FOUR
Sexual Self

WHEN WE LEARN THAT A BABY IS BORN, OUR FIRST question is almost always “girl or boy?” In every society, sex is a fundamental trait used to categorize individuals. It’s the first box to check on every form. When we meet someone new, we cannot help but seek to ascertain their sex. It’s a deep, subconscious drive that cannot be turned off. Sex is also the individual trait that we’re least likely to forget. You may not recall whether a person named Terry who you met briefly at a party two years ago had brown or black hair or worked as an accountant or a marketer, but you are very unlikely to forget his or her sex. No matter your culture, religion (or lack thereof), or politics, sex matters to all of us. That’s why people can get so upset when talking about it. Complicated and changing ideas about sex, including gender identities beyond the traditional female/male binary, can challenge the essence of who we are.

ON SEPTEMBER 22, 1938, an urgent message was transmitted from the police station in Magdeburg, Germany, to Berlin: “Women’s European high-jumping champion Ratjen, first name Dora, is not a woman, but a man. Please notify the Reich Sports Ministry at once. Awaiting orders by radio.” The Reich sport minister, Hans von Tschammer und Osten, didn’t want to believe this news—a deep embarrassment for the German state—and called for his own doctors to examine Dora. But their finding was the same. Dora Ratjen, nineteen, who had competed for Germany in Hitler’s showcase 1936 Berlin Olympics, and who had set a world record for the women’s high jump at the European Athletics Championship in Vienna just a few days earlier, was, in truth, a man—at least by the standards of Germany in 1938. Under Tschammer und Osten’s direction, Dora’s gold medal was quietly returned, her world record was struck from the books, and she was banned from sports competitions for life.
MALES TYPICALLY INHERIT ONE X and one Y sex chromosome, while females usually have two X sex chromosomes. One crucial gene on the Y chromosome, SRY, codes for an important protein that, by affecting the activation of other genes, guides male-typical development beginning early in embryonic life. In the presence of the SRY gene product, two small blobs of tissue are instructed to become testes, which secrete the hormone testosterone. Testosterone (or its metabolite dihydrotestosterone) then binds to specific receptor proteins in cells and has widespread effects throughout the body. It is a key signal driving male-typical development of everything from the genitalia (during embryonic development) to the Adam’s apple (much later, during puberty). In females, in the absence of SRY, other genes actively drive these same blobs of embryonic tissue to become ovaries, which secrete the key hormones estrogen and progesterone.1 Importantly, although testosterone is present in two different surges starting in early fetal life, the secretion of estrogen is suspended from the period shortly after birth to puberty. This means that, during certain critical stages of development, the main hormone difference is higher levels of circulating testosterone (and some testosterone-like hormones, collectively called androgens) in most males and lower levels in most females. However, females are not completely bereft of androgens as, starting around age eight, the adrenal glands secrete low levels of testosterone (and dihydrotestosterone and androstenedione), and these androgens are also important for normal development in females. In males, estrogen has a role in normal development and adult function as well, although some of the details have yet to be worked out.
In most people, sex determination is straightforward. You inherit an X chromosome from your mother and either an X or a Y chromosome from your father. If a Y-carrying sperm cell fertilizes the egg, you’ll be male; if it’s an X, you’ll be female. If you carry XX sex chromosomes, then you’ll develop ovaries, a vagina, and a typical vulvar region as you grow in utero. Later, typical female secondary sex characteristics like menstruation, rounded hips, and breast growth will emerge at puberty. If you carry XY sex chromosomes, then you’ll develop testes and penis in utero. Typical male secondary sex characteristics like a lowered voice, increased muscle mass, and body hair will emerge at puberty. However, there are several ways in which this process can become complicated, resulting in a range of conditions that we call intersex, defined as people born with sex characteristics, including internal and external genitals, that do not fit typical binary notions of male or female bodies. Most intersex traits are identified at birth, while others may not be discovered until puberty or even later in life.2
Rarely, intersex conditions result from chromosomal abnormalities. For example, Klinefelter syndrome—a condition in which all or some of the cells in the body have an extra X chromosome, yielding the pattern XXY—can sometimes produce an intersex condition. Individuals with XXY chromosomes have a penis and testes, but, in some severe cases, these organs are small and incompletely formed. XXY boys also have attenuated puberty, with reduced body hair and muscle mass and, sometimes, breast enlargement.
Most intersex conditions arise from alterations in hormone signaling in chromosomally typical (XX or XY) individuals. If an XY person carries a mutation that interferes with the function of the androgen receptor or its downstream biochemical signals, this results in a condition called androgen insensitivity syndrome. Depending on the severity of the mutation and its distribution in various cells of the body, the genitalia in XY individuals with androgen insensitivity syndrome can range from fully masculinized (rare) to fully feminized (most common). Secondary male sex characteristics like voice pitch, muscle mass, and body hair distribution can show a similar range of variation. A highly feminized person with XY chromosomes may have internal testes that secrete testosterone but may appear externally typically female, with a vulva and vagina. These individuals are almost always raised as girls with no suspicion of any underlying problem. At puberty, estrogen that is metabolized from testosterone secreted by the internal testes causes female-typical breast and hip development. Menstruation never begins because of the lack of ovaries or uterus, and often this is when the disorder is investigated and diagnosed.3
XY individuals carrying mutations in the gene encoding the testosterone-metabolizing enzyme called 5-alpha reductase have a particularly challenging intersex situation. This enzyme converts testosterone into the more active metabolite dihydrotestosterone, which is crucial for masculinization of the external genitals during fetal development. Its impairment results in external genitalia at birth that are either fully female-typical or of intermediate form. Many but not all people with 5-alpha reductase deficiency are raised as girls. Later, during puberty, there is a marked masculinization, and the affected individuals develop typical male features, including increased body muscle mass, deepening voice, testicular descent, and absence of breast development. In some cases, the external genitals resolve into a semi-functional penis.4 Most but not all people with 5-alpha reductase deficiency who were raised as girls come to identify as male after puberty.
Another type of intersex condition, congenital adrenal hyperplasia, occurs in XX people in whom the adrenal glands secrete an unusually large amount of testosterone due to a recessive gene mutation.5 Again, there is a range of effects, depending on just how much testosterone is secreted and a few other factors. In severe cases, both the external and internal genitalia become ambiguous, often with an enlarged clitoris and a shallow vagina. In milder cases, the genitalia are mostly female-typical, but male-typical secondary sex characteristics often emerge, including body hair, increased muscle mass, and suppressed menstruation.6 To complicate matters further, there are some women who produce unusually large amounts of testosterone but also carry androgen-receptor mutations, so their extra testosterone has no biological effects. The overall conclusion: Traditionally, most cultural ideas have reinforced the notion that biological sex is a clear and immutable binary trait. But in about one in three thousand live births, nature does not draw such a bright line between male and female bodies.7
WHEN MRS. RATJEN GAVE birth to her fourth child on November 20, 1918, there was some confusion in the room. Later, recalling the birth, her husband, Heinrich Ratjen, said, “I was not standing at my wife’s bedside during delivery. Rather I was in the kitchen at the time. When the child was born the midwife called over to me, ‘Heini, it’s a boy!’ But five minutes later she said to me, ‘It is a girl, after all.’” The child had ambiguous genitalia, a penis with a fissure and opening on the underside, and the parents didn’t know what to do. So they followed the midwife’s advice, named the child Dora, and raised her as a girl. Dora attended a girl’s school and wore girl’s clothing, but around age ten she began to question why she felt and looked like a boy. As she told the doctors who interviewed her later in life, her concern mounted as she failed to develop breasts or other female-typical secondary sexual characteristics during puberty. Dora’s first ejaculation horrified her. She felt trapped by her situation and, because of the stifling social norms of her time and place, unable to ask questions or confide in anyone about her condition. Chromosomal and androgen-receptor testing was not yet developed, so we don’t know the genetic details of Dora’s condition. What we do know is that, despite being raised as a female, Dora felt like a male and had a body that was mostly, but not entirely, male-typical.
