‘There is growing evidence that a baby’s development before birth has a major impact on their health in later life. This means that the prevention of obesity needs to begin in the womb.’1
Neena Modi
HUMAN BABIES ARE the fattest animals on earth.
At birth between 15% and 16% of a baby’s body consists of fat; they continue piling on the pounds for the next twelve months, consuming some 220,000 calories in the process.2 By the time they blow out the candle on their first birthday cake, up to 30% of their body weight is made up of fat.3
Nor is this our species’ only peculiarity where fat is concerned. While other mammals, including our primate relatives, only produce white fat once they have started to suckle, human babies start doing so while still in the womb.4 This explains why, to the delight of their proud parents, they look so plump and feel so smooth to the touch. By contrast, other newly born primates have only 3% fat, and so look and feel, in the words of German anthropologist Adolph Schultz, ‘decidedly “skinny” and horribly wrinkled.’5
But why are human babies so much fatter than other primates? What function can this vast calorific investment in producing adipose tissue possibly serve? And what can fat infants tell us about the causes of adult obesity?
Mammals require large deposits of fat for two main reasons – to hibernate or as protection from the cold. Hibernating animals build up their fatty reserves when food is plentiful and live off it when food is scarce.
In animals exposed to constant cold, such as those living in the oceans or in frozen wastes, fat is present all year round. And although only a few humans are ever exposed to such extreme environments, we too maintain large stores of fat from one season to the next. While most animals deposit fat deep within the body – especially between the muscles and around the heart, kidneys and intestines – both humans and sea-dwelling mammals store a high proportion directly beneath the skin. If you were to remove the skin of a rabbit or an ape, much of the superficial fat would remain behind on the organs. Do the same with a human, a seal or a dolphin and a large proportion would come away with the skin. The differences between visceral (deposited around the organs) and subcutaneous (deposited beneath the skin) adipose tissue are extremely important and we will return to them later in this chapter.
A number of theories have been advanced as to why we humans are so very different from every other land animal in the way we synthesise and deposit fat. We must assume that there is a good reason for this; if storing fat in the way we do was simply detrimental to our health, evolution would surely have corrected this by now. In this chapter we will explore several possible theories for the distribution of our fat tissue, as well as considering our brain-to-body ratio (or ‘encephalisation quotient’); another key feature of human biology which makes us so unique among mammals. As we will see, the relationship between the brain and our evolved patterns of fat storage has critical relevance to the problem of obesity.
The naked ape theory proposes that fat provides insulation for human infants which, unlike every other primate, are born both hairless and chubby.
‘Fat [is plentiful] in the new-born baby,’ wrote David Sinclair, Professor of Human Anatomy, University of Aberdeen, ‘probably as an insulation against cooling.’6 At first sight this seems to make a lot of sense. Human infants have a large surface area per kilogram of body weight.7 A baby with an average birth weight of between 7 and 8 pounds, for example, will have a Body Surface Area (BSA) of around 5 square feet, about the same size as a picnic rug, which means they lose heat rapidly.
But while the ‘insulation’ theory sounds plausible, there is a problem with it. Although human babies have nearly seven times as much fat beneath their skin as elephant seals, and three times more than fur seals, they don’t really need it to stay warm.
Seal pups, after all, spend their infancy swimming in freezing oceans or sleeping on sheets of ice in sub-zero conditions, while most human infants are snugly wrapped against even the slightest chill. And the Inuit, who inhabit some of the earth’s most intensely cold regions, do not possess extra layers of fat beneath their skin to provide greater insulation. Even as babies they, and other people indigenous to the polar regions, have subcutaneous fat deposits either equal to, or even smaller than, those living closer to the tropics.8
This has led many to conclude it is the human’s ability to generate heat via their lean body mass, rather than to conserve it by means of adipose tissue, that enables infants, as well as adults, to maintain a steady core temperature of 37°C.
‘Humans are among the fattest of mammals,’ observes anthropologist Professor William Stini of the University of Arizona, ‘but do not adapt to cold climates to any significant degree through the acquisition of thick layers of subcutaneous fat.’9
So if layers of fat beneath the skin are not, as the naked ape theory proposes, to protect us from the cold, what are they there for?
