provided, which are extremely helpful to researchers for designing rigorous and clever fatigue studies.
Study 4. Giannesini, B., Izquierdo, M., Le Fur, Y., Cozzone, P.J., Fingerle, J., Himber, J., Kunnecke, B., Von Kienlin, M. and Bendahan, D. (2005). New experimental setup for studying strictly noninvasively skeletal muscle function in rat using 1H-magnetic resonance (MR) imaging and 31P-MR spectroscopy. Magnetic Resonance in Medicine, 54, 1058–1064.
This paper describes for the first time a new experimental setup allowing strictly non-invasive investigation of muscle function using MR techniques. The corresponding setup integrates two systems allowing muscle contraction by transcutaneous stimulation and force measurement with a dedicated ergometer. The experimental results provide a direct comparison between two different set-ups, and clearly demonstrate that longitudinal studies are now possible with this original setup.
Study 5. Mattei, J.P., Kozak-Ribbens, G., Roussel, M., Le Fur, Y., Cozzone, P.J. and Bendahan, D. (2002). New parameters reducing the interindividual variability of metabolic changes during muscle contraction in humans. A 31P MRS study with physiological and clinical implications. Biochimica et Biophysica Acta, 1554, 129–136.
Considering that inter-individual variations in skeletal muscle metabolism can be a problem for reliable comparative analyses, the authors describe in the present study an original normalisation method based on relationships between metabolic and work-rate indices recorded during a 31P MRS study. The study, conducted with a group of 65 subjects, demonstrates that the amplitude of mechanical performance accounts for 50% of the between-subjects variations. They report a linear relationship, independent of any anthropometric measurements, capturing more than 90% of the variability.
GLOSSARY OF TERMS
31P MRS | 31 phosphorus magnetic resonance spectroscopy |
ADP | adenosine diphosphate |
AMP | adenosine monophosphate |
ATP | adenosine triphosphate |
BMI | body mass index |
CV | coefficient of variation |
EMG | electromyographic activity |
H reflex | Hoffmann reflex |
H2PO4− | ion dihydrogenorthophosphate |
HPO42− | ion hydrogenorthophosphate |
IT | intra threshold |
MRI | magnetic resonance imaging |
MRS | magnetic resonance spectroscopy |
MVC | maximal voluntary isometric contraction |
NAD | nicotinamide adenine dinucleotide |
NIRS | near-infrared spectroscopy |
NMR | nuclear magnetic resonance |
PCr/Pi | phosphocreatine/inorganic phosphate ratio |
PCr | phosphocreatine |
Pi | inorganic phosphate |
RF | radio frequency |
T reflex | tendon reflex |
ACKNOWLEDGEMENTS
The research activity related to muscle at CRMBM (UMR CNRS 6612, headed by Professor P.J.Cozzone) is conducted in collaboration with B. Giannesini, J.Gondin, S.Guis, Y.Le Fur, G.Layec, J.P.Mattei, C.Vilmen, A.Tonson and K.Yashiro. Financial support has been obtained from CNRS (Centre National de la Recherche Scientifique), ANR (Agence Nationale pour le Recherche), AFM (Association Frangaise contre les Myopathies) and PHRC (Programme Hospitalier de Recherche Clinique).
REFERENCES
Ackerman, J.J., Grove, T.H., Wong, G.G., Gadian, D.G. and Radda G.K. (1980). Mapping of metabolites in whole animals by 31P NMR using surface coils. Nature, 283, 167–170.
Argov, Z., Lofberg, M. and Arnold, D.L. (2000). Insights into muscle diseases gained by phosphorus magnetic resonance spectroscopy. Muscle and Nerve, 23, 1316–1334.
Arnold, D.L., Matthews, P.M. and Radda G.K. (1984). Metabolic recovery after exercise and the assessment of mitochondrial function in vivo in human skeletal muscle by means of 31P NMR. Magnetic Resonance in Medicine, 1, 307–315.
Avela, J., Kyrolainen, H. and Komi, P.V. (1999). Altered reflex sensitivity after repeated and prolonged passive muscle stretching. Journal of Applied Physiology, 86, 1283–1291.
Bangsbo, J., Johansen, L., Quistorff, B. and Saltin, B. (1993). NMR and analytic biochemical evaluation of CrP and nucleotides in the human calf during muscle contraction. Journal of Applied Physiology, 74, 2034–2039.
