L’homme, artisan de communautés biologiques artificielles
Petites causes, grands effets. Les équilibres de la Nature sont à la merci d’une chiquenaude.
Roger Heim
Comme s’il voulait parfaire la création, l’homme a transporté délibérément plantes et animaux à travers le monde, soit pour satisfaire un plaisir sentimental en reconstituant les communautés naturelles familières de la patrie lointaine d’où il avait émigré, soit pour augmenter la productivité des régions où il s’est établi.
À côté de ces tentatives, certaines déjà anciennes, il convient de ne pas oublier l’introduction des animaux domestiques, susceptibles de provoquer des ruptures d’équilibre analogues à celles dont sont responsables les animaux libérés à l’état sauvage ; leur impact est même plus grave encore, car l’homme les protège artificiellement contre toute compétition réelle.
À ces actions délibérées de l’homme, il faut ajouter les transports involontaires, que l’accélération et l’augmentation du volume du trafic dans les temps modernes ont rendus beaucoup plus fréquents. Ils concernent plus particulièrement les végétaux sous forme de semences et les animaux de petite taille. Les séquelles de ces acclimatations véritables sont parfois incalculables.
L’homme s’est tout d’abord préoccupé de l’introduction d’oiseaux et de mammifères, et depuis longtemps on en connaît les conséquences spectaculaires ; l’introduction d’éléments de la microfaune, dont le rôle biologique est fondamental, a de ce fait été négligée.
Sans entrer dans le détail de ces opérations, il faut signaler l’importance du transport et de l’introduction des micro-organismes du sol. Les terres sont caractérisées par un équilibre entre des millions de végétaux et d’animaux microscopiques ou de très petite taille ; leur fertilité et leur stabilité en dépendent d’une manière très directe. Or par le transport de végétaux et de la terre qui les accompagne, l’homme a, sans même s’en rendre compte, introduit des éléments exogènes, dont la prolifération a pu modifier profondément les équilibres pédologiques. Cela est vrai dans le cas des Nématodes et même dans celui des Lombrics, les vulgaires « vers de terre », qui jouent un rôle dans la transformation des sols.
Les animaux et les végétaux occupent dans leur milieu d’origine des niches écologiques bien déterminées et se trouvent en équilibre avec les autres éléments de la biocénose. Leurs populations sont contrôlées par les effets de la compétition et de la prédation. En revanche leur introduction dans une biocénose nouvelle, où ils sont étrangers, est le plus souvent pleine de dangers, par suite de l’absence très fréquente de compétiteurs et d’ennemis naturels susceptibles de limiter leurs effectifs. Leur acclimatation est de ce fait suivie d’une alternative : ou bien ils n’arrivent pas à prendre pied et disparaissent rapidement sans faire souche, étant en quelque sorte « étouffés » par le milieu ; ou bien ils réussissent pleinement, prolifèrent et deviennent alors des « pestes ». Leur succès explosif tourne à plus ou moins longue échéance à la catastrophe pour les habitats naturels, pour les animaux et les végétaux autochtones, et souvent même pour l’économie humaine.
Le succès des animaux introduits s’explique de différentes manières. L’espèce transportée par l’homme peut faire irruption dans une biocénose où une niche écologique susceptible de lui convenir est vacante, donc occupée immédiatement et sans peine. Elle peut également se comporter comme un compétiteur plus robuste et mieux armé que les homologues autochtones qui se trouvent ainsi éliminés. Elle peut enfin être un prédateur vis-à-vis duquel les espèces indigènes, constituant les proies, se trouvent désarmées. Notons d’ailleurs que la plupart du temps les espèces introduites font preuve d’une grande souplesse écologique et ne s’établissent pas dans la niche écologique que les responsables de l’acclimatation espéraient leur voir occuper. Elles modifient souvent leur mode de vie, notamment en ce qui concerne leur régime alimentaire163. Ce changement, souvent en rapport avec des conditions écologiques différentes, peut aussi résulter de modifications génétiques, en raison du nombre relativement réduit de géniteurs introduits, ce qui entraîne une redistribution du patrimoine héréditaire164.
Les acclimatations tentées dans les temps anciens ont été renouvelées à maintes reprises dans les temps modernes. Avec de nombreuses introductions involontaires, elles ont accentué le déséquilibre consécutif à d’autres facteurs, et provoqué la ruine de certaines communautés naturelles. Peu ont été bénéfiques à l’homme, excepté celles de quelques végétaux, les méfaits se manifestant d’une manière très variée. Quelques-unes de ces acclimatations seront évoquées à titre d’exemples pour avoir une vue d’ensemble de la situation actuelle165.
1. Transport et acclimatation de végétaux
Depuis des temps immémoriaux, l’homme a transporté de nombreux végétaux à travers le monde, les plantes cultivées étant bien entendu les plus anciennement acclimatées de toutes. Parallèlement aux défrichements, elles ont contribué à créer des milieux entièrement artificiels ; leur importance est de ce fait primordiale. Nous ne nous attacherons cependant pas à cet aspect du problème, quel que soit l’impact des plantes cultivées dans le monde vivant, car leur culture exige des transformations profondes des habitats auxquelles il a été fait allusion par ailleurs.
L’homme a simultanément modifié les habitats naturels par l’acclimatation de plantes sauvages. De nombreuses plantes familières des campagnes d’Europe et d’Amérique du Nord semblent se trouver dans leur patrie d’origine, faire partie du paysage depuis des générations. Et pourtant beaucoup ne sont que des intruses, venues à des époques diverses, parfois très reculées. L’homme a ainsi fabriqué de toutes pièces des communautés entières, souvent stables, ayant toutes les apparences de communautés naturelles ; l’influence humaine devient cependant manifeste dès que l’on entreprend une étude floristique approfondie. À part certaines associations de haute montagne et les habitats véritablement arctiques, il ne reste certainement aucune communauté végétale naturelle en Europe ; si ce fait provient en partie d’une modification profonde de l’équilibre des espèces autochtones, l’acclimatation de végétaux importés y a joué un rôle très important. Il en est de même en Amérique du Nord. Comme le rapporte Clark (1956), les associations végétales de Californie ne comprennent plus qu’un faible pourcentage de plantes autochtones, les espèces introduites étant même dominantes dans des formations relativement stables. L’homme a ainsi modelé le monde végétal sur une grande partie de la planète. On peut évidemment se demander ce qui arriverait si notre espèce disparaissait et si les plantes étaient abandonnées aux lois de l’équilibre naturel. Il est certain que dans bien des cas les espèces autochtones reprendraient le dessus ; mais les associations en seraient néanmoins profondément modifiées et les formations primitives ne se rétabliraient qu’au terme de temps très longs, parfois jamais.
L’influence humaine apparaît avec une particulière netteté dans le cas des formations graminéennes (Foury, Vernet, 1960). Depuis longtemps les agronomes ne peuvent plus « admettre que tout est pour le mieux dans le meilleur des mondes », même si en tant que biologistes ils sont « émerveillés par la perfection des équilibres naturels » (Vernet). Si le naturaliste est partisan de la plante spontanée, mieux adaptée aux conditions locales, l’agronome ne pense qu’à la productivité en fourrage et désire améliorer les pâturages par l’introduction d’espèces exogènes, souvent même après sélection en champ expérimental. Il remplace donc les associations naturelles par des formations entièrement artificielles à plus haut rendement ; les espèces introduites y occupent une place de premier plan.
La liste des espèces graminéennes ou herbacées introduites dans diverses parties du monde est fort longue et nous ne pouvons songer à la dresser ici. Parmi les principales Graminées fourragères cultivées à travers le monde, soit une quarantaine, 24 sont originaires d’Eurasie, 8 d’Afrique orientale, 4 d’Amérique du Sud, et 4 des autres régions du globe. La plupart des Légumineuses viennent d’Europe occidentale et méditerranéenne. Si l’Amérique n’a donné qu’une seule plante importante, le Maïs-fourrage, elle en a importé de nombreuses autres d’Europe. Divers Pâturins (blue grasses, Poa) et Agrostis (bent grasses) ont été largement répandus et se sont maintenant intimement incorporés à la flore nord-américaine. La plupart des plantes fourragères rencontrées actuellement dans les pâturages d’Australie et de Nouvelle-Zélande viennent de la région méditerranéenne. L’acclimatation de plantes similaires dans les zones intertropicales, beaucoup plus récente, est en pleine expansion ; on tend vers une uniformisation des pâturages, du Brésil à l’Afrique et à l’Indonésie. Notons que l’introduction de plantes fourragères s’est souvent accompagnée de celle d’un cortège de « mauvaises herbes » dont les graines se trouvaient mélangées à celles de l’espèce recherchée par l’homme, en dépit des triages les plus sévères.
Ces diverses introductions ont donc considérablement modifié les équilibres naturels des associations graminéennes. Leurs conséquences sur le plan de la conservation des habitats primitifs sont considérables.
Les forêts, surtout celles des zones tempérées, se sont trouvées elles aussi profondément transformées par l’introduction d’essences exogènes, autant que par la sélection artificielle des essences autochtones les plus intéressantes pour l’homme et par les méthodes de la sylviculture moderne. De très nombreux arbres ont été transportés à travers le monde.
Une mention spéciale doit être faite aux Eucalyptus, Myrtacées originaires d’Australie où ils constituent l’élément dominant de la végétation, maintenant acclimatés dans une grande partie des zones chaudes et tempérées (Penfold et Willis, 1961). Si les premières semences arrivèrent à Paris en 1804 et si aux alentours de 1810 des Eucalyptus prospéraient à la Malmaison, c’est seulement à partir de 1857 que des plantations véritables furent établies en Europe méridionale et en Afrique du Nord. Dès 1823, ces arbres étaient introduits au Chili, en 1828 en Afrique du Sud, dans l’Inde en 1843, en Californie en 1853 et en Argentine en 1857. De nombreuses espèces parmi les 500 qui forment le genre font l’objet de culture sur environ 1,4 million d’hectares à travers le monde. Rien qu’au Brésil, où l’Eucalyptus fut introduit entre 1855 et 1870, ces plantations occupent une surface de 800 000 ha et comprennent environ 2 000 millions d’arbres dont 1 200 millions dans le seul État de São Paulo. Les plantations d’Eucalyptus sont actuellement partout en pleine expansion, notamment sous l’impulsion de grands organismes internationaux.
Les Eucalyptus, dont les multiples espèces répondent à des besoins et à des exigences différents suivant la nature des sols et les climats, présentent d’incontestables avantages. Ils permettent de régénérer des terres abîmées par l’homme, comme ce fut notamment le cas dans le sud-est du Brésil, ravagé par la déforestation et les mauvaises pratiques culturales. En raison de leur croissance rapide, ils procurent à court terme une quantité appréciable de bois166 (bien que de qualité médiocre dans le cas de certaines espèces) constituant ainsi une ressource naturelle de valeur dans les pays déforestés où ils jouent un grand rôle économique. Cela est particulièrement important à l’époque actuelle en raison de la réduction des forêts et d’une demande croissante en produits ligneux. Mais les Eucalyptus ont été considérés à tort comme une panacée. À long terme le reboisement en autres essences est souvent plus profitable. Le biologiste ne trouve guère de satisfactions dans leurs forêts, de véritables déserts quant à leur peuplement animal.
Bien d’autres essences ont été transportées par l’homme en particulier dans le monde des Conifères. L’Amérique du Nord (surtout l’ouest) et le Japon, étant spécifiquement plus riches que l’Europe, ont tenté nos forestiers ; ils en ont importé une longue série d’espèces dont beaucoup se sont parfaitement adaptées à leur nouveau milieu, souvent mieux que les espèces autochtones. La vogue actuelle de l’« enrésinement » a été en grande partie satisfaite par l’introduction de ces essences à croissance rapide qui assurent un haut rendement aux forêts et ont permis la reconstitution ou le maintien d’un couvert végétal important dans de nombreuses régions du globe. Mais le remplacement des feuillus par des conifères a de multiples inconvénients sur le sol (dégradation, action défavorable sur la balance hydrique) et sur la faune, y compris le gibier.
Il faut bien entendu se garder de condamner a priori l’introduction de ces diverses plantes arborescentes, souvent suivie de réelles et durables réussites économiques ; il ne faut pas non plus partir en guerre d’une manière systématique contre l’aménagement des forêts qui augmente la productivité en bois de haute qualité. Il en est de même de l’amélioration des pâturages par l’introduction de nouvelles espèces graminéennes. Il y a néanmoins conflit entre le forestier et l’agronome d’une part, le conservateur de la nature d’autre part. L’homme a délibérément détruit certaines associations végétales par l’introduction de plantes exogènes favorisées au détriment des plantes autochtones. Et les méthodes modernes de la sylviculture et de l’agrostologie s’apparentent étroitement à celles de l’agriculture : on cultive des arbres ou des graminées fourragères comme des céréales ou des Betteraves. Cela aboutit de ce fait à la création de milieux d’autant plus artificiels que les espèces dominantes sont le plus souvent exogènes. Sans aucun doute le naturaliste doit comprendre la nécessité de la mise en valeur des terres et de leur transformation en vue d’une productivité maximale. En revanche l’économiste doit admettre la nécessité de conserver certaines parcelles dans leur état originel, de manière à assurer la survie, au moins à l’état de témoin, des communautés naturelles ; ces parcelles doivent être protégées avant tout contre l’intrusion de toute plante non autochtone qui viendrait en rompre l’équilibre. Ce n’est que par la compréhension et la bonne volonté de tous que l’on arrivera à concilier ces deux points de vue différents, mais parfaitement respectables l’un et l’autre.
Un grand nombre d’autres acclimatations végétales, volontaires ou non, se sont traduites par de véritables catastrophes.
Certaines plantes importées dans les régions tropicales pour aménager des barrières vivantes dans les zones d’élevage sont devenues des pestes en envahissant les pâturages. En Nouvelle-Calédonie (Barrau et Devambez, 1957), le Lantana Lantana camara, originaire d’Amérique tropicale et introduit au début de la colonisation comme plante d’ornement, fut préconisé comme barrière, ses longs rameaux épineux devant s’opposer au passage du bétail. Il ne tarda pas à se multiplier d’une manière incontrôlable, envahissant des pâturages tout entiers167 en compagnie d’autres plantes tout aussi prolifiques : Acacia farnesiana, la Sensitive géante Mimosa invisa (parvenue dans les années 1930 et ayant passé faussement pour une Légumineuse fourragère de grande valeur), et le Goyavier Psidium guajava importé d’Amérique. Ces plantes ont envahi les terres d’utilisation pastorale du versant occidental de l’île, au climat sec. Si elles contribuent parfois à protéger le sol contre l’érosion, leur multiplication a diminué considérablement le rendement des pâturages, au détriment de plantes autochtones plus intéressantes.
L’introduction des Opuntias en Australie fut plus catastrophique encore. Un seul pied d’Opuntia inermis importé en 1839 dans la Nouvelle-Galles du Sud s’y multiplia avec une vitesse accélérée. À la fin du siècle ce Cactus couvrait 4 millions d’hectares ; en 1920, environ 24 millions, et il continuait à progresser de 4 millions d’hectares par an. Les meilleures terres d’élevage disparaissaient sous ce fléau. Divers procédés de lutte furent essayés sans effet. Ce n’est qu’en 1925 qu’il vint à l’idée d’importer d’Uruguay et du nord de l’Argentine un petit Papillon, Cactoblastis cactorum, dont la chenille dévore les raquettes, y creusant des galeries par où pénètrent des agents de décomposition, bactéries et champignons. Le résultat fut spectaculaire, car ce Cactus disparut aussi vite qu’il avait progressé168.
Une autre cause d’introduction provient de l’établissement à l’état subspontané de plantes ornementales. Au moins quelque 20 000 espèces végétales font l’objet d’une pratique horticole quelconque. Bien entendu beaucoup d’entre elles ne peuvent vivre qu’en serre sous certains climats, et n’ont de ce fait aucune chance de se répandre dans la nature. Mais d’autres peuvent s’échapper de « captivité », s’adapter à la vie sauvage, entrer en compétition avec des espèces autochtones et tendre à les éliminer, voire à devenir de véritables « pestes » végétales. Le meilleur exemple est fourni par la Jacinthe d’eau Eichhornia crassipes, que certains n’ont pas hésité à appeler le « fléau vert ».
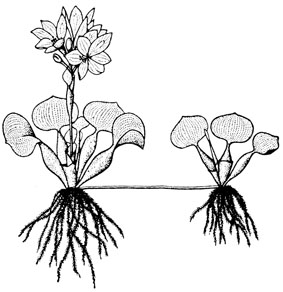
Fig. 41. Pied fleuri de Jacinthe d’eau Eichhornia crassipes avec jeune plante développée par stolon. D’après Robyns, 1955.
Monocotylédone de la famille des Pontédériacées, elle se présente comme une plante aquatique flottante, aux pétioles vésiculeux disposés en rosette et au rhizome immergé à faible profondeur (fig. 41). Ses fleurs forment de grands épis pourpres ou bleu mauve très décoratifs. La multiplication se fait cependant habituellement d’une manière végétative169.
Cette plante originaire de l’Amérique tropicale (ses centres de dispersion se trouvent surtout en Guyane et aux confins du Brésil et du Paraguay) fut introduite dans différentes régions chaudes du monde pour l’ornementation des bassins170. Elle ne tarda pas à s’en échapper et à coloniser la nature sauvage (Robyns, 1955).
La première apparition hors de son habitat normal se place dans le sud des États-Unis. Introduite en 1884 en Louisiane, puis en Floride en 1888, elle envahit tout le sud des États-Unis, jusqu’en Virginie et même en Californie, et devint rapidement une « peste »171, gênant même la navigation sur le Mississippi.
Cultivée dès 1894 au célèbre jardin botanique de Bogor (Buitenzorg), elle se répandit rapidement à Java, puis dans toute l’Indonésie, les Philippines, l’Australie, une partie des îles du Pacifique (îles Fidji et Hawaii entre autres). En 1902, elle était importée à Hanoï, d’où elle envahissait la péninsule indochinoise, l’Inde, y compris Ceylan (arrivée en 1905, elle y était déjà devenue un fléau en 1907).
En Afrique, bien qu’elle ait été présente à l’état subspontané au Zaïre dès 1910, c’est dans les temps récents (vers 1952) qu’elle se mit à pulluler et envahir le bassin du Congo et de ses affluents ; même à la hauteur de Kinshasa, le fleuve, devenu énorme, charrie des amas de Jacinthe d’eau, tandis que les bras plus étroits et les affluents sont obstrués au point que la navigation y est impraticable. La plante est en train de coloniser une partie de l’Afrique orientale (Kenya, Rhodésie) et a pénétré au Zaïre et même au Cameroun. À partir de 1958, elle envahit le bassin du Nil, et dès 1959 se rencontrait au Soudan, de Juba à Khartoum.
La Jacinthe d’eau a donc envahi la quasi-totalité des régions paléo-tropicales, avec une vitesse stupéfiante, grâce à son formidable pouvoir de reproduction par stolons. Une plante peut produire un nouvel individu en deux semaines. On a calculé en Louisiane que 10 plantes mères peuvent donner 655 360 nouvelles plantes en une saison de végétation (soit du 15 mars au 15 novembre), et il est certain que la Jacinthe d’eau se multiplie toute l’année dans les régions tropicales proprement dites. Ce fait explique l’envahissement rapide et total des eaux, qui se trouvent entièrement obstruées par ces associations monophytiques.
Cette plante décorative, qui fit la joie des amateurs, est responsable de ruptures d’équilibre par suite de la compétition avec des plantes aquatiques autochtones qui se trouvent refoulées ou éliminées. Par ailleurs elle crée de graves préjudices à la navigation, gêne les pêcheries et entraîne la raréfaction des poissons par modification des frayères.
Devant cette menace, divers moyens de lutte ont été préconisés (De Kimpe, 1957). La destruction mécanique (par bandes transporteuses qui enlèvent les Jacinthes et les rejettent sur les berges ou les entraînent vers des broyeurs) se révéla fort décevante. On entreprit alors une vaste campagne de destruction par pulvérisation d’herbicide, notamment 2,4-D. À partir de 1955, on arriva à un contrôle de l’extension de la Jacinthe d’eau et à son éradication d’une partie de son aire de distribution. Mais le coût des opérations est très élevé. Quelques foyers épars peuvent subsister, points de départ d’une nouvelle invasion172.
L’histoire de la naturalisation de cette espèce végétale montre combien une introduction inconsidérée, à première vue inoffensive, peut être dangereuse. Comme l’a dit Robyns (1955) : « Les ruptures d’équilibre biologique qui en résultent peuvent modifier entièrement la flore et la faune autochtones, en détruire certains composants et jeter même la perturbation dans l’existence normale des habitants. » Cet exemple doit être médité par tous ceux qui projettent des acclimatations dont on ne peut pas prédire les conséquences.
Remarquons enfin que l’homme a contribué par le transport de diverses souches végétales à fabriquer de nouvelles « espèces », notamment en provoquant l’hybridation d’espèces autochtones avec des espèces voisines importées. Si plusieurs cas sont connus en Amérique du Nord, le meilleur exemple se rencontre en Europe dans le cas d’une Graminée halophile, Spartina Townsendii, hybride naturel entre une espèce européenne, Sp. maritima, et une espèce nord-américaine, Sp. alternifolia, importée dès le début du XIXe siècle en Grande-Bretagne173. Cet hybride polyploïde, découvert en 1870, ne commença à se répandre que vers le début de ce siècle ; son expansion en bordure de mer dans les régions marécageuses saumâtres s’accéléra alors aussi bien en Angleterre que dans la partie nord de la France. Cette Spartine ne s’est sans doute pas montrée nuisible à l’homme ; mais son implantation a contribué puissamment à modifier les habitats, surtout en consolidant les étendues de vases mobiles. Elle remplace localement les prairies de Zostères (Zostera) mélangées à des Algues (Enteromorpha entre autres), milieu d’élection des Bernaches cravants Branta bernicla, dont ces plantes constituent la nourriture de base. Le changement des conditions écologiques dû à l’introduction et à la propagation de cette Spartine a donc été éminemment préjudiciable aux Bernaches, aux effectifs déjà réduits, comme à de nombreux Canards et petits échassiers (Ranwell, 1962). On ne peut non plus manquer de signaler les dangers que présenterait certainement l’introduction, le long des côtes européennes, de l’Algue géante Macrocystis pyrifera. Cette espèce, voisine de nos Laminaires et originaire de la côte orientale du Pacifique et des mers australes, a des frondes pouvant atteindre 65 m de longueur. Si son acclimatation, en vue de disposer d’algues capables de fournir en abondance tous les produits que l’on tire de ces végétaux, réussissait, les risques seraient grands de modifier l’équilibre écologique de nos côtes, notamment en ce qui concerne les poissons, et d’occasionner de sérieuses perturbations à la pêche et à la navigation côtière.