Fearing detection, Dora avoided dancing or swimming but soon found some solace in her love of sport. By age fifteen she was a regional high jump champion and a contender for the 1936 German Olympic team. When the Nazi government passed over the top German women’s high jumper, Gretel Bergmann, who was inconveniently Jewish, this created a spot on the team for Dora. She may have appeared unusually deep voiced and lean, but her fellow athletes never suspected her secret. Years later, Gretel Bergmann recalled, “In the communal shower we wondered why she never showed herself naked. It was grotesque that someone could still be that shy at the age of 17. We just thought—She’s strange. She’s odd.”
Dora finished fourth at the Berlin Olympics, just out of the medals, but continued to improve in the years to follow, breaking the women’s world record in high jump just two years later. It was on a train home from that victory in Vienna that her secret was revealed. A conductor on the train suspected that Dora was a man dressed in women’s clothing (cross-dressing was illegal in Germany at that time). When the train stopped in Magdeburg, the conductor alerted a policeman to his suspicions and the policeman confronted Dora. After a brief denial, and a presentation of her women’s ID from the recent European championships, Dora admitted that she had always felt like a man, a fact that was then confirmed by medical examinations. “Ratjen openly admits to being happy that the cat has been let out of the bag,” stated a police officer after her arrest. Although fraud charges were initially filed against Dora, they were later dropped when the prosecutor concluded that there was never any intent to deceive, just a terrible misunderstanding caused by well-meaning but confused adults when Dora was a newborn. Dora’s name was changed to Heinrich and he lived out the rest of his life, quietly, as a man.

FIGURE 9. Dora Ratjen at a high jump competition in 1937. Photo from the Bundesarchiv, Bild 183-C10379. Used with permission.
The case of Dora Ratjen is similar to those in the 1960s and ’70s, in which boys born with penile malformation received gender reassignment surgery as infants and have been raised as girls from the time of birth. The misguided theory that drove this ill-fated decision held that infants are a blank slate, and so chromosomal males could be raised to feel female. In fact, this completely failed. As they grew up, almost all reassigned boys reported that they felt male and almost all grew to become sexually attracted to women.8 As a result, the medical community has changed the standard of care from encouraging parents to choose a gender for intersex children soon after birth to encouraging them to wait until the children express a clear gender identity.
DORA RATJEN’S CASE WAS one of several cited by the International Olympic Committee (IOC) when they began mandatory screening of athletes seeking to compete in women’s events. The rationale for this practice has always been to catch male athletes masquerading as women. Remarkably, this has never happened.9 Instead, screening has entirely served to humiliate and exclude people with intersex conditions.
The athletes called the first mandatory femininity screening, which began at the 1966 European championships, “the nude parade.” In this situation, women who did not appear to the assigned panel of male doctors to be entirely female-typical could be called out of line and be made to spread their legs for closer examination. There was never any screening for athletes seeking to compete as male. In 1968, in response to complaints by female athletes, this degrading practice was replaced by a cheek swab to collect cells for chromosomal testing. The new rule held that only persons carrying XX chromosomes could compete as females. Not surprisingly, because sex is determined by a confluence of chromosomal and nonchromosomal factors, this method had problems.
One famous case was a Spanish hurdler named Maria José Martínez-Patiño, who had XY chromosomes and a profound androgen insensitivity syndrome. Her face and body were externally female-typical. She had breasts, a vulva, and a vagina, but no uterus or ovaries. She had always felt herself to be female and was raised as female. Her androgen insensitivity mutation assured that her body could not be affected by the testosterone produced by her internal testes. When her chromosome test was publicized, the response was immediate and brutal. Her medals and records were revoked and she was thrown off the Spanish team, losing her living allowance and her apartment. Her boyfriend left her and strangers pointed at her in the street. Later, she wrote: “If I hadn’t been an athlete, my femininity would never have been questioned. What happened to me was like being raped. It must be the same sense of violation and shame. Only in my case, the whole world watched.”10 She appealed her case, arguing correctly that her body received no competitive advantage from androgens produced by her internal testes. Eventually she won, but the process took three years, and by then her hurdling career was over.11 The strict XX chromosomal standard to compete as a female had clearly failed.
That said, there are still questions about advantages conferred in females who are XY with complete androgen insensitivity syndrome. This is suggested by the observation that the incidence of this syndrome in XY individuals is about 1 in 20,000 in the general population but about 1 in 420 in elite female athletes competing at the Olympic Games.12
We know that SRY-driven development of the testes and subsequent testosterone production is far from the only consequence of having a Y chromosome.13 That chromosome has about two hundred genes, of which at least seventy-two have been confirmed to direct the production of proteins. Some of those genes may confer advantages over XX athletes in certain sports in a manner that is independent of testosterone, perhaps by increasing height or lean muscle mass.14 But, at present, there is no evidence that female XY athletes with complete androgen insensitivity syndrome develop any physical attribute important for athletic performance that is not also present in at least some XX females.
In 2013, the International Olympic Committee announced a new rule: athletes seeking to compete as female could do so only if their blood testosterone levels were below ten nanomoles per liter, with exceptions for cases of androgen insensitivity. If an athlete wanted to compete and exceeded the ten nanomole limit, then she would either have to have surgery (to remove internal testes), take androgen-suppressing drugs to meet the cutoff, or compete as a man. Only 0.01 percent of women have natural testosterone levels exceeding ten nanomoles, so one might imagine that few athletes would be affected by this rule.15 However, the fraction of elite female athletes exceeding the IOC’s testosterone standard is about 1.4 percent, 140 times higher than the general population. This suggests that naturally high testosterone might indeed confer an advantage for some female athletes who do not also have androgen insensitivity. In recent years, several elite female athletes have been barred from competition on this basis, including Caster Semenya, a middle-distance runner from South Africa, and Dutee Chand, a sprinter from India. Chand appealed her ban to the Court of Arbitration for Sport in Lausanne, Switzerland. She argued that she was born and raised as a woman and that she had not doped or cheated in any way. Why should she be forced to undergo surgery or take drugs in order to compete as a woman?