Barker, A., Welsman, J., Welford, D., Fulford, J., Williams, C. and Armstrong, N. (2006). Reliability of 31P-magnetic resonance spectroscopy during an exhaustive incremental exercise test in children. European Journal of Applied Physiology, 98, 556–565.
Barker, A.T., Jalinous, R. and Freeston, I.L. (1985). Non-invasive magnetic stimulation of human motor cortex. Lancet, 1, 1106–1107.
Bendahan, D., Confort-Gouny, S., Kozak-Reiss, G. and Cozzone, P.J. (1990). Heterogeneity of metabolic response to muscular exercise in humans. New criteria of invariance defined by in vivo phosphorus-31NMR spectroscopy. FEBS Letters, 272, 155–158.
Bendahan, D., Jammes, Y., Salvan, A.M., Badier, M., Confort-Gouny, S., Guillot, C. and Cozzone, P.J. (1996). Combined electromyography-31P-magnetic resonance spectroscopy study of human muscle fatigue during static contraction. Muscle and Nerve, 19, 715–721.
Bendahan, D., Mattei, J.P., Ghattas, B., Confort-Gouny, S., Le Guern, M.E. and Cozzone, P.J. (2002). Citrulline/malate promotes aerobic energy production in human exercising muscle. British Journal of Sports Medicine, 36, 282–289.
Bernus, G., Gonzalez de Suso, J.M., Alonso, J., Martin, P.A., Prat, J.A. and Arus, C. (1993). 31P-MRS of quadriceps reveals quantitative differences between sprinters and long-distance runners. Medicine and Science in Sports and Exercise, 25, 479–484.
Bigland-Ritchie, B., Jones, D.A., Hosking, G.P. and Edwards, R.H. (1978). Central and peripheral fatigue in sustained maximum voluntary contractions of human quadriceps muscle. Clinical Science and Molecular Medicine, 54, 609–614.
Bilodeau, M. (2006). Central fatigue in continuous and intermittent contractions of triceps brachii. Muscle and Nerve, 34, 205–213.
Bishop, D., Spencer, M., Duffield, R. and Lawrence, S. (2001). The validity of a repeated sprint ability test. Journal of Science and Medicine in Sport, 4, 19–29.
Bosco, C., Belli, A., Astrua, M., Tihanyi, J., Pozzo, R., Kellis, S., Tsarpela, O., Foti, C., Manno, R. and Tranquilli, C. (1995). A dynamometer for evaluation of dynamic muscle work. European Journal of Applied Physiology and Occupational Physiology, 70, 379–386.
Bosco, C., Luhtanen, P. and Komi, P.V. (1983). A simple method for measurement of mechanical power in jumping. European Journal of Applied Physiology and Occupational Physiology, 50, 273–282.
Boyas, S., Nordez, A., Cornu, C. and Guével, A. (2006). Power responses of a rowing ergometer: mechanical sensors vs. Concept2 measurement system. International Journal of Sports Medicine, 27, 830–833.
Cairns, S.P., Knicker, A.J., Thompson, M.W. and Sjøgaard, G. (2005). Evaluation of models used to study neuromuscular fatigue. Exercise Sport Sciences Reviews, 33, 9–16.
Chance, B., Eleff, S. and Leigh Jr, J.S. (1980). Noninvasive, nondestructive approaches to cell bioenergetics. Proceedings of the National Academy of Sciences, 77, 7430–7434.
Chilibeck, P.D., McCreary, C.R., Marsh, G.D., Paterson, D.H., Noble, E.G., Taylor, A.W. and Thompson, R.T. (1998). Evaluation of muscle oxidative potential by 31P-MRS during incremental exercise in old and young humans. European Journal of Applied Physiology and Occupational Physiology, 78, 460–465.
Cieslar, J.H. and Dobson, G.P. (2001) Force reduction uncoupled from pH and H2PO4− in rat gastrocnemius in vivo with continuous 2-Hz stimulation. American Journal of Physiology Regulation, Integrative and Comparative Physiology, 281, R511–518.
Coggan, A.R., Spina, R.J., King, D.S., Rogers, M.A., Brown, M., Nemeth, P.M. and Holloszy, J.O. (1992). Histochemical and enzymatic comparison of the gastrocnemius muscle of young and elderly men and women. Journal of Gerontology, 47, B71–76.
