4
ORPHEUS
Cadences
At one point in the previous chapter we encountered a collection of birds in an arena of tall trees behind the house I live in. The tensions between these animals, extending through the long sightlines of air, were illustrative of life on land—out of the sea, anyway. Another conspicuous feature of the behavior of those birds, and many others, is the noise.
The sounds in this bowl-like aviary mostly come from parrots of various species, along with honey-eaters and a few others. From the Rose-breasted Cockatoos (or Galahs), a relatively gentle cheep; the Sulfur-crested Cockatoos issue a multi-frequency wrenching shriek. King Parrots emit single soprano notes of a half-second or so when they are sitting, and a great mixed-up blast of sound when flying. Among these come the pretty two- and three-note cadences of Crimson Rosellas, placed and repeated (forever auditioning for a Philip Glass opera), along with the laser-like cry of Rainbow Lorikeets, matching their neon riot of color, when visiting from lower altitudes. And sometimes present but elusive, in the background, sits a beautiful charcoalgray parrot called a Gang-gang—gray except that the males’ heads are coral-red. They have a call like the squeak of a rusted gate, or a cork being pulled from a bottle.
When all are going at once, the air fills with shrieks, cheeps, a few soprano cadences, and, as if in celebration, corks being pulled from wine bottles, over and over again.
Avian Life, Communication
Birds are an escape pod from—or for, rather—the dinosaurs. Their evolution began in the Jurassic, the middle part of the dinosaur reign, about 160 million years ago. A group branched off from other dinosaurs and slowly explored an unusual body plan—small, lightweight, feathered, warm-blooded, and with wings. Their evolutionary path included animals with much of a modern bird form, but claws on their wings and a snout with teeth. Feathers, including the modern “vaned” form as opposed to scrappy tufts, arose early, even before animals that looked in other ways much like birds. Early birds diversified in the Cretaceous, among the flowering plants of the previous chapter, and some of these small, high-energy, mobile dinosaurs then survived the mass extinction that wiped out their larger cousins, along with much else, about 66 million years ago.
It’s not that the huge tyrannosauruses and their like died while birds all snuck through. In a chart representing this period, a lot of evolutionary lines within the birds end abruptly with the asteroid as well. But some birds did make it through, and these were the only dinosaurs that did. Once on the other side in the post-asteroid world, they embarked on a rapid, exuberant multiplication.
The evolutionary relationships for birds that are alive now look in some ways as you might guess them to be. A deep branching, very old, separates large flightless birds like ostriches and emus from just about all the rest. Then, within the bigger branch, a second split leads to ducks, geese, and other fowl on one side, and another diverse collection on the other side. We walk onward through a great cluster of branches (pelicans, hawks …), and then encounter a huge group called the passerines. This group contains more than half of all bird species, including robins, crows, and the like. Right alongside them, as a small branch of their own, are the parrots. Much of this cluster unfolded quite quickly after the asteroid, as birds took over a good part of the world they had inherited.
The diagram on the next page is the first of several “tree of life” diagrams in this book, each showing fragments of the network of genealogical relationships among living things. In each case, time runs up the page to the present day. Segments are not drawn to scale and many branches are omitted. Labeled branches sometimes represent just a few species (or one), and sometimes a larger group. This first drawing also includes the asteroid impact putting an end to Tyrannosaurus rex and the Enantiornithes, early birds who had claws on their wings, and teeth.
Back in the previous chapter we looked at the evolutionary history of action, especially in animals. I divided animal action into five rough categories: movement, feeding, interaction between individuals (other than feeding), engineering, and information gathering. A conspicuous behavior within the third of these is communication. Communication has borders that are hard to mark out, and it fades and merges into other forms of interaction. Communication is common throughout life; its role will become enormous, almost overwhelming, when we begin, in the next chapter, to consider humans.

We might initially say that an action is a case of communication when its goal is to get a message across, or convey information. But which interactions involve messages and which don’t? We might try this: In communication, something is done by one organism to be perceived by another, and done with the aim (conscious or not) of affecting the other’s actions or responses in some way. Yes, but some cases that would be included in that category seem distinct from communication—handing over food to share, for example, or physically blocking someone’s path. Often, though, cases like that are mixtures, with a communicative aspect to them and other aspects as well. If so, what is the communicative aspect?