Testifying in support of the IOC’s testosterone standard, champion British marathon runner and sports official Paula Radcliffe contended that elevated testosterone levels “make the competition unequal in a way greater than simple natural talent and dedication.” She continued, “The concern remains that their bodies respond in different, stronger ways to training and racing than women with normal testosterone levels, and that this renders the competition fundamentally unfair.” However, testosterone is far from the whole story; it’s not as if female athletes who win Olympic medals all have naturally high testosterone. One recent study suggested that high testosterone in elite women athletes conferred an average 2 percent advantage for middle-distance runners and a 4 percent advantage for hammer throwers.16 This is a real effect, but is much smaller than the typical 10 to 12 percent gap in performance between elite male and female athletes in clearly quantifiable sports like running or jumping (as opposed to judged sports like mogul skiing or figure skating).17
Chand’s appeal was successful. She competed at the 2016 Olympics in Rio without surgery or testosterone-blocking drugs, but she failed to advance past the first round in the women’s one hundred meters. Caster Semenya competed as well and took gold in the eight-hundred-meter race. These disparate outcomes support the idea that high natural testosterone is not a uniquely potent special sauce for female athletic success. In its 2015 ruling, the court noted, “While the evidence indicates that higher levels of naturally occurring testosterone may increase athletic performance, the panel is not satisfied that the degree of that advantage is more significant than the advantage derived from the numerous other variables which the parties acknowledge also affect female athletic performance: for example, nutrition, access to specialist training facilities and coaching and other genetic and biological variations.”18
That last point is particularly notable. Elite athletes, male and female and intersex, often carry rare gene variants that contribute to their athletic performance. A swimmer with an average physique who is just as dedicated to training as Michael Phelps is unlikely to be able to overcome the physical advantages bestowed by Phelps’s long limbs and enormous feet. Presently, we haven’t identified the gene variants that endow Phelps with his unusual physiology, but they are very likely to contribute strongly to his athletic success.
However, there are some rare cases where making the genetic link to elite sports performance is possible. In the 1960s, the Finnish athlete Eero Mäntyranta dominated Nordic skiing. He won seven medals in three different Winter Olympics. Decades later, genetic testing of his extended family revealed a mutation in the gene coding for the erythropoietin receptor that increased red blood cell growth and survival. As a result, Eero and other affected members of his family had about a 50 percent increase in oxygen-carrying hemoglobin in his blood, a clear advantage in his chosen sport.
Why do we, as a society, easily accept Eero Mäntyranta’s genetic advantage in sport as natural talent but argue about Caster Semenya’s? It’s not that we believe that athletic success should only reflect dedicated effort and not hereditary factors—no one is suggesting, for example, that we ban the tallest basketball players. And it’s not that we believe that fairness requires equal access to nutrition or specialized training; we don’t see anyone proposing an equalizing handicap for athletes from poor backgrounds in sporting events. The reason that sex categories in sports are unusually divisive is because they occupy a point where complicated biology and deeply held cultural ideas about both sex and fairness collide.
NOT ALL ORGANISMS REPRODUCE sexually.19 Asexual reproduction by splitting (also called binary fission) is found in bacteria, certain plants, and some invertebrate animals like hydra—a tiny, drifting, freshwater critter distantly related to jellyfish. Why don’t more animals reproduce by binary fission?20 Wouldn’t it be faster and easier to dispense with mate finding entirely and just divide and make a genetically identical copy of yourself?21
There are two main benefits of sexual reproduction. First, when you inherit two copies of a gene, one from each parent, then the presence of a loss-of-function mutation in one copy is less likely to create a biological problem, as the presence of the other intact copy can usually compensate. Second, and more importantly, the mixing of parental gene variants in each generation of sexually reproducing animals produces more individuality through recombination than can be produced in animals that make exact genetic copies of themselves (clones). Another way to think about this is that genetic diversity in asexual animals can come about only through mutations in DNA, while animals that reproduce sexually have both mutations and the recombination of parental gene variants. Sexual reproduction creates a wider range of genetic variation and thereby forms a broader substrate upon which selective evolutionary pressures can act.22
If you’re going to reproduce sexually, you need a way of ensuring that two cells from the same individual don’t fuse to create offspring—that would defeat the whole advantage of sexual reproduction. So, the specialized reproductive cells, called gametes, must come in two flavors, egg and sperm, and they must be engineered such that egg cannot fuse with egg nor sperm with sperm. This arrangement most commonly requires two different types of organism: males, which make only sperm, and females, which make only eggs.23 Thus, females and males need specialized egg- or sperm-producing organs: ovaries and testes, respectively. Because sperm are small and motile and fertilized eggs are larger and most often develop inside the female, then males and females will also need other specializations of the reproductive tract: the uterus, vagina, penis, and so on. And, in mammals like us, the females need milk glands to nourish the young.
What’s obvious when we look at male and female humans is that, on average, they are different in many ways that are not just directly related to the requirements of copulation, pregnancy, childbirth, and breastfeeding. Adult males are typically taller (figure 10), heavier, leaner, and more muscular, with bigger bones, thicker skulls, and hairier faces. On average, adult females have reduced body hair and higher-pitched voices, with a greater percentage of body fat distributed to breasts, buttocks, and hips. Most of these average differences between males and females are also seen in the fossil record of our hominin ancestors. It’s worthwhile emphasizing that we’re talking about average differences here. Of course, there are individual women who are more muscular than the average man and individual men whose voices are higher-pitched than the average woman.
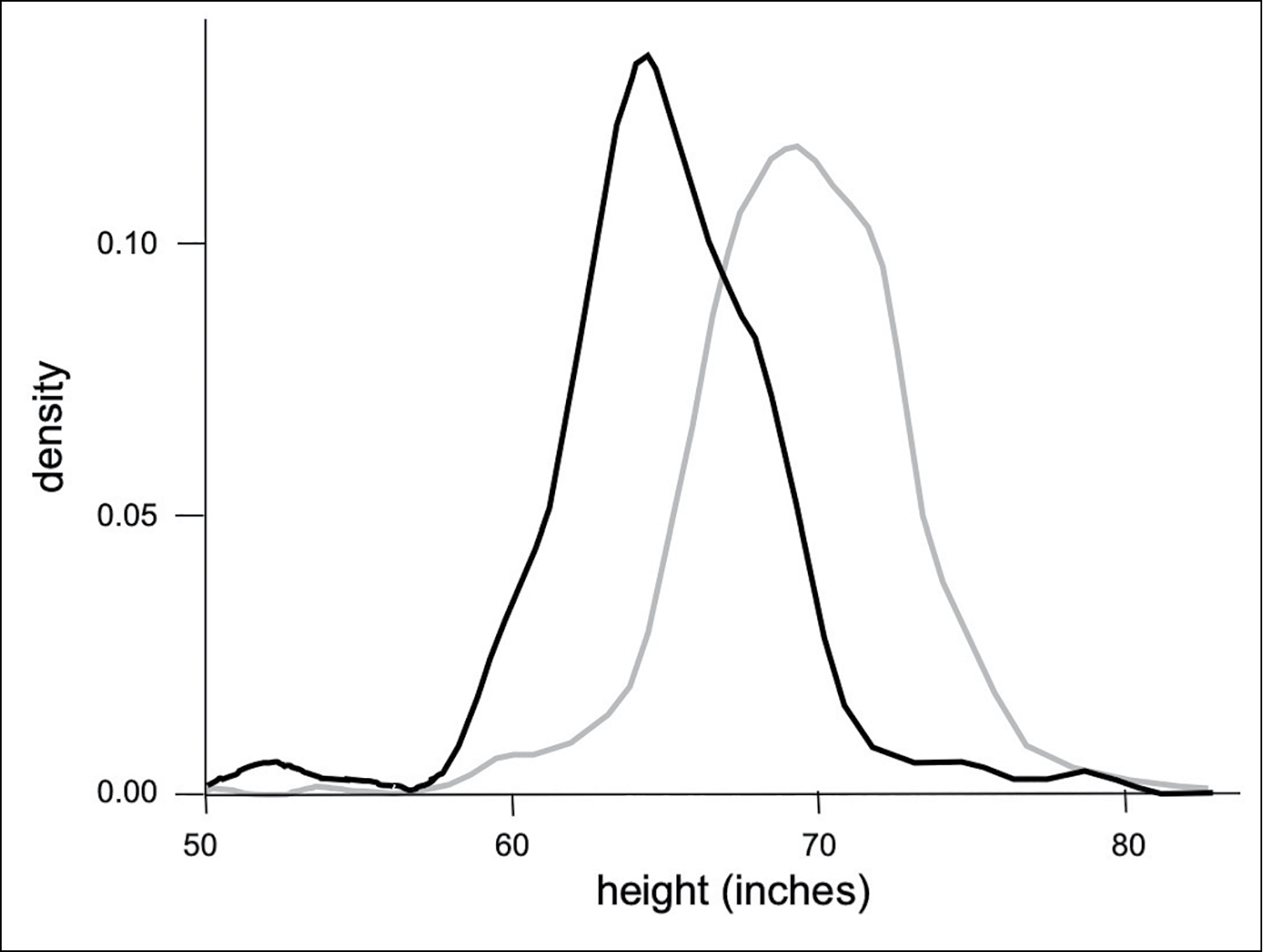
FIGURE 10. This graph of the distribution of adult heights in the United States shows that, on average, men (gray) are taller than women (black). However, there is considerable overlap. Also note that the distribution of height in men is a bit wider, with more men at the extremes and fewer at the average. The degree of overlap between male and female height distributions can be quantified by a statistical measure called standard deviation. The sex difference in height is about 2 standard deviations (d = 2).