Collins, D.F. (2007). Central contributions to contractions evoked by tetanic neuromuscular electrical stimulation. Exercise Sport Sciences Reviews, 35, 102–109.
Conley, K.E., Jubrias, S.A. and Esselman, P.C. (2000). Oxidative capacity and ageing in human muscle. Journal of Physiology, 526, 1, 203–210.
Cozzone, P.J., Vion-Dury, J., Bendahan, D. and Confort-Gouny, S. (1996). Future path of magnetic resonance spectroscopy in clinical medicine. La Revue du Praticien, 46, 853–858.
Desmedt, J.E. and Hainaut, K. (1968). Kinetics of myofilament activation in potentiated contraction: staircase phenomenon in human skeletal muscle. Nature, 217, 529–532.
Duchateau, J. and Hainaut, K. (1985). Electrical and mechanical failures during sustained and intermittent contractions in humans. Journal of Applied Physiology, 58, 942–947.
Edwards, R.H., Hill, D.K., Jones, D.A. and Merton, P.A. (1977). Fatigue of long duration in human skeletal muscle after exercise. Journal of Physiology, 272, 769–778.
Enoka, R.M. (2002). Neuromechanics of Human Movement. Champaign: Human Kinetics.
Farina, D. (2006). Interpretation of the surface electromyogram in dynamic contractions. Exercise Sport Sciences Reviews, 34, 121–127.
Farina, D., Merletti, R. and Enoka, R.M. (2004). The extraction of neural strategies from the surface EMG. Journal of Applied Physiology, 96, 1486–1495.
Fitzsimons, M., Dawson, B., Ward, D. and Wilkinson, A. (1993). Cycling and running tests of repeated sprint ability. Australian Journal of Science and Medicine in Sport, 25, 82–87.
Foley, J.M. and Meyer, R.A. (1993). Energy cost of twitch and tetanic contractions of rat muscle estimated in situ by gated 31P NMR. NMR in Biomedicine, 6, 32–38.
Fowler, M.D., Ryschon, T.W., Wysong, R.E., Combs, C.A. and Balaban, R.S. (1997). Normalised metabolic stress for 31P-MR spectroscopy studies of human skeletal muscle: MVC vs. muscle volume. Journal of Applied Physiology, 83, 875–883.
Francescato, M.P. and Cettolo, V. (2001). Two-pedal ergometer for in vivo MRS studies of human calf muscles. Magnetic Resonance in Medicine, 46, 1000–1005.
Funato, K., Yanagiya, T. and Fukunaga, T. (2001). Ergometry for estimation of mechanical power output in sprinting in humans using a newly developed self-driven treadmill. European Journal of Applied Physiology, 84, 169–173.
Gandevia, S.C. (2001). Spinal and supraspinal factors in human muscle fatigue. Physiological Reviews, 81, 1725–1789.
Giannesini, B., Izquierdo, M., Le Fur, Y., Cozzone, P.J., Fingerle, J., Himber, J., Kunnecke, B., Von Kienlin, M. and Bendahan, D. (2005). New experimental setup for studying strictly noninvasively skeletal muscle function in rat using 1H-magnetic resonance (MR) imaging and 31P-MR spectroscopy. Magnetic Resonance in Medicine, 54, 1058–1064.
Glaister, M., Stone, M.H., Stewart, A.M., Hughes, M. and Moir, G.L. (2004). The reliability and validity of fatigue measures during short-duration maximal-intensity intermittent cycling. Journal of Strength and Conditioning Research, 18, 459–462.
Gonzalez de Suso, J.M., Bernus, G., Alonso, J., Alay, A., Capdevila, A., Gili, J., Prat, J.A. and Arus, C. (1993). Development and characterization of an ergometer to study the bioenergetics of the human quadriceps muscle by 31P NMR spectroscopy inside a standard MR scanner. Magnetic Resonance in Medicine, 29, 575–581.
Gupta, R.K., Gupta, P., Yushok, W.D. and Rose, Z.B. (1983). On the noninvasive measurement of intracellular free magnesium by 31P NMR spectroscopy. Physiological Chemistry and Physics and Medical NMR, 15, 265–280.
