Microscopic Structure of Muscle
Microscopic Structure of MuscleIN THIS CHAPTER:
Microscopic Structure of Muscle
Muscle Contraction
Macroscopic Structure of Muscle
Solved Problems
There are three types of muscle tissue: smooth, cardiac, and skeletal. Each type has a different structure and function, and each occurs in a different location in the body.
Muscle functions include:
• Motion. Body movements such as walking, breathing, and speaking, as well as movements associated with digestion and the flow of fluids.
• Heat production.
• Posture and body support.
Because muscle cells resemble tiny threads, they are called muscle fibers. Each skeletal muscle fiber is a multinucleated, striated cell containing a large number of rodlike myofibrils that extend the entire length of the cell. Each myofibril is composed of still smaller units, called myofilaments (or filaments). Thin myofilaments are composed primarily of the contractile protein actin and thick myofilaments contain primarily the contractile protein myosin.
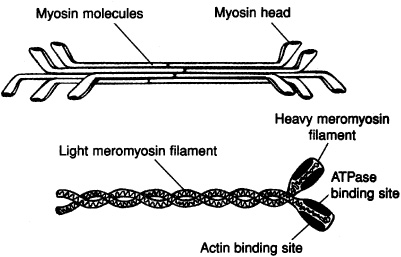
Thick filament: Shaped like a golf club, each myosin protein has a long rod portion, and a globular head. The myosin head contains an actin binding site and a myosin ATPase binding site. The strands of the rod portion bind together with their globular head projecting outward to form the thick filaments that lie between the thin filaments (Figure 7-1).
Thin filament: The thin filament is composed of the proteins actin, tropomyosin, and troponin. Two long strands of actin molecules form the backbone of the thin myofilaments. Long, thin, threadlike tropomyosin proteins spiral around and cover the myosin binding sites on the actin helix. The troponin molecule fastens the ends of the tropomyosin molecule to the actin helix (Figure 7-2). In skeletal and cardiac muscle the thick and thin myofilaments overlap within the myofibril in a distinctive pattern called a sarcomere. The sarcomere is the structural and functional unit of the myofibril.
Figure 7-1. Structure of thick myofilaments.
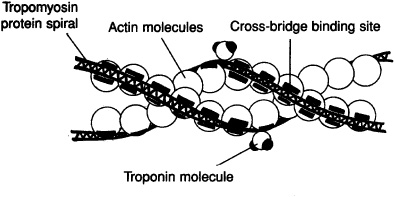
Figure 7-2. Structure of thin myofilaments.
The regular overlapping pattern of thin and thick filaments within the myofibrils is responsible for the cross-banding striations seen in skeletal and cardiac muscle. The dark bands contain the thick filaments and are called A bands. The lighter bands, the I bands, are regions where only thin filaments occur. The I bands are bisected by dark Z lines where the thin filaments of adjacent sarcomeres join.
The sarcolemma (cell membrane) of a muscle fiber encloses the sarcoplasm (cytoplasm). The sarcoplasm is permeated by a network of membranous channels, called the sarcoplasmic (endoplasmic) reticulum, that forms sleeves around the myofibrils. The longitudinal tubes of sarcoplasmic reticulum empty into expanded chambers called terminal cisternae. Calcium ions (Ca2+) are stored in the terminal cisternae and play an important role in regulating muscle contraction.
Transverse tubules (T tubules) are internal extensions of the sarcolemma that extend perpendicular to the sarcoplasmic reticulum. The T tubules pass between adjacent segments of terminal cisternae and penetrate deep into the interior of the muscle fiber to allow the action potential from the cell surface to be delivered into the center of the fiber.
In the sliding filament theory of contraction, a skeletal muscle fiber, together with all of its myofibrils, shortens by movement of the insertion toward the origin of the muscle. Shortening of the myofibrils is caused by shortening of the sarcomeres, which is accomplished by sliding of the myofilaments. The mechanism that produces the sliding of the thin (actin) myofilaments over the thick (myosin) myofilaments during contraction is outlined in the steps below.

1. Stimulation via the neurotransmitter acetylcholine across the neuromuscular junction initiates an action potential on the sarcolemma of the muscle fiber. This action potential spreads along the sarcolemma and is transmitted into the muscle fiber through the T tubules.
2. The T tubule potential causes the terminal cisternae of the sarcoplasmic reticulum to release calcium ions (Ca2+) in the immediate vicinity of each myofibril.
3. Ca2+ ions bind to and change the protein structure of the troponin molecules attached to the tropomyosin molecules on the thin filaments. This change causes the tropomyosin to move aside, exposing the actin binding sites.
4. Myosin cross bridges bind to actin. Upon binding, the cocked (energized) myosin head (HMM) undergoes a conformational change, causing the head to tilt. This pulls the actin filament over the myosin filament in an action called a power stroke.
5. After the power stroke, ATP binds the HMM, causing detachment of the cross bridge from the actin binding sites. The enzyme ATPase within the HMM cleaves ATP to ADP + energy; the energy is used to recock the HMM. The HMM can then bind with another actin site (if still exposed due to the presence of Ca2+) and produce another power stroke.
6. Repeated power strokes successfully pull in the thin filaments. This sliding with a ratchet mechanism involves numerous actin binding sites and myosin cross bridges and constitutes a single muscle contraction.
