Theories pass. The frog remains.
JEAN ROSTAND, INQUIÉTUDES D’UN BIOLOGISTE
“MAN, A WITNESS OF THE FLOOD”
In the early eighteenth century, scholars were still divided over the origin and nature of fossils and offered many explanations for the presence of these strange objects found in rocks. The word “fossil” comes from the Latin term fossilis (obtained by digging), so anything dug out of rocks (including crystals, concretions, and many other nonbiological objects) were originally called fossils. Some scientists thought that fossils were works of the devil, placed in rocks to confuse the faithful and spread doubt. Others argued that they grew in rocks under the influence of mystical “plastic forces” (vis plastica) or that some creatures had crept into crevices, been crushed, and died, leaving their skeletons encased in stone. Only a minority of scholars connected the fossilized shells of clams and snails to their modern descendants.
Many fossils were simply unrecognizable at the time because they looked like no extant creature. The strange triangular objects known as “tongue stones” (
glossopetrae) were thought to have fallen from the sky and to have magical properties, including the ability to heal snake bites and detoxify poisons. But in 1669, the Danish doctor Niels Steensen (known to us by his Latinized name, Nicholas Steno) saw “tongue stones” in the mouth of a shark and realized that they were teeth. Most people thought that ammonites were the remains of coiled snakes because the chambered nautilus would not be discovered until the early nineteenth century. The stem pieces or columnals of crinoids were believed to be stars that had fallen from the heavens.
Figure 11.1
Johann Scheuchzer’s “Homo diluvii testis,” displayed at the Teylers Museum in Haarlem, Netherlands. (From Donald R. Prothero, Bringing Fossils to Life: An Introduction to Paleobiology, 3rd ed. [New York: Columbia University Press, 2013], fig. 1.4)
In particular, the Bible still influenced ideas about fossils. In 1726, for example, the Swiss naturalist Johann Scheuchzer described a fossil as “the bony skeleton of one of those infamous men whose sins brought upon the world the dire misfortune of the Deluge.” It was a large skeleton, about 1 meter (3.3 feet) long from the head to the hip bones; had a skull and arms and a backbone; and had been found in the rocks. Therefore, it must be a human who had died in Noah’s flood. Scheuchzer named it
Homo diluvii testis (Man, a witness of the Flood) (
figure 11.1). But in 1758, the pioneering naturalist Johannes Gesner disagreed, believing it to be a catfish! Then in 1777, Petrus Camper argued that it was a lizard. In 1802, Martin van Maur bought the specimen for the Teylers Museum in Haarlem, where it still resides. In 1836, it was formally named
Andrias scheuchzeri, which translates to “Scheuchzer’s image of man.”
The mistake was not rectified until almost a century after Scheuchzer first described it. After Napoleon annexed the Netherlands, the specimen found its way to Paris, where the great Baron Georges Cuvier, the founder of vertebrate paleontology and comparative anatomy, got to work on it. He prepared the skeleton in the slab to better expose the bones and found much more detail than had been visible originally, especially in the arms. In addition, he had spent his professional life studying comparative anatomy, so he knew at once that it was not a human skeleton. A few comparisons, and Cuvier realized that it was not even a primate or a mammal—but a gigantic salamander!
Such gigantic salamanders are not extinct. Two species in Japan and China are even larger than Scheuchzer’s fossil (
figure 11.2). The Chinese giant salamander is almost 2 meters (6.6 feet) long and can weigh as much as 36 kilograms (80 pounds)! It is placed in the same genus as Scheuchzer’s fossil, but is named
Andrias davidianus. It lives in rocky hill streams and lakes with clear water, usually found in forested regions, as well as at altitudes of 100 to 1500 meters (330 to 4920 feet). The Japanese species is named
Andrias japonica, is slightly smaller than the Chinese giant salamander, and inhabits a similar environment. Both species are endangered, since their habitats are being destroyed and such large aquatic animals need a lot of territory to survive. In addition, they are being poached for traditional Chinese medicine, which is already driving rhinoceroses, tigers, pangolins, and many other animals to extinction as well.
Figure 11.2
The Chinese giant salamander. (Photograph courtesy Luke Linhoff)
LIVING ON BOTH SIDES
In
chapter 10, we saw how amphibians arose from lobe-finned fish in the Late Devonian. But how did they evolve into the familiar groups of living amphibians, especially the frogs, toads, and salamanders? Once again, the fossil record has produced some amazing specimens that show the stages of this evolutionary history.