2. Un Mollusque nuisible : l’Achatine
Les Achatines appartiennent à un groupe de Mollusques pulmonés voisin des Escargots, riche en espèces et réparti dans les régions tropicales de l’Ancien Monde ; particulièrement bien diversifiées en Afrique où elles habitent surtout les forêts humides, quelques-unes d’entre elles peuplent le sud-est de l’Asie, la Malaisie et l’Indonésie. Une seule retiendra notre attention, Achatina fulica, distribuée originellement à travers l’Afrique orientale depuis l’Abyssinie jusqu’au Mozambique : il s’agit d’une espèce de grande taille dont la coquille atteint 13 cm et le corps étendu 20 cm (fig. 42). Volontiers arboricole, elle grimpe aux arbres où elle se nourrit, surtout dans son jeune âge, de pousses et de bourgeons. Si peu de dégâts ont été signalés dans son aire d’origine, il n’en est pas de même dans les régions tropicales où elle a été introduite, volontairement ou non, par l’homme (Mead, 1961).
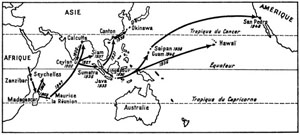
Fig. 42. Achatine d’Afrique orientale Achatina fulica.
Ces acclimatations sont pour certaines déjà anciennes. Sans aucun doute non indigène à Madagascar, où sa présence remonte à la nuit des temps, cette Achatine fut pour la première fois signalée hors d’Afrique à l’île Maurice, en 1803, puis vers 1821 à La Réunion : le gouverneur de l’époque l’avait fait venir de Madagascar et l’élevait dans son jardin, son épouse ayant un faible pour le potage à l’escargot qui passait pour guérir les poitrinaires. En 1847 le malacologiste W. H. Benson transporta cette Achatine de Maurice aux Indes, où il la libéra dans le jardin de la Bengal Asiatic Society.
L’homme l’a disséminée ensuite largement à travers les régions tropicales de l’Ancien Monde (fig. 43). On la signala aux environs de 1840 aux Seychelles, aux Comores en 1860, à Ceylan en 1900, à Pérak et en Malaisie en 1928. Ce Mollusque commettait déjà un peu partout des dégâts aux cultures, notamment aux plantations de Thé et d’Hévéa. En 1931 il faisait son apparition en Chine méridionale, en 1935 à Java, en 1936 à Sumatra et au Siam en 1937.
Les Japonais introduisirent délibérément cette Achatine aux îles Mariannes (Saipan et Tinian) en 1938, afin de l’utiliser à des fins culinaires ; elle se répandit ensuite ailleurs dans l’archipel à la faveur du transport de matières végétales (notamment à Guam en 1946). Elle a également pris pied dans de nombreuses autres îles d’Océanie, aux Hawaii (elle y arriva en 1936) où elle paraît cependant peu répandue (sauf à Oahu). Elle a même fait son apparition aux États-Unis en 1947, envahissant certains districts de Californie (probablement transportée à l’état d’œufs ou de très jeunes individus collés à des véhicules militaires rapatriés du Pacifique après la guerre) ; le climat californien ne paraît cependant pas convenir à ce Mollusque propre aux habitats humides des tropiques. Il en disparut rapidement, mais en 1967 fut introduit fortuitement en Floride, à la suite d’une imprudence. L’Achatine s’est répandue à travers le sud-est des États-Unis et menace de s’étendre à travers toutes les parties chaudes de l’Amérique.
Fig. 43. Transport et acclimatation volontaire ou non de l’Achatine Achatina fulica.
D’après R. Tucker Abbott, 1949
Cette Achatine est donc maintenant répandue à travers une large aire dans les parties tropicales de l’Ancien Monde. Elle s’est mise à y pulluler, grâce à son extraordinaire faculté de reproduction174 et à sa grande souplesse écologique. À Ceylan, on observa 227 individus sur un seul tronc de Cocotier. À Java, un colon en récoltait plus de 400 chaque matin dans un petit jardin. Dans les Mariannes, elles sont si nombreuses qu’elles occasionnent des accidents d’autos, les pneus patinant sur les Mollusques écrasés sur les routes.
D’une manière très curieuse, elle a accentué ses tendances arboricoles dans tous les pays qu’elle a conquis. Elle attaque les bourgeons et les pousses d’une longue série de plantes cultivées (surtout Bananiers, Cacaoyers, Papayers, Hévéas, Agrumes et plantes de couverture) et y commet des dégâts qui prennent localement des proportions de catastrophe, en particulier en Malaisie et aux Mariannes. Les planteurs lui reprochent de plus de transporter les spores et les germes de nombreuses maladies végétales (mosaïques par exemple) grâce à la large sole sur laquelle elle se déplace comme nos Escargots.
Devant cette grave menace pour l’agriculture, plusieurs moyens de lutte ont été préconisés, le plus simple étant la collecte systématique des Achatines ainsi mises à prix : par moins d’un demi-million d’adultes et quelque 20 millions d’œufs ont ainsi été récoltés à Singapour, sans pour autant faire disparaître l’espèce de l’île. Les appâts empoisonnés n’ont guère eu plus de succès.
Les biologistes ont alors tenté la lutte biologique. Les populations d’Achatines paraissent en équilibre en Afrique, du fait de l’action limitante d’ennemis naturels qui font défaut dans les aires où elles ont été introduites. Des missions scientifiques furent envoyées au Kenya pour y rechercher les ennemis naturels de ce Mollusque. À part quelques Insectes, elles en ramenèrent surtout 2 Mollusques Gastéropodes carnassiers de la famille des Streptaxidae : Gonaxis kibweziensis et Endentulina affinis. Une première tentative fut faite dans l’île d’Agiguan, dans les Mariannes. Introduits en 1950, les Gonaxis avaient détruit 20 % des Achatines en 1952, et 60 % en 1954. Devant le succès de cette expérience, celle-ci fut renouvelée dans la plupart des îles du Pacifique, et notamment aux Hawaii. Des quelque 30 espèces qui y furent introduites, 5 se montrèrent réellement efficaces et plus particulièrement Gonaxis quadrilateralis. Celui-ci détruit les pontes et les jeunes Achatines et n’hésite pas à s’attaquer à des adultes de 7 à 10 cm. Aux Hawaii, les Mollusques déprédateurs ont diminué dans une proportion de 80 % et l’état de leurs populations qui ne comprennent plus que des individus de grande taille montre que celles-ci ne se renouvellent plus. Notons qu’un autre Mollusque carnivore originaire de Floride, Englandina rosea, contribue lui aussi au contrôle des Achatines. Les tentatives faites avec d’autres espèces, et notamment Gonaxis kibweziensis, ont été abandonnées.
D’autres prédateurs on fait l’objet d’essais analogues, et notamment des insectes : Carabidae (Tefflus d’Afrique orientale) et Lampyridae (Lamprophorus de l’Inde). Leur action est nettement moins efficace.
On dispose donc maintenant d’un moyen de lutte biologique, qui, joint à l’usage bien compris de molluscicides, permet, sinon d’éradiquer, du moins de contrôler les populations d’un Mollusque particulièrement dangereux pour les cultures et l’équilibre naturel (Abbott 1949, 51 ; Mead, 1961 ; Petitjean, 1966).
3. Poissons et autres animaux aquatiques
Depuis fort longtemps, l’homme s’est efforcé de transporter d’innombrables poissons hors de leur aire de distribution naturelle. Certains de ces essais ont été couronnés d’un plein succès et ont permis de résoudre des problèmes alimentaires en mettant à la disposition des populations humaines des ressources appréciables en protéines animales. Mais d’autres n’ont pas été sans entraîner une modification profonde des équilibres biologiques, au détriment d’espèces autochtones parfois plus intéressantes sur le plan économique (Vibert et Lagler, 1961). De véritables succès ont sans aucun doute été enregistrés, telle l’acclimatation de diverses Truites dans les rivières de montagne d’Extrême-Orient ou d’Afrique orientale, des Saumons en Nouvelle-Zélande et des Aloses (Alosa sapidissima) sur la côte pacifique de l’Amérique du Nord ; ces succès s’expliquent par le fait que ces espèces semblent « préadaptées » aux milieux où on les a introduites, y occupant des niches écologiques demeurées vacantes, sans élimination d’espèces autochtones et sans prolifération exagérée (fig. 44).
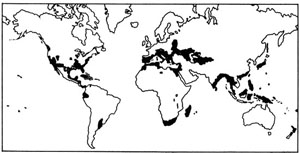
Fig. 44. Distribution des Gambusies Gambusia affinis à travers le monde. L’aire d’origine est marquée par des hachures. Partout ailleurs ce poisson a été introduit par l’homme en vue du contrôle des Moustiques dont les larves aquatiques constituent sa nourriture. Bien que bénéfique dans l’ensemble, cette acclimatation a provoqué en quelques points des ruptures d’équilibre consécutives à la pullulation. D’après Krumholz, Ecol. Monogr. 18, 1948.
L’introduction et l’élevage intensif du Tilapia, Cichlidé originaire d’Afrique, maintenant objet de pratiques piscicoles très prospères en Extrême-Orient, sont également à considérer comme largement bénéficiaires. Ce poisson, véritable providence des régions tropicales carencées en protéines animales, n’a dans l’ensemble pas eu d’impact dans les communautés naturelles, car il n’est guère sorti des milieux artificiels, étangs et rizières, où l’homme l’a confiné. Son établissement à l’état sauvage est en revanche susceptible d’entraîner de profondes répercussions sur les équilibres, ne serait-ce qu’en raison de son extraordinaire pouvoir de multiplication.
Mais dans d’autres cas l’acclimatation peut mener à une véritable pullulation qui engendre le nanisme de l’espèce introduite – donc manque de rentabilité économique –, l’élimination d’espèces autochtones par compétition ou prédation, et même la transformation des habitats.
Fig. 45. Crabe chinois Eriocheir sinensis. La carapace peut atteindre des dimensions de 9 sur 7,5 cm.
Un exemple de prolificité excessive est fourni par notre Carpe dont l’introduction en Amérique du Nord a failli tourner à la catastrophe. Un stock originel de 345 individus amené en 1876 aux États-Unis et élevé dans des établissements de pisciculture fut répandu à travers le pays. Il y devint la souche d’une population démesurée qui se multiplia aux dépens d’espèces plus intéressantes, probablement par suite de sa rusticité et d’une meilleure utilisation de la nourriture. Les Carpes modifient de plus les milieux originels en détruisant la végétation aquatique et en troublant l’eau par la boue soulevée par leurs activités. Certaines étendues d’eau ont été ainsi entièrement privées de leur végétation par ces poissons responsables d’un « surpâturage aquatique ». La Carpe a par ailleurs été introduite dans le sud-est du Brésil, notamment dans le rio Iguazu, s’attirant la vindicte des pêcheurs privés de poissons appréciés, éliminés par un compétiteur plus entreprenant. Il en est de même en Afrique du Sud (Bigalke, 1937).
L’Europe a été elle aussi le théâtre d’acclimatations malheureuses, en particulier celle du Poisson-chat Ameiurus nebulosus, introduit en France en 1870. Renommé pour la délicatesse de sa chair, il est en réalité difficile d’accommoder pour la table en raison de ses nageoires piquantes ce poisson qui prend la place d’espèces plus intéressantes. Il en va de même de la Perche soleil Eupomotis gibbosus, acclimatée dès 1886 en France et installée maintenant dans les eaux du pays presque entier, aux dépens d’espèces dont elle n’a de loin pas la valeur alimentaire.
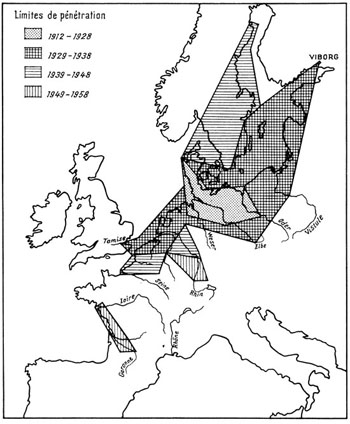
Fig. 46. Extension du Crabe chinois Eriocheir sinensis en Europe de 1912 à 1958. S’il a par ailleurs pénétré en Méditerranée, il semble avoir disparu de Suède et de Finlande où seuls quelques spécimens isolés ont été signalés dans les temps récents. D’après Hoestlandt, 1959.
Nous ne pouvons manquer de rapporter ici l’introduction sans doute involontaire du Crabe chinois Eriocheir sinensis, signalé pour la première fois en 1912 en Allemagne du Nord, sur un affluent de la Weser, et amené accidentellement d’Extrême-Orient, voyageur clandestin dans les ballasts emplis d’eau d’un bateau naviguant sur son lest (fig. 45). De là, ce Crabe thalassotoque (il passe sa vie en eau douce, mais se reproduit dans les eaux saumâtres et salées) se répandit à partir de 1923 sur une bonne partie de l’Europe, grâce à son tempérament et à ses déplacements périodiques entre les eaux douces et salées. En fait l’espèce pullule175 maintenant dans une vaste zone, de la mer Baltique au bassin de la Gironde et même en Méditerranée. Il a pénétré loin à l’intérieur des terres, puisqu’on le trouve le long de l’Elbe et de la Moldau jusqu’à Prague (fig. 46).
Du fait de son extrême prolificité, ce Crabe s’est montré très nuisible en modifiant les habitats aquatiques (destruction des plantes submergées), en entrant en compétition avec certains poissons176 (concurrence alimentaire), en minant les berges et les digues et provoquant des éboulements (éminemment fouisseurs, ces Crabes creusent des terriers et des galeries atteignant 80 cm de profondeur et 12 cm de diamètre ; on en a compté jusqu’à 30 au m3) et gênant la pêche (ils endommagent les filets et obturent les nasses). Aucun moyen de contrôle n’a encore donné des résultats satisfaisants, en dépit de captures massives réalisées grâce à des barrages et à des fosses lors de ses migrations : sur une rivière près de Berlin, 2,5 millions d’individus furent recueillis en 5 mois ; près de Magdebourg, 355 t furent collectées en 1932 dans l’Elbe. Cette destruction s’est montrée inopérante et aucun moyen n’a encore été découvert pour endiguer son extension. Ce Crabe aura donc contribué à détruire l’équilibre naturel des eaux douces d’Europe tout en causant de graves préjudices à l’homme (André, 1947).
Si les introductions de Poissons végétariens ou planctonophages ne sont pas exemptes de dangers, celles-ci sont beaucoup plus graves dans le cas des espèces carnivores dont certaines peuvent devenir de redoutables prédateurs dans le milieu où ils ont été imprudemment libérés177.
C’est notamment ce qui s’est passé dans les rivières et les lacs des étages tempérés et montagnards des Andes où furent introduits plusieurs Salmonidés nord-américains. Or ces eaux pauvres en poissons sont peuplées de Siluridés du genre Trichomycterus et de Cyprinodontidés du genre Orestias (fig. 47), différenciés en un nombre assez considérable d’espèces, adaptées chacune à un mode de vie différent, notamment au point de vue des spécialisations alimentaires (les unes sont végétariennes, d’autres se nourrissent strictement de Crustacés, d’autres de petits Mollusques). Il s’agit de groupes ayant littéralement « éclaté » dans leur évolution, et occupant une série de niches écologiques demeurées vacantes par suite de la réduction des souches établies sur les hauts plateaux andins.
Fig. 47. Orestias agassizi du lac Titicaca. D’après Neveu-Lemaire, Lacs des hauts plateaux de l’Amérique du Sud, Paris, 1906.
Cette remarquable expérience naturelle est actuellement gravement menacée par l’introduction des Salmonidés prédateurs qui font disparaître les espèces autochtones devenues leurs proies. Plusieurs espèces sont au bord de l’extinction. Si l’intérêt économique de cet acclimatement n’apparaît pas encore clairement, la catastrophe scientifique est en tout cas manifeste.
Un exemple similaire se rencontre à Célèbes où, en 1939, on découvrit de la manière la plus inattendue un poisson siluroïde, Clarias batrachus, inconnu jusqu’alors à l’est de Java et de Bornéo. L’enquête révéla qu’un particulier en avait rapporté quelques individus de Java et les avait libérés dans les eaux célébésiennes, en raison de son goût pour ce poisson qui lui rappelait sa jeunesse. Il s’agit en réalité d’un dangereux prédateur qui se nourrit de frai et qui risque de causer des ravages parmi des poissons de grande importance économique. Il n’a pu être éradiqué de Célèbes où il constitue donc une menace permanente (Schuster, 1952).
L’homme est parfois indirectement l’artisan de l’introduction de certains poissons. Le meilleur exemple est certainement celui de la Lamproie Petromyzon marinus dans les Grands Lacs américains (East 1949). On sait que ce « poisson » bizarre – membre d’un groupe de Vertébrés très particuliers, celui des Agnathes – se livre à des migrations régulières qui le mènent de la mer, où il passe la majeure partie de sa vie vers les rivières où il se reproduit. Dans le passé, s’il s’était établi naturellement dans le lac Ontario et dans quelques lacs de l’est des États-Unis (Finger Lakes), les fameuses chutes du Niagara constituaient jusqu’alors un obstacle majeur à son extension. En 1829, on aménageait le canal Welland pour permettre à la navigation de contourner les chutes ; avec un retard de 100 ans, la Lamproie sut en profiter, pénétra dans le lac Erié, sans toutefois y proliférer, puis ne tarda pas à faire irruption dans la rivière Saint-Clair (en 1930), dans les lacs Huron et Michigan (en 1937) et jusque dans la partie occidentale du lac Supérieur (en 1946). Cette invasion prit dès lors l’allure d’une véritable explosion. Les Lamproies se multiplièrent rapidement dans les rivières s’écoulant vers les grandes étendues d’eau et commencèrent à exercer leurs ravages dans ces véritables mers intérieures, où la pêche représente une ressource économique de quelque 12 millions de dollars. Pas moins de 90 % des Truites grises Salvelinus namaycush178 ramenées par les pêcheurs portaient de graves morsures de Lamproies, qui s’attachent à leurs proies et les dévorent vivantes. Pour chaque victime ayant survécu aux attaques de ces dangereux carnivores, des centaines et peut-être des milliers meurent sur le coup sans jamais être décelées par l’homme. Le stock de Truites et de Corégones Coregonus clupaeformis, objets de la pêche commerciale, a donc régressé dans des proportions catastrophiques, par suite d’une prédation à laquelle ces populations n’étaient pas adaptées. La production totale des eaux des lacs Huron et Michigan appartenant aux États-Unis tomba de 8,6 millions de livres de Truites à moins de 26 000 livres. Une industrie prospère périclitait donc par suite d’une introduction dont l’homme est indirectement responsable, ayant créé les voies de migration par lesquelles avaient passé les prédateurs.
Cette introduction si désastreuse provoqua la création d’un comité chargé d’étudier les moyens de combattre le fléau (Sea Lamprey Committee, organisé par le Fish and Wildlife Service des États-Unis, en collaboration avec les organismes similaires du Canada). Une vaste campagne fut entreprise et l’on mit à l’essai divers procédés de destruction des prédateurs : arrêt par grilles électriques des reproducteurs se rendant vers leurs frayères dans les rivières au cours rapide ; destructions des larves par toxiques (notamment usage de roténones qui tuent souvent la totalité des poissons d’un cours d’eau) ; usage de pièges spéciaux. L’invasion n’est cependant de loin pas arrêtée ni même limitée à l’heure actuelle. Après l’essai de plus de 6 000 produits, on semble néanmoins disposer maintenant de certaines substances sélectives capables d’exterminer les Lamproies sans causer de dommages aux poissons.
Cet exemple montre à quel point un acte à première vue sans importance – l’aménagement d’un canal navigable – peut avoir d’influence sur l’équilibre de la nature et créer un problème grave pour la conservation des espèces et la protection d’une richesse naturelle dont vivent des milliers d’hommes.
4. Transport et acclimatation d’Insectes
Les Insectes sont manifestement les animaux terrestres dominants à l’époque actuelle. Certains vivent en association avec l’homme ; d’autres, bien plus nombreux, sont liés à ses cultures et à ses industries. Il n’y a de ce fait même rien d’étonnant à ce que l’homme ait assuré volontairement ou non le transport de beaucoup d’entre eux à travers le monde.
Le nombre et la variété des insectes transportés ont beaucoup augmenté dans les temps récents du fait de la fréquence accrue des échanges commerciaux, des plus grandes facilités de voyages et aussi de l’accélération de ceux-ci, rendant possible le transport d’animaux vivants auxquels la lenteur des voyages ne permettait pas d’arriver autrefois à bon port.
Sur un bateau chargé de riz, venant de Calcutta et de Rangoon et se rendant à Cuba, pas moins de 42 espèces d’Arthropodes (parmi lesquels des Bostryches, des Ténébrionides, des Curculionides et des Pyrales) ont été rencontrées, ayant survécu à des fumigations et à des désinsectisations répétées (Myers, 1934). Les services sanitaires des États-Unis ont découvert des insectes très variés à bord de 28 852 avions parmi les 80 716 qui furent inspectés de 1937 à 1947. Ces données permettent à elles seules de juger de l’importance du transfert d’insectes par les moyens de transport modernes, et aussi de leurs chances de s’établir, en dépit de la surveillance de services d’inspection spécialisés, notoirement insuffisants dans beaucoup de pays et en tout cas incapables d’autre chose que de limiter parmi les insectes le nombre de « passagers et d’immigrés clandestins »179. Comme dans le cas des végétaux, l’homme a, volontairement ou non, créé de nouvelles communautés d’Arthropodes, au détriment d’espèces autochtones et surtout au préjudice de ses propres intérêts.
Cet impact dans la faune est surtout évident dans les régions insulaires, où l’équilibre naturel a été profondément déplacé. D’après Zimmermann (1948), sur 5 000 espèces d’insectes signalées aux îles Hawaii, pas moins de 1 300 ont été amenées par l’homme. La présence de la plupart de ces éléments exogènes est consécutive à des acclimatations involontaires, notamment du fait du transport de produits alimentaires (riz, céréales, farine) ou de matières végétales (y compris des matériaux d’emballage) ; un nombre relativement réduit a été acclimaté volontairement, notamment en vue de leur emploi dans la lutte biologique. Beaucoup des insectes introduits ont été à l’origine de la disparition ou du moins de la régression massive de certaines populations d’insectes autochtones, principalement dans les régions de basse et moyenne altitude. Une Fourmi, Pheidole megacephala, est en particulier à incriminer en tant que prédateur ; presque ubiquiste en dehors des régions montagneuses encore couvertes de forêts, elle a fait disparaître la plupart des insectes endémiques vis-à-vis desquels elle se comporte comme un véritable fléau. Parmi les Hyménoptères Euménides, les Odynerus indigènes, dont les populations se comptaient par millions, ont à peu près disparu par suite de la compétition avec certains Hyménoptères Ichneumonides introduits, qui les ont privés des chenilles de Lépidoptères qu’ils parasitent. La lutte paraît particulièrement inégale entre espèces endémiques et espèces introduites du fait de la spécificité parasitaire des premières et de la grande souplesse des autres.
Ces deux cas ne sont pas uniques aux Hawaii, dont la faune entomologique a régressé dans une notable proportion par suite de la prédation et de la compétition avec les espèces introduites180 (en plus bien entendu de la destruction des habitats et de la végétation indigène que nous n’avons pas à envisager ici). Ces faits se sont reproduits dans beaucoup d’autres parties du monde, où l’acclimatation a modifié profondément l’équilibre des populations d’insectes autochtones et sans aucun doute fait disparaître certaines espèces.