In the early 1940s, Max Westenhöfer, a medical doctor from the University of Berlin, described what he termed ‘aquatile Mensch’.10 Influenced by the mood of National Socialism of the time, Westenhöfer disagreed with aspects of the Darwinian theory of kinship between ape and man. Instead, he looked to the sea to describe traits unique to man, disputing the idea that we evolved strictly from plains-based primates. However, his theory was riddled with inconsistencies, and did not reach a large audience.
Some twenty years later a marine biologist, named Alister Hardy proposed a similar theory. ‘The suggestion I am about to make may at first seem far-fetched,’ he told an audience of SCUBA diving enthusiasts in Brighton, ‘yet I think it may best explain the striking physical differences that separate Man’s immediate ancestors (the Hominidae) from the more ape-like forms (Pongidae) which have each diverged from a common stock of more primitive apelike creatures which had clearly developed for a time as tree-living forms.’11
Hardy went on to expound his own version of what became known as the Aquatic Ape Theory. He argued that around 5 million years ago, due to competition from other animals our ancestors were forced to leave the plains to search for food in the oceans. He claimed that the length of our legs and our lack of hair are signs of us having adapted to an aquatic environment. He also pointed to similarities between the adipose tissue deposits in humans and ocean-dwelling mammals, such as dolphins.
While superficially persuasive, the Aquatic Ape Theory is rejected by the majority of scientists today. A more popular explanation is that rather than helping to streamline the body for swimming, our lack of hair can be explained as the result of our need to control body temperature within fairly tight limits.12
Sweating is a relatively wasteful mechanism for temperature control, in that it uses up valuable water. In a hot environment such as a savannah, a furry animal would require even more water for cooling, since the fur would naturally cause it to retain heat and therefore perspire more. It seems likely that a perspiring animal that evolved in a hot climate would therefore gradually lose its fur. However, the loss of fur would likely lead to some kind of compensatory tissue, as the animal would still have to contend with cold temperatures upon occasion. This is where other scientists postulate our hairless appearance and our concentrations of subcutaneous fat unique in land mammals, have their origins – they allow us to stay reasonably cool when it is warm, and reasonably warm when it is cool.
Subcutaneous fat as a method for insulating an organism (rather than external fur) makes a lot of sense from an evolutionary perspective and we certainly should not discount the theory. However, there is also a strong possibility that it is our brains rather than our bodies that require the extra glucose which stored fat represents. When we consider our physiology in more detail, particularly in comparison to other mammals, there is good reason to think this may be the case.
The average human brain is 1,000 grams larger than would be expected for a mammal of our size and weight.13 At 20 watts, the metabolic cost of our brain is also more than four times greater than that of the typical mammal, which is around 3 watts.
‘A human child under the age of 5 years uses 40% to 85% of resting metabolism to maintain his/her brain,’ according to William Leonard and Marcia Robertson of the University of Guelph. ‘Therefore, the consequences of even a small caloric debt in a child are enormous given the ratio of energy distribution between brain and body. Hence, the prolonged period of growth in humans may be partly an adaptation to limit the already high total and brain energy requirements during childhood.’14
Although these demands decline as the child develops (the adult brain consumes about 20% of total energy production), this still represents a considerable investment. The brain makes up just 2% of our body weight yet it consumes more than twenty-two times the amount of energy required by our muscles.15
Which raises the question of how humans are able to afford the metabolic cost of possessing such a large brain.16
Where does the energy required to fuel the brain come from?17
In 1995, Leslie Aiello, a Reader in Psychological Anthropology at University College London, and Peter Wheeler, Director of Biological and Earth Sciences at John Moores University, Liverpool, suggested humans were able to develop larger and more efficient brains because they also evolved smaller and more effective guts.18
Known as the Expensive Tissues Theory, this groundbreaking idea arose from a realisation that the larger the brain, the greater its appetite for energy in the form of glucose. Assuming you are fairly inactive, the energy provided by one fifth of all the food you consume goes entirely to fuelling your brain. Aiello and Wheeler asked themselves what special features humans possess that enable them to deliver more glucose to the brain than other animals. One possibility might have been that we burn up energy at an exceptionally fast rate. It turns out, however, that a person at rest burns energy at almost exactly the rate predicted for any primate of our body weight. This ruled out the possibility that our big brains are powered by inordinate amounts of energy passing through the body.19
This left only one possible explanation. The metabolic cost of brain tissue is approximately 240 kcal/kg per day (at rest), which is similar to other organs such as the liver (200 kcal/kg per day), and around half that of the heart (440 kcal/kg per day). Yet, as we have seen, the brain accounts for approximately 20% of total energy usage in adult humans, compared to 8–10% in primates and 3–5% in all other mammals. The immense energetic demands of the brain, therefore, can only be met by a corresponding reduction in the amount being consumed by other organs in the body.