This question can be approached by looking more closely at the behaviors “on each side” of an action, display, or mark—the production side and the interpretation side. I’ll start with one setup, which can be seen as central in some ways. In these cases, a conspicuous feature is that the behavior seen on the receiver or interpreter side is not much concerned with the intrinsic physical nature of whatever was perceived, but is directed onto something else, perhaps some distance away. Vervet monkeys, for example, give alarm calls to one another, and they have different alarm calls for different threats: eagles, leopards, snakes. If a receiver hears the eagle call, they look up; if they hear a snake call, they look down, and so on. This receiver-side behavior is directed on something else, the threat. The receiver’s behavior only makes sense, though, if the behaviors on the sender side are set up the right way. It only makes sense to look up when you hear a particular sound if the senders make that sound when they’ve seen an eagle, not a snake. In a stable communication system, the habits on the sender side and the habits on the receiver side are aligned; they fit together. In this case, the different calls do not seem to be modeled on anything about snakes, leopards, or eagles themselves. You could do things that way, but it’s not necessary, and the vervets apparently don’t. If senders and receivers are “on the same page,” then the particular sound or sign used to indicate something does not matter, and to be “on the same page” is to have that interlocking relationship between production and interpretation.
In that first case, the behavior on the receiver side that anchors the setup is a behavior directed on something else in the environment. In another group of cases, the interlocking behaviors on each side are different. Rather than coordinating behavior in relation to something else, they coordinate or initiate behavior between the animals themselves, as in a “Shall we dance?” signal, or something like that. Again, a “Shall we dance?” signal might be derived from something seen in actual dancing, but need not.
The idea that nearly anything could be used as a sign for nearly anything else in a communication system is described in linguistics and semiotics as the “arbitrariness” of signs. This is not always present; sometimes it does matter what a message is physically made of. The communicative side of a food-sharing action requires that what is handed over is food rather than something else, and a display made in a mating context might be chosen as a natural indicator of vigor or health. The medium can matter. But the core phenomenon is, I hope, starting to become clear. In communication, there is an interaction mediated by something produced by one party and seen (heard, smelled …) by another. What makes the interaction a communicative one is the fact that the behaviors on each side of the sign, sound, or mark are aligned, perhaps coordinating actions that are directed on some other object or situation, perhaps coordinating other actions of the sender or receiver themselves. (It might be both.)
There are mixed cases, where an interaction has some of this communicative flavor plus something else as well. Communication also extends outside the realm of action or behavior itself, as it includes colors, marks, and other permanent features of an animal’s body. Bright feathers and long tails are designed to be seen, to get a message across and induce a response. The clear, recognizable phenomena of communication emerge from vaguer, partial forms—just as we saw, in previous chapters, in the cases of action, organisms, goals, and engineering.
Birds are enthusiastic participants in send-receive systems of this kind, and their calls have a variety of roles—threats, alarms, advertisements. Many of the bird sounds in the fish-bowl aviary behind my house are probably “contact calls” between pairs or members of a group; they are staying in touch, registering one another’s presence.
Communication, like other forms of action, is older than animals. Bacteria communicate by releasing and absorbing chemicals. Some of what they do may be faintly analogous to those contact calls in birds; bacteria use chemical communication to work out how many other cells of the same kind are around (this is called “quorum sensing”). The sea, home of early animal life, is not really as quiet as it can seem; shrimp click and fish croak, doing this to impress size and vigor on rivals or potential mates. But unless you are lucky enough to find yourself immersed in whale song, the sea’s soundscape is nowhere near as dramatic as the choruses arising from birds, frogs, or insects such as cicadas. In fish, courtship often works through dances and visual displays.
Octopuses, who appeared in chapter 3 as maestros of manipulation, are not very social. But given their behavioral abilities, there is a lot they can do in this area if they want to. In the unusual circumstances of Octopolis and Octlantis, the high-density sites I mentioned a few times in that chapter, we have seen signals and displays. One we have called the Nosferatu—tall, dark, and cloaked, and, unlike its cinematic vampire namesake, featuring the rear end of the animal raised over its head. This seems to be an indication of aggression: “I’ve seen you and I’m coming for you.” We’ve become interested also in a less common behavior, informally christened the “arms to heaven” display. Two of the arms, not the front ones but the second pair, are raised over the head in a great arc and held there for a second or so. The body seems to be stabilized by having the front-most pair of arms held down so the next pair can reach high. The drawing on this page makes this clearer. This might look like an odd pose for an octopus, but the drawing is traced from a video frame—this is really how the behavior looks.