The main explanation for how these physical differences between men and women arose comes from the theory of sexual selection, first proposed by Charles Darwin and since elaborated on and refined by Robert Trivers and many others. It holds that males and females don’t mate randomly, but rather seek to mate with those that appear genetically fit, so that their offspring are as healthy and successful as possible. Mating is a big investment, and in most mammals the investment in terms of gametes, pregnancy, childbirth, and child-rearing falls disproportionately on the female. For females, another aspect of this investment is that they are out of the mating business while they are pregnant and for some time after childbirth (such as during breastfeeding), a time when males can potentially mate again. Also, in humans and some other species, the upper bound of the reproductive years is higher for males than females.
All this means that at any given time there are fewer reproductive-age females available for mating than reproductive-age males. There are two main consequences of this difference. First, males must often fight with each other for access to scarce fertile females, hence their larger size and increased bone density and musculature. Second, males must display characteristics that attract females. These may include some of the same traits that allow males to successfully fight or intimidate other males, such as large size or a low-pitched voice. In many species, there are also ornamental male traits that appear to attract females but that are not obviously related to fighting or intimidating other males. The most famous of these, and a favorite of Darwin, was the huge elaborate tail that graces the peacock but not the peahen. Other male signals for sexual selection can involve elaborate behaviors, including calls, gift giving, dances, and nest building.24 In general, females are much less likely to show such ornaments or behaviors because they face less competition for mates.
Sexual selection theory has been used to explain not just the physical differences between men and women but also both sexual and nonsexual behaviors. It has been claimed that sexual selection has driven men to be promiscuous, risk-taking, aggressive, and violent, while women, with their greater investment in parenthood, are choosy about sexual partners, cooperative, and nurturing. Is this idea true, or is it just a story that has been concocted to reinforce traditional, socially constructed roles for men and women? Whenever there is an explanation that could be used—and indeed has been used—to justify historical and ongoing oppression of women by men, it behooves us to subject it to careful scrutiny.25
For sexual selection to operate, it requires that some individuals be more successful in mating and having children than others. If everyone were equally successful at making babies, then there would be no basis to weed out the sexual losers. If sexual selection theory is right, then the variance in reproductive success among males should be greater than that among females, reflecting that males must compete to mate with a limited pool of available, fertile females. The first experimental tests of this idea were performed by geneticist Angus Bateman in the 1940s, using fruit flies, and appeared to show that males do indeed have a greater variation in reproductive success than females. This happens for two reasons. The first is that males show more variation in the number of sexual partners than females. The second is that, when males mate with more partners, they increase the number of offspring produced per additional partner at a higher rate than when females do.
In recent years, Bateman’s experiments have been justifiably criticized for flaws in both experimental design and statistical analysis.26 For some scholars, these critiques are sufficient to toss out the entire edifice of sexual selection. In their view, Bateman’s experiments are the basis for sexual selection theory, and so their invalidation destroys the foundation on which subsequent experiments have been built. But this isn’t really true. Subsequent experiments and field observations have been inspired by Darwin and Bateman’s theory, but they do not rely on Bateman’s experimental findings and so should stand or fall on their own merits.
Bateman’s predictions have now been tested in a wide variety of animals, ranging from mollusks to insects to fish to mammals. The overall result is that, in the sixty-two different species examined, most but not all males showed higher variance in reproductive success and also more reproductive success as sexual partners were added.27 Even more informative are the special cases where these results did not hold. For example, species in which the males provide most parental care—like the seahorse or the pipefish (males carry the pregnancy in a special brood pouch, outdoing even the most dedicated Brooklyn hipster dad)28 and a shore bird called the wattled jacana (males incubate the eggs and raise the chicks)29—show stronger sexual selection in females, and the females tend to be larger and more ornamented.30 These are truly the exceptions that prove the rule. Bateman’s experiments may have been flawed, but the Darwin/Trivers/Bateman hypothesis has survived, albeit with certain important caveats.
Another exception that runs counter to the ideas of Darwin, Trivers, and Bateman are species in which females gain reproductive success from mating with multiple males.31 There are many group-living species in which one or a few dominant females are the only ones to successfully breed. In some cases, like the marmoset or the mole rat, the dominant female physiologically suppresses the reproductive cycles of her subordinates. In others, the subordinate females are allowed to mate but their newborns are then killed by dominant females, as in wild dogs and meerkats. In yet other animals, like savanna baboons, lions, and langur monkeys, most females mate with several males. The anthropologist Sarah Blaffer Hrdy, who first described this behavior in langurs, suggests that such female promiscuity may serve to muddle the paternity of baby langurs, thus reducing the chance that the dominant male will kill newborns that he thinks he did not sire.32 In recent years, as DNA testing has come into wider use, it has become clear that, in many species that were thought to be monogamous, both males and females are getting a bit on the side. The original Darwinian notion that females are always sexually choosy is incorrect; it holds for many species, but there are more than a few exceptions. Most importantly, the cases where the Darwin/Trivers/Bateman model fails are not random, but fall into particular situations that are well explained by male parental investment, female social structure, infanticide, and a few other factors.33
WE’VE JUST SPENT SOME time going over the evidence in critters for and against the hypothesis that sexual selection operates more strongly in males than in females. That is useful to set the stage, but what we really care about are humans. In considering whether sexual selection can explain some of the structural and behavioral differences between men and women, it’s worthwhile to note that, among our mammalian cousins, humans are sexually unusual in that we exhibit long-term biparental care, social monogamy, accurate assignment of paternity, and concealed ovulation.