Among primates, the size of most organs is determined by inescapable physiological rules. A species whose body weighs twice that of another, for example, requires a heart weighing almost twice as much. The sole organ that could be reduced in size without significant cost to health was the digestive tract – provided it could be supplied with foods rich in calories and nutrients.
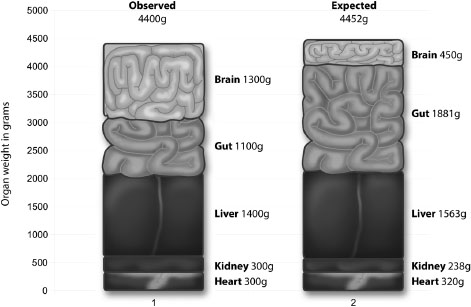
As Figure 4.1 shows, while our liver, kidneys and heart are of the size we would expect in an animal of our size, the brain is three times larger and the gut about 60% smaller than expected.20 Reducing the size of our gut allowed us to expand our brain, while keeping the total weight of our five key organs at approximately 4,400 grams, which is almost exactly in line with the 4,452 grams we would expect them to be in a primate of our size.
‘The relationship between relative brain size and relative gut size is perhaps not unexpected,’ comment Leslie Aiello and Peter Wheeler. ‘Of all the expensive tissues only the brain and the gastrointestinal tract have a significant latitude to vary in size in relation to the overall body size of the animal.’21
The size of an animal’s gut depends on the quality of its diet. Watch cows grazing in a field and you’ll quickly realise that’s almost all they ever do. For six hours a day they chomp and chew grass, hay and any other vegetation that comes their way.
While this diet provides a certain amount of carbohydrates and a little protein, the fact that their food is full of fibre, lignin (the organic substance which supports plant and wood cells) and even some silica, makes it difficult to digest. For every 24 calories a cow consumes, less than 1 calorie (0.95) is converted into new tissue, while 15 calories are expelled as faeces, urine and gas.
To achieve even this meagre energy conversion cows, in common with other grass-eating ruminants, possess four stomachs. Here the food can be partially digested before being returned to the mouth for further chewing, after which it is returned to the stomachs once more. It takes a great deal of grass to provide ruminants with the nourishment they need, which explains why most grazing animals are fairly large – they have to be able to accommodate their complex digestive systems, which can include up to 170 feet of intestine. This compares to an average length of 22 feet 6 inches (6.9 metres) in a human male and 23 feet 4 inches (7.1 metres) in a female.22
Aiello and Wheeler estimated the number of calories a species is able to save by having a small gut, and showed that the number nicely matched the extra cost of a larger brain. They concluded that because primates have smaller intestines and therefore expend less energy fuelling them, they are able to allocate more to power their bigger brains than other animals. Humans have proportionally the largest brains and smallest digestive systems of any primate. The brain’s energy requirements, and how we ‘learned’ to get greater nutrient value from food, sets up the next critical phase of understanding the brain in the context of obesity.
Some evolutionary biologists claim that the pressure for large brains, and the greater intelligence these make possible, arises chiefly from the need to outwit social rivals. The late Richard Alexander, from the University of Michigan, argued that because brainpower is critical for planning raids and winning battles, possessing a higher intellect could lead to greater success during the long history of human conflict.23
Another theory proposed that species hunting over great distances required superior brainpower to create mental maps of their territories. But while it is true that early hunter-gatherers covered distances significantly greater than those of apes, there is no discernible correlation between range size and brain size.24
A more definitely observable advantage of possessing a larger brain is that it enables more complex social groups to develop. It allows individuals to develop both ‘social intelligence’ and a ‘Theory of Mind’ – that is, the ability to recognise that other individuals think much as they do, which leads to feelings of empathy. With the finding that more sophisticated primates with bigger brains and a greater amount of tissue in the neocortex (frontal regions of the brain that contribute to long-term decision-making) live in larger groups, have more close relationships and use strategic planning more effectively than smaller brained primates, evolutionary psychologist Robin Dunbar proposed this ‘social brain’ hypothesis.25
But leaving aside the undoubted benefits of the increased intelligence which a larger brain affords, one of the most important questions is how we obtained sufficient energy to grow more brain tissue in the first place. Brain tissue is extremely ‘expensive’; it has enormous energy demands, and therefore needs a lot of fuel to keep it functioning. Where did that energy come from?