The word “amphibian” comes from the Greek term amphibion (living on both sides)—that is, both in water and on land—and “living on both sides” is one of the distinguishing features of amphibians. Most have the ability to thrive in both environments, as long as they can get moisture. Desert toads have adapted to adapt to a world with almost no water and eke out an existence underground, keeping cool and moist. However, most amphibians still need moist places in which to lay their eggs and complete their life cycle (although a handful actually give birth to live young and skip the egg stage altogether).
The living amphibians are tremendously diverse, with over 5700 known species. More than 4800 of them are frogs and toads, but only 655 are salamanders and newts. In addition, there are about 200 species in a third group of amphibians: the apodans, or caecilians. The legless apodans burrow underground mostly in tropical soils of South America, Africa, and Asia. They have tiny eyes that can sense light and dark, and some have eyes at the tip of sensory tentacles, but most are blind. To the nonspecialist, they look almost like giant earthworms.
Amphibians range enormously in size, from the tiny New Guinean frog
Paedophryne amanuensis, which is only 7.7 millimeters (0.3 inch) long, to the huge Chinese giant salamander. Salamanders and newts retain the simple elongate body form, with a long tail and four simple limbs, of the most primitive amphibians (such as
Tiktaalik,
Ichthyostega, and
Acanthostega [
chapter 10]).
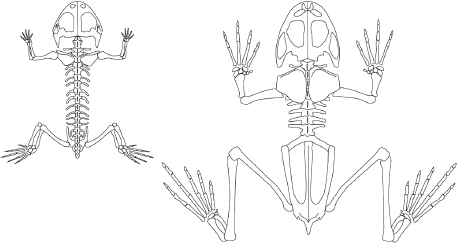
Figure 11.3
Comparison of the skeleton of Triadobatrachus (left) with that of a modern frog (right). Although they look superficially similar, Triadobatrachus was much more primitive than any modern frog in having many trunk vertebrae, small simple hips rather than an elongate hip structure, small fore- and hind limbs that did not allow it to jump, a slightly longer tail, and a much more primitive skull. (Drawing by Mary P. Williams)
Frogs are the most spectacularly divergent from this ancestral body plan of all the living amphibians. As anyone who has dissected a frog in high-school biology class knows, they are truly unique in their body design (
figure 11.3). Although adult frogs and toads have no tail, their larvae (tadpoles) hatch with a tail that is resorbed into their body as they mature. The head of frogs is short, with a blunt broad snout that allows them to open their mouth wide as they capture food (often using a long sticky tongue). Their very long muscular hind legs enable them to make huge leaps (both to catch prey and to escape predators) as well as swim with great power. The trunk of the frog skeleton is also short, with tiny stumpy ribs and very elongated hip bones to support the hind-leg muscles. Since frogs cannot use their ribs for breathing, they use an inflatable pouch in their throat that can pump air in and out (as well as make a variety of sounds). Frogs range tremendously in size, from the tiny New Guinean frog to the Goliath frog, which is more than 300 millimeters (12 inches) long and weighs 3 kilograms (7 pounds). It is so big that it eats birds and small mammals, as well as insects.
If the Goliath frog were not impressive enough, in 1993 a group of scientists working in the Upper Cretaceous rocks of Madagascar found the fossil of an even bigger frog. After 15 years of fitting all the pieces together (including most of the skull from 75 fragments), they published a description of it in 2008. They named it Beelzebufo ampinga (devil’s toad). The genus name is a composite of Beelzebub (Lord of the Flies), another name for the devil, and Bufo, the genus of common toads; the species name is Malagasy for “shield.” It was a ceratophrynine, a member of the group known as the “horned toads” of South America, so this family once extended across Gondwana, which included most of the present-day Southern Hemisphere. Its most remarkable feature was its size. Based on the nearly complete skeleton, it was 40 centimeters (16 inches) long and weighed 4 kilograms (9 pounds)—one-third again as large as the Goliath frog! It had a very large head and a wide mouth, and it is speculated that it could eat even baby dinosaurs, which roamed Madagascar at the time.