When we were working on a paper about the Nosferatu behavior, a critic said: “That is not a signal; that is just a preparation to jet toward another octopus, a gearing-up—and the other octopus can predict what is likely to follow, so it reacts.” It is true that displays can arise from the “ritualization” of other behaviors, like preparation for an attack, and there won’t be a sharp divide between displays and more “ordinary” behaviors. In the Nosferatu case, we do think it has become a display, at least in part. In any case, the arms-to-heaven gesture does not seem to have an alternative interpretation of the kind that was offered for Nosferatu. There’s no other action that this sudden raising of the two second arms resembles, as far as we know. The closest might be a reaching-out, a probing—octopuses do a lot of that. But as the drawing shows, the arms-to-heaven is not much like a probe. The action is disarmingly (!) human, with arms skyward and curled slightly in. What does it mean? We are not sure, but it seems sometimes to be an “I’ve seen you!” or “Here I am!” signal, often done as another octopus makes its way closer, but without the aggression of Nosferatu. (I like David Scheel’s suggested translation: “See me here!”)
We can scour the seas for such cases, but—again with the mammalian exception of whale song—none of it is in the league of bird calls. I was sitting with some friends on a deck over the garden of Dave Pye, carpenter, musician, and cultivator of orchids. Suddenly there was a great racket. At least six or seven cockatoos began shrieking, and some other birds (bowerbirds) joined in with sounds of their own. Many or all of the cockatoos were close by each other, high in one tree. Dave said: There’s probably a snake down there, or a goanna (a large Australian lizard); they are sounding an alarm. So we walked down under the birds, and, sure enough, a diamond python was curled up at the base of a bush—looking, I imagined, a little sheepish as the racket went on above him.
Display and Evaluation
Some actions are communicative; they exist within sender-receiver setups, and their goal is to affect the behaviors of those who perceive the communicative act or the mark it leaves. Within this category is another. Some actions and displays are made to be evaluated by the other side. Am I attracted? Am I impressed? Often I might not actually be impressed, but that can still be the goal that the behavior had. That is why the other animal spoke, sang, wrote, painted, or danced in the way they did.
I mean this category of “evaluation” broadly, to cover all sorts of responses that include some assessment of good or bad, attractive or repulsive. Evaluation or appreciation is a special kind of receiver-side behavior. I say it is “receiver-side” although some of what we evaluate was not produced for that purpose. A lot of what we evaluate was produced for a reason of that kind, though, and a huge amount of effort by animals is aimed at getting particular evaluative responses to occur.
Song is a case in point. In human life, the Orpheus myth, coming down to us in different versions from the ancient world, has encapsulated the power of music. Orpheus, who could charm not only people but animals and stones themselves with his lyre, failed to rescue his lover Eurydice from the Underworld after she was bitten by a snake (perhaps not heeding the cockatoos’ warning). After his rescue attempt, Orpheus wandered, spurning others, until his mourning prompted his murder by the maenads, female followers of Dionysus. The earliest two operas that have survived were both written about Orpheus. In different versions of the myth, the preoccupations of local social contexts, especially sexual concerns, are layered onto the old story. But through all this, a central thread is the uncanny and sometimes uncontrollable power of music.
The power of this combination of behaviors—display, evaluation, attraction—is not restricted to human life. It is a life-shaping, and hence Earth-shaping, combination. Evaluation is subjective; it is tied to the goals and values of a receiver, and how something appears to them—how it strikes them, whether it is found attractive. Though it is subjective in this way, evaluation has wide-ranging effects on what is around us. The ornithologist Richard Prum uses the phrase “aesthetic evolution” for evolution that is driven by displays and evaluative responses to those displays.
Birds, the focus of Prum’s book The Evolution of Beauty, show us what aesthetic evolution can do, with their plumage, displays, songs, and body forms. Flowers are another example. Flowers are made to be seen or smelled, made to be drunk from, made to attract insects and other animals. They are not just made to be perceived, but made to appeal. Attracting insects, birds, and bats allows flowering plants to interact, plant to plant, over long distances. The previous chapter noted how these plant-plant interactions are routed through the behaviors and nervous systems of animals; they are also routed through those animals’ values and goals. Forests of flowering plants depend on the interaction of displays and evaluation. Thor Hanson, in his book Buzz, contrasts forest scenes of a typical kind with the vegetation on the Juan Fernández Islands off the coast of Chile. These islands are, and have long been, almost completely without bees. As a result, the flowers are mostly small, greenish-white, and lacking the symmetrical displays of petals that bee-visited flowers tend to have. I said back in chapter 3 that to imagine an early flower, one might picture a magnolia. We should picture it also as white. Magnolias evolved before bees did, and they evolved to be fertilized by beetles. Multicolor clusters of flowers, especially blue and yellow, came with bees. (According to Hanson, most bees can’t see the color red; that is a bird-attracting color.)