Two million years ago it is likely our diet comprised almost exclusively plant material – fruits, rhizomes, nuts, potatoes and other root vegetables, seed pods and tree gums – and perhaps meat obtained through random scavenging or occasional hunting. It was an existence characterised by brief periods of prosperity interspersed with times of extreme hardship, and even famine.26
Because the length of time an animal can survive without food increases with body size, large animals are more tolerant of variation in food availability. Seals and whales, for example, are capable of surviving off their own body reserves for extended periods, and can transfer a significant amount of energy to their offspring via their milk. Where humans are concerned, an advantage of our comparatively larger body size lies in the ability to consume low-quality ‘fallback’ foods such as potatoes and root vegetables, bulbs and corns. Meat is higher quality but harder to come by. So digestive adaptability enabled humans to intersperse grazing and consumption of plant foods with consumption of animal tissue, giving them multiple potential sources of energy. Such flexibility would have aided survival when hunting trips failed and undoubtedly helped to provide the demanding brain with the energy it needed.
However, many researchers believe there was another critical catalyst for the growth of our larger brain – at some point in our evolutionary history, we humans learned to cook.
In 1942, nutritionist Victor Lindlahr published a book entitled You Are What You Eat. The phrase quickly became a popular piece of folk wisdom. It is, however, not entirely accurate. Rather, we are what we ingest and digest, with the emphasis less on ingestion than digestion.27 The way our bodies process the nutrients we put into them is absolutely crucial.
People who eat only uncooked fruits and vegetables have to consume many more of them to prevent weight loss, since they can fully digest only a portion of what they ingest. Cooking softens hard-to-digest foods such as root vegetables, making them easier to chew and to absorb nutrients from. Heat also breaks up the long molecular chains present in many toxins, whilst also killing bacteria and other parasites in meat.
Exactly when and how mankind, the only species to cook its food, first worked out how to do so remains uncertain. One idea, which must in the absence of firm evidence be regarded as highly speculative, is that, between 700,000 and 800,000 years ago, a wildfire accidentally cooked some animals. Scavenging people that came across these found them so tasty they adopted the procedure for themselves.28
It is possible, but again mere speculation, that the first cooking involved simply tossing the raw meat onto an open fire. Some time later spit-roasting likely became the favoured method; one can imagine hunters returning with their prey on the end of a spear and holding it over an open fire, turning the meat to ensure it was cooked evenly. The invention of sharp tools, first in stone and later in metal, enabled them to cut meat into smaller pieces to cook it faster. Food might also have been boiled, using the shells of large molluscs or turtles as vessels; clay pots were only created in about 5000 BCE.
‘Cooking increased the value of our food. It changed our bodies, our brains, our use of time, and our social lives,’ says Richard Wrangham in his book Catching Fire: How Cooking Made Us Human. He believes that the ‘transformative moment that gave rise to the genus Homo, one of the great transitions in the history of life, stemmed from the control of fire and the advent of cooked meals.’29
The most widely accepted evidence for mankind’s first controlled use of fire comes from Gesher Benot Ya’aqov in Israel. At this waterlogged site on the shores of Lake Hula, in the northern Dead Sea Rift, archaeologists identified early hearths, on which willow, poplar, ash and wild olive had been burned.
Evidence was also found for the roasting of oats, wild grapevine and barley as well as meat. The animals consumed included horses, deer, rhino, hippo and birds. Bones were unearthed showing cut marks and patterns of breakage indicating the extraction of marrow. On the basis of this evidence, Professor Naama Goren-Inbar and her colleagues from the Institute of Archaeology at Hebrew University concluded that: ‘The hominins who frequented the shores of the lake for over 100,000 years knew how to use fire and exercised that knowledge repeatedly throughout much of the . . . period. The domestication of fire by hominins surely led to dramatic changes in behavior connected with diet, defence and social interaction.’30 By increasing the efficiency with which food could be digested, cooking freed our early ancestors from the need to spend so much time foraging, chewing, and digesting. The higher-quality diet that resulted from cooking enabled proto-humans to absorb energy from food more efficiently. This meant they were able to get by with a smaller gut, which allowed an increase in the size and sophistication of their brains.