Hamaoka, T., Iwane, H., Shimomitsu, T., Katsumura, T., Murase, N., Nishio, S., Osada, T., Kurosawa, Y. and Chance, B. (1996). Noninvasive measures of oxidative metabolism on working human muscles by near-infrared spectroscopy. Journal of Applied Physiology, 81, 1410–1417.
Hogan, M.C., Ingham, E. and Kurdak, S.S. (1998). Contraction duration affects metabolic energy cost and fatigue in skeletal muscle. American Journal of Physiology, 274, E397–402.
Hoult, D.I., Busby, S.J., Gadian, D.G., Radda, G.K., Richards, R.E. and Seeley, P.J. (1974). Observation of tissue metabolites using 31P nuclear magnetic resonance. Nature, 252, 285–287.
Hug, F., Bendahan, D., Le Fur, Y., Cozzone, P.J. and Grelot, L. (2004). Heterogeneity of muscle recruitment pattern during pedaling in professional road cyclists: a magnetic resonance imaging and electromyography study. European Journal of Applied Physiology, 92, 334–342.
Hughes, M.G., Doherty, M., Tong, R.J., Reilly, T. and Cable, N.T. (2006). Reliability of repeated sprint exercise in non-motorised treadmill ergometry. International Journal of Sports Medicine, 27, 900–904.
Hultman, E., Sjöholm, H., Jäderholm-Ek, I. and Krynicki, J. (1983). Evaluation of methods for electrical stimulation of human skeletal muscle in situ. Pflugers Archives, 398, 139–141.
Iotti, S., Frassineti, C., Alderighi, L., Sabatini, A., Vacca, A. and Barbiroli, B. (2000) In vivo (31)P-MRS assessment of cytosolic [Mg(2+)] in the human skeletal muscle in different metabolic conditions. Magnetic Resonance Imaging, 18, 607–614.
Jubrias, S.A., Esselman, P.C., Price, L.B., Cress, M.E. and Conley, K.E. (2001). Large energetic adaptations of elderly muscle to resistance and endurance training. Journal of Applied Physiology, 90, 1663–1670.
Keenan, K.G., Farina, D., Maluf, K.S., Merletti, R. and Enoka, R.M. (2005). Influence of amplitude cancellation on the simulated surface electromyogram. Journal of Applied Physiology, 98, 120–131.
Kemp, G.J., Roussel, M., Bendahan, D., Le Fur, Y. and Cozzone, P.J. (2001). Interrelations of ATP synthesis and proton handling in ischaemically exercising human forearm muscle studied by 31P magnetic resonance spectroscopy. Journal of Physiology, 535, 901–928.
Kemp, G.J., Taylor, D.J., Thompson, C.H., Hands, L.J., Rajagopalan, B., Styles, P. and Radda, G.K. (1993). Quantitative analysis by 31P magnetic resonance spectroscopy of abnormal mitochondrial oxidation in skeletal muscle during recovery from exercise. NMR in Biomedicine, 6, 302–310.
Kent-Braun, J.A. and Le Blanc, R. (1996). Quantitation of central activation failure during maximal voluntary contractions in humans. Muscle and Nerve, 19, 861–869.
Kent-Braun, J.A. and Ng, A.V. (2000). Skeletal muscle oxidative capacity in young and older women and men. Journal of Applied Physiology, 89, 1072–1078.
Kent-Braun, J.A., McCully, K.K and Chance, B. (1990). Metabolic effects of training in humans: a 31P-MRS study. Journal of Applied Physiology, 69, 1165–1170.
Kohrt, W.M., Malley, M.T., Coggan, A.R., Spina, R.J., Ogawa, T., Ehsani, A.A., Bourey, R.E., Martin, W.H. and Holloszy, J.O. (1991). Effects of gender, age, and fitness level on response of  to training in 60–71 yr olds. Journal of Applied Physiology, 71, 2004–2011.
to training in 60–71 yr olds. Journal of Applied Physiology, 71, 2004–2011.
Kriketos, A.D., Pan, D.A., Lillioja, S., Cooney, G.J., Baur, L.A., Milner, M.R., Sutton, J.R., Jenkins, A.B., Bogardus, C. and Storlien, L.H. (1996). Interrelationships between muscle morphology, insulin action, and adiposity. American Journal of Physiology, 270, R1332–1339.