At this stage, it is worth taking a moment to ask a basic question: What are colors? Whether colors are real features of the “external world” or exist merely in our minds has been debated by philosophers for centuries. I think that the habits we have of talking about colors as features that have to be located somewhere, inside or out, are a bit at odds with what is actually going on.
The full story about color, in outline, looks like this. Visible light is a form of electromagnetic radiation. That radiation exists on a wide spectrum, from radio waves to gamma waves, with color in the middle. The differences along this spectrum involve the wavelength, the distance between peaks in the wave. This to me is such a surprise—that radio waves, visible light with its colors, X-rays, and gamma rays are all essentially the “same stuff,” differing just in wavelength (and with definite scare quotes around “stuff”). In the range that is visible to us, finer wavelength differences are associated with colors. The surfaces of objects around us, as a result of their makeup, reflect some wavelengths of light and absorb others, and our eyes then absorb what reaches us. (Jackie Higgins comments in her book Sentient that a “red” flower is, in a sense, not red but anti-red, rejecting red and absorbing the rest.) The phenomenon of color, then, involves objects reflecting and radiating light, the radiation itself, and the responses in the eyes and brains of animals, which can differ between us, other mammals, insects, and so on. That whole is what the coloring of the world involves, where it lives.
In any behavior, preferences of some kind must be in the picture. Having detected something nearby, do you approach or avoid it? Here is sugar: Is that a good thing or a bad thing? Any behavioral choice reflects an agenda and goals, but these may be implicit, or “wired in,” and not felt by the organism. Even bacteria prefer some chemical environments to others. Especially in cases with insects and some other animals, we might wonder whether attraction and preference are experienced at all—whether the insect finds the flower pleasing or exciting, as opposed to just fitting a specification or a search image. The question perhaps has two sides. First, we can ask whether an assessment is experienced in any way at all (remember the bacteria), and a further question is whether the response is felt in something like an emotional way, or more dispassionately (“Hmm, very suitable”). Prum, in The Evolution of Beauty, tends to press ahead, assuming that evaluations are indeed felt. His book is mostly about birds and mammals, where the question of feeling is not too vexed. Once we put insects, and their all-important relation to flowering plants, into the picture, the question does press.
Even if an evaluation is not emotionally felt, it is still tied to the values and point of view of some agent, and subjectivity in this sense can be a powerful force. Back near the start of this chapter, I sketched the evolutionary history of birds and said that their most distinctive feature—feathers—arose even before the dinosaurs in question looked very bird-like. Why did feathers appear so early? Their most conspicuous function now is flight, but feathers, it is thought, appeared initially in animals that had no chance of flying. They must have had some other role. Insulation, the control of temperature, used to be the standard hypothesis on this matter. When I was a student, this was often used as an example of change in function in evolution; feathers first had the function of insulation, and then flight. They initially had one sensible, practical role, and then another. But now it is thought likely that the initial role of feathers, at least in part, was something more extravagant: display. Prum supports this view, and so do others.
Nature has a mix of happenstance beauty and designed, crafted beauty. Some things are made to be appreciated, even if they were not made for us. Others ended up in a form that we humans, or some of us, or us plus perhaps some nonhumans, find beautiful even though they were not made to be seen at all. I once posted a photo of an intense sunset, up in the far north of Australia, on social media and a person commented, “Beauty for its own sake.” It would be more accurate to say: beauty for no sake.
On the “beauty for no sake” side of the divide we find not only sunsets, but that marine analogue of a flower-filled forest, a coral reef. Healthy reefs have an abundance of colors—dark greens, cornflower and neon blues, turquoise, orange, and pink. Why are reefs colored? I had always thought the colors, and their diversity, came from the light-collecting symbionts living inside the coral polyps. Light-gatherers of this kind do not have to be green. But in this case, all the symbionts in corals are fairly closely related and their own color is a green or brown-green, not much different from plants in a forest. The colors in a coral reef—the blues and pinks, the turquoises—instead tend to be largely, at least, a result of chemicals made by the coral animals themselves. These chemicals seem to be provided by the polyps as a kind of sunscreen for their symbionts, to prevent them from being overloaded, and also to organize their light-collecting activities.