Un autre problème intéressant directement l’homme est celui des insectes se comportant comme des « pestes » vis-à-vis des cultures. De nombreux insectes ont profité du déplacement d’équilibre consécutif à la mise en culture et de la quantité supplémentaire de nourriture mise ainsi à leur disposition pour pulluler et causer de graves dégâts aux récoltes, soit sur pied, soit stockées en attendant leur consommation. Il est à remarquer que beaucoup des insectes nuisibles appartiennent à des espèces introduites. En 1939 déjà, Smith faisait remarquer qu’aux États-Unis, parmi 183 espèces préjudiciables aux cultures, pas moins de 81, soit 44 %, avaient été importées par l’homme. Celles-ci semblent particulièrement aptes à devenir des fléaux économiques, leur pullulation n’étant pas limitée du fait de l’absence de leurs prédateurs et compétiteurs naturels.
Pour les Européens, le meilleur exemple est celui du Doryphore Leptinotarsa decemlineata, Coléoptère originaire de l’ouest des États-Unis, du Colorado au Mexique. Ce parasite des Solanées vivait sur des espèces sauvages (Solanum rostratum entre autres) et se trouvait en équilibre avec son environnement. Quand la mise en valeur de l’Amérique du Nord gagna vers l’ouest et que l’extension des cultures de Pommes de terre (Solanum tuberosum) eut atteint les limites de l’aire naturelle du Doryphore, il se produisit un transfert de cet insecte vers l’espèce cultivée. Il montra aussitôt un extraordinaire potentiel d’adaptation et se mit à pulluler d’autant plus que les prédateurs qui en limitaient les effectifs sur les plantes sauvages ne le suivirent pas sur la plante cultivée.
À partir de 1859, il commença à se répandre vers l’est et avait atteint la côte orientale des États-Unis vers 1874, à la suite d’une extension somme toute naturelle. De là il fut transporté par l’homme à travers l’Atlantique. En 1876, un premier foyer d’infestation européen fut stoppé en Allemagne, de même que plusieurs tentatives de colonisation ultérieures. Mais en 1920, son implantation dans la région de Bordeaux ne put être endiguée et c’est de là que le Doryphore envahit toute la France (1935), puis le reste de l’Europe. Seules les îles Britanniques ont pu être protégées de son envahissement par les services de protection des végétaux. Les dégâts ne purent être réduits qu’au prix d’une lutte chimique onéreuse, sans graves conséquences toutefois sur l’équilibre de la faune. L’histoire du Doryphore montre comment une plante importée du Nouveau Monde par l’homme, transformée en Europe par les pratiques culturales, puis réexportée vers l’Amérique, a favorisé l’extension naturelle d’un parasite hors de son habitat d’origine, puis dans toute l’aire de culture de la plante nourricière après transport artificiel à travers l’Atlantique (Elton, 1958).
La liste des espèces d’insectes ainsi acclimatées est fort longue ; le mécanisme d’extension, aussi varié soit-il dans le détail, se ramène au fond toujours au même schéma général. L’irruption dans un nouveau milieu est presque toujours suivie d’une pullulation effrénée ; la population peut ensuite se stabiliser à un niveau plus bas, sans toutefois cesser de poser des problèmes qui restent graves dans bien des cas et exigent une lutte coûteuse à l’aide d’insecticides, eux-mêmes générateurs de ruptures d’équilibre.
Si l’introduction d’insectes porte surtout de graves préjudices à l’agriculture, elle a aussi parfois de profondes répercussions sur le plan médical. Involontairement, l’homme a transporté des insectes vecteurs de maladies épidémiques, à l’origine de catastrophes lourdes de conséquences. Le meilleur exemple en est sans aucun doute celui de la malaria dans le nord-est du Brésil (Soper et Wilson, 1943).
Vers la fin de 1929 ou au début de 1930, un aviso français arrivait de Dakar à Natal, porteur du courrier. Il amenait avec lui des passagers clandestins : quelques Anophèles de l’espèce Anopheles gambiae, Moustique répandu sur une bonne partie de l’Afrique et un des deux principaux vecteurs de la malaria. Ces Moustiques jouissent de ce triste privilège du fait qu’ils sont très étroitement adaptés à l’homme. Contrairement à d’autres espèces d’Anophèles, et en particulier à ceux qui habitent naturellement le nord-est du Brésil. Ils se tiennent de préférence à l’intérieur des habitations humaines et leur cycle tout entier se déroule au voisinage de celles-ci ; leurs larves peuvent se développer dans de minuscules collections d’eau, même des débris de vaisselle comme on en trouve partout autour des maisons181.
Ces Anophèles s’acclimatèrent immédiatement et dès mars 1930, on collectait 2 000 larves à Natal, d’où ils se répandirent dans les environs de la ville. Dès la fin d’avril la malaria était devenue d’une manière inattendue un problème sérieux à Natal. En janvier 1931, 10 000 cas étaient signalés parmi les 12 000 habitants d’un faubourg ouvrier de Natal (Alecrim) et l’épidémie gagnait les districts environnants.
Puis, de 1932 à 1937, la malaria sembla régresser. Il y eut une période de silence consécutive à l’éradication d’Anopheles gambiae à Natal. Cet insecte s’était cependant établi dans d’autres localités, et dès 1937 on le signalait dans une partie des États de Ceara et de Rio Grande do Norte (fig. 48). En 1938, une terrifiante épidémie de malaria s’abattait sur la région avec un taux de mortalité très élevé comme dans toute forme épidémique ; les cas se chiffraient par centaines de mille et l’on estime que plus de 20 000 habitants succombèrent à la maladie182. Les conséquences furent catastrophiques sur le plan matériel ; la vie économique tout entière fut paralysée dans les régions contaminées, qui se rangent parmi les plus pauvres du monde. Incapables de travailler, les habitants qui vivent au jour le jour se trouvèrent subitement dans un état de complet dénuement. Selon un auteur brésilien (P. A. Sampaio) : « Comme des hordes de Huns sanguinaires les Moustiques avançaient laissant une traînée de deuil et de destruction dans leur sillage. »
Devant cette catastrophe, le gouvernement fédéral brésilien commença une vaste campagne de lutte avec l’aide de la Rockefeller Foundation.
La progression du Moustique fut d’abord stoppée ; puis on commença à l’éliminer de la zone contaminée : en novembre 1940, il était entièrement éradiqué du Brésil. Cette lamentable expérience avait coûté des milliers de vies humaines, un peu plus de misère pour une population au niveau de vie déjà très bas et plus de 2 millions de dollars.
Toute l’Amérique avait néanmoins été protégée de ce fléau, car l’Anopheles gambiae aurait vraisemblablement pu s’étendre sur une aire allant du sud des États-Unis au nord de l’Argentine. Cette catastrophe risque cependant de se répéter dans le futur, ne serait-ce qu’en raison de l’augmentation rapide du trafic entre l’Afrique et l’Amérique et de la difficulté du contrôle sanitaire et entomologique des moyens de transport, notamment des avions.
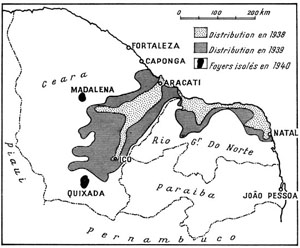
Fig. 48. Distribution du Moustique africain Anopheles gambiae dans le nord-est du Brésil. Cet insecte s’est répandu dans la vallée du Jaguaribe, de l’Assu et de l’Apodi, aux conditions écologiques particulièrement favorables ; en revanche sa progression vers l’ouest et le sud a été gênée par des zones peu propices à son établissement. D’après Soper et Wilson, 1943.
Cette acclimatation involontaire démontre une fois de plus le danger de l’irruption dans une partie du monde d’un animal de la taille d’un minuscule insecte, transporté d’une région où il est en relatif équilibre avec son milieu, vers une région où il risque de provoquer une catastrophe.
La fièvre jaune offre un exemple similaire en Amérique du Sud, où elle est arrivée en même temps que son principal vecteur, un Moustique encore, Aedes aegypti ; son origine est vraisemblablement à chercher en Afrique d’où elle a cheminé avec les convois d’esclaves noirs. Ces moustiques se sont acclimatés en Amérique tropicale et vivent au voisinage des habitations, entretenant ainsi la fièvre jaune urbaine. Mais des Moustiques autochtones ont été infectés à leur tour par le virus, notamment des Haemagogus ; ils sont susceptibles d’entretenir une forme sylvatique de la fièvre jaune, très répandue à travers les forêts du Bassin amazonien et même au-delà, de nombreux Singes servant de réservoir de virus. La fièvre jaune importée dans le Nouveau Monde, où elle est transmise par un insecte lui aussi acclimaté, a opéré un transfert vers des animaux autochtones et peut dorénavant se transmettre par un cycle ne faisant appel qu’à des représentants de la faune indigène.
L’aspect pathologique des acclimatations ne doit pas être négligé, le transport accidentel d’une souche d’insectes vecteurs pouvant être lourd de conséquences sur le plan de l’hygiène publique.
5. L’Étourneau et quelques autres envahisseurs ailés
L’Étourneau Sturnus vulgaris, notre vulgaire Sansonnet, figure parmi les oiseaux les plus entreprenants du monde ailé. Largement répandu à travers la région paléarctique, il y est d’ailleurs en pleine extension, sans doute en rapport avec les transformations apportées par l’homme aux biotopes originels, sa fécondité élevée, ses facilités d’adaptation et son anthropophilie marquée.
Il a été acclimaté dans diverses parties du monde, en particulier en Amérique du Nord, en Afrique du Sud, en Australie et en Nouvelle-Zélande, où il n’a pas manqué de provoquer des ruptures d’équilibre au sein des populations d’oiseaux autochtones, tout en commettant des dégâts dans les cultures, se révélant parfois une véritable plaie dont il est difficile, voire impossible, de se débarrasser.
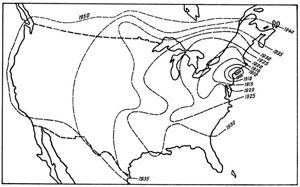
Fig. 49. Extension de l’Étourneau Sturnus vulgaris en Amérique du Nord. Les lignes interrompues marquent les limites approximatives de distribution aux dates indiquées. D’après Wing, 1943.
L’envahissement de l’Amérique du Nord est particulièrement spectaculaire (fig. 49). Après plusieurs essais d’acclimatation infructueux, 60 individus furent lâchés en 1890 et 40 en 1891 à Central Park, en plein New York. Pendant 6 ans, l’Étourneau resta confiné au district urbain new-yorkais ; puis ses populations augmentant, il se répandit à travers le nord-est des États-Unis. Dès lors son extension commença vers l’ouest et le sud. En 1926, il avait atteint la vallée du Mississippi, et partout son établissement était suivi d’une prolifération de l’espèce, dont les vagues partaient ensuite coloniser les zones demeurées vierges.
À l’heure actuelle l’Étourneau a envahi la totalité des États-Unis, jusqu’à la côte pacifique où il a niché dès 1959 en Californie. Il est établi par ailleurs dans le nord du Mexique, au Canada jusqu’aux rives de la baie d’Hudson et même récemment en Alaska.
L’établissement de l’Étourneau à travers l’Amérique du Nord a donc présenté les caractères d’une véritable explosion ; en quelque 70 ans, cet oiseau a presque peuplé un continent. En 1943, ses populations étaient estimées à environ 50 millions d’individus, soit environ 1 % des populations nicheuses totales des États-Unis. Toutes proviennent d’un stock initial d’une centaine d’individus, ce qui démontre l’extraordinaire prolificité de cet oiseau et sa grande souplesse d’adaptation.
En Afrique du Sud, l’Étourneau a été introduit dès 1899, par Cecil Rhodes en personne, au Cap, d’où il s’est répandu rapidement dans les districts avoisinants. En Australie, 36 individus ont été lâchés dans le Victoria en 1863 ; ils ont fait souche en se multipliant rapidement et en envahissant tous les districts habités, sauf apparemment l’ouest du continent. En Nouvelle-Zélande, 17 individus ont été importés aux environs de 1862 ; puis de nouveaux contingents ont été lâchés au cours des années ultérieures par les soins de sociétés d’acclimatation et de particuliers. L’Étourneau s’est multiplié au point d’envahir toute la Nouvelle-Zélande, et même des îles aussi éloignées que Kermadec et Macquarie.
Le cas de l’Étourneau montre bien comment un oiseau actif, capable de s’adapter à toutes les circonstances, peut faire irruption dans des communautés naturelles après son introduction artificielle par l’homme. Les conséquences sont très variées. Insectivore, il est incontestablement utile, surtout au moment de sa nidification, en consommant une grande quantité de larves et d’insectes, mais ses ravages, surtout dans les vergers, les vignobles et même les champs (semences à la germination) sont parfois très importants.
L’Étourneau a par ailleurs eu une action manifeste sur les communautés d’oiseaux autochtones, sa compétition en limitant les effectifs. Il occupe les sites de nidification et accapare la nourriture du fait d’une dominance très nette. En Nouvelle-Zélande notamment les effectifs de Pipits, de Martins-pêcheurs et de Tuis Prosthemadera novae-hollandiae ont considérablement diminué en partie en raison de sa concurrence.
Les mêmes faits se sont reproduits avec un autre Sturnidé, le Martin triste, « Merle des Molluques » ou Mina Acridotheres tristis, originaire d’une large zone allant du Turkestan à la péninsule indochinoise, et introduit dans nombre de régions chaudes du globe, depuis l’Afrique du Sud jusqu’au Pacifique. Comme son cousin, ce compétiteur peut être rendu responsable de la raréfaction de certaines espèces autochtones, en particulier en Nouvelle-Calédonie (Barrau et Devanchez, 1957).
À l’île Maurice où pas moins de 23 espèces aviennes ont été acclimatées, la plupart asiatiques (Carié, 1916), un cas similaire est offert par le Bulbul Orphée Pycnonotus jocosus, originaire de l’Inde, de la Chine et de la péninsule indochinoise. Six individus introduits en 1892 ne tardèrent pas à se multiplier au point que, 8 ans plus tard, leur descendance occupait déjà une aire de quelque 100 km2 et que dès 1910 elle avait envahi presque toute l’île où ces oiseaux pullulent à l’heure actuelle. En plus des déprédations que causent ces Passereaux frugivores aux cultures (fruits, légumes délicats, Caféiers) ils attaquent et évincent les oiseaux autochtones, par exemple les Oiseaux à lunettes Zosterops curvirostris, devenus ainsi rares et cantonnés maintenant aux forêts épaisses.
6. Le Lapin à l’assaut des continents
Le Lapin Oryctolagus cuniculus, animal d’apparence si sympathique et si utile à beaucoup de points de vue, est un des fléaux de l’humanité à laquelle il a coûté en définitive beaucoup plus cher qu’il n’a rapporté.
Ce Rongeur duplicidenté, originaire de la région méditerranéenne et principalement d’Espagne et des îles de la Méditerranée occidentale183, semble avoir été importé en Italie vers 230 avant J.-C., ainsi qu’en Grèce où seul le lièvre était connu des auteurs classiques. C’est au cours du Moyen Âge qu’il fut acclimaté en Europe moyenne et septentrionale (il n’est signalé d’une manière certaine qu’à partir du XIIIe siècle en Angleterre) ; d’ailleurs il n’aurait pu prospérer en dehors de la région méditerranéenne avant que les défrichements et la déforestation ne créent les habitats ouverts qui lui sont nécessaires.
Ses populations augmentèrent rapidement comme l’attestent de nombreux témoignages historiques. Bien installé dans toute l’Europe, le Lapin ne donne plus l’impression d’y être un intrus acclimaté par l’homme au départ d’une aire de répartition originellement très réduite.
Cette invasion de l’Europe sous la protection et l’aide directe ou indirecte de l’homme ne constitue néanmoins que la première étape de la conquête de vastes espaces à travers le monde ; le Lapin est sans doute le Mammifère terrestre ayant la répartition la plus diversifiée (fig. 50), mis à part bien entendu les Rats et Souris devenus commensaux de l’homme.
De toutes les introductions tentées à travers le monde, c’est en Australie qu’elles ont le plus « brillamment » réussi. Les premiers Lapins y arrivèrent dès 1787, sans donner naissance à des populations importantes, Ceux qui peuplent actuellement le continent descendent de 24 individus sauvages amenés d’Angleterre sur le clipper Lightning et libérés près de Geelong, dans l’État de Victoria en 1859. Les Lapins ont réussi à coloniser les deux tiers de l’Australie, s’adaptant à des conditions écologiques très variées, y compris celles du semi-désert. Ils y sont devenus le plus grand fléau de ce continent sur le plan scientifique comme sur le plan économique.
En Nouvelle-Zélande, après plusieurs essais infructueux, le Lapin fut introduit aux environs de 1864-1867, puis à plusieurs reprises au cours des décennies suivantes. Il commença à pulluler vers 1874 et a envahi depuis tous les habitats favorables de l’île du Sud, et une bonne partie de l’île du Nord où les facteurs climatiques et écologiques lui sont toutefois moins propices ; là aussi il est devenu un véritable fléau.
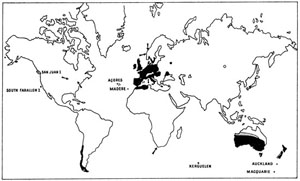
Fig. 50. Distribution du Lapin Oryctolagus cuniculus à la suite des introductions par l’homme. Les flèches marquent l’emplacement de foyers limités. La zone entourée de pointillés en Sibérie indique un lieu d’introduction d’où le Lapin semble avoir disparu. Enfin en Australie, la zone hachurée n’est peuplée que par une population clairsemée.
Le Lapin (forme domestique) a été introduit par ailleurs dans la partie chilienne de la Terre de Feu, aux environs de 1910, et dès 1947 y était devenu une plaie. Il s’est répandu sur le continent sud-américain, aussi bien au Chili qu’en Argentine. En dépit du danger, ce Rongeur a été libéré en Amérique du Nord, sur quelques îles au large de l’État de Washington (San Juan Islands) et de la Californie. Il risque maintenant d’être acclimaté accidentellement sur le continent lui-même sous le nom fallacieux de « San Juan rabbit », des contingents ayant été envoyés notamment dans l’Ohio, le Wisconsin et la Pennsylvanie. Notons par ailleurs que le Lapin a été introduit en Afrique du Sud sur un îlot près de Cape Town (Robben Island). Il est à souhaiter que ces tentatives ne réussissent pas, car l’Amérique du Nord et l’Afrique pourraient être dans le futur les théâtres d’invasions aussi désastreuses qu’en Australie184.
On ne saurait oublier les introductions malheureuses du Lapin dans certaines îles subantarctiques, et notamment à Auckland, à Macquarie et à Kerguelen. Dans cette dernière île, les souches domestiques libérées par l’expédition anglaise du « Passage de Vénus » en 1874 ont vite pullulé. Ces Rongeurs sont responsables d’une très grave érosion du sol et de la disparition presque complète du Chou de Kerguelen Pringlea antiscorbutica, Crucifère très caractéristique de Kerguelen qui ne se maintient que sur les îlots et les falaises inaccessibles au Rongeur ; cette disparition a entraîné la réduction dramatique d’une communauté d’Invertébrés hautement spécialisés.
Si le Lapin se trouve maintenant largement dispersé à travers le monde entier (fig. 50), c’est en Australie et en Nouvelle-Zélande que son introduction a été la plus malheureuse ; la véritable « explosion » de ses populations eut comme conséquences une série de catastrophes à l’échelle de continents entiers. Le succès de cette acclimatation s’explique par la grande souplesse écologique du Lapin et par son extraordinaire pouvoir de reproduction, dont la renommée n’est pas usurpée ; une femelle, en âge de se reproduire à 15 semaines, est susceptible de donner naissance à 8 portées de 6 petits en moyenne par an.
Mais le succès du Lapin s’explique surtout par son irruption dans les territoires dépourvus de prédateurs et de compétiteurs susceptibles d’en limiter le nombre : les mammifères indigènes d’Australie sont en majorité des Marsupiaux à première vue au moins incapables de lutter contre un rival aussi bien armé. La pullulation du Lapin dépasse en fait tout ce que l’imagination peut concevoir : pas moins de 428 millions de peaux ont été exportées d’Australie de 1945 à 1949, ce qui donne une idée de l’importance des populations.
Les mêmes faits se sont reproduits en Nouvelle-Zélande comme le montre notamment l’augmentation des exportations de peaux de Lapin : 33 000 en 1873 ; plus d’un million en 1877 ; plus de 9 millions en 1882 ; et environ 17,6 millions en 1945.
Ces Rongeurs constituent un véritable fléau grevant l’économie de l’Australie et de la Nouvelle-Zélande. Ils ravagent la végétation en prélevant des quantités considérables de Graminées, entrant de ce fait en compétition avec les Moutons, principale richesse agricole de ces pays. Si l’on tient compte du fait que de 7 à 10 Lapins consomment autant d’herbe qu’un Mouton, les Lapins exportés en 1945 de Nouvelle-Zélande équivalent à eux seuls à 2 millions de Moutons, dont la vente aurait produit un revenu de plus de 2 millions de livres, au lieu d’environ 1,3 million rapportées par la vente des peaux de Lapin, soit une perte nette d’environ 700 000 £. Or ces chiffres ne concernent que les Lapins exportés, et non pas l’ensemble de leurs populations. Ces animaux coûtent certainement des millions de livres chaque année à la Nouvelle-Zélande. En Australie le cheptel ovin et bovin pourrait être augmenté de 25 % si ce Rongeur disparaissait. Dans la partie occidentale de la Nouvelle-Galles du Sud la capacité-limite des terres a baissé de 50 % depuis son introduction, ce district comptant plus de 15 millions de moutons en 1891 et moins de 7 millions en 1951.
Il y a plus grave encore, car le Lapin détruit systématiquement la végétation, non seulement graminéenne, mais aussi arbustive et arborescente dans ses formes jeunes (il est grand consommateur d’écorce). Modifiant le couvert végétal, qu’il appauvrit, il est à l’origine d’une érosion d’autant plus redoutable qu’il mine les sols de ses terriers. Son introduction a été la cause principale de la dégradation des habitats, perte irrémédiable pour l’Australie comme pour la Nouvelle-Zélande, intraduisible en chiffres.
L’influence du Lapin sur la faune australienne est elle aussi de la plus extrême gravité. Ce Rongeur a en effet modifié profondément l’équilibre naturel et a éliminé tous les Marsupiaux herbivores par suite de la compétition185.
La nocivité du Lapin provoqua la mise en application en Australie d’une série de mesures destinées à favoriser la lutte contre ce fléau186. Quelques-unes furent particulièrement malheureuses, comme l’introduction du Renard d’Europe. Ce Carnivore au régime alimentaire très éclectique se multiplia rapidement et, s’il préleva quelques Lapins, il contribua surtout à la raréfaction, voire à l’extinction de beaucoup de Marsupiaux.