While our distant ancestors might have lived in deprivation, always unsure where their next meal was coming from, today, in the developed world at least, consumers enjoy unlimited access to brain fuel in the form of HED (high energy-dense) foods. Indeed, the chief challenge in the twenty-first century is to avoid consuming too much glucose. So what happens when we no longer need our guts to work so effectively, when we have more fuel than our primitive ancestors could have ever dreamed was possible?
Half a century ago, James Neel, a professor in the department of Human Genetics at the University of Michigan Medical School, published a controversial paper entitled, ‘Diabetes Mellitus: A “Thrifty” Genotype Rendered Detrimental by “Progress”?’31 In it he sought to address a medical paradox that had long puzzled health workers. As mentioned in Chapter 3, diabetes is a serious condition, and therefore impairs evolutionary success by making sufferers less likely to reproduce. This seems contrary to basic Darwinian principles – why should such harmful genes have been favoured by natural selection? The answer, suggests Neel in his ‘thrifty genotype’ hypothesis, is that they once aided survival.
‘During the first 99 per cent or more of man’s life on earth, while he existed as a hunter and gatherer, it was often feast or famine,’ he points out. ‘Periods of gorging alternated with periods of greatly reduced food intake. The individual whose pancreatic responses minimized post-prandial glycosuria (the passing of glucose in the urine) might have, during a period of starvation, an extra pound of adipose reserve.’32
So Neel argues that insulin resistance, which allows for huge blood sugar spikes and ultimately favours fat storage, would actually have been beneficial for our distant ancestors. It would limit the liver’s ability to absorb and metabolise glucose, ultimately leading to increased fat storage. This additional fat would be immensely useful as a source of energy during periods of famine. However, in more luxurious circumstances this same trait contributes to an increased risk of developing Type II diabetes, which is what we are seeing happen today. No matter how helpful these genes might have been during the Palaeolithic era, they are distinctly unhelpful in conjunction with a twenty-first century diet.
As we have explained, between birth and twelve months babies become increasingly fat. Later, between one and five, when children grow taller, they no longer require large stores of adipose tissue, and gradually become slimmer. Then, around the age of six, the so called ‘adiposity rebound’ occurs. The child starts putting on weight again, laying down fat deposits once more. Exactly when this rebound occurs is of critical importance to the child’s health in later life. For reasons that are still not fully understood, low weight gain during the early years triggers an early adiposity rebound. And an early adiposity rebound is a strong predictor of both Type II diabetes and obesity in adulthood.33
More than twenty years ago, David Barker, Professor of Clinical Epidemiology at the University of Southampton, and one of the twentieth century’s most influential epidemiologists, demonstrated that babies with a below-average birth weight are at greater risk of developing a wide range of diseases later in life.34 He was not only concerned with the effect of malnutrition due to poverty on babies, but to that of any diet unbalanced in macronutrients and deficient in micronutrients. Diets, in fact, very similar to those followed by millions of people worldwide today.
This ‘foetal programming hypothesis’, which the British Medical Journal later named the ‘Barker Hypothesis’, transformed thinking about the causes of obesity and obesity-related disorders such as coronary heart disease, stroke, hypertension and diabetes.35 Challenging the notion these can be explained by a combination of bad genes and unhealthy lifestyle, Dr Barker argued that the root of all these health problems is to be found in the womb. He claimed it is the nutritional environment in which foetus and infant develop which permanently ‘programs’ metabolism and determines the ‘pathologies of old age’.
‘The nourishment a baby receives from its mother, and its exposure to infection after birth’, he says, ‘determine its susceptibility to chronic disease in later life.’36 The very start of life is the critical developmental window on which the individual’s future health and wellbeing depend.
To understand why things go wrong, let’s focus on the role of insulin as the baby develops in the womb.
Manufactured by beta cells in the pancreas, insulin is the sole hormone responsible for energy storage in vertebrates, including humans. Only the brain and liver do not require it in order to utilise glucose. Because of insulin’s unique role, a great many metabolic problems arise if there are any difficulties over its release.