When I was young, we were given brightly colored zinc sunscreen to stripe on our faces. The colors, other than white, were not functional; they were added to encourage us to wear it. Sunscreen now tends to be transparent; the fashion has not persisted. So with a layer of protection over photosynthetic cells, a coral reef is a bit like a forest covered in brightly colored sunscreen.
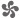
In relation to the mix of designed and undesigned beauty (beauty for no sake) in nature, seahorses are an interesting case. They are beautiful animals, at least to many people, with their implausible, delicate bodies. They form long-lasting bonds, and pairs in some species engage in elaborate dance-like greetings each morning. These dances are meant to be seen and recognized, and perhaps appreciated. Their role, expressed in human terms, might be some approximation to, “Yes, we’re still here, still together, still glad to be.”[1] Seahorses can change color on both rapid and slower timescales, and some of their color choices may figure in those morning dances (they do have color vision).
Those implausible bodies look like they were designed on a whim by a jeweler—until you see them in their milieu, in their element. I was once diving at Nelson Bay, the site described in the forest-and-reef section of the previous chapter, and spent a little too long in the calm water with an especially beautiful radiant yellow seahorse, when the tide turned and the water began rushing out. Both the seahorse and I started to twist, and in a moment it was clear who was at home there and who was not. I soon had trouble keeping any kind of position and control at all. As I was battling, a swarm of stingrays came past. They were high in the water, managing to swim against the tide, but doing so by swimming very hard. These were cow-nosed rays, a name that does no justice at all to their diamond-like form. They looked like a flock of gigantic starlings. (I noted in the previous chapter that a flock of starlings is called a “murmuration”; a collection of stingrays is called a “fever,” and that is very much how they looked.) Then they were gone, up toward the river whose tidal release was sending all this water down on us.
Soon it was not possible for me to retain any sort of position. I let myself go, and went off like a balloon with the rushing tide. As I went, I passed a tiny and even more delicate yellow seahorse on a sponge—this one can’t have been more than an inch and a half long. With her slender tail, curled like a lock of hair, she was hanging on calmly, effortlessly, and doing just fine as I careened past toward the exit.
Song
Many birds, such as the parrots of the opening of this chapter, can make calls but not song—not something sustained and organized. Most of the elaborate avian singers belong to a particular group called songbirds (or oscines) within the passerine birds, the large group I mentioned earlier that makes up a majority of all bird species. Not all of the oscines actually sing, but they share a distinctive vocal organ that enables some to reach remarkable sonic heights.
At the moment, it appears that the whole enormous branch of passerines, and the songbirds themselves, originated in Australia (though it was a larger version of Australia, with other, now separate landmasses attached). Debates about bird classification and origins can be quite fraught. As Tim Low notes in his book Where Song Began, the idea that Australia might be the birthplace of so much of the splendor of bird life faced considerable resistance (including from Ernst Mayr, a bird-resembling evolutionist who perched astride much of twentieth-century ornithology), as if Australia was too scruffy and disreputable a place to be the first home of the group that produced nightingales and larks. In belated recognition, a huge branch of the bird tree, one containing a majority of all bird species, including songbirds, has recently been designated the Australaves.
Australia today includes living representatives of the earliest known branchings in the genealogical tree of songbirds. Some products of those branchings are my next topic.
The first split within the songbird tree gave rise, on one side, to just a handful of birds alive now, the lyrebirds and scrubbirds, and to a huge collection of birds on the other side. That spindly lyrebird/scrubbird branch has just four species living today. The tree diagram on the next page shows some of these relationships. We are zooming in on the right-hand side of the first tree diagram for birds here.
It’s often said that lyrebirds are “ancient” birds—the most ancient songbirds. This way of talking is confused in some respects; a lyrebird today is the product of as many years of evolution as any other living bird—or you, for that matter. Lyrebirds are distant cousins of other songbirds, but your more distant human cousins are not more “ancient” than your nearer ones. Lyrebirds are also products of a branching that led to very few survivors today. That also does not make them ancient; it makes them distant cousins of other birds, off by themselves without a lot of close living relatives of their own.

Still, lyrebirds do look old. They have a dinosaur-like appearance, at least to me. Their fossil record also goes back a fair way, and they have at least one feature that is probably genuinely old: Lyrebirds and scrubbirds have a different kind of syrinx, or voice box, than other songbirds.