When a human baby is born, its brain is about four hundred cubic centimeters in volume, about the same as that of an adult chimpanzee. It develops at an enormous rate until age five and at a slower rate until about age twenty, when it finally reaches its mature state, with a volume of twelve hundred cubic centimeters. As any mother will tell you, that four hundred cc head just barely fits through the birth canal, and sometimes it doesn’t quite make it. In fact, death during childbirth is almost a uniquely human phenomenon. We need that huge head to house our big brain and be so clever. Why, then, didn’t the human birth canal just evolve to be wider, to accommodate big ol’ human heads? The most likely reason is that it would require reconfiguring the female pelvis in a way that would interfere with upright posture and so was an evolutionary nonstarter.
Then, the baby is born and is helpless for a really long time. Humans have the longest childhood of any animal. There’s no other critter where a ten-year-old cannot reliably make its own way in the world. This means that when the father contributes care, protection, or resources to his children, it’s a huge help. By contrast, in most animals, the male is entirely out of the picture, makes no contribution to the care of the offspring, and it’s OK.
While many female animals advertise their fertility with swellings, odors, or stereotyped behaviors, human ovulation is mostly (but not entirely) concealed. Despite what perfume manufacturers have told you, we have yet to find any human pheromones, much less those that signal ovulation to males (more on this in chapter 5). This means that most human sex is happening outside of the female’s fertile time and is thus recreational, not reproductive. It also means that if a man is to be confident in his paternity, he must monopolize the female sexually throughout her cycle. Not to take all the romance out of the deal, but this is part of the reason why heterosexual marriage (with all of its variants) is such a widespread cross-cultural institution.
This monogamous (or at least serially monogamous) arrangement seems to work out. DNA testing has shown that, contrary to what you might have gleaned from watching Jerry Springer, the assignment of paternity to a woman’s husband or long-term partner is accurate about 98 percent of the time. This finding holds widely across cultures.34 False paternity happens, but it is not widespread. It’s worthwhile to keep in mind that all of these unusual aspects of the human mating system (long-term biparental care, social monogamy, accurate assignment of paternity, and concealed ovulation) are factors that would appear to reduce sexual selection in men.35
By using genealogical records and surveys, we can count people’s children and thereby estimate the variance in reproductive success and how it differs between men and women. On average, in a recent meta-analysis of eighteen different human populations across the globe, males did have a larger variation in reproductive success than females, consistent with the Darwin/Trivers/Bateman model.36 But dig a little deeper into the data and some fascinating details emerge. First, the variance of the effect across populations was large. The ratio of male reproductive success variance to female reproductive success variance was 0.70 in Finns but 4.75 among the Dogon people of Mali. Second, in general, the populations with monogamous marriage systems like those in Finland, Norway, the United States, and Dominica had ratios near 1, indicating no difference in sexual selection in men and women. The populations with polygynous marriage systems (one man, multiple wives), as found among the Dogon, the Aché of Paraguay, and the Yanomami of Venezuela, had much higher ratios, indicating greater sexual selection in men.37 The most likely conclusion is that sexual selection has indeed operated more strongly in men than in women for much of our human history, and that has given rise to increased height, muscle mass, and physical aggression in males, largely from competition between them.38
It may well be that contemporary polygynous societies are a better model of ancestral human social groups than monogamous ones. But that difference is likely eroding as monogamous marriage slowly comes to predominance across the world. That said, even men in most monogamous societies have somewhat higher variation in reproductive success than women, in part because divorced men are more likely than divorced women to remarry and start a second family.39
If we look at an affluent, mostly socially monogamous population with access to contraception, such as young adults in the United States, are women, on average, choosier about their sexual partners than men? A study on this topic was conducted by psychologists Russell Clark and Elaine Hatfield.40 Usually the part of a scientific paper where the methods are described is mind-numbingly dull, but I was amused by Clark and Hatfield’s:
The confederates stood on one of five college quadrangles, and approached members of the opposite sex, who were total strangers.… Once a subject was selected, the requestor approached him/her and said: “I have been noticing you around campus. I find you to be very attractive.” The confederate then asked subjects one of three questions: “Would you go out with me tonight?” “Would you come over to my apartment tonight” or “Would you go to bed with me tonight?”… The requestor carried a notebook which had one of the three requests written on a separate page. The type of request was randomly determined. After the selection of a subject, each requestor flipped a page in the notebook to see what type of request was to be made.… Subjects were not approached between class periods or during rainy weather. Subjects were debriefed and thanked for their participation.41
This much-cited study was performed at Florida State University in 1978 and was repeated with the same design in 1982. The results of the two studies were almost the same, so I’ll report those for the 1978 version: 50 percent of the men and 56 percent of the women agreed to a date and, strikingly, 75 percent of the men and 0 percent of the women agreed to go to bed with the requestor. Yes, that’s right. More men were willing to have sex with a stranger than to go on a date. There have since been several replications of the main result in different countries.42 This study became so well known that the requestor’s lines even became the basis for the hit song “Would You…?” by the British electropop group Touch and Go in 1998.43 One can argue with some details of the experimental design and about the size of the male-female difference, but the overall finding is clear: on average, men are much more willing to have sex with strangers.
Clark and Hatfield suggest that the basis for this striking difference between men and women is that, while women and men are equally interested in sex with strangers, women are holding their desires in check out of a fear of violence, impregnation, or social disapproval. Science historian Cordelia Fine, in her book Testosterone Rex, amplifies these points and adds that it may be rational for women to decline stranger sex with men because there is a low chance (11 percent) of them experiencing an orgasm in a casual hookup situation.44
So let’s do an optimistic thought experiment, in which sexual violence is rare, so-called slut shaming has been banished, and women regularly enjoy orgasms with casual sex partners. Would this situation then reveal that, on average, women and men have equal interest in sex with strangers? We can’t really know, but I suspect that men would still be somewhat more interested in casual sex. After all, masturbation is a safe, private, and reliable route to orgasms for both men and women. Yet, even in anonymous surveys, women across the lifespan report less frequent masturbation than men.45 Among lesbians, the fear of unwanted pregnancy and orgasm-free sex is reduced, but, on average, lesbians report about the same amount of interest in and practice of sex with strangers as straight women (and much less than gay men).46 There is no question that we have lived and continue to live in patriarchal societies, with the attendant physical and social risks to women engaging in sex with strangers. But the statistics on women’s masturbation and lesbian sexual behavior lead me to suspect that, on average, there is a significant biological difference between men and women in casual sexual behavior that has been driven by sexual selection and would remain even if those traditional risks to women were diminished.