7. Once the action potential ceases, the sarcoplasmic reticulum actively transports Ca2+ from the cytoplasm into the terminal cisternae. Without calcium ions, the troponin molecule resumes its original shape so that the tropomyosin is pulled back over the myosin binding sites of the thin filament. The thin filaments slide back to their noncontracted position, and the muscle is relaxed.
Stimulation from a motor neuron initiates the contraction of skeletal muscle. The space between the axon terminal of a motor neuron and the cell membrane of a muscle fiber is called the neuromuscular (myoneural) junction (Figure 7-3).
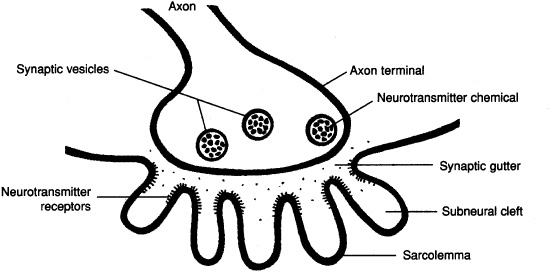
Figure 7-3. The neuromuscular junction.
The action potential travels along the motor neuron to the axon terminal, where it causes an influx of calcium ions. The calcium ions cause synaptic vesicles to release acetylcholine, which diffuses across the synaptic gutter and combines with specific receptors on the sarcolemma. An action potential radiates over the sarcolemma, initiating the sequence of events described above.
A motor unit consists of a single motor neuron together with the specific skeletal muscle fibers that it innervates. A large motor unit serves many muscle fibers, a small motor unit serves relatively few muscle fibers. Contraction of a skeletal muscle requires recruitment of motor units. Few motor units are recruited when fine, highly coordinated movements are being performed. Many motor units are recruited when strength is needed. Individual muscle fibers of a motor unit respond to an electrical stimulus in three phases (Figure 7-4):
1. the latent period between stimulation and the start of contraction;
2. the contraction period, or duration when work is being accomplished; and
3. the relaxation period, or recovery of the muscle fiber.
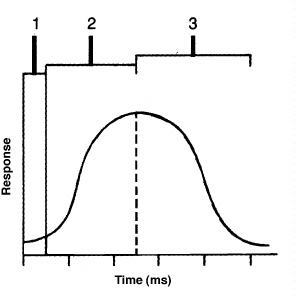
Figure 7-4. The activity of a muscle fiber in response to a stimulus.
Types of Skeletal Muscle Fibers
Fast-twitch fibers: Large fibers with large amounts of glycogen; low amounts of O2 carrying pigment myoglobin; anaerobic pathways to produce ATP; power and speed
Slow-twitch fibers: Small fibers with low glycogen content; high amount of myoglobin; aerobic pathways to produce ATP; resist fatigue, endurance.
Intermediate fibers: Intermediate in size and glycogen content; high amount of myoglobin; both aerobic and anaerobic pathways to produce ATP.
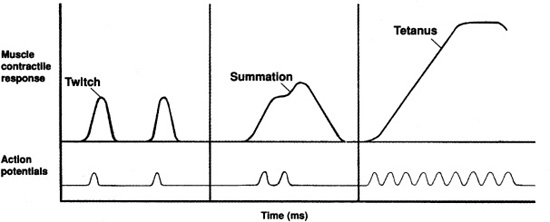
A single action potential to the muscle fibers of a motor unit produces a muscle twitch, a rapid, unsustained contraction (Figure 7-5). If the impulses are applied to a muscle in rapid succession through several motor units, one twitch will not have completely ended before the next begins. Since the muscle is already in a partially contracted state when the second twitch begins, the degree of muscle shortening in the second contraction will be slightly greater than the shortening that occurs with a single twitch. The additional shortening due to the rapid succession of two or more action potentials is termed summation. At high stimulation frequencies, the overlapping twitches sum to one strong, steady contraction call tetanus.
Figure 7-5. Patterns of muscle twitch, summation, and tetanus.
Skeletal muscle tissue, in association with connective tissue, is organized into muscle bundles. This muscle architecture determines the force and direction of the contracting muscle fibers. Muscle fibers may be organized with parallel fibers, convergent fibers, pennate (feather-shaped) fibers, or as a sphincter (circular) muscle.
Loose fibrous connective tissues bind muscles at various levels to unify the force of contraction. Surrounding each muscle fiber is a connective tissue called the endomysium. A group of individual muscle fibers is bound together by another connective tissue, the perimysium, to form a fasciculus. Many fasciculi make up an individual muscle. Each muscle is surrounded by a third connective tissue, the epimysium. These three connective tissues are continuous with the tendon that secures the muscle to bone.
Muscles are attached to the skeleton at two locations. The origin of a muscle is the more stationary attachment of the muscle; the insertion the more movable attachment. In the appendages, the origin is generally proximal in position and the insertion is distal in position.
• the difference between muscle fibers, myofibrils, and myofilaments.
• the mechanism of muscle contraction and the importance of Ca2+ and ATP in this process.
• the organization and function of skeletal muscles: motor units, fiber types, types of contractions, and gross structure.
Matching

True or False
____ 1. Actin is found only in the striated fibers of cardiac and skeletal muscle tissues. (False)
____ 2. Fasciculi are enclosed in a covering of perimysium. (True)
____ 3. Thin myofilaments are primarily composed of myosin proteins. (False)