Lyrebirds are quite large, around the size of a pheasant, with dark feathers and, especially on males, long tails. They have a prominent forehead that can look a bit Mesozoic. Lyrebirds spend most of their time on the ground but can fly when they want to—although it often looks like an approximation of flight, as if an early attempt. They are agile, moving quickly with flappy strides and leaps through thick forest. Their long legs have large claws, used to rip up the ground as they rummage for food.
Lyrebirds are fairly common in the forest that introduced chapter 3, the forest reached by descending stairs and ladders into a valley in Australia’s Blue Mountains (the species is the Superb Lyrebird, the only one I’ve seen). I was sitting quietly down there once, in a clearing around ten in the morning, when I heard a couple of sudden, unexpected sounds. Turning, I found a lyrebird. I didn’t see her arrival, but there she was, quite close. The lyrebird had descended noisily from a tree—they sleep there at night—and this one was keeping impressively relaxed hours.
Standing up, she made exactly the sound you would make if you were a smallish dinosaur who had woken up, jumped out of a high tree, landed awkwardly, and were trying to pull yourself together: a deepish grunt and suppressed squawk.
A few minutes later, another appeared suddenly, though I again did not see the fall. Thump. And a third.
I have followed lyrebirds for some time through the forest, once they are on the ground. They rove around tearing things up. Sometimes much smaller birds follow closely, evidently looking for stray bits of food kicked up by those big claws. I saw one of these little wrens get so close underfoot that it was thrown out the back in a great tumble, head over heels. It reminded me of octopuses hunting and small fish following them to see what gets unearthed.
I saw one lyrebird in the early evening, after a several-course and very messy dinner, go and clean herself off in a stream. First she cleaned her legs, then nearly her whole body, splashing around. She then went and sat on a branch above the ground. There, she appeared to dry off and preen. A bath after dinner.
Lyrebirds are also, many agree, the most spectacular songbirds of all. Their mimicry is astounding. Males produce a cascade of near-perfect copies of the calls of other species, in quick succession. These are concatenated, sometimes jumbled, and mixed in also with some mechanical-sounding calls that are not mimicking anyone. Females mimic also, and seem to mimic different species than males.
One study found that even other birds can’t always tell lyrebird counterfeits from genuine calls of their own species. Sometimes you can tell; sometimes the mimicry seems too “formal”—like an operatic soprano too carefully singing a folk song. Their sound also projects more, sometimes, than the original might. But that is a rather fussy review! They are just so impressive. At least in males, it appears that much or most of their repertoire is picked up from other lyrebirds, although they can add new sounds on their own. Males are calling to impress and attract mates. The function of female songs is less clear; some singing appears to involve competition with other females for territory.
In the next chapter, we’ll encounter some ideas about human behavior and evolution that were developed by Kim Sterelny, a philosopher. He and his partner, Melanie, a historian, have a house in an Australian forest with a large population of lyrebirds. This can engender a fair amount of sonic confusion. A flock of cockatoos found their calls answered unexpectedly, and seemed to lapse into an irritated silence. Melanie is a committed gardener, continually planting and restoring, and often hammering large metal stakes into the ground to make protective cages around new plants. One day, Kim had been imploring her to take a break. She agreed that she might have been overdoing it and went inside. A little later, Kim heard a telltale ringing sound, metal on metal, of more hammering of stakes. He went outside to berate, or at least plead. It was a lyrebird. This sound, Kim said, was distinct from the abrupt mechanical sounds that lyrebirds also make, though perhaps it was a modification of such a sound.
Bowers
In songbird evolution, the lyrebirds branch off early. Then there’s another early split, leading on one side to another small cluster of species alive today, and on the other side to thousands. The smaller cluster includes another group of birds that are notable for their actions, especially in the light of the classification of actions I gave in the previous chapter. These are the bowerbirds.
Lyrebirds have that slightly dinosaurian look, but bowerbirds just look like birds. They do move with an awkward hop, as if (on analogy with lyrebird flight) walking is the project that evolution is still working on here. Their great forte is collecting and engineering. Male bowerbirds build a nest-like structure on the ground, but it is just for display. Constructed from plant stems, it has, in many species, two gracefully arched walls, meeting at the top or not quite, forming a tunnel. Other species build what look like huts or maypoles. A collection of objects will be placed out front.