There are average differences between the behavior of men and women in the nonsexual realm, but generally the effects are few and of only moderate size. Most measures of personality traits, social interaction, and cognition do not show significant average differences between women and men.47 Also, as Thekla Morgenroth and her coworkers have noted, we need to be vigilant about cultural assumptions creeping into such assessments. For example, men are culturally stereotyped as being more risk-taking, and this position has been supported by survey data.48 But if the behaviors we choose to measure to determine risk-taking are already male-associated—like wagering, drug use, and participation in dangerous sports—but do not include female-associated risks—like childbirth (which is, statistically speaking, much riskier than extreme sports) and organ donation (which women do at a higher rate than men)—we’ve biased the outcome.49
Both observational studies in the lab and personality assessments show that men are, on average, more physically and verbally aggressive than women. But the effect is small, about 0.6 standard deviations.50 Women, on average, appear to be more empathetic (about 0.8 standard deviations), as measured by surveys and observational assessments. Recall that the sex difference in average height was about 2 standard deviations, so aggression and empathy, measured in these ways, are notably smaller effects. However, the assessments by psychologists probably underestimate the real-world sex difference in aggression, as 96 percent of homicides worldwide are committed by men and 78 percent of homicide victims are men (not counting wars).51 This is unlikely to be due to mere socialization of human males, as a similar sex bias was found in a study of eighteen chimpanzee communities, where 92 percent of killings were committed by males and 73 percent of the victims were males.52 In the cognitive realm, while there is no significant average difference between women and men in IQ test scores, women tend to do a bit better than men on tests of verbal fluency (0.5 standard deviations) and men do a bit better on tests of spatial perception and mental rotation of objects (0.6 standard deviations).53
By far the largest difference between human males and females in nonsexual behavior is found in children’s play. Children spend most of their waking hours playing and, on average, boys prefer to play with object toys like trucks, while girls prefer to play with social toys like dolls. And, on average, boys engage in more rough-and-tumble play. These distinctions are present early in life, are broadly cross-cultural, and, unlike most behavioral differences between men and women, they are large. A composite measure of children’s play yields a sex difference of about 2.8 standard deviations—even larger than that for adult human height. How do these notable differences in children’s play come about? Traditional explanations from developmental psychology hold that children acquire sex-typical patterns of behavior through social learning. Indeed, studies have shown that children tend to pick toys that have been designated for their sex by adults or that they have seen others of their sex choose. From the moment of birth, boys are awash in blue, dinosaurs, and trucks, while girls are inundated with pink and dolls. There’s no question that social learning is an important influence on children’s play, but is it the whole story?
There are several ways to test the hypothesis that sex differences in children’s behavior also have an innate, biological component. One is to look at newborns, who have not yet had the opportunity to be influenced through social learning. In a well-known study by Jennifer Connellan and coworkers, 102 babies (average age = 37 hours) were presented separately with either a real human face (Connellan’s) or a mobile consisting of jumbled bits of a photo of her face with a small ball attached, and their reactions were videotaped. The tapes were then cropped to show just the eyes of the baby and were analyzed by independent experimenters who did not know the baby’s sex. The result was that boys looked at the mobile more than girls did, and girls looked at the face more than they looked at the mobile.54 This result has been taken to mean that some behavioral average sex differences are, at least in part, biological in origin. This is an important claim and one that cries out for careful replication.55
If the observed differences in children’s play behavior have a biological component, then it is likely to result from the exposure of the male nervous system to higher levels of androgens in utero. Indeed, childhood (before puberty) is a time when there are almost no gonadal steroid hormones circulating in the body, so the die must have been cast before then. It is not ethical to manipulate fetal hormones in early development just for the purpose of research, so the fallback has been to study naturally occurring disorders of fetal hormone signaling. Girls with congenital adrenal hyperplasia, who were exposed to elevated concentrations of androgens in utero, show decreased female-typical play and increased male-typical play. Similarly, girls whose mothers were prescribed androgens during pregnancy for medical reasons showed increased male-typical play, including toy choice. Tellingly, the opposite effects on play were produced by fetal exposure to androgen-blocking drugs.56
These findings argue strongly for an organizing role for early androgen exposure in determining sex differences in behavior. But we must recall that girls with increased fetal androgen signaling, from either congenital adrenal hyperplasia or maternal androgens, will often have partially masculinized external genitalia. It has been suggested that this appearance will cause the parents to treat these daughters more like sons, thereby affecting their play behavior through social learning.57 However, observational studies indicate that this is not the case: in fact, if anything, parents tend to encourage female-typical behavior to a greater degree in girls with ambiguous genitalia as a compensatory mechanism.58
If normal fetal hormone exposure influences play styles, then we might be able to see sex differences in play behavior in other mammalian species. Indeed, in both rats and rhesus monkeys, males show much more rough-and-tumble play, consistent with preparation for later competition for mates. And in both of these species, treating females with androgens in utero or shortly after birth causes them to play like males.59 Remarkably, a study of vervet monkeys found sex-typical toy preferences similar to those in human children, with female play appearing to involve preparation for childcare behaviors (figure 11). These preferences are seen even though vervet monkeys lack the social transmission of toy preference found in humans. Indeed, sex-based preferences in the young monkeys were evident with the very first exposure to the toys and without observation of other monkeys interacting with them.60

FIGURE 11. Reminiscent of human children, female vervet monkeys prefer playing with dolls (left), while male monkeys prefer object toys like cars (right). The female monkey seems to be performing anogenital inspection of the doll, similar to the way vervet mothers inspect their infants. From Alexander and Hines (2002). Used with permission of Elsevier.
One explanation for these findings, consistent with the vervet monkey results, is that early androgen exposure alters the brain in a way that determines a preference for male-typical play. However, we also know that boys model their behavior on other boys and men, and that girls model their behavior on other girls and women. So perhaps what androgens are really doing is influencing the brain to attend to and imitate behaviors of males, a form of hormone–social experience interaction.61
THERE ARE MANY DISORDERS of the nervous system that have different incidence or severity between males and females. These include diseases that emerge early in life, like autism spectrum disorder, early onset schizophrenia, dyslexia, stuttering, ADHD, and Tourette’s syndrome and related tic disorders. Sometimes these effects can be large: autism spectrum disorders are about five times more likely to occur in boys and Tourette’s is about three times more likely. Other neuropsychiatric diseases with sex differences typically manifest after puberty and include anorexia nervosa, multiple sclerosis, late-onset schizophrenia, Parkinson’s disease, and major depression. Again, sex-related incidence can vary from large (anorexia is about fourteen-fold higher in girls) to moderate (on average, the onset of Parkinson’s is about two years later in women). And it can be complicated: multiple sclerosis in about four-fold more common in women but is typically more severe in those men who are affected. This pattern does not allow for a simple explanation, like a multiple-sclerosis-protective factor encoded on the Y chromosome.62
Like any trait, we should not assume that the sex differences in neuropsychiatric diseases are all caused by innate biology. It’s extremely likely, for example, that the much higher incidence of anorexia among females results, in large part, from societal objectification of women’s bodies. And the higher incidence of some adult-onset neuropsychiatric disorders in men, like Parkinson’s disease, might relate to higher exposure to environmental toxins in certain industrial workplaces where more men are present.63
Similarly, the measures we use to calculate the incidence of some of these diseases depend on the willingness of people to seek treatment and hence be counted in the statistics. Women are treated for major depression at a higher rate than men, but it is unclear if that’s because women suffer more depression or because, on average, they are more willing to seek help from a doctor or psychotherapist. And it could easily be that the rate of depression in women (and intersex people) is higher because of ongoing oppression, in much that same way that it is higher among poor people compared to the middle class.64
The five-fold higher incidence of autism spectrum disorders in boys and their typical emergence in early childhood has suggested to some, most notably psychologist Simon Baron-Cohen, that exposure to androgens in utero is a significant risk factor for that condition. In his view, informed by experiments measuring testosterone in samples of amniotic fluid and then following those children through early development, autism spectrum disorders are produced when there are unusually high levels of androgens present in utero, resulting in a case of “extreme male brain.”65 At present, however, there has been a notable failure to replicate this basic finding.66 It may still be that Baron-Cohen’s idea is correct, but that testosterone levels measured in amniotic fluid at a single time point are just not a good indicator of developmental androgen exposure.67 Or it may be that variation in other (SRY- and testosterone-independent) genes on the Y chromosome are most important for conferring the increased autism risk in boys. However, to date, the candidate gene variants contributing to autism risk are not located on the Y chromosome.