On tenta aussi d’endiguer la progression du Rongeur en aménageant des barrières en grillage, dont certaines se rangent parmi les plus longues du monde entier. Telle est en particulier celle qui, édifiée de 1902 à 1907, rejoignait Port Hedland sur l’océan Indien à Hopetown sur la côte méridionale, longue de 2 150 km et destinée à mettre à l’abri l’Australie occidentale et ses terres de culture. D’autres s’étendaient en Australie orientale pour tenter de limiter l’extension vers le nord. Ces barrières, dont l’étendue totale fut d’environ 11 000 km, ne réussirent malheureusement pas à contenir le Lapin, soit qu’elles aient été construites trop tard, soit que le Rongeur ait réussi à les franchir en profitant de brèches.
Divers procédés de destruction furent par ailleurs mis en œuvre : battues organisées, furetage, enfumage, gaz toxiques et surtout empoisonnement par appâts mélangés de phosphures, de strychnine et d’arsenic (ces procédés contribuèrent d’ailleurs à la destruction de beaucoup de Marsupiaux tentés par les appâts). Le fléau ne put être endigué en dépit de ces efforts, qui coûtèrent une fortune à l’Australie et à la Nouvelle-Zélande (de 1943 à 1945, le coût de ces opérations se chiffrait annuellement à environ 170 000 livres néo-zélandaises).
En 1950, on eut l’idée de répandre une maladie épizootique qui décimait les Lapins domestiques au Brésil : la myxomatose, due à un virus spécifique, sans danger pour l’homme187. Après plusieurs tentatives infructueuses, les Australiens réussirent à contaminer une très vaste aire, l’extension de la maladie impliquant un transport du virus par divers parasites et par les moustiques. L’épizootie atteignit d’énormes proportions, et l’on estime qu’elle tua les quatre cinquièmes des Lapins du sud-est de l’Australie. La mortalité fut de 99,5 % dans certains cas. Le résultat était presque miraculeux.
C’est ce qui donna l’idée d’introduire la myxomatose en Europe, où en 1952 on procéda en France, dans l’Eure-et-Loir, à un lâcher de Lapins contaminés. L’épizootie se propagea rapidement à travers toute l’Europe occidentale et y détruisit une proportion très importante de Lapins. Cette initiative fut d’ailleurs suivie d’une polémique véhémente, car, en France notamment, le Lapin, bien que nuisible à la sylviculture et aux cultures agricoles, a une très grande importance cynégétique.
Nous n’insisterons pas sur la propagation de cette maladie qui pour la première fois a conduit à un contrôle effectif de populations de Lapins. En éliminant virtuellement ce Rongeur sur de très vastes surfaces, elle a permis de mesurer l’impact de celui-ci dans la nature (voir à ce sujet les rapports de la 6e réunion de l’UICN, Édimbourg, 1956). L’aspect le plus spectaculaire concerne la végétation. En Australie, des régions auparavant désertiques se couvrirent de végétation, et pour les années 1952-1953, l’augmentation de la production agricole est évaluée à 50 millions de livres. En Europe, le paysage végétal changea notablement, en particulier en France où l’on assista dans les forêts dégradées à une repousse de la végétation arbustive et à des semis naturels de diverses essences forestières, partout en voie de régénération.
Peut-on considérer pour autant le péril du Lapin écarté ? Sans doute pas, car l’extermination n’est jamais totale. Dans les années qui suivirent l’introduction de la myxomatose en Australie on observa une diminution du taux de mortalité, due à une atténuation du virus, et surtout à la sélection de souches de Lapins réfractaires à la maladie. Il est probable que l’on s’achemine vers un équilibre entre les Lapins et une maladie qui persiste à l’état endémique dans les régions infectées. Sauf si une souche de Lapins complètement réfractaires est sélectionnée et se lance à nouveau à l’assaut des zones désertées à l’heure actuelle, des mesures de contrôle très attentives devraient suffire à conserver un juste équilibre entre le Lapin et son environnement et à maintenir ces déprédateurs à un niveau supportable188.
Il est à souhaiter que l’on parvienne à ce résultat qui préserverait les intérêts de tous189. Sinon le gentil Lapin redeviendra le fléau qu’il a été à travers le monde, en partant d’une zone très réduite de la Méditerranée occidentale, d’où l’a transporté l’homme inconscient des facultés destructrices de cet animal d’aspect si débonnaire.
7. Les Mammifères, alliés ou complices de l’homme
Du fait de leur grand intérêt économique, les Mammifères ont fait l’objet de très nombreuses tentatives d’acclimatation à travers le monde (De Vos, Manville et Van Gelder, 1956). On ne compte pas moins de 200 espèces dont la naturalisation a été tentée avec des succès très divers190, se rangeant principalement parmi les Lagomorphes, les Rongeurs, les Carnivores et les Ruminants.
Certaines de ces acclimatations ont été accidentelles, comme c’est avant tout le cas de celles des Rats et des Souris transportés par l’homme partout à travers le monde, au point que leur engeance est maintenant ubiquiste. Leur impact dans les communautés naturelles est considérable. Ils ont ravagé le couvert végétal et se sont comportés comme de dangereux prédateurs. Les lamentables exemples des Râles du Pacifique, éliminés en grande partie par les Rats, et des oiseaux de mer des îles antarctiques, décimés par le même fléau, sont là pour nous permettre de juger de l’importance de ces Rongeurs. Notons qu’ils ont de même éliminé les compétiteurs autochtones, comme on le constate notamment aux îles Galapagos dont les Cricétidés endémiques ont disparu partout où se sont établis les Rats (Brosset, 1963).
D’autres introductions ont été délibérées, comme celles des animaux-gibier ou des animaux à fourrure. On peut y ajouter l’acclimatation d’animaux domestiques redevenus sauvages, s’intégrant dès lors au sein des communautés biologiques et y provoquant des ruptures d’équilibre.
Beaucoup de ces acclimatations ont échoué. D’autres en revanche n’ont que trop bien réussi, les animaux introduits étant devenus rapidement des « pestes ». Il convient cependant de ne pas porter de jugements généraux, car les conséquences d’une introduction varient souvent largement à l’échelle locale.
C’est notamment le cas du Rat musqué Ondatra zibethica, Rongeur Microtiné (Campagnols) de la taille du Lapin, originaire d’Amérique du Nord, aux mœurs rappelant un peu celles du Castor. Sa valeur comme animal à fourrure détermina son introduction en Europe, où il fut acclimaté pour la première fois en 1905 en Tchécoslovaquie aux environs de Prague. En 1914, toute la Bohême était déjà colonisée par des effectifs estimés à 2 millions. Puis la Bavière fut envahie, la progression de l’animal étant d’environ 50 à 70 km par an, principalement le long des systèmes hydrographiques offrant des conditions écologiques favorables à ce Rongeur aquatique. En 1933, quelque 200 000 km2 étaient occupés par le Rat musqué en Europe moyenne.
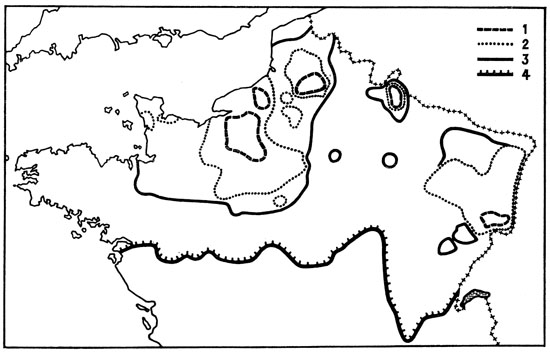
Fig. 51. Progression de l’aire de répartition du Rat musqué Ondatra zibethica en France. 1. Situation en 1932 ; 2. Situation en 1951 ; 3. Situation en 1954 ; 4. Situation en 1963. D’après Dorst et Giban, 1954, complété.
En 1930, le Rat musqué, élevé en France dans des fermes d’élevage depuis 1920 environ, fit son apparition dans une douzaine de foyers répartis dans la moitié nord du pays. Quelques-uns de ceux-ci s’éteignirent immédiatement, d’autres au contraire s’étendirent, notamment en Normandie, dans la Somme, les Ardennes et l’Alsace. À l’heure actuelle, le Rat musqué, en voie d’extension, occupe une large aire dans le nord de la France (fig. 51) ; il fait par ailleurs partie intégrante de la faune mammalienne d’une vaste partie de l’Europe, de la France à la Russie.
Il a aussi été acclimaté aux environs de 1929 dans les îles Britanniques où des colonies s’établirent de l’Écosse au sud de l’Angleterre. Une campagne d’éradication à grande échelle fut organisée et en 1937 on pouvait estimer qu’il avait été extirpé des îles Britanniques.
Le Rat musqué fut enfin introduit en Finlande (dès 1922), en Suède (1944) et surtout en URSS, où les acclimatations qui remontent à 1927 furent renouvelées à maintes reprises aussi bien en Russie qu’en Sibérie où les lâchers furent effectués en de multiples points (Lavrov, 1960) (fig. 52).
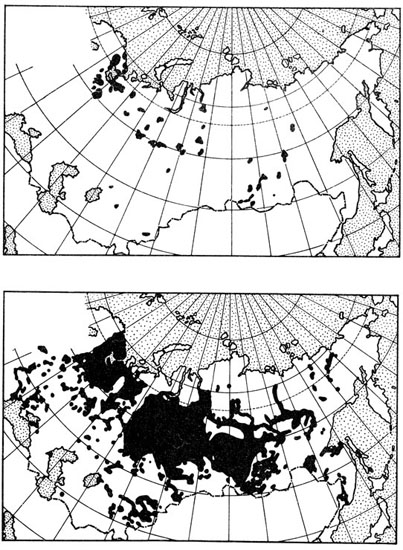
Fig. 52. Extension du Rat musqué Ondatra zibethica introduit en URSS. En haut : situation en 1936 ; en bas : situation en 1956. D’après Lavrov, 1960.
Ce Rongeur est donc maintenant établi dans toute l’Eurasie, de la France au Kamtchatka, et sa distribution est devenue holarctique grâce à l’intervention de l’homme. Son impact dans la nature a été jugé de manière très diverse. En Europe occidentale, il est considéré comme une véritable peste. Il creuse en effet des terriers de grandes dimensions dans les berges et les digues qui se trouvent très dangereusement minées. De plus, il semble volontiers compléter son régime végétarien par des proies animales, certains individus devenant même franchement carnivores. On lui reproche de ce fait de commettre des dégâts parmi les poissons et les Crustacés dulçaquicoles. Son influence sur les populations de poissons est cependant beaucoup plus importante par la transformation des habitats (colmatage des étendues d’eau, transformation des eaux courantes en marécages). Ces dégâts, nullement compensés par les revenus de la vente de la fourrure, font que dans cette partie de son nouvel habitat l’homme lutte à grands frais contre son extension et sa multiplication exagérée, sans espoir de parvenir à son éradication.
Il n’en est pas de même en URSS et en Finlande où ce Rongeur ne semble pas avoir été nuisible aux habitats. Cela tient sans doute au fait que la nature a été moins bouleversée par l’intervention humaine et que le Rat musqué y tient une place un peu comparable à celle que lui a assignée la Nature. La « résistance » du milieu y est plus grande et les dommages aux industries humaines plus réduits du fait du maintien d’un équilibre voisin de l’état originel. Cela explique pourquoi les autorités soviétiques assurent la propagation et l’exploitation rationnelle du Rat musqué devenu une richesse naturelle. En 1954, environ 649 000 peaux ont été collectées en URSS.
Ces variations dans l’impact d’un animal introduit dans un nouvel habitat montrent combien il faut mesurer le jugement porté sur une acclimatation. Elles s’expliquent par des facteurs écologiques sur lesquels nous aurons l’occasion de revenir.
Notons par ailleurs que l’Europe a été colonisée par le Ragondin Myocastor coypus, grand Rongeur semi-aquatique dont l’aire de répartition s’étend sur l’Amérique du Sud, du sud du Brésil à la Patagonie. Introduit dès 1926 comme animal à fourrure, il s’est établi dans les îles Britanniques, en France, aux Pays-Bas, au Danemark, en Allemagne, en Suède, en URSS et même au Japon. Le Ragondin ne s’est pas multiplié à la même vitesse que le Rat musqué, dont il n’a de loin pas la nocivité. Il est même considéré comme utile par les pisciculteurs, car il faucarde la végétation aquatique devenue envahissante, nettoie les surfaces d’eau dormante couvertes d’une végétation dense et augmente la productivité des étangs (accélération de la minéralisation des vases) (Ehrlich, 1959). Cette action n’est toutefois bénéfique que si les effectifs de Ragondins sont bas, faute de quoi on aboutit à une destruction de l’habitat191.
En revanche, l’introduction en Angleterre192 de l’Écureuil gris Sciurus carolinensis fut des plus malheureuses et se solde par de graves préjudices aux forêts et par une rupture d’équilibre aux dépens de l’Écureuil européen Sciurus vulgaris, qui régresse devant l’envahissement de son rival américain (Shorten, 1954).
L’Écureuil gris, originaire de l’est des États-Unis, fut introduit dès 1876 dans le Cheshire, puis dans différentes parties du sud de l’Angleterre entre 1890 et 1929. Il ne tarda pas à se répandre à partir des divers foyers initiaux, et, dès 1930, il occupait une aire d’environ 26 000 km2, surtout dans le sud-est de l’Angleterre ; l’extension était partout rapide, avec des régressions temporaires dues à des épizooties (coccidiose) ou à des hivers rigoureux. En 1945, tout le sud de l’Angleterre était envahi, la progression de l’Écureuil américain n’étant arrêtée que par des fleuves difficilement franchissables, des barrières écologiques (landes et zones marécageuses dépourvues d’arbres) et les districts industriels. À l’heure actuelle l’Écureuil gris occupe la majeure partie de l’Angleterre, une partie de l’Écosse et une large aire en Irlande, où il a été introduit en 1913 (fig. 53). Partout il se multiplie d’une manière très rapide.
L’Écureuil gris est considéré comme une vraie peste par les biologistes britanniques, dont certains n’hésitent pas à affirmer que seul le Surmulot le dépasse en malfaisance. Sans doute tous les Écureuils sont quelque peu nuisibles aux forêts en attaquant les arbres qu’ils écorcent et dont ils endommagent le cambium, infligeant de sévères blessures par lesquelles pénètrent les Champignons pathogènes. De plus ils se montrent nuisibles aux pépinières et aux jeunes plantations. Mais l’Écureuil gris se montre particulièrement malfaisant, en témoignant d’une prédilection affirmée pour l’attaque des arbres ; sa nocivité est de plus décuplée par une densité plus forte que celle de l’Écureuil européen.
L’invasion de l’Angleterre par ce Rongeur a provoqué dès 1931 l’organisation d’une campagne de destruction à l’échelle nationale, renforcée par diverses mesures législatives (par exemple le Grey Squirrel Order de 1947). La lutte est entreprise par tir, piégeage, appâts empoisonnés et destruction des nids. En dépit de l’importance des moyens mis en œuvre, il est peu probable que plus d’un tiers de la population soit exterminé chaque année, ce qui ne permet pas d’espérer l’éradication de l’Écureuil gris, tout au plus son contrôle.
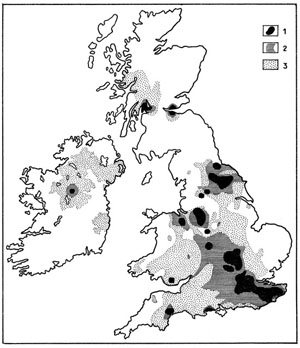
Fig. 53. Extension de l’Écureuil gris Sciurus carolinensis dans les îles Britanniques. 1. Aire occupée aux environs de 1920 ; 2. Aire occupée en 1930 ; 3. Aire occupée en 1952. D’après Shorten, 1954, modifié.
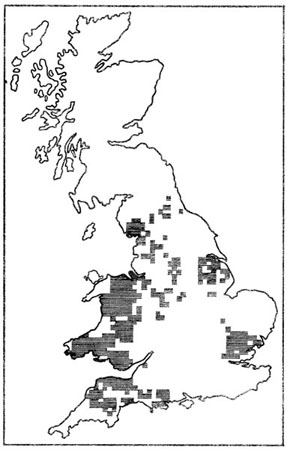
Fig. 54. Réduction de l’aire de distribution de l’Écureuil européen Sciurus vulgaris en Angleterre (la carte ne mentionne pas la répartition en Écosse), d’après le résultat des enquêtes menées de 1945 à 1952. Remarquer par comparaison avec la figure 53 que cette espèce a pratiquement disparu de l’aire peuplée par l’Écureuil gris. D’après Shorten, 1954, modifié.
L’introduction et l’extension de l’Écureuil américain à travers les îles Britanniques ont exercé une profonde influence sur l’équilibre des populations d’Écureuils européens, disparaissant progressivement de toutes les zones envahies pour des causes très partiellement connues (fig. 54). Certains ont affirmé qu’il y avait compétition et lutte entre les deux espèces, l’Écureuil introduit n’hésitant pas à attaquer et à tuer l’Écureuil autochtone ; il ne s’agit sans doute que de cas isolés. La compétition alimentaire n’est pas non plus flagrante, pas plus qu’un taux de reproduction favorisant l’intrus. La concomitance des deux phénomènes est cependant trop parfaite pour qu’ils ne soient pas liés d’une manière quelconque. Il suffit que deux espèces manifestent des différences écologiques à première vue minimes pour que l’équilibre se déplace au profit de la mieux armée. Une sensibilité différente aux maladies, une meilleure métabolisation des ressources alimentaires, une dominance, même peu affirmée, dans les comportements, une plus grande rapidité des populations à « récupérer » après une période défavorable suffisent à expliquer de telles ruptures d’équilibre.
En revanche l’Écureuil européen paraît mieux adapté que son cousin américain à la vie dans les forêts de Conifères ; en fait il s’est maintenu dans le nord de son habitat, notamment en Écosse où l’espèce introduite ne s’est pas étendue avec la même vitesse que dans le Sud, où elle a surtout colonisé les forêts de feuillus (elle montre une prédilection marquée pour les hêtres). Il est possible qu’un nouvel équilibre s’établisse à la faveur de l’enrésinement des forêts qui se manifeste en Angleterre comme partout ailleurs en Europe. Il s’agit en tout cas d’une expérience fort intéressante que l’homme paye chèrement sur le plan économique.
Nous ne ferons qu’une brève mention aux autres acclimatations de Mammifères à travers le monde. Certaines, relatives à des animaux-gibiers, ont été couronnées de succès, comme celle du Cerf Sika Sika nippon en France et en Angleterre où il s’est fort bien adapté au milieu et se montre moins destructeur que le Cerf élaphe. Son acclimatation s’est néanmoins faite au détriment de ce dernier et l’on peut se demander si même dans le cas de telles introductions l’intérêt n’est pas plutôt d’accroître les populations d’espèces autochtones.
D’autres acclimatations concernent des Carnivores. L’introduction de Mangoustes, notamment dans les Antilles, a été évoquée précédemment, en même temps que les véritables catastrophes qu’elle a entraînées pour la faune endémique. Le Vison américain Mustela vison échappé de fermes d’élevage en Scandinavie et en Islande y a commis des déprédations parfois graves. L’introduction du Vison et du Raton laveur Procyon lotor dans quelques îles au large de la côte pacifique du Canada (Lanz Island, Cox Island), réalisée en 1938, a entraîné dès 1950 une multiplication de ces prédateurs qui font des ravages au sein des riches colonies d’oiseaux marins. Cet exemple doit être médité par ceux qui projettent d’acclimater le Vison dans les îles subantarctiques ; si cette introduction réussissait, elle aurait de grandes chances de ruiner rapidement les colonies reproductrices d’oiseaux de mer, Manchots entre autres, qui en constituent la plus belle parure.
Notons enfin que l’introduction de Mammifères apparentés à des formes autochtones est parfois suivie d’une fâcheuse hybridation des deux stocks de populations. Plusieurs formes du genre Capra ont été introduites à partir de 1901 dans les Tatras, en particulier le Bouquetin C. ibex, la Chèvre sauvage C. aegagrus et le Bouquetin de Nubie C. nubiana ; une invraisemblable hybridation en a résulté, qui a mené la population tout entière à une ruine totale, notamment en raison d’un dérèglement de la période de reproduction, les jeunes naissant en plein cœur de l’hiver. En Tchécoslovaquie, le Chevreuil de Sibérie Capreolus pygargus s’est hybridé avec le Chevreuil autochtone C. capreolus. Beaucoup de femelles de ce dernier, saillies par des brocards sibériens de plus grande taille, sont mortes au cours de parturitions rendues très difficiles par la taille des fœtus à terme. Ces conséquences parfois inattendues doivent rendre encore plus méfiant dans le domaine de l’acclimatation, d’autant plus qu’elles peuvent se produire dans le cas d’animaux appartenant à d’autres groupes que les Mammifères193.
8. La Nouvelle-Zélande dévastée par les animaux introduits
Les exemples que nous venons d’évoquer ont tous trait à des cas spécifiques. Il convient parallèlement, en prenant une unité géographique, de voir comment l’introduction de multiples souches animales et végétales peut y provoquer de graves ruptures d’équilibre. L’exemple de la Nouvelle-Zélande semble particulièrement bien désigné pour mesurer l’impact d’une série d’acclimatations mal comprises au sein d’une même biocénose.
Sans doute des facteurs très complexes sont-ils intervenus dans la dévastation de ces îles dont la faune et la flore comprennent une grande proportion d’endémiques, évolués du fait de l’isolement géographique. La mise en valeur à des fins agricoles et pastorales a certes entraîné la disparition d’une vaste partie des habitats primitifs par action directe de l’homme : au XIVe siècle, avant l’arrivée des Maoris, on estime que la majeure partie du pays, soit 25 millions d’hectares, était boisée ; les forêts sont maintenant réduites à 5,8 millions d’hectares, le reste ayant été converti en pâturages ou en terres cultivées.
Mais l’impact humain s’est aussi manifesté par l’introduction de multiples espèces végétales et animales exogènes. La Nouvelle-Zélande constitue à ce point de vue le meilleur exemple de rupture d’équilibre fragile d’une faune insulaire sous l’influence de compétiteurs et de prédateurs étrangers.
Comme de nombreuses régions insulaires, la Nouvelle-Zélande est à l’état naturel d’une très grande pauvreté floristique et faunistique. Sa flore hautement spécialisée se groupe principalement en deux types d’associations : des savanes herbeuses et des forêts humides (formées d’essences très particulières, tels les Podocarpus, Dacrydium, Metrosideros et Nothofagus). Quant à la faune, elle ne comprenait à l’origine aucun Mammifère, hormis deux Chiroptères (un Rat, Rattus exulans, et le Chien ont été amenés par les Maoris venus de Polynésie centrale au XIVe siècle). Les oiseaux comprenaient des espèces hautement spécialisées, en particulier des formes aptères, dont les Dinornis ne sont que les plus connus avec les Kiwis ou Apteryx.