Kufel, T.J., Pineda, L.A. and Mador, M.J. (2002). Comparison of potentiated and unpotentiated twitches as an index of muscle fatigue. Muscle and Nerve, 25, 438–444.
Lakomy, H.K.A. (1984). An ergometer for measuring the power generated during sprinting. Journal of Physiology, 354, 33.
Larson-Meyer, D.E., Newcomer, B.R., Hunter, G.R., Hetherington, H.P. and Weinsier, R.L. (2000a). 31P MRS measurement of mitochondrial function in skeletal muscle: reliability, force-level sensitivity and relation to whole body maximal oxygen uptake. NMR in Biomedicine, 13, 14–27.
Larson-Meyer D.E., Newcomer B.R., Hunter G.R., McLean J.E., Hetherington H.P. and Weinsier R.L. (2000b). Effect of weight reduction, obesity predisposition, and aerobic fitness on skeletal muscle mitochondrial function. American Journal of Physiology, Endocrinology and Metabolism, 278, E153–161.
Layec, G., Bringard, A., Vilmen, C., Micallef, J.P., Le Fur, Y., Perrey, S., Cozzone, P.J. and Bendahan, D. (2008). Accurate work-rate measurements during in vivo MRS studies of exercising human quadriceps. Magma, 21, 227–235.
Lodi, R., Muntoni, F., Taylor, J., Kumar, S., Sewry, C.A., Blamire, A., Styles, P. and Taylor, D.J. (1997). Correlative MR imaging and 31P-MR spectroscopy study in sarcoglycan deficient limb girdle muscular dystrophy. Neuromuscular Disorders, 7, 505–511.
Lodi, R., Rajagopalan, B., Bradley, J.L., Taylor, D.J., Crilley, J.G., Hart, P.E., Blamire, A.M., Manners, D., Styles, P., Schapira, A.H. and Cooper, J.M. (2002). Mitochondrial dysfunction in Friedreich’s ataxia: from pathogenesis to treatment perspectives. Free Radical Research, 36, 461–466.
McCartney, N., Heigenhauser, G.J., Sargeant, A.J. and Jones, N.L. (1983). A constant-velocity cycle ergometer for the study of dynamic muscle function. Journal of Applied Physiology, 55, 212–217.
McCreary, C.R., Chilibeck, P.D., Marsh, G.D., Paterson, D.H., Cunningham, D.A. and Thompson R.T. (1996). Kinetics of pulmonary oxygen uptake and muscle phosphates during moderate-intensity calf exercise. Journal of Applied Physiology, 81, 1331–1338.
McCully, K.K., Kent, J.A. and Chance, B. (1988). Application of 31P magnetic resonance spectroscopy to the study of athletic performance. Sports Medicine, 5, 312–321.
Maffiuletti, N.A., Bizzini, M., Desbrosses, K., Babault, N. and Munzinger, U. (2007). Reliability of knee extension and flexion measurements using the ConTrex isokinetic dynamometer. Clinical Physiology and Functional Imaging, 27, 346–353.
Margaria, R., Aghemo, P. and Rovelli, E. (1966). Measurement of muscular power (anaerobic) in man. Journal of Applied Physiology, 21, 1662–1664.
Marsh, G.D., Paterson, D.H., Thompson, R.T. and Driedger, A.A. (1991). Coincident thresholds in intracellular phosphorylation potential and pH during progressive exercise. Journal of Applied Physiology, 71, 1076–1081.
Mattei, J.P., Kozak-Ribbens, G., Roussel, M., Le Fur, Y., Cozzone, P.J. and Bendahan, D. (2002). New parameters reducing the interindividual variability of metabolic changes during muscle contraction in humans. A 31P MRS study with physiological and clinical implications. Biochimica et Biophysica Acta, 1554, 129–136.
Merton, P.A. (1954). Voluntary strength and fatigue. Journal of Physiology, 123, 553–564.
Meyer, R.A., Prior, B.M., Siles, R.I. and Wiseman, R.W. (2001) Contraction increases the T(2) of muscle in fresh water but not in marine invertebrates. NMR in Biomedicine, 14, 199–203.