TOGETHER, THESE RESULTS—FROM SEXUAL behavior to children’s play to neuropsychiatric disease susceptibility—indicate that there are some disorders and behaviors that are, on average, significantly different between men and women, and there are a subset of those that appear to have a strong biological component. If this is true, then we would expect to find some important average differences in the function and perhaps the structure of men’s and women’s brains. The problem that currently limits all human brain research is that we can rarely do invasive studies on living humans.68 We can take DNA samples from cheek swabs, look in detail at the cellular structure of cadaver brains, and use brain-scanning machines on living people. But these machines are very crude tools. They cannot see individual neurons, nor can they measure the electrical activity of individual neurons or the strength of connections between them. For those crucial experiments, we need lab animals.
In rats, mice, and monkeys, we have solid evidence that, in many different brain regions, there are important sex-related differences in the function of neural circuits. For example, some neurons fire nearly twice as fast in females as in males. Some types of synapses are more easily changed by experience in males. Other neurons have their electrical or chemical properties change over the course of the female estrus cycle in an estrogen-dependent manner. There are more and more of these examples that are accruing as researchers begin to pay attention to sex differences.69 Importantly, these sex-related differences are not only found in brain regions known to influence sexual behaviors. There are also important differences in the circuits involved in many functions, including motor control, memory, pain, stress, and fear. Sometimes, these differences can be seen in the size of a brain region: There’s an area called the medial preoptic area (MPOA) that’s involved in sexual behavior and is, on average, larger in males than females. Another region, the anteroventral periventricular nucleus (AVPV), is larger in females, and it’s electrically inhibited by a nearby region called the bed nucleus of the stria terminalis (BNST), which, in males, sends ten times more inhibitory nerve fibers to the AVPV. Sorry for the alphabet soup here. The precise names of these regions aren’t crucial. The point is that there are many brain circuits that are, on average, functionally different in male and female mammals. What’s more, these differences can be changed by experimental manipulation of hormones during development: Males with impaired androgen signaling will have a smaller, female-like MPOA. Females with disrupted estrogen signaling will have a smaller, male-like AVPV. Importantly, while there are many locations in the brain that are, on average, sexually dimorphic in their electrical, chemical, or connection-map properties, there are many others that aren’t. These types of experiments are fairly new, and our understanding of sex differences in fine-scale brain function are at an exciting but early stage.70
In humans, current technology does not allow us to make these cellular-scale measurements in the intact brain, but there’s every reason to believe that the story is similar. For example, the human equivalent of the MPOA is called the third interstitial nucleus of the hypothalamus (INAH3) and, on average, it is also larger in males than in females (in both sexes it’s small enough that it can only be measured in human autopsy tissue). In a recent brain-scanning study of 2,838 adults, on average, the gray matter volume of the amygdala (an emotion-processing center) and the hippocampus (a center for memory of facts and events, particularly spatial memory) is slightly larger in males, while that of the prefrontal cortex (involved in self-control and executive function) and the posterior insula is slightly larger in females.71 An important limitation of these studies is that the brain is plastic, and certain experiences can cause various brain regions to slightly shrink or swell (as we discussed in chapter 3). So, these sex differences in the size of adult human brain regions reflect inborn sex differences convolved with the plastic effects of different experiences of life as a woman or a man. That’s why it’s particularly useful to study fetal brains, before they are impacted by the long reach of culture. One recent study scanned the brains of 118 fetuses during later pregnancy (twenty-six to thirty-nine weeks gestation) and found significant differences in the resting state connectivity between males and females.72 This interesting finding awaits replication.
Recently, neuroscientist Daphna Joel and her colleagues posed the question: If there are truly different male and female brains, can we look at a brain scan and accurately determine sex? After all, aside from a small group of individuals with intersex conditions, we can do this easily by looking at the external genitals. To address this question, they examined a large data bank of brain scans of adult women and men. They measured the sizes of many structures and the connections between them and found extensive overlap between the distributions of females and males. In addition, they found that most brains are composed of unique mosaics of features, some more common in females than males, some more common in males than females, and some equally common in both. They concluded that “human brains do not belong to one of two distinct categories: male brain/female brain.”73
In my view, there are several problems with this conclusion. First, it’s a mistake to make a fundamental statement about brains based on an imaging technology with such poor resolution. Their conclusion was not, “With this limited view afforded by present-day brain scanners, we cannot accurately categorize human brains as male or female.” Rather, it was a definitive statement about the underlying biology. Second, as has been pointed out by others, the failure of Joel and coworkers to distinguish male and female brains from scans was due to an underpowered statistical design with insufficient comparisons of the various brain measurements. When Adam Chekroud and his colleagues attacked a similar data set of brain scans and applied appropriate multivariate statistics, they were able to classify scans as male or female with 93 percent accuracy.74 Kevin Mitchell has pointed out that this problem is akin to face recognition.75 If we take any particular characteristic of a face—the size or shape of the nose or the bushiness of the eyebrows—one will not be able to sort male faces from female ones. Even several of these measurements combined might not be sufficient to make an accurate determination. Yet, when we look at a human face, taking into account many different parameters, we can ascertain sex quite well—similar to the 93 percent accuracy of Chekroud’s method. It’s not as accurate as a peek in the underpants, but it’s quite close.
Third, and perhaps most importantly, I find the whole construction of the question unhelpful. It turns out that we can determine sex reasonably well from an individual’s brain scan (and some future high-resolution brain scan will undoubtedly be even better). But, even if we couldn’t, so what? The important point is that there are some true average sex differences in behavior and brain function between men and women, and a subset of those are likely to be biologically influenced and hence subject to evolutionary forces. The accuracy with which we can assign sex based on an individual’s brain scan using present or future imaging technology is unimportant to the larger issue of average sex differences in the human brain.