Une région aussi pauvre sur le plan faunistique ne pouvait donc que tenter ceux qui voulaient y acclimater des animaux, soit pour recréer leur home lointain, soit pour en accroître les ressources naturelles (Clark, 1949 ; Wodzicki, 1950-1961 ; Murphy, 1951 ; Elton, 1958 ; Howard, 1964).
Les premiers essais remontent au capitaine Cook, qui dès 1774 et 1777 acclimata le Chou, la Betterave, la Pomme de terre, ainsi que la Chèvre, le Mouton et le Porc, dans le but de ravitailler en vivres frais les équipages des navires faisant relâche.
L’impact de l’homme s’accentua au siècle suivant, quand la Nouvelle-Zélande devint une colonie de peuplement. La préoccupation essentielle des colons fut alors d’introduire le plus grand nombre possible d’animaux en accord avec la tendance si répandue partout à travers le monde à cette époque. Les mesures législatives traduisent bien cet état d’esprit194, de même que la fondation de nombreuses sociétés d’acclimatation. Si au début du XXe siècle ce zèle manifesta un ralentissement sérieux et si l’on commença à prendre les premières mesures visant à la protection de la faune indigène et à la lutte contre les animaux introduits devenus envahissants, on se trouvait déjà en face d’une situation très sérieuse.
Jusqu’en 1950, environ 53 espèces de mammifères et 125 espèces d’oiseaux avaient fait l’objet de tentatives d’acclimatation, parmi lesquelles respectivement 34 et 31 avaient réussi. Près de la moitié des mammifères et 72 % des oiseaux ont été introduits entre 1860 et 1880. L’Europe venait largement en tête quant à l’apport faunistique et la proportion est encore beaucoup plus forte dans les réussites.
Parmi les mammifères ayant réussi à s’acclimater en Australie figurent 6 Kangourous, un Opossum, le Hérisson d’Europe, l’Hermine, la Belette, le Furet, le Lapin, le Lièvre, le Thar, le Chamois, le Cerf élaphe, le Cerf Axis, le Cerf Sambar, le Wapiti, le Cerf Sika, le Daim, le Cariacou de Virginie et l’Élan. Il faut y ajouter les animaux domestiques redevenus sauvages. Parmi les oiseaux figurent la Bernache du Canada, le Cygne noir, le Canard colvert, le Faisan de chasse, le Colin de Californie, la Perdrix Chukar, l’Alouette des champs, la Grive musicienne, le Merle noir, l’Étourneau, le Martin triste, le Freux, le Corbeau flûteur Gymnorhina hypoleuca, le Moineau, le Verdier, le Pinson, la Linotte, le Chardonneret et le Bruant jaune.
Cette liste permet à elle seule de présager de l’étendue des dévastations causées par les animaux introduits. Beaucoup de niches écologiques étaient vacantes, ce qui explique la véritable « explosion » de certaines espèces. La compétition et la prédation firent peu à peu reculer les formes autochtones, notamment parmi les oiseaux, les animaux dominants des biocénoses sauvages de la Nouvelle-Zélande195.
Les extensions les plus spectaculaires concernent les grands mammifères. Le Cerf élaphe Cervus elaphus fut introduit pour la première fois en 1851, près de Nelson, dans l’île du Sud ; cette introduction fut suivie de nombreuses autres196, au point que maintenant l’espèce est répartie d’une manière presque continue dans toute la Nouvelle-Zélande (fig. 55). Les habitats créés par l’homme – une alternance de forêts, de taillis et de pâturages – lui conviennent particulièrement. Le Daim Dama dama a une distribution plus circonscrite et ne s’est guère étendu autour des points de libération ; mais sa densité y est extrêmement forte par suite d’une véritable explosion de ses populations (on en a tué jusqu’à 6 000 sur une aire de 8 000 ha).
Le Chamois Rupicapra rupicapra, importé d’Autriche en 1907 et en 1913 (10 individus libérés au mont Cook) s’est répandu tout au long des Alpes de l’île du Sud. Parmi les animaux domestiques redevenus sauvages, la Chèvre et le Porc ont le mieux prospéré en raison de leurs grandes facultés d’adaptation.
L’Opossum Trichosurus vulpecula d’Australie introduit comme animal à fourrure en 1858 s’est immédiatement multiplié avec une terrifiante vitesse ; il est réparti maintenant sur toute la Nouvelle-Zélande à l’exception de quelques zones limitées. Contrairement à ce qui se passe dans son habitat australien, il s’est montré très dévastateur dans les forêts, se nourrissant de jeunes pousses et de bourgeons, et arrivant de ce fait à tuer les arbres en quelques années. Il commet par ailleurs de graves dégâts dans les vergers. L’accroissement de ses populations peut se mesurer au nombre moyen de peaux collectées par porteur de licence d’abattage, qui a passé de 163,8 en 1921-1925, à 299,7 en 1935-1940 et à 597,2 en 1940-1945. Dans certains districts sa densité dépasse 600 individus au km2.
Les importantes populations de mammifères introduits ont eu une profonde influence sur les sols et le couvert végétal. L’Opossum dévaste les strates supérieures, du fait de sa vie arboricole, tandis que les Ongulés et le Lapin dégradent les strates inférieures. Les forêts ont donc régressé, accélérant les progrès d’une érosion déclenchée par la déforestation et le surpâturage dû aux animaux domestiques.
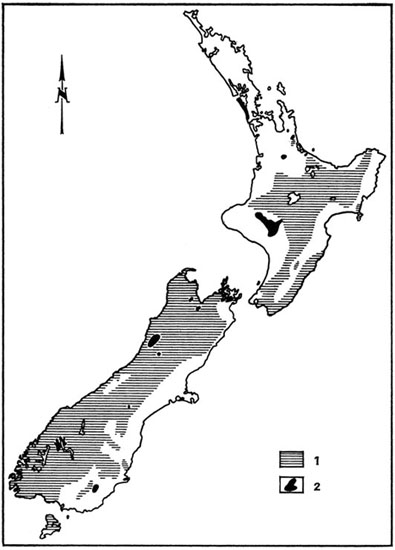
Fig. 55. Distribution actuelle (1) du Cerf élaphe Cervus elaphus et (2) du Daim Dama dama introduits par l’homme en Nouvelle-Zélande. Contrairement au Cerf, le Daim s’est cantonné au voisinage des points de lâcher, quitte à y pulluler. D’après Wodzicki, 1961.
Cet accroissement numérique sans mesure pose de nombreux problèmes pratiques. Après une première phase de protection des espèces introduites, jusqu’aux environs de 1930, on commença à les soustraire à celle-ci, puis à distribuer des primes d’abattage. Des équipes de destruction systématique furent créées, et au cours des années qui suivirent la Seconde Guerre mondiale, les chasseurs professionnels abattirent chacun jusqu’à 40 Cerfs par jour, jusqu’à 2 000 au cours d’un seul été. Entre 1932 et 1954, 512 384 Cerfs furent abattus au total par les chasseurs appointés par le gouvernement. Si l’on tient compte des prélèvements par les particuliers et en se basant sur l’exportation de peaux de Cerfs, on peut estimer qu’au minimum 1,4 million d’individus ont été abattus pendant cette période, mais le chiffre réel est vraisemblablement 2 fois plus élevé.
Les Opossums donnent lieu à une chasse au moins aussi intensive, car en 1945, pas moins de 922 088 ont été prélevés. Nous avons évoqué ailleurs les opérations dirigées contre le Lapin.
Il faut bien entendu tenir compte des ressources économiques tirées de l’exploitation de ces animaux, à inscrire à leur crédit. Mais le déficit reste énorme, surtout si l’on tient compte de la dégradation des terres, dont les effets permanents vont en s’aggravant.
Notons enfin que de nombreux végétaux ont été acclimatés. Allan (1936) les estime à 603 espèces, mais seules 48 entrent véritablement en compétition avec les espèces autochtones. Il faut remarquer que, contrairement aux animaux, les plantes introduites ne prospèrent que sous la protection humaine ou dans des habitats plus ou moins modifiés, soit par l’homme soit par les animaux acclimatés. Il est probable que si l’homme et les animaux introduits venaient à disparaître, les végétaux indigènes reprendraient le dessus et élimineraient la plupart des espèces exogènes197.
Les diverses tentatives d’acclimatation ont donc été des plus préjudiciables à l’équilibre de la nature en Nouvelle-Zélande. Les efforts des colons pour construire un monde vivant à l’image de celui qu’ils avaient connu en Europe ont abouti à une caricature de celui-ci. Ils ont compromis l’avenir de beaucoup d’espèces autochtones et ruiné les habitats. Ils ont créé de graves problèmes économiques consécutifs à la pullulation de certaines espèces. La Nouvelle-Zélande constitue un lamentable exemple de rupture d’équilibre dû à l’introduction d’éléments exogènes multipliés sans frein.
Cet exemple se retrouve dans d’autres parties du globe, surtout dans les régions insulaires, d’un équilibre particulièrement fragile. C’est notamment le cas des îles Hawaii, des Galapagos, de la plupart des Antilles, sans compter les îles subantarctiques où l’introduction de la Souris, du Rat, du Lapin, a tourné à la catastrophe pour la flore et la faune autochtones.
Les quelques exemples d’acclimatations évoqués dans ces pages et choisis parmi beaucoup d’autres montrent avant tout que les importations volontaires ou non d’espèces animales et végétales se soldent par des échecs totaux ou par des proliférations excessives. Hormis quelques remarquables exceptions, la plupart des « réussites » ont tourné au désastre.
Il faut se méfier de l’amateurisme en matière d’acclimatation, comme l’ont prouvé avec éloquence les expériences du siècle passé, où l’homme s’est amusé à transporter les animaux et les végétaux les plus inattendus à travers le monde. Mais il faut se méfier tout autant de l’acclimatation « scientifique ». Sauf dans des cas très particuliers, nos connaissances sur la biologie des êtres vivants sont encore trop rudimentaires pour que nous puissions anticiper sur les conséquences d’une introduction et juger à l’avance de l’avenir des espèces transportées et de leur impact dans la nature.
L’introduction d’un animal ou végétal dans un milieu où il est étranger bouleverse l’équilibre entre les espèces autochtones et crée de nouvelles chaînes alimentaires. Tout l’écosystème se trouve perturbé et est sujet à de graves ruptures d’équilibre. Cela apparaît avec une netteté particulière quand une niche écologique est vacante et que l’espèce introduite peut ainsi rapidement devenir envahissante : le Lapin en Australie, le Cerf en Nouvelle-Zélande, le Crabe chinois en Europe, l’Étourneau aux États-Unis, le prouvent d’une manière éclatante. Cela apparaît aussi quand l’espèce introduite est mieux armée dans la lutte pour la vie, qu’une espèce autochtone écologiquement similaire, alors éliminée ; la concurrence entre le Lapin et les Marsupiaux herbivores en Australie, celle entre la Carpe et les poissons autochtones aux États-Unis ou celle entre l’Écureuil gris et l’Écureuil européen ne constituent que quelques cas probants parmi tant d’autres.
Réalisant son erreur, l’homme tente alors parfois de rétablir l’équilibre en introduisant une espèce prédatrice, principe même de la lutte biologique. Mais si celle-ci a abouti à de remarquables réussites en ce qui concerne les insectes, les échecs ont été particulièrement néfastes dans le cas des Vertébrés : le Chien, introduit aux Galapagos pour lutter contre les Chèvres, a détruit les jeunes Reptiles ; le Chat, introduit dans certaines îles subantarctiques pour contrôler les Rats et les Souris, a ravagé les colonies d’oiseaux ; le Renard introduit en Australie pour combattre les Lapins, a mené beaucoup de Marsupiaux à l’extinction.
Les acclimatations sont ainsi suivies de réactions en chaîne, dont l’homme ne peut prévoir ni le déroulement ni les conséquences. On remarquera que la plupart des catastrophes se sont produites soit dans des îles naturellement appauvries ou dans des régions profondément transformées par l’homme, phénomène explicable par des considérations écologiques brillamment exposées par Elton (1958). Les écosystèmes insulaires sont relativement simples, par suite de la réduction naturelle du nombre des espèces, donc des chaînes alimentaires. Quant aux zones modifiées par l’homme et ses industries, les biocénoses y ont été considérablement appauvries et se trouvent réduites à l’état de véritables caricatures.
Une espèce s’étend et se multiplie en fonction inverse de la résistance que lui offre le milieu. Or la résistance des habitats naturellement ou artificiellement simplifiés est à son minimum. L’espèce introduite peut ainsi y proliférer au-delà de toute limite et devenir rapidement une « peste ». C’est ce qui s’est produit en Nouvelle-Zélande, dépourvue de mammifères, dans les Antilles et dans beaucoup de régions insulaires aux éléments faunistiques très réduits. C’est également ce qui s’est produit en Europe et en Amérique du Nord, continents où de larges superficies ont été entièrement modifiées par l’homme et où les transformations profondes ont abouti à un appauvrissement des milieux naturels, rendus éminemment vulnérables. Les introductions dans des milieux complexes, aux chaînes écologiques innombrables, par exemple la forêt hygrophile, n’ont au contraire que des chances réduites de réussir, et encore moins d’avoir des conséquences dramatiques.
Cela constitue une raison supplémentaire de conserver autant que possible aux habitats un semblant d’équilibre naturel, garant d’une résistance accrue du milieu vis-à-vis d’espèces exogènes arrivées par suite d’une action volontaire ou non de l’homme.
Pillage ou exploitation rationnelle des ressources des mers
Ô mer, nul ne connaît tes richesses intimes…
Charles Baudelaire
Bien que les ressources des mers ne soient pas inépuisables, elles représentent néanmoins une masse énorme dont l’homme tire parti depuis les temps immémoriaux. Les eaux salées prises dans leur ensemble se rangent parmi les milieux naturels ayant la plus haute productivité ; une partie des protéines animales consommées par l’humanité en provient d’ailleurs198.
Le volume de ces ressources vient avant tout de l’énorme surface des mers, qui s’étendent sur 71 % du globe, et de leur richesse exceptionnelle en éléments minéraux parmi lesquels tous les corps simples sont représentés sous forme de sels ou d’ions libres.
La vie y pullule, aucune partie des eaux salées n’en étant dépourvue, bien qu’elle soit surtout concentrée dans les couches superficielles. La strate « utile » est cependant beaucoup plus épaisse que sur la terre : les végétaux marins, en particulier les Algues microscopiques, occupent une épaisseur de quelque 20, parfois 100 m, alors que la couche productive du sol n’atteint que quelques dizaines de centimètres.
La biomasse totale des végétaux et des animaux marins dont on a décrit de véritables galaxies est de ce fait hors de proportion avec les biomasses relevées sur terre. Les eaux marines forment des biocénoses extraordinairement complexes et particulièrement prospères, aux chaînes alimentaires innombrables, des Algues microscopiques et des animalcules du plancton aux géants des mers, poissons, Calmars et Cétacés.
Par ailleurs, en dépit des progrès de la pêche et de son industrialisation, le prélèvement par l’homme est encore peu important en valeur relative. On estime qu’à l’heure actuelle de 60 à 70 millions de tonnes (65,7 millions en 1973) sont pêchées tous les ans dans le monde, contre 21,1 millions de tonnes en 1938, et 33,3 en 1958. En 1973, le tonnage se répartit comme suit en milliers de tonnes métriques :
| Afrique | 4 800 |
| Amérique du Nord | 4 940 |
| Amérique du Sud | 4 260 |
| Asie | 30 240 |
| Europe | 12 530 |
| Océanie | 260 |
| URSS | 8 620 |
Certains secteurs, comme une partie des côtes africaines et les mers de l’hémisphère austral, ne sont encore que très peu touchés par la pêche et ses industries annexes, dont les aires privilégiées restent l’Atlantique et le Pacifique nord.
En dépit de ces faits, les ressources marines sont cependant d’ores et déjà exploitées d’une manière trop intensive en de nombreux points du globe, surtout au voisinage des grandes concentrations humaines ; le perfectionnement des moyens de capture et une demande croissante en aliments tirés de la mer expliquent cette situation qui devient chaque année plus préoccupante.
Il est sans aucun doute a priori impossible de penser qu’une espèce marine puisse être exterminée par suite de la seule action humaine, sauf en ce qui concerne les mammifères : la Rhytine en a fourni un lamentable exemple, et certaines Otaries à fourrure ont été amenées au bord d’une éradication totale. Cela est impensable dans le cas de tous les autres animaux marins, aussi bien poissons qu’Invertébrés. La mer est si vaste et la fécondité des organismes qui y vivent si démesurée qu’une telle probabilité n’est pas à envisager en pratique. Les poissons pondent des millions d’œufs, dont une énorme proportion disparaît même quand la population est en parfait équilibre naturel.
Le danger d’une surexploitation concerne bien plus la rentabilité commerciale des pêcheries, qui risque d’être gravement atteinte quand la pression de la pêche devient trop forte. L’homme constitue un élément entièrement étranger à l’équilibre normal des populations animales marines. Quand des prélèvements sont effectués, un nouvel équilibre s’établit, pas nécessairement défavorable à l’ensemble de la biocénose marine ; bien plus que dans le cas des écosystèmes terrestres, il existe un énorme surplus de populations, éliminé normalement par suite des effets de la prédation et de la compétition, et dans lequel vient puiser l’homme sans dommage pour l’ensemble des stocks. Des prélèvements massifs et répétés risquent en revanche de compromettre la rentabilité commerciale, non point tant en exterminant les reproducteurs, comme dans le cas des animaux terrestres et en particulier des oiseaux et des mammifères, car il en restera presque toujours suffisamment, mais en prélevant des individus n’ayant pas encore atteint une taille suffisante.
Le phénomène est particulièrement bien étudié en ce qui concerne les poissons exploités par les pêcheurs. Dans aucun autre domaine des sciences de la nature on n’a accumulé autant de renseignements concernant la structure et la dynamique des populations ; l’abondance des données provient des multiples études faites en raison de l’importance économique du problème et de l’énorme matériel à la disposition du biologiste et du statisticien.
Sans entrer dans le détail de ces recherches qui ont abouti à de solides connaissances quant à la théorie des pêches, rappelons au préalable plusieurs faits indispensables à la compréhension des lois régissant une exploitation rationnelle des stocks de poissons.
Comme chez tous les animaux, une quantité identique de nourriture assure en grande partie la croissance chez les jeunes, alors que chez les individus âgés elle sert presque uniquement à leur maintien en vie, sans gain de poids appréciable199. Si les poissons âgés dominent, ils accaparent la nourriture au détriment des plus jeunes, dont la croissance se trouve donc ralentie et qui périssent en grand nombre. Un prélèvement de sujets adultes augmente donc les chances de survie des individus plus jeunes, parmi lesquels la croissance est plus rapide, ce qui a pour effet d’augmenter la biomasse totale. On arrive donc à un résultat à première vue paradoxal, mais qui s’explique parfaitement par les lois de la dynamique des populations : dans certaines conditions, une exploitation du stock de poissons par l’homme augmente la biomasse de l’ensemble de la population par suite de la croissance différentielle selon l’âge.
Par suite du prélèvement par l’homme, tant que celui-ci ne dépasse pas un certain seuil, les populations de poissons se trouvent donc dans un meilleur état. D’une manière remarquable, la productivité des districts exploités par la pêche est supérieure à celle des districts « vierges », consécutivement à une modification profonde de la structure de la population, les individus âgés étant éliminés au profit de ceux qui n’ont pas encore entièrement terminé leur croissance200.
Mais au contraire quand le prélèvement par l’homme augmente en importance, il élimine alors des tranches de population dont les individus n’ont pas encore achevé leur croissance. Cela aboutit à une diminution graduelle et accélérée de la biomasse totale : c’est ce que l’on appelle l’overfishing201 (fig. 56).
Fig. 56. Un exemple d’overfishing : le Haddock dans les mers d’Islande. En haut : par suite du développement de la pêche à la Morue (que traduit la courbe ascendante indiquant la durée des opérations de pêche), tous les poissons furent surexploités ; au milieu : fluctuations du tonnage de Haddock débarqué ; en bas : fluctuations des prises par unité d’effort, en baisse accentuée. On remarque que le tonnage diminue en dépit d’un effort de pêche accru, signe caractéristique d’une exploitation exagérée. D’après Graham, 1956.
L’ensemble des études théoriques et pratiques réalisées dans ce domaine montre qu’un prélèvement excessif diminue la productivité et par conséquent le volume de la collecte. Dans un exemple précis, on a montré qu’en prélevant 80 % des individus des différentes classes d’âge, on obtenait un poids de 106 102 unités ; en en prélevant seulement 50 %, ce chiffre s’élevait à 161 138 unités par suite d’une répartition différente des tailles des poissons capturés. Cela signifie que pour un effort de pêche moindre (pêchant à 50 % au lieu de 80 %), la collecte était de moitié plus importante, l’équilibre de la population s’établissant à un niveau plus élevé (Hardy, 1959).
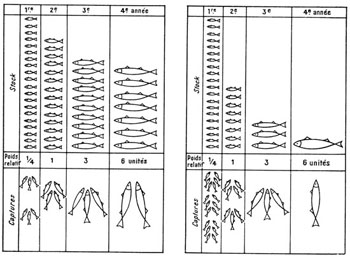
Fig. 57. Effet des efforts de pêche sur les prises de poissons (compte tenu de la mortalité naturelle estimée à 5 %). À gauche : effort de pêche atteignant par an 30 % du stock ; pour le même poids total (soit en tout 26 1/2 unités) on prend plus de gros poissons, ceux-ci ayant eu le temps de croître, et moins de petits ; à droite : effort de pêche atteignant par an 90 % du stock ; pour le même poids total (26 1/2 unités) on prend surtout des poissons de petite taille. Ce schéma montre que l’on a intérêt à laisser les individus atteindre une certaine taille, ce qui permet d’obtenir le même volume de prises avec un effort de pêche moindre.
D’après E. S. Russel, in Graham, 1949.
Comme le montre par ailleurs le classique diagramme publié par Graham (1948) (fig. 57), si l’effort de pêche atteint 30 % du stock par an, on obtient le même poids total de poissons qu’avec un effort de pêche de 90 % – soit 3 fois plus élevé –, par suite de l’élimination des individus de taille moyenne et une prédominance d’individus de petite taille. Dans le dernier cas, on compromet l’avenir de la population et on est sans cesse obligé d’augmenter l’effort de pêche – donc de diminuer la rentabilité de la pêcherie – pour tenter de maintenir le tonnage débarqué au même niveau ; on aboutit rapidement à la ruine du stock exploité.
De multiples définitions ont été données de l’overfishing et certaines font appel à des connaissances approfondies de l’équilibre des populations. La meilleure et la plus pratique précise qu’il y a overfishing chaque fois qu’il y a combinaison d’un effort de pêche accru et d’un rendement moindre.
La rentabilité de la pêche repose donc sur des notions assez subtiles de dynamique des populations et de variations de la biomasse de celles-ci. Le stock de reproducteurs est en règle générale toujours assez important, de même que le nombre de poissons de petite taille, par suite de l’extraordinaire fécondité des poissons et des autres animaux marins. Mais l’homme n’a pas intérêt à prélever les poissons avant qu’ils n’aient atteint leur taille optimale, qui n’est pas leur taille maximale étant donné la dynamique de leurs populations et les lois de leur croissance pondérale202 (fig. 58).