Different species of bowerbirds often prefer different colors. Where I live, the Satin Bowerbird is the local species, and it collects bright blue objects. To find a bower in the undergrowth, look for those objects. (It’s like finding an octopus—look for what the animal has done.) The blue objects, which often now tend to be human-made, such as plastic pegs and bottle lids, will be mixed in with some bright yellow ones. That combination seems to have a special role, and a male will sometimes hold, in its beak, a blue object plus the petal of a small yellow flower. These, or just a yellow petal, are presented to any interested female.
Males don’t just add continually to their bower. There’s a lot of rearranging, and what looks like some careful subtraction. The collections of colored objects are also tended and modified. The best collection I’ve seen had a regular array of slender yellow grass strands and feathers underneath, dozens of blue objects on top, plus—in a Gothic touch—a single, small white skull.
Why do they choose blue? Color choices vary across species, and do so in intriguing ways. The blue-collecting Satin Bowerbird has intense blue eyes, and blue-black feathers on mature males. The Great Bowerbird, up in the north of Australia, ignores blue objects and collects white and other pale-colored ones, with a few other colors, including red and green, sprinkled in. That species has pale gray feathers, and in the only individual I’ve seen, its silver-gray-white feathers were quite close in color to its ornaments. Males of that species also have a lilac-pink crest. The Spotted Bowerbird, which I’ve never seen, is said to have no particular color preference in its collections, and its body, also, is mixed colors—browns, grays, green, and again a lilac crest. As you can see, I am curious about resemblances between the ornaments and the bodies; although the principle seems far from firm, matches in some cases seem unlikely to be coincidental.
In the case of preferences for blue, rarity has been conjectured as an important factor. Before all that plastic was available, blue objects would have been rare in many bowerbird environments. Far from civilization, the blue objects seen in bowers tend to be parrot feathers and flowers. An interest in rare colors makes sense—until humans degrade the currency. Studies of some other species have suggested that rare objects are not preferred in those cases. However, there’s no need to assume that the various bowerbird species are all doing the same thing; each species has a system of its own, and they could have evolved in different directions. In the case of Satin Bowerbirds, I keep coming back to that match with the birds’ brilliant blue eyes.
What about those combinations of blue and yellow that figure in the Satin Bowerbird displays? As the botanist Katherine Preston noted to me, yellow and blue are “complementary” colors, at least in some color schemes, and their contrast might be especially strong. Yellow also matches the Satin Bowerbird’s beak color in adult males, and several things might be going on here at once. Jared Diamond, in the course of some intrepid work on a spectacular bower-building species in the mountains of New Guinea, saw hints of local “cultural traditions”—different preferences and styles in different nearby locations, probably due to the copying of older birds by younger ones. This is now looking possible in several species.
Whatever account is given of their color choices, the bottle caps, petals, and other collected objects here are tools, in the sense introduced in the previous chapter of this book. They are tools not for building, but for social interaction. This is not usually how they’re described, but I think that’s because of our habitual association of tools with more utilitarian projects. The bowerbird’s objects fit the requirement that the animal is manipulating one object with the goal of having an effect on something else, in this case another bowerbird.
The effect these tools are aimed at—the bowerbird’s collection of objects, and the bower itself—is evaluation and appreciation. They are intended to impress. A bird guide I traveled with, Laurie Ross, watched a bowerbird laboriously construct what looked like a fine bower. The bird then saw his product fail to get any response from several females in a short period, and he tore it down to build another.

Bowerbirds are not just collectors and builders, but in all ways tremendous displayers and advertisers. They hold out their objects, dance, and sing in a low, sliding coo. Like lyrebirds, they do some vocal mimicry. Their courting behaviors seem to employ every possible means to make an impression: body, sounds, found objects, built artifacts—everything.
Bowerbirds are an extreme illustration of the way birds have been shaped by the linked activities of display and evaluation, and they have taken these activities in an unusual direction. It’s surprising to me that just this one small group of birds, out of all the species, engages in this kind of collecting and building. A few other birds use tools in their displays. The Palm Cockatoo of far north Australia drums rhythmically with a branch—a drumstick that has been cut and shaped by the bird—on dry wood to attract a mate. But there seems to be nothing else like these elaborate structures built by bowerbirds. Gerald Borgia, after studying the birds extensively, has suggested that bowers themselves originated as places where females could view male displays and ornament collections while avoiding the possibility of being subjected to an unwanted copulation. To initiate a mating, the male has to leave his main display area, at the front of the bower, and go around behind it; this gives the female a chance to make an exit. From there, bowers were elaborated. In addition to evaluating displays and the rest, females began to assess a male’s quality by the bower itself. In this view, the close inspection of ornaments and displays came first, the first bowers arose as a protected area like an opera box, and only later became something the females assessed aesthetically as well. All this still leaves open the question of why collecting ornaments is itself so rare.