Miller, R.G., Boska, M.D., Moussavi, R.S., Carson, P.J. and Weiner, M.W. (1988) 31P nuclear magnetic resonance studies of high energy phosphates and pH in human muscle fatigue. Comparison of aerobic and anaerobic exercise. Journal of Clinical Investigation, 81, 1190–1196.
Miller, R.G., Carson, P.J., Moussavi, R.S., Green, A., Baker, A., Boska, M.D. and Weiner, M.W. (1995). Factors which influence alterations of phosphates and pH in exercising human skeletal muscle: measurement error, reproducibility, and effects of fasting, carbohydrate loading, and metabolic acidosis. Muscle and Nerve, 18, 60–67.
Miller, R.G., Giannini, D., Milner-Brown, H.S., Layzer, R.B., Koretsky, A.P., Hooper, D. and Weiner, M.W. (1987). Effects of fatiguing exercise on high-energy phosphates, force, and EMG: evidence for three phases of recovery. Muscle and Nerve, 10, 810–821.
Millet, G.Y. and Lepers, R. (2004). Alterations of neuromuscular function after prolonged running, cycling and skiing exercises. Sports Medicine, 34, 105–116.
Millet, G.Y., Lepers, R., Maffiuletti, N.A., Babault, N., Martin, V. and Lattier, G. (2002). Alterations of neuromuscular function after an ultramarathon. Journal of Applied Physiology, 92, 486–492.
Nygren, A.T. and Kaijser, L. (2002). Water exchange induced by unilateral exercise in active and inactive skeletal muscles. Journal of Applied Physiology, 93, 1716–1722.
O’Brien, T.D., Reeves, N.D., Baltzopoulos, V., Jones, D.A. and Maganaris, C.N. (2008). Assessment of voluntary muscle activation using magnetic stimulation. European Journal of Applied Physiology, 104, 49–55.
Park, J.H., Vansant, J.P., Kumar, N.G., Gibbs, S.J., Curvin, M.S., Price, R.R., Partain, C.L. and James Jr, A.E. (1990). Dermatomyositis: correlative MR imaging and P-31 MR spectroscopy for quantitative characterization of inflammatory disease. Radiology, 177, 473–479.
Paton, C.D. and Hopkins, W.G. (2001). Tests of cycling performance. Sports Medicine, 31, 489–496.
Pierrot-Deseilligny, E. and Mazevet, D. (2000). The monosynaptic reflex: a tool to investigate motor control in humans. Interest and limits. Clinical Neurophysiology, 30, 67–80.
Pincivero, D.M., Gear, W.S. and Sterner, R.L. (2001). Assessment of the reliability of high-intensity quadriceps femoris muscle fatigue. Medicine and Science in Sports and Exercise, 33, 334–338.
Place, N., Maffiuletti, N.A., Martin, A. and Lepers, R. (2007). Assessment of the reliability of central and peripheral fatigue after sustained maximal voluntary contraction of the quadriceps muscle. Muscle and Nerve, 35, 486–495.
Place, N., Yamada, T., Bruton, J.D. and Westerblad, H. (2008). Interpolated twitches in fatiguing single mouse muscle fibres: implications for the assessment of central fatigue. Journal of Physiology, 86, 2799–2805.
Polkey, M.I., Kyroussis, D., Hamnegard, C.H., Mills, G.H., Green, M. and Moxham, J. (1996). Quadriceps strength and fatigue assessed by magnetic stimulation of the femoral nerve in man. Muscle and Nerve, 19, 549–555.
Quistorff, B., Nielsen, S., Thomsen, C., Jensen, K.E. and Henriksen, O. (1990). A simple calf muscle ergometer for use in a standard whole-body MR scanner. Magnetic Resonance in Medicine, 13, 444–449.
Racinais, S., Girard, O., Micallef, J.P. and Perrey, S. (2007). Failed excitability of spinal motoneurons induced by prolonged running exercise. Journal of Neurophysiology, 97, 596–603.
Raymer, G.H., Marsh, G.D., Kowalchuk, J.M. and Thompson, R.T. (2004). Metabolic effects of induced alkalosis during progressive forearm exercise to fatigue. Journal of Applied Physiology, 96, 2050–2056.
Rodenburg, J.B., de Boer, R.W., Jeneson, J.A., van Echteld, C.J. and Bar, P.R. (1994). 31P-MRS and simultaneous quantification of dynamic human quadriceps exercise in a whole body MR scanner. Journal of Applied Physiology, 77, 1021–1029.