Fig. 58. Captures de Plies en 1/4 d’heure de chalutage en mer du Nord. En haut : en 1938 ; en bas : en 1945. Remarquer que la guerre a permis au stock de se reconstituer et que la taille moyenne des poissons a considérablement augmenté. D’après Graham, 1949.
Fig. 59. Évolution de la pyramide d’âge d’une même population de poissons au cours d’une quinzaine d’années d’exploitation. La disparition des individus de grande taille est due à une surexploitation des classes d’âge supérieures. La diminution de la biomasse proportionnelle à la surface des graphiques est nettement apparente. D’après Huntsman, 1948.
Certains spécialistes ont avancé que ces faits ne présentent pas en pratique de gravité. D’après eux, les pêcheries donneraient lieu à des phénomènes d’autorégulation mettant à l’abri les populations de poissons faisant l’objet des captures ; la rentabilité commerciale jouerait le rôle de frein dans les prélèvements. La pêche devient de plus en plus compliquée à mesure que se raréfient les populations (fig. 59) ; on est alors obligé d’acquérir un armement plus dispendieux, d’aller plus loin en mer pour atteindre d’autres secteurs ; la pêche perd ipso facto sa rentabilité. Le prix auquel le poisson est susceptible d’être commercialisé serait donc un frein pour la pêche, dont la pression diminue immédiatement dès qu’elle cesse d’être rentable. Il s’écoule ensuite un laps de temps considérable avant que la pêche ne reprenne, par suite de l’inertie de l’armement, ce qui permet aux populations de poissons de se reconstituer.
Ces affirmations ne sont cependant que très partiellement exactes du fait de deux facteurs très importants : d’une part la demande croissante en poissons sur un marché mondial loin d’être saturé ; d’autre part le perfectionnement actuel des moyens techniques qui permettent une pêche plus efficace qu’avec les moyens plus primitifs utilisés jusqu’à des temps relativement récents, tout en conservant et même en augmentant son rendement. La pêche industrielle a certes ses impératifs économiques qui en limitent la rentabilité au-delà d’un certain seuil ; mais celui-ci est beaucoup plus élevé que dans le cas de la pêche artisanale et tend à augmenter à mesure que se perfectionne la technique. Aussi l’overfishing se manifeste-t-il en divers points du globe et doit-il retenir l’attention de tous ceux qui se préoccupent d’une exploitation rationnelle des ressources naturelles.
Ce problème s’est posé avec une acuité croissante corrélativement au développement du chalutage hauturier. Vers 1880, l’apparition du chalutier à vapeur, puis, vers 1894, du chalut à plateaux bouleversa l’équilibre de la pêche en permettant l’exploitation de secteurs de plus en plus éloignés des côtes. De plus la taille des chalutiers ne cessa d’augmenter : le tonnage moyen des chalutiers anglais est passé de 177 t en 1906, à 231 en 1926, à 284 en 1937 et à 410 en 1966. Les bateaux devenus de plus en plus grands et très perfectionnés sont ainsi capables d’aller de plus en plus loin en mer et d’exploiter des secteurs demeurés jusqu’alors « en friche ».
En règle générale, le tonnage de poissons pêchés augmente tout d’abord en fonction de l’accroissement des moyens mis en œuvre ; puis il commence à diminuer, rapidement même dans certains cas. Symptôme caractéristique de l’overfishing, que nous allons préciser en évoquant quelques exemples particulièrement démonstratifs, et en montrant aussi comment on a réussi dans certains cas à rétablir par une sage réglementation de la pêche maritime une situation tout d’abord compromise.
Certes la déplétion de certains stocks de pêche ne s’explique pas uniquement par des prélèvements excessifs de l’homme. De nombreuses causes naturelles sont en effet susceptibles de modifier les équilibres de populations, voire d’en provoquer la rupture en dehors de toute influence humaine. Les poissons sont liés, surtout au moment de leur frai, à des conditions océanographiques très strictes, notamment en ce qui concerne la température et la salinité, agissant directement ou par modifications de la quantité d’aliments disponibles. Le jeu des transgressions et des régressions marines explique dans certains cas la diminution massive des populations de poissons, sans intervention de l’homme. Il ne faut pas automatiquement incriminer l’overfishing dans le cas d’une déplétion des mers.
Un exemple caractéristique est offert par les pêcheries au large des côtes péruviennes, dans les eaux froides baignées par le courant de Humboldt. La productivité de cet écosystème marin est normalement très élevée, ce que traduit aussi l’abondance des oiseaux (voir p. 429). Certes des phénomènes océanographiques, consécutifs à des transgressions répétées à plusieurs reprises au cours de l’histoire récente, expliquent un changement des conditions physiques de la mer, et par voie de conséquence une diminution sensible de l’abondance des poissons. L’homme a cependant contribué dans une très large mesure par des prélèvements excessifs à la régression massive des stocks et à la ruine des pêcheries. Son activité s’est exercée surtout aux dépens d’une espèce de petite taille, l’anchoveta (Engraulis ringens), que l’on convertit en farine de poisson. Une surexploitation prolongée a produit un effondrement des populations que traduit la diminution massive des tonnages débarqués. Le Pérou occupait en 1969 la première place avec 9 243,6 milliers de tonnes, pour un tonnage mondial de 62 700 milliers de tonnes. Ce chiffre s’éleva même à 12 612,9 en 1970, avant de tomber à 2 299,3, soit moins du cinquième, en 1973. Une surexploitation manifeste est venue conjuguer ses effets à ceux d’un phénomène naturel.
Cela doit être pour l’homme une raison supplémentaire de gérer sagement les ressources naturelles des océans, et de ne pas ajouter un facteur de destruction artificiel à des causes qui échappent à tout contrôle.
1. La pêche au Flétan et ses vicissitudes
Tout autour du cercle arctique, aussi bien dans l’Atlantique que dans le Pacifique, se rencontre un poisson d’excellente qualité, le Flétan Hypoglosses ou « Halibut », le plus grand de tous les Pleuronectes (dont la Sole est le mieux connu), pouvant dépasser une longueur de 3 m et un poids de 300 kg. Sa croissance est lente et sa maturité sexuelle tardive. Les populations de l’Atlantique ont été considérablement appauvries par des prélèvements excessifs, surtout celles d’individus trop jeunes et d’une taille insuffisante, particulièrement appréciés dans certains pays anglo-saxons. Il en subsiste sans doute encore dans les mers du Groenland à la Scandinavie, mais les stocks survivants ne peuvent plus donner lieu à une pêche industrielle régulière, sauf au large de la Norvège où une réglementation stricte protège les Flétans et en assure l’exploitation rationnelle.
Les populations de Flétans du Pacifique nord auraient suivi une évolution semblable sans les mesures de conservation prises par le Canada et les États-Unis. Elles furent en effet exploitées avec une intensité croissante au début de ce siècle ; 1,5 million de livres furent débarquées en 1888 et 50 millions en 1908. Mais les tonnages ne tardèrent pas à baisser d’une manière inquiétante, en dépit de l’exploitation de nouveaux fonds de pêche. Dans un district de la côte pacifique, la prise par unité d’effort passa de 320 livres en 1915, à 143 livres en 1920, et à 79 livres en 1927. Pour l’ensemble des secteurs, ces chiffres n’atteignirent en 1927 que le tiers des valeurs relatives en 1915. L’ensemble de la population de Flétans avait été réduit de deux tiers pendant cette période et présentait d’indéniables signes d’overfishing (fig. 60).
Cette situation obligea les gouvernements du Canada et des États-Unis à prendre des mesures. Une première convention passée en 1923 ferma la pêche pendant la période de reproduction. D’autres accords furent signés en 1930 et 1937, et complétés en 1953 pour réglementer la pêche.
Ces mesures ne tardèrent pas à produire leurs effets, car dès 1931, le tonnage global et les prises par unité d’effort augmentèrent à nouveau ; ces dernières passèrent dans les bancs de pêche à l’ouest du cap Spencer, dans la partie la plus méridionale de l’Alaska, de 65 en 1930, à 86 en 1934, à 112 en 1937, à 121 en 1940 et à 151 en 1944.
Depuis 1931, l’abondance des Flétans a augmenté de 150 % et le tonnage débarqué annuellement dans les ports dépasse maintenant couramment de 50 à 60 millions de livres ; en 1960, il a atteint le record de 71,9 millions de livres. Une commission (International Pacific Halibut Commission) veille à la conservation des stocks et en permet l’utilisation rationnelle. Le Pacifique est divisé en secteurs à l’intérieur desquels la pêche est réglementée, les captures limitées et les périodes d’ouverture de la pêche très étroitement circonscrites (fig. 61). Ces mesures sont soumises à révision en fonction de l’état des populations, ce qui assure une exploitation d’une bonne rentabilité économique.
Fig. 60. Variations du rendement de la pêche au Flétan sur la côte pacifique de l’Amérique du Nord, traduites par le tonnage des prises par unité d’effort (en livres). D’après Burkenroad, 1948.
Il est probable que les effets de l’overfishing sont venus se superposer à des fluctuations naturelles des populations de Flétans, car l’analyse des variations des tonnages prélevés et du déclin des populations suivi de leur accroissement fait apparaître certaines discordances (Burkenroad, 1948). Cela n’enlève rien à la valeur de l’argument ; bien au contraire, cela montre combien l’homme doit se montrer prudent dans l’exploitation d’un stock de poissons soumis à des fluctuations qui échappent totalement à son contrôle. Le Flétan du Pacifique nord reste l’exemple classique d’une ressource naturelle marine que des mesures adéquates ont permis de restaurer après avoir été sérieusement menacée par une exploitation excessive.
Fig. 61. Secteurs définis en 1950 par l’International Pacific Halibut Commission sur la côte pacifique de l’Amérique du Nord pour la conservation des stocks de Flétans et leur exploitation rationnelle. D’après Monod, 1956.
2. Le problème de la Sardine du Pacifique
La Sardine du Pacifique Sardinops caerulea peut sans aucun doute être elle aussi citée comme un exemple de surexploitation d’un animal marin (fig. 62). L’abondante littérature consacrée à ce problème envisagé aussi bien sous son angle scientifique que pratique témoigne de ses incidences sur l’économie humaine.

Fig. 62. Sardine du Pacifique Sardinops caerulea.
Ce petit poisson vit en bancs énormes, groupant souvent un million d’individus, parfois plus de 10 millions, le long des côtes de l’Amérique du Nord, de l’Alaska à la Basse-Californie. Habitant les eaux littorales, il peut s’écarter jusqu’à 350 milles des côtes au moment du frai, qui a lieu dans les eaux profondes (Gates, 1960). La pêche de cette Sardine a commencé vers la fin du siècle dernier (la première conserverie fut établie en 1899 en Californie). Le tonnage débarqué annuellement a oscillé entre 300 et 2 000 t jusqu’en 1912 ; il augmenta ensuite pendant la Première Guerre mondiale, pour se stabiliser quelque peu, avant de s’accroître à nouveau dans des proportions extraordinaires : en 1916-1917 il atteignait 27 000 t ; en 1924-1925, 174 000 t et pas moins de 800 000 t furent débarquées dans les ports américains répartis de la Colombie britannique à San Diego pendant la saison de pêche 1936-1937. Ces tonnages se maintinrent à un niveau voisin pendant les années 1930 et les premières années de la décennie suivante, faisant de l’exploitation de cette Sardine la première de tout le Nouveau Continent par le tonnage débarqué et la troisième en valeur commerciale (après celles du Thon et du Saumon) : la valeur marchande dépassait annuellement 10 millions de dollars. Au cours de cette extraordinaire prospérité, ces Sardines étaient utilisées non seulement par les conserveries, mais aussi en vue de la fabrication d’engrais et de farine de poisson destinée à l’alimentation du bétail.
À partir des premières années 1940, le tonnage débarqué subit des fluctuations importantes, puis diminua d’une manière dramatique (fig. 63) : en 1953-1954 seulement 4 460 t furent débarquées, condamnant à la ruine une industrie prospère et obligeant les armateurs à désarmer les bâtiments et à fermer les usines de transformation. Les premiers symptômes de la raréfaction de l’espèce se manifestèrent au Canada, où le tonnage débarqué passa de 34 000 t pendant la saison 1945-1946 à moins de 500 en 1947-1948. Puis ce fut le tour du nord des États-Unis, avant que la Californie, le principal centre d’exploitation, ne fût touchée à son tour vers 1951. La production est demeurée à un niveau très bas depuis cette époque203.
Dès 1940, cependant, les biologistes s’étaient inquiétés de cette exploitation abusive, dont les symptômes se manifestaient déjà par une proportion moindre de Sardines adultes et une augmentation du nombre de jeunes, montrant que l’on exploitait les immatures et les subadultes. L’effet de l’overfishing demeurait encore masqué par un effort de pêche accru et par un accroissement du tonnage de la flotte.
La surexploitation des Sardines est rendue responsable de cette raréfaction accentuée, aboutissant à la disparition de la rentabilité commerciale des opérations de pêche, bien que des causes naturelles aient pu accélérer le phénomène. La diminution du stock d’individus reproducteurs semble à incriminer, en raison des captures massives d’adultes, puis de jeunes encore immatures prélevés avant d’être en âge de se reproduire. L’action de l’homme est sans doute venue ajouter ses effets à une fluctuation naturelle de grande amplitude de ces populations, au moment où elles se trouvaient en pleine décroissance.
Fig. 63. Fluctuations des tonnages de Sardines du Pacifique débarqués dans les ports américains de la côte ouest. 1. Début des pêcheries en Colombie britannique ; 2. Début des pêcheries dans l’Oregon et le Washington ; 3. Abandon des pêcheries en Colombie britannique ; 4. Abandon des pêcheries dans l’Oregon et le Washington ; 5. Abandon des pêcheries à San Francisco. D’après Calif. Dept. of Fish and Game, Report, 1957.
La pression de la pêche a diminué par la force même des choses et on peut espérer que les stocks se reconstitueront à la faveur de la législation adoptée pour leur protection (saison de pêche limitée, contingentement du tonnage, constitution de réserves où toute pêche est interdite). Il n’en est pas moins vrai qu’une industrie prospère s’est trouvée ruinée au moment où les pêcheries d’autres parties du monde (peut-être tout aussi imprévoyantes) ont accaparé un marché où l’industrie californienne se trouve handicapée. Cet exemple montre clairement qu’une économie humaine se doit de ménager les stocks constituant sa matière première, faute de quoi elle ne peut qu’être vouée à l’échec en dépit d’une euphorie momentanée, cause même de la dilapidation du capital naturel qu’elle représente.
Le Merlu Merlucius merlucius, connu sur les marchés sous le nom de « colin », voisin des Morues dans la classification et propre aux mers tempérées ou chaudes, se rencontre particulièrement de l’Irlande aux côtes du Maroc et du Sénégal. Il a depuis fort longtemps fait l’objet d’une pêche intensive et constitue, surtout depuis la Première Guerre mondiale, un poisson très recherché en Europe.
La demande accrue a provoqué une exploitation exagérée des lieux de pêche, relativement limités en étendue. Dès 1910, on constatait une diminution sensible des apports des chalutiers pêchant au bord du plateau continental atlantique. Les pêcheurs étaient amenés à se rendre de plus en plus vers le sud, au large des côtes d’Espagne et du Portugal d’abord, du Maroc, de Mauritanie et du Sénégal ensuite. Les zones de pêche s’appauvrirent rapidement et bientôt se posa le « problème du Merlu ». L’examen des chiffres est particulièrement suggestif et montre que les mauvaises pêches sont dues à l’épuisement des stocks (fig. 64). La guerre, en ralentissant, voire en arrêtant la sortie des chalutiers, permit cependant à ceux-ci de se reconstituer. Les apports montrent une augmentation du tonnage de Merlu, lente d’abord et sujette à fluctuations, puis très rapide quand la pêche fut reprise à la fin de la guerre. Dans le secteur du plateau continental exploité par l’armement de La Rochelle, le tonnage moyen annuel débarqué par « marée » (campagne de 12 jours) par un chalutier à vapeur augmenta de plus de 21 t entre 1938 et 1946 ; le Merlu dépassait de plus de 13 t la moyenne d’avant-guerre avec des apports de 23,3 t par marée. La proportion de Merlus s’était accrue dans une plus importante proportion que celle des merluchons (Merlus de moins de 880 g, ayant de 3 à 6 ans), témoignant du vieillissement de la population dont la biomasse s’était considérablement accrue par suite d’un nouvel équilibre (Letaconnoux, 1951).
Fig. 64. Fluctuations des tonnages de Merlu débarqués à La Rochelle (total des pêches par marée de 12 jours). Le trait horizontal marque le tonnage moyen au cours de la période envisagée. D’après Letaconnoux, 1951.
Ce stock accru fut cependant rapidement décimé et la prospérité de courte durée. Dès 1949, le tonnage ramené aux ports retombait en dessous de la moyenne générale des 30 dernières années : 3 ans d’une mauvaise exploitation des populations de Merlu auront suffi pour ruiner le bénéfice dû à l’arrêt de la pêche pendant les hostilités ; une simple augmentation du maillage des filets aurait sans doute permis de maintenir la population à un niveau élevé et d’assurer à la pêche un rendement satisfaisant.
En 1949, le rendement des chalutiers ne fut que de 14,4 t en moyenne par marée, soit le même que celui des années 1937-1939 (14,7 t). Cet effondrement se poursuivit pendant les années suivantes. Simultanément s’observait une diminution du stock des autres poissons, eux aussi gravement menacés. L’examen de la répartition par taille et la structure de la population par classes d’âge montre qu’il s’agit manifestement d’un overfishing consécutif à un épuisement du stock dont la guerre avait permis la reconstitution.
Ces résultats ont été entièrement confirmés par des constatations similaires faites dans d’autres zones de pêche, et notamment dans la portion de l’Atlantique située au sud-ouest des îles Britanniques (Hickling, 1946). Si l’on se base sur les tonnages rapportés à Milford Haven, dans le Pays de Galles, on assiste à un ralentissement notable de la pêche pendant la guerre, ramenant dans les années 1941-1943 le tonnage débarqué à 39 % de ce qu’il était avant les hostilités. Depuis la fin de la guerre, un accroissement notable du tonnage a été enregistré, dû principalement à une taille supérieure des Merlus (le nombre d’Églefins et de Raies augmentait simultanément). Le rendement des chalutiers par heure de travail s’accrut de 200 % dans certains cas. En 1945, une flotte de 10 000 t captura 2,5 fois autant de Merlus qu’une flotte 3 fois plus importante en 1932, ce qui témoigne de la vitesse avec laquelle un stock de poissons est capable de se reconstituer. Mais cette amélioration n’aura pas eu de lendemain, car ces populations n’ont pas tardé à être elles aussi décimées par des prélèvements excessifs.
L’homme a donc compromis l’avenir d’une population reconstituée grâce à des circonstances fortuites. Aucune amélioration n’est à espérer tant que des mesures autoritaires ne seront pas prises pour laisser aux populations le temps d’accroître leurs effectifs et d’atteindre la biomasse maximale, parmi laquelle un contingent optimal pourra être prélevé.
Si les populations de poissons de fond (ou poissons démersaux) paraissent particulièrement sensibles aux effets d’un overfishing dangereux, on s’accordait à penser qu’il n’en est pas de même des poissons de surface (ou poissons pélagiques), et en particulier du Hareng Clupea harengus, le poisson le plus important au monde sur le plan économique, au point que la possession de ses lieux de pêche a même occasionné des guerres. Il semble cependant que certaines de ses populations présentent des signes indéniables d’une surexploitation de la part de l’homme.
Sans doute existe-t-il, comme dans le cas d’autres poissons, des fluctuations naturelles très importantes (dues notamment à des variations du taux de survie au cours du stade larvaire), dont il faut tenir compte pour apprécier l’influence de l’homme.
Un overfishing semble néanmoins exister, comme l’a notamment avancé Ancelin (1953) dans le cas des secteurs sud de la mer du Nord et de la Manche orientale. L’étude de la composition des populations qui font l’objet de la pêche dans ces régions le montre clairement, en particulier le remplacement anormal des classes constitutives du stock. La comparaison avec des espèces de poissons pêchés en excès, tels le Merlu et la Plie, est particulièrement suggestive. Les professionnels de la pêche se plaignent d’ailleurs du fait que les rendements actuels, pourtant favorisés par l’accroissement des moyens techniques, sont loin d’atteindre ceux qui ont été enregistrés après la guerre, indice probant en l’absence de statistiques établies sur des tonnages par heure de pêche. Les prises totales baissèrent de 364 à 111 millions de livres de 1950 à 1965. En 1960, la majorité des harengs pêchés avait un âge de 3 à 7 ans, en 1973, elle appartenait à des classes d’âge de 2 ans et moins.
Il faut donc admettre que les poissons de surface ne sont pas à l’abri d’un overfishing, en dépit de leur apparition saisonnière, de leur caractère pélagique et de la densité de leurs bancs.
Il n’est pas impossible que la pêche au chalut ait une influence sur l’état des populations de Hareng, les lieux de pêche étant exploités au moment de leur ponte qui se dépose au fond ; la destruction des œufs pourrait avoir une répercussion fâcheuse sur les effectifs des populations. Mais les prélèvements excessifs par capture au filet sont, avant tout, les responsables de la diminution. Les pronostics que l’on avait faits depuis longtemps se sont malheureusement révélés exacts (Garreau, 1970). Depuis 1965 on assiste à une diminution considérable des tonnages. En mer du Nord les prises tombèrent de 1,5 million de tonnes de Harengs en 1965 à 820 000 t en 1968. Alors qu’en 1960 on pêchait des Harengs de 3 à 7 ans, la majeure partie des prises a maintenant 2 ans. On capture de plus en plus d’immatures et une faible proportion du stock est en état de frayer. Si des quotas très stricts ne sont pas fixés et appliqués, on risque d’assister à l’anéantissement des populations de Harengs en mer du Nord, et aussi d’autres poissons, notamment des Maquereaux, sur lesquels les pêcheurs ont fait porter leurs efforts du fait de la baisse du rendement des Harengs.
5. La pêche dans la mer du Nord
La mer du Nord a été depuis longtemps une des zones les plus prospectées, pas moins de douze nations y envoyant leurs flottes de chalutiers hauturiers.
Cette exploitation intensive a mené les stocks de poissons de la mer du Nord à une diminution inquiétante. Pour l’Églefin Gadus eglefinus, le tonnage débarqué par jour d’absence des chalutiers dans les ports anglais est tombé de 7,8 quintaux en 1906, à 4,6 en 1914, pour remonter à 15,8 en 1919 et redescendre ensuite à 4,6 en 1933 et à 1,9 en 1935. Aux Pays-Bas, l’apport total d’Églefin est passé de 32 634 t en 1915 à 10 355 t en 1928. Simultanément les individus de petite taille devenaient proportionnellement plus nombreux de 1910 à 1913 : représentant 39 % des captures, la proportion passait à 69 % au cours de la période 1923-1930. Pour les îles Britanniques, les proportions progressaient de 42,5 % en 1913 à 81,7 % en 1928. Des constatations analogues concernent les Morues et les Plies. De 1925 à 1938, et à nouveau depuis 1950, les chalutiers opèrent avec un rendement minimal.