When they are elaborate, bowers are reminiscent of peacock tails and other expensive-looking aids to display in some male birds. Biology has seen a long-running and sometimes tense debate about features of this kind. One view has it that these features are used by males to advertise health and vigor, in a way that is hard to fake. Only healthy males can afford to produce such extravagant features. An alternative is that these features are the result of a “runaway” process where a bodily feature in males, and an initially somewhat arbitrary preference for that feature in females, become genetically associated with each other. As this association becomes established, each feature becomes more and more extreme. These much-debated differences don’t affect my larger themes here: the sheer importance of display, evaluation, and attraction as elements of the living world, especially of animal life. Thinking specifically about birds, it’s also appealing to think that all this elaborate communication and display was in place, in such extravagant styles and in a separate evolutionary line, well before humans started doing any of these things.
Orpheus in the Undergrowth
Orpheus, the mythological figure who could charm all who heard him, including stones, and later prompted the first operas, played the lyre. That the world’s most accomplished songbird carries a lyre on its body is a glorious, lyrical fact, a true piece of evolutionary alchemy.
That name—lyrebird—comes from the feathers on the tails of males. A lyre, the musical instrument, usually has curved, bow-like wooden arms that connect the main body of the instrument to a crossbar, and strings run between the crossbar and body. A male lyrebird’s tail has two long curved and banded feathers (the “lyrates”) that sit on the outside and are extended outward when displaying, with many finer and pale feathers between them.
On one of the first occasions when I spent any time with a lyrebird, I was walking a rainforest path and heard what sounded like a whipbird. These are small and elusive birds, with a call-and-response between pairs that includes a sharp, impressive whip sound—a sizzling, ascending snap. The call I heard was very much like a whipbird’s, but a bit too clear, too well projected (as in my earlier comment about some lyrebird mimicry being overly operatic). I left the track and started walking up into the bush.
The path I’d been on was a standard and well-traveled one. A minute or so off it, I was in a completely different place. The walk became dense and steep. About half the things I touched seem to prick or slightly sting me. I found Orpheus standing on a low stump. In quick succession, he was a whipbird, a black cockatoo, a Laughing Kookaburra, a magpie, and many I didn’t know. I moved a little toward but also around him, trying to give him space. Initially he edged away, but after a while he seemed not to mind and I was able to get quite close. I stayed there for about half an hour.
In a campground parking lot in the same area (Dorrigo Rainforest), I locked the car, which beeped, and heard an immediate echo. I unlocked and did it again—the same echo, not matching the car exactly, but close. I never saw the imitator, but suspect it was a lyrebird. The scientific literature is somewhat skeptical about claims of mimicry of human-made sounds, at least in the wild. This may be partly because of some exaggerated early tales. If lyrebirds living in captivity count, then the case is clear. A lyrebird in Sydney’s zoo has been recorded producing various human-origin sounds. Its rendition of a baby crying is amazing; at least to me, it’s indistinguishable from a human cry.
I also said earlier that male lyrebirds acquire their repertoires not, primarily, by directly copying the calls of other species and combining their sounds, but by copying the organized songs of other lyrebirds. The initial copying of a whipbird or parrot has to happen at some stage among the birds in some region, but the sound then stays in place through copying. Other birds are good mimics, especially some parrots and starlings, but do any others pass sounds from bird to bird in this way? If this capacity of lyrebirds is old, then this is, I suppose, the oldest recording medium on Earth.
This system can preserve a sound for a considerable time, too. A whipbird had attracted me to Orpheus, the lyrebird I’d watched in the undergrowth. Whipbirds are not found in Tasmania, the island at the very bottom of Australia, but lyrebirds were introduced there in the 1930s, and they brought with them the sizzling snap of the whipbirds. In 1964, the lyrebirds down there were clearly reproducing that call. A (human) recording made in 1984 still had a “barely recognizable” remnant of the sound. For thirty years the lyrebirds’ mental record remained intact, and faded a few decades on. Lyrebirds can live for twenty years, so this preservation need only have covered a couple of generations, though it might have been more. If another, younger lyrebird was listening to Orpheus along with me, somewhere nearby, then his song, though not preserved in its particular details, might live on as a contribution to others, carried forward through time.