Russ, D.W. and Kent-Braun, J.A. (2004). Is skeletal muscle oxidative capacity decreased in old age? Sports Medicine, 34, 221–229.
Ryschon, T.W., Fowler, M.D., Arai, A.A., Wysong, R.E., Leighton, S.B., Clem Sr, T.R. and Balaban, R.S. (1995). A multimode dynamometer for in vivo MRS studies of human skeletal muscle. Journal of Applied Physiology, 79, 2139–2147.
Saab, G., Thompson, R.T. and Marsh, G.D. (2000). Effects of exercise on muscle transverse relaxation determined by MR imaging and in vivo relaxometry. Journal of Applied Physiology, 88, 226–233.
Smith, S.A., Montain, S.J., Matott, R.P., Zientara, G.P., Jolesz, F.A. and Fielding, R.A. (1998). Creatine supplementation and age influence muscle metabolism during exercise. Journal of Applied Physiology, 85, 1349–1356.
Tarnopolsky, M.A. and Parise, G. (1999). Direct measurement of high-energy phosphate compounds in patients with neuromuscular disease. Muscle and Nerve, 22, 1228–1233.
Taylor, D.J. (2000). Clinical utility of muscle MR spectroscopy. Seminars in Musculoskeletal Radiology, 4, 481–502.
Thomas, C.K., Woods, J.J. and Bigland-Ritchie, B. (1989). Impulse propagation and muscle activation in long maximal voluntary contractions. Journal of Applied Physiology, 67, 1835–1842.
Thorstensson, A. and Karlsson, J. (1976). Fatiguability and fibre composition of human skeletal muscle. Acta Physiologica Scandinavica, 98, 318–322.
Todd, G., Taylor, J.L. and Gandevia, S.C. (2003). Measurement of voluntary activation of fresh and fatigued human muscles using transcranial magnetic stimulation. Journal of Physiology, 551, 661–671.
Tong, R.J., Bell, W., Ball, G. and Winter, E.M. (2001). Reliability of power output measurements during repeated treadmill sprinting in rugby players. Journal of Sports Sciences, 19, 289–297.
Treuth, M.S., Butte, N.F. and Herrick, R. (2001). Skeletal muscle energetics assessed by (31)P-NMR in prepubertal girls with a familial predisposition to obesity. International Journal of Obesity and Related Metabolic Disorders, 25, 1300–1308.
Trimble, M.H. and Harp, S.S. (1998). Postexercise potentiation of the H-reflex in humans. Medicine and Science in Sports and Exercise, 30, 933–941.
Vergès, S., Maffiuletti, N.A., Kerherve, H., Decorte, N., Wuyam, B. and Millet, G.Y. (2009). Comparison of electrical and magnetic stimulations to assess quadriceps muscle function. Journal of Applied Physiology, 106, 701–710.
Vøllestad, N.K. (1997). Measurement of human muscle fatigue. Journal of Neuroscience Methods, 74, 219–227.
Wade, A.J., Marbut, M.M. and Round, J.M. (1990). Muscle fibre type and aetiology of obesity. Lancet, 335, 805–808.
Ward, K.M., Rajan, S.S., Wysong, M., Radulovic, D. and Clauw, D.J. (1996). Phosphorus nuclear magnetic resonance spectroscopy: in vivo magnesium measurements in the skeletal muscle of normal subjects. Magnetic Resonance in Medicine, 36, 475–480.
Weidman, E.R., Charles, H.C., Negro-Vilar, R., Sullivan, M.J. and MacFall, J.R. (1991). Muscle activity localization with 31P spectroscopy and calculated T2-weighted 1H images. Investigative Radiology, 26, 309–316.
Whipp, B.J., Rossiter, H.B., Ward, S.A., Avery, D., Doyle, V.L., Howe, F.A. and Griffiths, J.R. (1999). Simultaneous determination of muscle 31P and O2 uptake kinetics during whole body NMR spectroscopy. Journal of Applied Physiology, 86, 742–747.
Wilson, G.J. and Murphy, A.J. (1996). The use of isometric tests of muscular function in athletic assessment. Sports Medicine, 22, 19–37.