Les deux guerres mondiales apportèrent également dans cette zone des améliorations au stock des populations, en raison du ralentissement, voire de l’arrêt complet de la pêche (fig. 65). Le rendement par jour de la pêche des chalutiers anglais, pour l’ensemble des captures, qui s’élevait à 17,6 q en 1906, remonta à 30,6 q en 1919 ; il diminua ensuite régulièrement, s’établissant à 12,8 en moyenne de 1934 à 1937.
On conçoit l’importance de ce problème pour l’ensemble des pêcheries dont l’activité s’exerce dans cette partie des mers. Cette situation a provoqué des mesures conservatrices sur lesquelles nous allons revenir.
6. Les remèdes à la surexploitation des populations de poissons
Dans l’ensemble donc l’état actuel des populations est très alarmant. Si certaines mers ne sont encore que très peu prospectées (par exemple les mers australes et notamment celles qui bordent l’Australie), d’autres sont le théâtre d’un overfishing caractérisé, dramatique parfois dans ses proportions. La situation est d’autant plus grave qu’une demande sans cesse accrue provoque une pression de plus en plus vive sur les effectifs.
Fig. 65. Accroissement des stocks de poissons dans la mer du Nord et le nord-est de l’Atlantique, par suite de l’arrêt de la pêche pendant les deux guerres mondiales. L’augmentation est indiquée (par la zone hachurée) en pourcentage par rapport aux tonnages débarqués l’année précédant les hostilités (comme l’explique le graphique encadré à gauche). La surface de chaque diagramme est proportionnelle au tonnage débarqué. L’accroissement est partout très sensible. D’après Graham, 1949.
Le prélèvement intensif, grâce à des moyens techniques en voie de perfectionnement constant, permet d’accroître le volume des apports. La pêche hauturière est autant à incriminer que la pêche littorale, rendue responsable de la destruction d’un grand nombre de jeunes poissons, en particulier de Pleuronectes. La pêche crevettière est également considérée comme meurtrière en raison des mailles serrées des filets utilisés, où viennent se faire prendre d’innombrables jeunes poissons sans aucune valeur marchande, inutilement retranchés des populations aux dépens desquelles s’effectue la grande pêche.
En face de ces graves dangers, de nombreuses mesures ont été préconisées par les législateurs sur la recommandation des océanographes et des techniciens de la pêche.
L’une des premières vise à la réglementation du maillage des filets, de manière que les poissons d’une taille inférieure puissent s’échapper. Cela pose évidemment un grand nombre de problèmes techniques, dus aux caractéristiques des filets et aux tailles des poissons en cause204.
Par ailleurs on s’est employé à limiter les périodes d’ouverture de la pêche, à constituer des réserves où les populations de poissons peuvent se reconstituer et à délimiter des secteurs exploités par rotation de manière à assurer aux poissons qui s’y trouvent le temps d’atteindre leur taille optimale.
L’ensemble de ces mesures repose sur une connaissance précise de l’écologie et de la dynamique des populations de poissons exploitées. Ces exigences font qu’à travers le monde ont été créés des laboratoires océanographiques de recherches pures ou appliquées, dont les activités sont tournées uniquement vers cette étude.
D’autres problèmes sont d’ordre juridique. La haute mer au-delà de la limite des eaux territoriales échappe en effet aux juridictions nationales ; la réglementation de son exploitation met donc en jeu des règles très délicates de droit international. Un effort de compréhension a été fait par toutes les nations et s’est concrétisé par la signature d’une série de conventions du plus haut intérêt. Les hautes parties contractantes s’engagent à prendre des mesures visant les unes à favoriser l’accroissement des populations de poissons (par exemple interdiction de pêche dans le temps et dans l’espace, contingents des captures), les autres à limiter les prélèvements afin d’assurer le rendement optimal de pêche (par exemple interdiction de certains engins, maillage des chaluts, fixation des tailles marchandes).
La première convention internationale, relative à la pêche en mer du Nord, a été signée le 6 mai 1882. Depuis cette date on compte environ 150 accords bilatéraux, une dizaine d’accords régionaux et un plus petit nombre d’accords portant sur une espèce de poisson déterminée, avec parfois combinaison de ces deux données. On compte une douzaine de « grandes » conventions, intéressant 42 États (Gros, 1960).
Les conventions qui réglementent la pêche dans la mer du Nord et dans le nord-est de l’Atlantique figurent parmi les plus importantes du fait que de multiples nations y envoient leurs flottes et que ces mers constituent des terrains de pêche très anciennement exploités, d’un exceptionnel intérêt vu leur richesse et leur proximité des grands centres de consommation d’Europe. Les problèmes d’overfishing y sont particulièrement graves.
Fig. 66. Secteurs définis par la convention des pêcheries de l’Atlantique nord-est. Convention de Londres (1946). D’après Monod, 1956.
Une première conférence fut tenue à Londres en novembre 1936 et en mars 1937 ; elle aboutit à la signature de la convention internationale du 23 mars 1937, s’appliquant à l’Atlantique nord et aux mers adjacentes (une partie de la Baltique et la Méditerranée exclues). Cette convention fixait essentiellement le maillage des chaluts en fonction des zones envisagées et les tailles marchandes minimales de divers poissons.
En 1946, une nouvelle conférence, tenue à Londres pour réviser la convention de 1937 et l’amender compte tenu de l’évolution des stocks de poissons (fig. 66), aboutit à la signature de la convention internationale de l’overfishing, le 5 avril 1948205. Le maillage minimal était augmenté de même que les tailles minimales marchandes des poissons auxquels deux espèces étaient ajoutées (Merlan et Limande). Ces dispositions sont devenues effectives à partir de 1954. De plus, la convention a prévu la création d’une commission permanente qui groupe les délégués des pays signataires et est chargée de l’application de l’accord.
Au cours de cette conférence fut discuté par ailleurs, sur proposition de l’Angleterre, un projet de contingentement des tonnages prélevés, comme cela a été appliqué dans ce pays dès 1937 par accord volontaire des armateurs et par intervention du gouvernement. L’attribution de quotas fut écartée, mais peut-être l’imposera-t-elle dans le futur.
Notons aussi qu’une autre convention internationale concerne les zones de pêches de l’Atlantique nord-ouest (Washington, 8 février 1949), surtout en vue de préserver les stocks d’Églefins et de Morues, les plus importants des poissons de cette zone206. Nulle part ailleurs la pression sur les stocks n’a été aussi forte, les apports étant passés de 1,8 million de tonnes en 1954 à 4,2 millions de tonnes en 1970, grâce au perfectionnement des méthodes de pêche (Chaussade, Penn ar Bed, 75, 1973).
Des conventions similaires concernent par ailleurs d’autres régions du globe, notamment celle à laquelle il a été fait allusion quant au Flétan dans le nord-est du Pacifique. L’exploitation du Thon a fait aussi l’objet d’accords entre divers pays, notamment dans les mers baignant le Nouveau Monde (Inter American Tropical Tuna Commission).
Si ces mesures constituent de très sérieux progrès, elles n’ont cependant pas eu les résultats attendus. Aussi après des discussions techniques préalables se réunit en 1959 à Genève une conférence des Nations unies, afin de parvenir à un accord mondial sur l’exploitation des ressources marines. Elle aboutit à la rédaction de la convention sur la pêche et la conservation des ressources biologiques de la haute mer. Cette convention représente un pas important vers une réglementation de la pêche hauturière, car, pour la première fois, il est insisté sur la nécessité de la conservation des ressources, résultant d’un équilibre dynamique propre à assurer un rendement optimal constant. Au point de vue du droit international, cette convention est également très originale. Au cours des débats se manifesta en effet une nette opposition entre les partisans des vieux principes libéraux du droit international classique et ceux qui plaidaient en faveur d’une sorte de droit préférentiel dont profiteraient les nations riveraines, en contradiction flagrante avec le droit traditionnel de la haute mer207. Une place importante est ainsi faite aux droits économiques et sociaux des États en voie de développement, aux dépens de vieilles économies basées sur des pêches lointaines. Cette convention, signée après de laborieuses discussions par une trentaine de nations (avec des oppositions farouches de certains pays), n’est cependant pas encore ratifiée. Il est à souhaiter qu’elle le soit, car son application constituerait un progrès sensible vers une exploitation rationnelle des ressources de la mer (Gros, 1960).
L’urgence du problème n’échappe à personne. Il convient sans tarder de renforcer les mesures déjà en vigueur afin de faire face à la grave déplétion qui s’accentue dans maintes zones de pêche. La création de vastes réserves protégeant les frayères ou les zones où les poissons viennent croître devrait également être envisagée. Une rotation des districts et l’attribution de quotas à chaque État devront sans doute être instituées un jour prochain pour soulager la pression de la pêche.
Nous nous heurtons cependant alors aux innombrables difficultés des règlements supraétatiques, au problème des moyens de répression et de contrainte en cas d’infraction, très difficiles à appliquer comme les événements politiques de la dernière décennie nous l’ont appris sur bien d’autres plans. Il est à souhaiter que les hommes réussissent, pour une fois, à s’entendre sur le plan international. Il n’est plus question de leur prestige et de leur chauvinisme, car c’est l’utilisation rationnelle d’une ressource naturelle en péril qui est en jeu. Derrière les idéologies et les intérêts économiques de communautés et de particuliers, il ne faut pas oublier qu’il y a des mobiles « écologiques », car c’est en définitive la faim qui pousse les hommes à revendiquer les mers et à les exploiter d’une manière exagérée. Ce milieu n’aura le meilleur rendement – et il peut l’avoir à brève échéance – qu’en fonction d’une entente entre hommes de bonne volonté.
Notons par ailleurs que l’on a commencé l’élevage de poissons de mer, comme celui de la Morue en Norvège, de la Sole en France et de la Plie en Écosse208. Malgré l’ancienneté de certains de ces essais de « mariculture » (certains remontent à 1884), on en est encore au stade expérimental, de même qu’en ce qui concerne l’introduction artificielle d’espèces marines (par exemple l’acclimatation de l’Alose Alosa sapidissima de la côte orientale des États-Unis en Californie), dont il faut cependant se méfier a priori, car ces tentatives sont susceptibles d’entraîner des ruptures d’équilibre naturel. L’aquaculture marine est prometteuse dans les milieux côtiers et même dans les eaux littorales, en dépit de difficultés inhérentes à la complexité des écosystèmes marins et à des rendements souvent médiocres. Mais la haute mer restera sans doute encore longtemps, voire toujours, un milieu naturel sans aucune modification de la part de l’homme, si ce n’est sa prédation. L’humanité a asservi la terre dans sa quasi-totalité ; la mer demeure sauvage et les interventions de l’homme se bornent à y prélever, souvent inconsidérément hélas, ce qu’il estime nécessaire à sa consommation. Sur les mers, nous en sommes encore à la cueillette, en dépit de nos moyens de collecte hautement perfectionnés.
7. La chasse à la Baleine et sa réglementation
Nous avons évoqué précédemment quelques-unes des grandes étapes de la chasse à la Baleine à travers le monde et l’histoire de la raréfaction progressive de certaines espèces.
À l’époque actuelle, les baleiniers disposent d’un matériel leur permettant de capturer tous les Cétacés quelle que soit leur taille, y compris les Rorquals que leur puissance et leur vitesse mirent pendant longtemps hors d’atteinte209. Limitée à ses débuts à l’hémisphère Nord, la chasse s’étendit à partir de 1905 aux mers antarctiques (Budker, 1957). Tout d’abord entreprise à partir de navires-usines servant de stations flottantes sédentaires, cette chasse devint pélagique à partir de 1925, après l’intervention du slip-way, rampe inclinée du bateau permettant de hisser la Baleine sur le pont. Le nombre de Cétacés tués dans le monde suivit alors une courbe ascendante rapide, comme les quelques données suivantes permettent de s’en rendre compte (d’après International Whaling Statistics, Oslo, 1966) :
| 1900 : | 1 635 | 1935 : | 39 311 |
| 1905 : | 4 592 | 1938 : | 54 835 |
| 1910 : | 12 301 | 1951-1952 : | 49 794 |
| 1915 : | 18 320 | 1956-1957 : | 58 990 |
| 1920 : | 11 369 | 1961-1962 : | 66 090 |
| 1925 : | 23 253 | 1964-1965 : | 64 680 (dont 32 563, soit 50 %, dans l’Antarctique) |
| 1930 : | 37 812 |
Bien que l’exploitation des Baleines ait été planétaire, l’Antarctique est maintenant le théâtre principal de cette chasse ; pendant la saison 1930-1931, pas moins de 38 navires-usines et 184 bateaux chasseurs y pratiquèrent la chasse pélagique. Ce nombre fut de 9 navires-usines et 120 bateaux chasseurs pour la saison 1966-1967 (parmi lesquels 3 soviétiques avec 55 bateaux chasseurs).
Sans doute y eut-il des fluctuations dans le nombre de Cétacés capturés, par suite d’une surproduction et d’un effondrement consécutifs des cours, notamment en 1932, où le nombre de Cétacés tués passa à 12 988 contre 43 129 en 1931. Et les compagnies baleinières commençaient déjà à cette époque à prendre conscience du danger de déplétion.
Des ententes limitées furent prises de ce fait entre grandes nations baleinières et même entre les compagnies qui commencèrent à réduire leurs captures en se fixant des quotas. Dès 1932, une première convention passée entre la Norvège et la Grande-Bretagne limita le nombre de baleines et la production d’huile, de même que la période de chasse. Cette convention fut reconduite les années suivantes, tandis que la Norvège promulguait des lois baleinières concernant son armement. Certaines compagnies restaient toutefois en dehors de ces accords.
En 1937 fut accompli un pas décisif, à savoir la signature à Londres par les principales nations intéressées (Afrique du Sud, Argentine, Australie, Allemagne, États-Unis, Grande-Bretagne, Irlande, Norvège, Nouvelle-Zélande) de la première convention baleinière internationale ; ses clauses fixaient les dates d’ouverture et de fermeture de la chasse, les tailles limites et l’interdiction de chasser les Baleines franches et les Baleines grises. Cette convention fut reconduite en 1938, avec un article additionnel important : la création d’une réserve totale, comprise entre 70° et 160° de longitude ouest.
Le stock de Baleines se reconstitua pendant la Seconde Guerre mondiale. Mais la paix revenue, de graves carences alimentaires se faisaient sentir à travers le monde ; de ce fait on pouvait craindre un prélèvement excessif au fur et à mesure que les nations baleinières reconstituaient leur armement détruit pendant les hostilités.
Dès 1944, une première réunion se tint à Londres ; parmi les décisions les plus importantes figure l’introduction de la notion d’« unité Baleine bleue » (Blue Whale Unit, BWU). Les 4 Cétacés à fanons faisant l’objet des chasses sont de tailles très différentes. Comme il est difficile en pratique de limiter les captures par espèce, on fixa donc une limitation globale par des équivalences en tenant compte de la quantité d’huile fournie par les Cétacés et en prenant comme unité celle du plus grand, le Rorqual bleu. Cette équivalence s’établit comme suit : 1 Rorqual bleu (« Blue ») = 2 Rorquals communs (« Fin ») = 2 1/2 Mégaptères (« Jubartes ») = 6 Rorquals de Rodolphi (« Sei »).
On peut donc limiter le nombre de captures sans faire de distinction spécifique, formule pratique qui ne tient toutefois pas compte de l’importance des effectifs des différentes espèces. D’une manière arbitraire on fixa le contingent à 16 000 BWU pour la chasse pélagique dans l’Antarctique pour 1944 (avant la guerre la moyenne des captures était de l’ordre de 24 000 BWU).
En novembre 1945, une conférence baleinière réunit les délégués de 12 nations à Londres. Devant la pénurie de matières grasses à travers le monde, devenue particulièrement grave, il fut décidé de tout mettre en œuvre pour la pallier par la chasse des Cétacés, en limitant toutefois les captures pour ne pas compromettre l’avenir. Cette réglementation fut matérialisée par la signature par 19 nations de la convention internationale pour la réglementation de la chasse à la baleine à Washington, États-Unis, en date du 2 décembre 1946. Les clauses de cette convention, véritable charte de l’exploitation des Cétacés, concernent la protection des immatures (interdiction de capturer une femelle suitée, tailles limites), la limitation du nombre des Baleines capturées (quota fixé en BWU ou pour une espèce déterminée, limitation dans le temps par fixation des dates d’ouverture et de fermeture), la constitution de réserves intégrales (région antarctique comprise entre 70° et 160° de longitude ouest) et la protection d’espèces menacées (Baleines franches notamment). De plus la convention délimite une série de zones à travers le monde, où les modalités de la chasse sont réglementées (le navire-usine n’est autorisé qu’au sud de 40° de latitude sud et dans une partie du Pacifique nord en raison de son grand rayon d’action qui augmente ses facultés destructrices). Une commission baleinière permanente a été créée et est chargée (assistée de comités scientifiques) de l’application de ces clauses, notamment les pénalisations des chasseurs de Baleines enfreignant les règlements. Dix-sept nations ont adhéré à la convention de Washington210.
Si le nombre de Cétacés capturés annuellement reste très élevé, il tend néanmoins à diminuer par suite de la déplétion des stocks. En 1955-1956, on a capturé dans l’Antarctique 31 564 Cétacés parmi lesquels 27 958 Rorquals communs et 560 Rorquals de Rudolphi ; il faut y ajouter les captures effectuées par les stations à terre, non touchées par la limitation imposée aux navires-usines, et celles qui sont effectuées dans d’autres parties du monde. En 1964-1965, on ne capturait plus que 28 211 Cétacés dans l’Antarctique, parmi lesquels seulement 7 811 Rorquals communs et 20 380 Rorquals de Rudolphi beaucoup plus petits, chacun ne comptant que pour 1/6 d’unité BWU ; le Rorqual bleu n’entre plus que pour 20 dans ce chiffre, contre 5 130 en 1951-1952. En 1966-1967, le nombre de Cétacés tués dans le monde tombait à 51 593.
Ces chiffres sont inquiétants aux yeux de ceux qui se préoccupent non seulement de la protection de la nature, mais aussi d’assurer une exploitation rationnelle des ressources naturelles. D’après des travaux réalisés par le Discovery Committee, les Baleinoptères de l’Antarctique avaient des effectifs compris entre 142 000 et 340 000, avec une moyenne de l’ordre de 220 000 (la proportion des espèces est évaluée à 75 % de Rorquals communs, de 15 % de Rorquals bleus et de 10 % de Mégaptères). Les chiffres de captures paraissaient déjà nettement trop élevés, et l’on commençait à nourrir des inquiétudes quant à une surexploitation, notamment en ce qui concerne le Rorqual bleu et le Rorqual commun. L’espèce de loin la plus menacée apparaissait être le Rorqual bleu. Pendant les saisons de chasse (Antarctique) antérieures à 1937-1938, il formait l’espèce prédominante. Ainsi, de 1930 à 1933, si l’on considère l’ensemble des captures Rorqual bleu plus Rorqual commun, on constate que la proportion était environ de 80 % pour le premier et 20 % pour le second. Cette proportion s’est progressivement inversée, à tel point que bientôt le Rorqual bleu ne compta plus que pour 5 %. Le Rorqual commun a donc supporté l’essentiel de l’activité baleinière antarctique, et de ce fait a été décimé (entre les saisons 1955-1956 et 1963-1964, il aurait diminué des deux tiers d’après la commission baleinière). À l’heure actuelle, le même phénomène se renouvelle avec le Rorqual de Rudolphi, beaucoup plus petit, mais dont les captures furent près de trois fois plus importantes que celles de Rorqual commun en 1964-1965. Il diminue de ce fait rapidement dans l’Antarctique (de 30 % entre 1966-1967 et la saison de chasse précédente).
Le souci majeur des nations baleinières devrait donc, de plus en plus, être de rechercher les moyens de sauvegarder leur « matière première ».
Le comité scientifique de la commission baleinière internationale a, depuis plusieurs années, insisté sur la nécessité de prendre des mesures énergiques. Un comité de trois spécialistes de la dynamique des populations, auquel s’est joint ultérieurement un quatrième, a été formé en 1960 ; il s’est réuni à Rome puis à Seattle, et a présenté son rapport final à la dernière session de la commission. Les conclusions de ce rapport sont très pessimistes et confirment les avertissements que le comité scientifique de la commission baleinière n’a cessé de donner depuis plusieurs années.
L’abolition de l’unité Baleine bleue (BWU) serait très souhaitable, car celle-ci n’est en somme qu’un artifice permettant d’assigner une limite au chiffre des captures. L’idéal serait de prescrire une limitation par espèce : c’est le vœu du comité scientifique, comme celui du « comité des trois ». Malheureusement, l’application pratique comporte des difficultés considérables, et pour le moment, la BWU a été maintenue.
La commission a pris diverses mesures destinées à protéger les stocks de Cétacés, en interdisant d’abord la capture des Rorquals bleus dans une bonne partie de l’Antarctique et celles des Mégaptères dans l’hémisphère Sud.
En dépit de ces restrictions, les populations ont cependant continué de décliner. Nous avons vu que le Rorqual bleu est particulièrement menacé et l’on a pu craindre son extinction virtuelle. Ses effectifs antarctiques, qui s’abaissèrent à moins de 2 000, ne dépassent pas 6 000 individus actuellement, alors que la population optimale serait de l’ordre de 100 000 individus. Le comité des trois spécialistes mentionné ci-dessus concluait qu’il est possible que les populations aient régressé au-delà du minimum requis pour une reproduction normale et que l’espèce était de ce fait même condamnée. Les autres Rorquals se trouvent également en mauvaise posture, ne serait-ce qu’en raison d’une pression de chasse d’autant plus forte que le Rorqual bleu, d’un bien meilleur rendement vu sa taille, ne compte plus dans les captures. Or il existe un conflit à propos des baleines. En raison des impératifs de la rentabilité actuelle des campagnes de chasse et des besoins mondiaux en matières grasses, les captures annuelles continuent à être beaucoup trop élevées. On assiste ainsi progressivement à la déplétion des stocks. Les espèces risquent de se raréfier jusqu’à un point critique et une richesse naturelle d’une importance considérable est en voie d’être dilapidée dans les années prochaines.
Il est à souhaiter que de sages mesures soient prises et appliquées avec rigueur par les plus hautes instances internationales afin de ne pas compromettre définitivement l’avenir de cette ressource marine dont dépend une partie importante de l’humanité. Mais on a de sérieux doutes sur ce qui risque de demeurer un vœu pieux, surtout quand on constate que la commission baleinière a rejeté tout moratoire à l’exploitation des grands Cétacés, se contentant de réduire quelque peu des quotas bien trop élevés.
8. Le problème des Crustacés
Les Crustacés ne seront que brièvement mentionnés, bien qu’ils soient partout en diminution à travers le monde en raison de la déplétion des stocks consécutive à un prélèvement excessif. D’ailleurs les différentes « guerres de la Langouste » dont on a été témoin dans les temps récents, par exemple au large du Brésil, sont un signe de la raréfaction des Crustacés dans certaines zones de pêche, forçant les pêcheurs à se rendre vers d’autres, où ils se heurtent à des problèmes de concurrence compliqués de contestations juridiques.