Additionally, insulin controls the rate at which a foetus develops by matching speed of growth to the availability of food. If the mother is poorly nourished, so too is the foetus. In this situation an automatic survival mechanism kicks in. Muscle growth is sacrificed, since it is of relatively low importance to the developing baby. By keeping more sugar in the blood, rather than allowing it to be stored in the muscles, nature strives to protect the developing brain of the baby. After a while, in conditions of nutritional scarcity, this cautious approach to sugar management – the ‘thrifty genotype’ discussed earlier – becomes ‘hard-wired’ into the brain and so persists throughout life.
If, following birth, children who have developed in such conditions find themselves in an environment where food is freely available, they have an increased risk of developing diabetes, either because their bodies are incapable of producing sufficient insulin, or because their tissues do not respond to it in the normal manner.
A reduced ability to produce insulin, combined with a need for it in larger quantities because the body is less responsive to it, makes it impossible for the individual to maintain blood sugar levels within normal limits. The individual has developed what doctors term ‘insulin resistance’. If a person is insulin resistant, their muscle, fat and liver cells do not respond appropriately to the hormone. The body therefore needs to produce more insulin in the pancreas to aid the transfer of glucose to those cells. If it is unable to meet the demand, excess glucose accumulates in the bloodstream. If this glucose is not used by the cells, it is converted into fat. Thus, by compromising the body’s ability to utilise glucose, insulin resistance can cause weight gain.
A previously advantageous strategy for safeguarding the brain is now transformed into a life-threatening liability. The blood becomes flooded with sugar and weight gain occurs, which then increases the body’s resistance to insulin. When the body has too much adipose tissue and too much blood glucose over a prolonged period, diabetes develops.37 So we have good reason to think that the diet of a mother during pregnancy can have long-term adverse effects on a child’s ability to control their weight, and serious implications regarding their likelihood of developing diabetes.
It’s a fact that obese parents tend to have obese children. While only 10% of children with slender parents become obese, approximately 40% of those with one obese parent do. Where both parents are significantly overweight, this proportion rises to 70%.38
The question posed by Professor Alexandra Logue is: how much ‘is due to eating or exercise habits transmitted environmentally from parents to children, and how much is due to genes transmitted from parents to children?’39
To investigate the link between the mother’s weight and her unborn child’s risk of obesity, Professor Neena Modi and her colleagues, from the Department of Medicine at Imperial College London, used Magnetic Resonance Imaging to scan over a hundred newborn babies as they slept. We will have more to say about this powerful technique for imaging both body and brain in Chapter 7. The scans enabled them to measure the amount of fat in the infant’s liver cells as well as the total amount and distribution of fat in their bodies.
What the researchers found surprised and concerned them. Liver cell fat and total fat, especially around the abdomen, increased across the entire range of children in correlation with their mothers’ BMI; the more overweight the mother, the fatter her baby.40 This study produced the first evidence of biological changes which, combined with an unhealthy lifestyle, could put these infants’ health in jeopardy as they grew older.
‘Fatter women have fatter babies and there is more fat in the babies’ livers’, Neena Modi concluded. ‘If these effects persist through childhood and beyond, they could put the child at risk of lifelong metabolic health problems. There is growing evidence that a baby’s development before birth has a major impact on their health in later life. This means that the prevention of obesity needs to begin in the womb.’41 In short, the results of Professor Modi’s work bear out the Barker Hypothesis.
The accumulation of excess adipose tissue, fat, is not the result of greed or laziness. Rather, since glucose is the primary fuel of the brain, it may be the case that the intense psychological pressure and stress we feel on a daily basis causes the brain to cry out for more fuel. Thus, excess adipose tissue is perhaps better viewed as an unintended consequence of the brain’s chronic demand for more glucose. The next problem, of course, is that excess adipose tissue makes further demands on the brain by transmitting a signal that the body is in fact hungry, which is something we examine with the discovery of the hormone leptin, covered in the next chapter.
In this chapter we have explained some of the reasons that we strongly believe that obesity is not down to greed or laziness.42 Whilst poor diet choices and insufficient exercise play an important role in weight gain, ultimately it occurs in response to demands from the brain. The brain is, in turn, affected by hormones circulating in the bloodstream. To understand what these hormones are, and how they exert their influence over consumption, we must investigate the secret life of fat.