SO FAR, WE’VE BEEN talking about biological sex as determined by sex chromosomes, variations in hormone signaling, and the random nature of development: female, male, or intersex. Now, let’s switch to talking about gender. According to the World Health Organization, “Gender refers to the socially constructed roles, behaviors, activities, and attributes that a given society considers appropriate for men and women.” Being socially constructed, gender identity will vary across cultures and times—maleness means something different in modern Japan than it did in Spain during the Middle Ages. While sex is a biological phenomenon that doesn’t always sort easily into male and female, gender is even more variable.76
Most people are cisgender, meaning that their biological sex and their gender identity match. Recall that, on average, men and women differ in height by about 2 standard deviations. By contrast, men and women differ in gender identity by about 12 standard deviations. That’s another way of saying that most people identify with the biological sex they were assigned at birth. But for about 0.6 percent of adults in the United States (about 1 in 167),77 it’s more complicated; this situation is called transgender. Some transgender people feel themselves to be the opposite of their sex assigned at birth. Others feel no particular or lasting association with any gender and thereby identify as nonbinary, agender, gender-fluid, or one of the many other terms (including about seventy now available in the gender box on the Facebook registration page).
If one’s gender identity doesn’t match one’s biological sex, this often, but not always, produces feelings of gender dysphoria. Most transgender people report feeling gender dysphoric at some point during childhood, although for others it can manifest in puberty or adulthood or not at all. The intensity of gender dysphoria can range from mild to strong (the latter often accompanied by major depression or thoughts of self-harm). Depending on individual inclination, opportunity, and cultural practice, gender dysphoria may motivate its sufferers to seek sexual reassignment surgery or hormone therapy.78
Neuroscientist Ben Barres, in a letter written to me and many other colleagues as he began his transition to male in the mid-1990s, described his own experience with gender dysphoria:79
Ever since I was a few years old I have had profound feelings that I was born the wrong sex. As a child I played with boy’s toys and boys nearly exclusively. As a teenager I could not wear dresses, shave, wear jewelry, makeup or anything remotely feminine without extreme discomfort. I watched amazed as all of these things came easily to my sisters. Instead, I wanted to wear male clothing, be in the Boy Scouts, do shop, play sports with the guys, do auto mechanics and so forth.… It is not that I wish I were male, rather, I feel that I already am.
Because these terms can be confusing, I think it’s important to make an explicit point: about 0.03 percent of people are intersex, but about 0.6 percent of adults identify as transgender. This means that about 95 percent of people who identify as transgender have normal external and internal genitals. However, they feel that those genitals are not a match (or a consistent match) to their gender identity assigned at birth. So, if most transgender people have sex-typical genitals, how does gender dysphoria come about?
One possible clue comes from the converse statistic: although only about 5 percent of transgender people are intersex, intersex people are unusually likely to switch gender identity at some point in their lives. Recall the condition called 5-alpha reductase deficiency, in which XY people are unable to produce the key fetal masculinizing signal dihydrotestosterone, leading some of them to develop female-typical external genitals and be raised as girls, only later to undergo male-typical puberty. Remarkably, among those raised as girls, most (seventeen out of eighteen in one study) will change to live as men after male-typical puberty. It’s not absolute, however. For example, two affected siblings who carry the same mutation, and hence the same impairment of enzyme function, have chosen to live as different sexes, one as a man and the other as a woman.80
Since most intersex conditions result from alterations in steroid hormone signaling, then perhaps gender dysphoria results, at least in part, from some alteration in these processes within the brain. One possibility is that there are alterations in fetal steroid hormone signaling throughout the body that are below the threshold necessary to alter the genitals, either internal or external, but are sufficient to change brain circuits that influence gender identity. Another possibility is that steroid hormones are produced in the fetal or newborn brain and can have local effects there. Indeed, estradiol (a particular form of estrogen) can be locally synthesized in some particular brain regions and has been shown to act locally, but brain-derived estradiol has little or no effect in the rest of the body.81
There is some evidence from twin and sibling studies that gender dysphoria has a heritable component, with one estimate yielding 62 percent of variance attributed to genes.82 This number should be taken as a rough approximation, however, as the sample size was small relative to the incidence of gender dysphoria. To date, there is no convincing evidence implicating variation in any particular gene in gender dysphoria. As with most behavioral traits, the heritable component of gender dysphoria likely results from variation in many different genes acting together or in particular combinations.
Investigating the potential neural basis of gender dysphoria is challenging. It’s not amenable to study in laboratory animals. And many people with gender dysphoria who volunteer to be studied in the lab have already been taking hormone therapy or have had surgery, so it’s unclear if differences in their brain scans, for example, are a result of such treatment or predate it. One laboratory has suggested that gender dysphoria might be related to the size of the BNST brain region. In adult males, the BNST region is enlarged. Studying a small sample of autopsy tissue, researchers found that the BNST tended to be smaller in male-to-female transgender people compared to cisgender men.83 However, one problem with such a theory is the fact that the difference in BNST size between males and females seems not to emerge until adulthood, and most transgender people report experiencing gender dysphoria in childhood.84 Furthermore, these interesting findings have yet to be replicated by another lab. There are also a number of brain-scanning studies examining transgender adults, but the results are inconsistent and the number of subjects is small.85 In my view, it is likely that there are variants in the function of the brain that contribute to, but do not entirely explain the onset of, gender dysphoria, but, at present, we do not know what they are.
GENDER IS AN OVERWHELMING cultural force that pervades every aspect of human life from the forceps to the stone. And, as we discussed in chapters 2 and 3, we are built to be changed by experience, and so living in a culturally gendered world will inevitably influence our bodies and brains. It’s no secret that, even now, in societies that purport to be egalitarian, women, intersex people, and trans people are routinely objectified and denied equality of opportunity. There has been a long history of denying equality of opportunity for women by claiming innate differences in male and female brains and minds. These claims are not just Victorian artifacts. They persist to the present day. So, as a political device, it’s appealing to imagine a blank-slate brain, in which differences in the behavior of men and women are solely inscribed by a patriarchal culture. As a lifelong committed feminist, I would be very happy if that were true. But it isn’t.
Many claims that have been made, about Martian men and Venusian women and the like, are unsupportable nonsense. In most measures of cognition and personality, men and women are indistinguishable. But, as we have discussed, there remain average biological differences in the brains and behaviors of men and women that are real and significant, and a subset of those are innate. There’s much to be learned about innate sex differences in the human brain, and such work must be critiqued and debated and held to the highest standard. But the trend is clear: as we are able to probe at finer and finer scales, to the level of cells, biochemistry, and electrical signals, more average sex differences in brain function are revealed.
Importantly, these differences, whether neural or behavioral, are effects on populations. Even large sex differences, like propensity for physical violence or incidence of autism, do not allow us to make predictions about individuals. There are individual violent women and there are girls on the autism spectrum, even though they are less prevalent than boys. There are men who have multiple sclerosis. While we (and other animals) are built to stereotype and prejudge people based on their sex (as revealed in implicit-bias experiments), we all need to work to eliminate such prejudice against individuals in our thoughts.
Here’s what I believe with all my heart. The argument in favor of sex and gender equality, including intersex people and a spectrum of gender identities, must be a moral argument about the way things should be, not a biological argument about the way things are, either in humans or in critters. If tomorrow there were definitive proof of certain inborn average differences in the brain function of women or intersex people or trans people, that would not be an argument for maintaining a system that denies them equal opportunity. The moral argument for equality of opportunity for all can and should accommodate sexual selection theory and certain innate differences between female, male, and intersex people’s brains and behavior. Indeed, it is too important a goal to saddle with the ultimately indefensible argument for a blank-slate mind.