Cette évolution n’est pas seulement perceptible dans les mers bordant l’Europe, car d’indéniables symptômes d’overfishing ont été signalés dans des mers aussi éloignées que celles qui baignent les îles subantarctiques Saint-Paul et Amsterdam où une Langouste (Jasius lalandei) donne lieu à une exploitation régulière (Paulian, 1957). D’après les résultats des campagnes de pêche on constate que les tonnages pêchés ont sérieusement diminué en dépit d’une meilleure connaissance des fonds. La ponction considérable opérée a été excessive et l’on a provoqué, en dépit des avertissements des biologistes, une rupture d’équilibre fatale à toute exploitation. Il conviendra de suspendre toute opération de pêche pendant un temps, puis de limiter sévèrement le prélèvement, d’autant plus que le plateau continental est étroit et que les populations de Langoustes sont réduites, même si des mers plus profondes servent de « réservoirs ».
Des décisions semblables seraient à prendre dans bien d’autres régions du monde fixant notamment d’une manière plus étroite les limites des tailles marchandes (réglementation de la dimension des mailles des casiers), car partout les grands Crustacés accusent une diminution sensible des effectifs. La pêche crevettière dans divers secteurs, notamment au large de l’Afrique occidentale, est elle-même en déficit sérieux.
9. Le problème des Mollusques
Les Mollusques ne sont nullement à l’abri d’une surexploitation de la part de l’homme. Si l’ostréiculture, ayant pour but la production d’Huîtres comestibles, est devenue une véritable entreprise industrielle échappant dans un certain sens aux lois biologiques et ne dépendant plus qu’en faible proportion des bancs naturels211 – elle s’apparente souvent plus à l’élevage au sens agricole du terme –, il n’en est pas de même de l’exploitation des Mollusques producteurs de nacre. Celle-ci s’adresse en effet à des populations sauvages que l’homme a dévastées dans bien des lieux par ses prélèvements excessifs.
Le meilleur exemple se trouve en Polynésie française et plus spécialement aux îles Tuamotu, dont la principale ressource économique est précisément la nacre, avec le coprah et le phosphate (Ranson, 1962).
Les Mollusques producteurs sont des Huîtres perlières de la famille des Aviculidés appartenant à plusieurs espèces, dont la plus importante, et de loin, est Pinctada margaritifera, répandue de Tahiti aux Fidji et en Nouvelle-Calédonie ; des espèces voisines sont distribuées dans d’autres districts et notamment au Japon et dans l’océan Indien (elles étaient autrefois pêchées pour leurs perles, mais cette exploitation est aujourd’hui abandonnée). Leur coquille ovale, noirâtre ou verdâtre à l’extérieur, présente à leur intérieur dans la partie centrale une couche de nacre blanchâtre (aragonite) ; elle peut atteindre 30 cm de diamètre et peser jusqu’à 10 kg.
Dans les lagons des îles océaniennes où les plongeurs atteignent le fond, les stocks d’Huîtres nacrières se sont progressivement épuisés. Or ces Mollusques ont de faibles chances d’assurer leur descendance, en dépit de leur énorme fécondité. On estime que pour un million d’œufs expulsés (une femelle de grande taille en émet plusieurs dizaines de millions), de 1 à 10 seulement donneront des individus arrivant à l’âge adulte. L’extinction totale de la population est prévisible à plus ou moins brève échéance dès que le stock de reproducteurs tombera au-dessous d’une certaine limite. Il faut maintenir des dizaines de milliers de reproducteurs concentrés sur un espace relativement restreint pour assurer la continuité de l’espèce ; cette concentration est une condition impérieuse à la survie des populations, d’autant plus que les sexes sont séparés.
C’est ce que n’avaient pas compris les imprévoyants plongeurs polynésiens qui épuisèrent véritablement la plupart des lagons. Les chiffres globaux publiés concernant l’exportation de « nacres » ne donnent qu’une idée très approchée de l’évolution de la production en raison d’une commercialisation irrégulière.
Les renseignements recueillis sur des lagons précis sont beaucoup plus démonstratifs ; en voici quelques-uns, tous dans les Tuamotus212 :
Vers 1900 kg | Vers 1940-1950 kg | |
| Arutua | 60 000 | 1 500 |
| Kaukura | 40 000-50 000 | 0 |
| Manihi | 80 000 | 4 688 |
| Aratika | 80 000-100 000 | 911 |
| Hikueru15 | 1 000 000 | 528 401 |
| Takapoto | 400 000 | 211 000 |
Il ne restait que 6 centres importants de production en plus de 8 lagons ayant encore une rentabilité appréciable. Tous les autres, soit 35, ont une production nulle ou insignifiante, les populations y étant pratiquement épuisées.
En dépit des avertissements de certains observateurs, remontant à 1884, l’homme a donc réussi à appauvrir gravement les ressources naturelles d’îles réputées heureuses.
Les mesures propres à régénérer les effectifs sont avant tout de constituer dans chaque lagon des réserves naturelles, abritant des Huîtres pleinement adultes, donc capables d’émettre en grand nombre de produits génitaux viables ; la pêche n’est jamais ouverte dans cette zone d’où les larves émises se dispersent sur l’ensemble du lagon. Il convient de disposer ailleurs des supports (pierres ou fagots de bois) sur lesquels les larves viennent se fixer, en un mot d’appliquer les techniques classiquement employées en ostréiculture (fig. 67). On arrive ainsi à enrichir les lagons épuisés, le milieu nourricier où les Huîtres trouvent leur subsistance étant demeuré constant. La définition de secteurs exploités par rotation compléterait la réglementation, comme l’interdiction de la pêche de nacres ayant moins de 15 cm de diamètre.
Les principes généraux d’une exploitation rationnelle s’appliquent donc aux Huîtres perlières, à savoir la constitution d’une réserve intangible où se multiplie l’espèce, et la sage gestion d’un capital fragile, mais dont la fécondité du « cheptel » autorise d’espérer la régénération.
Fig. 67. Plan schématique du lagon d’Hikueru, îles Tuamotu, et délimitation des zones en vue d’une exploitation rationnelle des Huîtres nacrières. La réserve où la plongée est interdite est hachurée. Trois secteurs à exploiter par rotation sont délimités. Les collecteurs (destinés à favoriser la fixation des larves) sont représentés par des croix (fagots de bois) et des cercles (pierres). D’après Ranson, 1962.
En Nouvelle-Calédonie un problème similaire concerne le Troca Trochus niloticus, autre coquillage fournissant une nacre épaisse et résistante (Angot, 1959) (fig. 68). Bien qu’une abondante population soit établie sur le plateau récifal, le haut profit tiré de son exploitation a mis la rentabilité de l’opération en péril. Quand aux alentours de 1907 commença la pêche, le Troca était extrêmement abondant et se ramassait à la pelle et au râteau sur les trottoirs récifaux à marée basse ; dès 1910-1920 cependant il se raréfia dans ce milieu et commença à être exploité à la plongée. En dépit d’un effort de pêche accru et du perfectionnement des méthodes de collecte, dès 1930 la production moyenne était en diminution régulière, exemple caractérisé d’overfishing : 1 000 t furent exportées annuellement jusque vers 1913, 790 en 1916, 622 en 1922, 389 en 1924 et 358 en 1928.

Fig. 68. Troca Trochus niloticus.
En dépit des cris d’alarme lancés à cette époque, l’exploitation continua, avec une exportation moyenne de 400 t par an jusqu’à avant la Seconde Guerre mondiale. Après la réorganisation des marchés, en 1946, la surexploitation recommença de plus belle. De 1946 à 1948, l’effort de pêche demeura le même, mais le tonnage récolté fit une chute spectaculaire de 1 221 t (ce chiffre élevé s’explique du fait de l’interruption de la pêche pendant la guerre) à moins de 500 t. Après une stabilisation de 1948 à 1953, sur les bases d’un tonnage moyen de 500 t, un accroissement prodigieux de l’effort de pêche conduisit tout d’abord à une augmentation massive du tonnage prélevé (880 t en 1954), suivi d’un déclin tout aussi rapide : 723 t en 1955, 402 en 1956. Les pêcheurs avaient augmenté en nombre et exploitaient des zones de plus grande profondeur, demeurées vierges jusqu’alors ; malgré cet effort ils ne parvenaient plus à maintenir le tonnage collecté, le prix du Troca montant d’ailleurs en flèche (fig. 69).
Fig. 69. Variation du rendement de la pêche au Troca en Nouvelle-Calédonie de 1946 à 1956. Remarquer la diminution spectaculaire du tonnage en 1947-1948. De 1953 à 1956, on remarque une intensifi-cation des activités de pêche (traduite notamment par l’augmentation du nombre de pêcheurs), mais une baisse du tonnage récolté. (L’augmentation de la taille des coquilles, due à une exploitation en zones plus profondes, a de plus exigé un effort de pêche accru.) La comparaison de ces diverses courbes traduit l’overfishing.
D’après Angot, 1959.
Pour pallier les efforts si néfastes de l’overfishing, les autorités locales prirent un certain nombre de mesures, parmi lesquelles l’interdiction de la pêche pendant un an à partir de septembre 1956, pour restaurer le stock, et la fixation de la taille limite des Trocas négociables (10 cm de diamètre au minimum). La limite minimale de taille des Mollusques constitue à elle seule une mesure adéquate, car elle permet à de nombreux adultes de se reproduire et de repeupler ainsi les surfaces récifales d’où l’espèce a presque entièrement disparu. Des signes certains permettent d’espérer que la réglementation adoptée se soldera par un succès en permettant au stock de Trocas de se renouveler et d’assurer ainsi par son exploitation rationnelle un revenu constant à des communautés de pêcheurs dont la plongée est la seule ressource.
10. Les Tortues de mer
Les Tortues de mer font l’objet d’une exploitation très ancienne, en particulier pour leurs œufs. Ces Chéloniens viennent en effet à terre pour enfouir leurs pontes dans le sable ; les hommes les trouvent d’autant plus facilement que ces lieux d’étendue réduite sont visités d’année en année depuis des temps immémoriaux. Certaines espèces sont exploitées pour l’écaille de leur carapace ; d’autres, dont le plastron a une texture particulière213, servent à la confection de la soupe à la tortue, consommé considéré comme un mets de choix dans le monde « civilisé », notamment dans les pays anglo-saxons.
Ces diverses utilisations ont entraîné une surexploitation manifeste des diverses espèces de Tortues marines qui ont diminué dans des proportions considérables à travers les mers chaudes qu’elles fréquentent (Hendrickson, 1961 ; Parsons, 1962). La Tortue luth Dermochelys coriacea – le géant du groupe pesant jusqu’à 1 500 livres, véritable fossile vivant dépourvu de carapace remplacée par des plaques incluses dans la peau – n’est plus représentée que par des populations relictuelles au Costa Rica, en Guyane, en Afrique du Sud et en Malaisie (Trengganu), où moins de 1 600 femelles pondent annuellement dans ce qui est considéré comme le plus important lieu de reproduction. La collecte exagérée des œufs est responsable de la diminution rapide de cette espèce même considérée un moment comme éteinte.
La Tortue verte ou Tortue franche Chelonia mydas – qualifiée par Archie Carr, le spécialiste mondial des Tortues de mer, de « reptile le plus utile du monde » – est, elle aussi, exploitée pour ses œufs, notamment dans le Sud-Est asiatique ; la surexploitation a entraîné une diminution massive de la production, tombée pour les colonies du Sarawak, de 2 184 095 pour la période 1929-1936 à 1 038 129 pour la période 1955-1961 (Harrisson, 1962). De plus cette espèce jouit du triste privilège d’être parmi celles qui fournissent la matière première de la soupe à la tortue ; de ce fait, elle est pourchassée à travers le monde entier. Elle se trouve ainsi en grave danger, comme plusieurs autres espèces, principalement la Tortue à écaille Eretmochelys imbricata, de l’Atlantique, qui fournit une écaille renommée et un succédané pour la confection du fameux consommé.
Il convient donc de prendre des mesures pour assurer la survie des Tortues de mer et garantir l’exploitation rentable d’une richesse naturelle compromise par des collectes abusives. Les portions de plages où les Tortues pondent leurs œufs doivent être mises en réserve, comme cela a déjà été fait en Malaisie et au Costa Rica, et la collecte des œufs sévèrement contrôlée. Les essais de transfert des œufs dans des enclos spécialement aménagés, dont le succès est très prometteur, méritent d’être poursuivis et étendus (notamment en Malaisie ; Hendrickson).
11. Les dangers de la pêche sportive
et de la chasse sous-marine
La pêche sportive a pris une très grande extension au cours des dernières années, surtout dans le Nouveau Monde, et en particulier dans les mers chaudes bordant la Floride et les Antilles. Le nombre croissant d’amateurs a obligé les gouvernements intéressés à promulguer des réglementations afin de protéger les populations de poissons ainsi exploitées. Le grand intérêt économique de la pêche sportive, devenue une véritable industrie par les activités qu’elle suscite, telles que construction de bateaux et fabrication d’équipement sans compter l’hôtellerie (en Floride un visiteur sur quatre est un pêcheur sportif), justifie pleinement la protection des stocks aux dépens desquels elle s’effectue.
Par ailleurs il faut mentionner le développement rapide de la chasse sous-marine dans toutes les régions où la mer est suffisamment chaude et limpide, par exemple dans la région méditerranéenne. Les innombrables plongeurs armés de leurs engins peuvent contribuer à dépeupler les zones littorales en certains poissons. De divers côtés on a signalé la raréfaction des espèces particulièrement recherchées. Mais les chasseurs sous-marins n’exploitent cependant qu’une très étroite bande côtière, en raison de la faible profondeur qu’ils sont capables d’atteindre. Aussi les menaces ne sont-elles encore que peu sérieuses, bien qu’il conviendra de ne pas négliger ce problème à l’avenir.
On notera à ce point de vue la remarquable initiative prise par l’État de Floride qui a aménagé en bordure de son territoire le premier parc sous-marin du monde (fig. 70). Au sud-est de la péninsule, au large de Key Largo, s’étend une frange de récifs coralliens qui se prolonge le long des Florida Keys ; cette frange est formée de Coraux variés, au milieu desquels vivent des pléiades de poissons multicolores. Ce district était mis à sac par les fournisseurs de magasins de curiosités, qui utilisaient barres à mine et explosifs pour arracher les Coraux dont les squelettes blanchis sont vendus aux touristes en quantités énormes. De plus les jeunes poissons faisant l’objet de la pêche sportive viennent s’y abriter, et les populations risquaient fort de diminuer par suite des perturbations apportées dans leur « nursery ». Dès 1957, les biologistes se préoccupèrent de la sauvegarde de cette richesse naturelle. Des difficultés administratives retardèrent la création de la réserve qui fut finalement promulguée en 1960. La Pennekamps Coral Reef Preserve (ou Key Largo Coral Reef Preserve) s’étend sur 21 milles marins de long et 4 milles de large, soigneusement balisés. Cette réserve est ouverte aux visiteurs, habilités à y plonger, mais non à y chasser au harpon ou à prélever le moindre échantillon. Un programme de recherches scientifiques a été entamé dans cette zone désormais à l’abri des perturbations de la part de l’homme.
Fig. 70. Emplacement du parc sous-marin établi au large de la Floride.
Cette réserve n’est pas la seule de son genre, car il faut aussi citer les réserves sous-marines de Virgin Islands N. P., de Buck Islands, près de Saint Croix Island, des Dry Tortugas et à Exuma Cave aux Bahamas. D’autres projets visent à établir des zones de protection à Phosphorescent Bay, Porto Rico, et à Bucco Reef, Tobago.
Par ailleurs en mars 1968, le Kenya a établi deux parcs nationaux marins destinés à protéger l’habitat aquatique si spectaculaire de ses côtes. Couvrant une superficie d’environ 17,5 km2, ils sont complétés par des réserves nationales dont la plus grande, dans la région de Malindi, s’étend sur 181 km2.
On ne peut manquer d’insister sur l’importance de telles réserves naturelles. De nombreux parcs marins ont été créés depuis cette époque ou sont en voie de l’être. Une conférence tenue à Tokyo en mai 1975 a permis de définir une politique à l’échelle mondiale.
Il est urgent de préserver en haute priorité les écosystèmes coralliens menacés par les pollutions, l’extraction des sables, un tourisme sauvage et le vandalisme de collecteurs sans scrupule. Les magnifiques jardins sous-marins de Coraux, animés de poissons multicolores, sont d’une fragilité extrême et ne résisteraient pas à des aménagements mal conçus dans la zone littorale, générateurs de nuisances et de ruptures d’équilibre écologique.
12. Les ressources marines dans le futur
Les ressources marines prises dans leur ensemble ont fait et font donc encore souvent l’objet d’une exploitation irrationnelle, qui met leur pérennité même en danger. L’overfishing se fait sentir dans des régions très diverses du globe, parmi les plus riches cependant, motivé par des raisons le plus souvent économiques et financières, en un mot le profit.
La déplétion des zones de pêche classiques entraîne à travers le monde entier des « guerres du poisson » entre pêcheurs venus souvent de très loin. Il suffit de rappeler des incidents survenus dans les temps récents entre Russes et Américains sur le grand banc de Terre-Neuve et en Alaska ; entre Islandais et Britanniques au large de l’Islande ; entre Américains, Japonais, Coréens et Russes dans le Pacifique nord ; et entre Irlandais, Belges et Hollandais au large de l’Islande, pour être convaincu de l’âpreté de la lutte engagée par les différentes nations pour s’approprier le poisson au risque de dépeupler à jamais les zones de pêche les plus fréquentées et de rendre les opérations de moins en moins rentables. Le prélèvement anarchique continue à s’effectuer comme par le passé, mais ses conséquences sont beaucoup plus graves, étant donné l’efficacité des moyens mis en œuvre, sans commune mesure avec ce qu’ils étaient il y a peu de temps.
La gravité de la situation n’échappe à personne. En raison de l’accroissement de la population humaine, la pression de la pêche va aller en augmentant et l’on demandera de plus en plus à la mer, notamment en protéines animales, dont les océans restent la source principale. Il convient en conséquence de continuer et d’amplifier les efforts faits actuellement pour assurer un rendement élevé aux pêcheries, voire l’accroître, tout en sauvegardant le capital lui-même.
Il est impensable à l’heure actuelle que l’homme puisse modifier la haute mer et la « mettre en valeur » en la transformant comme il a fait des milieux terrestres. Il s’agira donc toujours de l’exploitation d’une richesse naturelle non transformée.
Dans leur ensemble les ressources marines restent peu exploitées, les mers de l’hémisphère boréal – mer du Nord, Atlantique et Pacifique nord – mises à part. Une augmentation du rendement global des mers est encore possible par la mise en exploitation de nouveaux secteurs, en particulier le long des côtes d’Afrique, d’Amérique du Sud (Argentine, Uruguay) et d’Australie. Les mers tropicales abritent de nombreux poissons pratiquement inexploités ; il est vrai que les coraux y rendent le chalutage impossible et que de nouveaux procédés de pêche (notamment pêche électrique) doivent être mis au point avant que ces secteurs puissent être ouverts à une utilisation à grande échelle.
Il y aurait aussi lieu d’élargir l’éventail des espèces utilisées dans la consommation humaine. Quatre-vingt-dix pour cent de la biomasse des animaux marins sont constitués par des Invertébrés, pratiquement inexploités, hormis quelques Mollusques et Crustacés. La plupart paraissent sans doute non comestibles, mais des recherches seraient à entreprendre dans ce domaine.
Même parmi les poissons, une très faible fraction est consommée par l’homme et donne lieu à une exploitation régulière. Seuls 6 % des espèces sont exploités et 2 % couramment. En pratique une douzaine d’espèces font l’objet de pêches commerciales ; si l’on prend le tonnage comme base d’estimation, 33 % sont constitués par des Clupéidés (Harengs, Sardines, etc.), 25 % par des Gadidés (Morues et espèces voisines), 9 % par des Scombridés (Maquereaux, Thons), 3 % par des poissons plats (Soles, Limandes, Turbots, etc.), le reste étant formé d’espèces diverses. Il serait donc souhaitable d’envisager l’exploitation d’autres types de poissons.
Cela n’irait sans doute pas sans de nombreuses difficultés. Tout d’abord il ne faut pas oublier que si l’homme s’est cantonné jusqu’à présent à quelques espèces, des raisons biologiques et pratiques ont présidé à ce choix, les populations étant particulièrement importantes et leur pêche présentant des facilités exceptionnelles. Il faudrait donc étudier de nouveaux engins et modifier les techniques de pêche en fonction de la biologie des espèces. De plus il faudrait faire l’éducation des consommateurs ; même dans des pays hautement évolués, de nombreuses barrières « psychologiques » empêchent la consommation de poissons auxquels le public n’est pas habitué. L’humanité devra cependant vaincre ces préjugés, le plus souvent sans fondement, si elle veut assurer sa subsistance. Ces poissons pourraient ne pas entrer directement dans le régime alimentaire de l’homme, mais servir à la fabrication de farines pour l’alimentation du bétail, comme cela se fait déjà pour un tiers des captures mondiales. Cela constitue cependant un énorme gaspillage, car l’animal domestique ne restitue qu’une faible part des protéines qu’il a ingérées. Il est navrant de constater que certains pays souffrant de carences alimentaires, comme le Pérou, pêchent pour fournir des surplus de luxe à des pays riches !
Rappelons enfin que certains ont préconisé l’exploitation directe du plancton. Sur le plan écologique, cela aurait pour effet de supprimer de nombreux stades intermédiaires dans les chaînes alimentaires ; il en résulterait un meilleur rendement : il faut au moins 10 kg de plancton pour faire 1 kg de poisson ; dans la mer du Nord 2 millions de tonnes de Harengs équivalent à 50-60 millions de tonnes de plancton. Au lieu de passer par l’intermédiaire des poissons de surface (Hareng, Sardine), l’homme consommerait ainsi directement, sous une forme à trouver, la nourriture planctonique de ceux-ci, d’où un meilleur rendement énergétique. Aussi tentante que soit à première vue cette solution, de nombreux problèmes se présentent immédiatement, en raison de la difficulté de collecter ces animaux et végétaux microscopiques en suspension dans l’eau. La consommation directe du plancton par l’homme est un rêve, et ce n’est pas demain que l’homme se substituera aux poissons de surface dans les chaînes alimentaires marines.
La mer conserve encore à l’heure actuelle sa haute productivité, et est même capable de régénérer assez rapidement les effectifs des animaux que l’homme a imprudemment décimés. Elle est une des grandes sources d’aliments du monde de demain. Une exploitation rationnelle de la part de l’homme peut assurer les moissons marines dont dépend en grande partie sa survie. Il convient sans tarder de replacer les prélèvements à leur niveau optimal et de permettre, par des mesures de protection, aux fonds de regagner leur potentiel biologique compromis par nos gaspillages.