RICHES OF THE RED BEDS
This is just a glimpse of the range of size and diversity of living amphibians. What about their fossil ancestors? Starting with “fishibians” (
chapter 10), there was a huge evolutionary explosion of different kinds of amphibians during the Carboniferous (355 to 300 million years ago) and Permian (300 to 250 million years ago). Most belong to three major groups that are extinct, but they were once the largest and most dominant animals on land until reptiles took over that role in the Early Permian.
By far the best place to collect Early Permian amphibians and contemporaneous land animals are the red beds of northern Texas, especially in the area around Wichita Falls and Seymour (and across the state line in Oklahoma). These incredible fossil deposits were discovered by the pioneering paleontologist Edward Drinker Cope in 1877. Working with just a horse and wagon and one or two local helpers, he found the ground literally covered with fragments of bone, along with skulls and skeletons. He collected a full wagonload in just a few days, thus beginning the long tradition of American paleontologists collecting in these rich deposits, and shipped them back to Philadelphia for study.
Almost every paleontologist who has published on the evolution of early reptiles and amphibians has collected in the red beds of Texas, including the giants of the field whose name every paleontologist knows well: Samuel Wendell Williston of the University of Kansas (in the 1890s) and the University of Chicago (until his death in 1918), Alfred S. Romer of the University of Chicago (in the 1920s) and Harvard (until the 1970s), and Everett “Ole” Olsen of the University of Chicago (and later UCLA).
The conditions for collecting are no picnic. The area is blazing hot in the summer, with windstorms that blow red dust into everything: food, beverages, equipment, and eyes and other sensitive areas. The groundwater is as hot as tea and nasty tasting, filled with pink mud and alkali, so those who drink too much of it get kidney stones. Once they find a good locality, collectors have to dig in deep and hunker down, trying to keep cool and avoid breathing the dust.
But the rewards are worth it! The most common animal in the red beds is the fin-backed, tiger-size predator
Dimetrodon, familiar from dinosaur plastic toy sets and children’s dinosaur books (
chapter 19). However,
Dimetrodon was not a dinosaur, but a very early member of the lineage that gave rise to the mammals, known as synapsids or “protomammals” (once called mammal-like reptiles, although synapsids were not reptiles). Most specimens reached 2 to 4 meters (7 to 14 feet) in length, weighed up to 270 kilograms (600 pounds), and had spines 1.2 meters (4 feet) tall on their back to support their fins. They were the top predator of their time, feeding on smaller fin-backed synapsids like the herbivore
Edaphosaurus, as well as a variety of primitive true reptiles, such as the lizard-size
Captorhinus, which was closely related to turtles.
But the synapsid and reptile denizens of the Texas red beds are only a tiny part of the story. Even though Dimetrodon ruled the planet in the Early Permian, amphibians reached their acme of size and diversity, and many of them were top predators that competed for food in this harsh landscape.
WHEN AMPHIBIANS RULED THE WORLD
The most abundant and impressive of the three groups of Late Paleozoic amphibians was the temnospondyls (formerly, labyrinthodonts). Most resembled fat crocodiles, with long trunks and tails as well as strong limbs that sprawled out to the sides. Unlike crocodiles, however, they had huge flattened skulls with eye sockets that pointed upward, and rows of sharp conical teeth arrayed around their large snouts. The head of some specialized temnospondyls known as archegosaurs superficially resembled that of crocodiles, with a long narrow snout. One of them was
Prionosuchus, from the Pedro do Fogo Formation in Brazil, which dates to the Middle Permian (270 million years ago).
Prionosuchus lived in lagoons and rivers, and had not only a crocodile-shaped body, but a long very narrow snout that was specialized for catching fish and other aquatic prey, as does the gavial (or gharial). If it was truly 9 meters (30 feet) long, as some claim,
Prionosuchus was the largest amphibian that has ever lived—and larger than any living crocodile as well—although others argue that the estimates of the tail and body are too long, and it may have been only 5 meters (16 feet) in length.
The earliest temnospondyls were only about 1 meter (3.3 feet) long, but by the Permian, they were among the largest land creatures the planet had ever seen. One of the commonest fossils in the Early Permian red beds of Texas is that of
Eryops, a big temnospondyl known from numerous complete skeletons (
figure 11.4A). It had a sprawling body more than 2 meters (6.6 feet) long, with a robust tail and limbs, and a skull well over 60 centimeters (2 feet) long in big individuals!
Eryops was one of the largest terrestrial animals of the Early Permian, capable of hunting prey both in water and on land. The slightly more primitive
Edops, also from Early Permian red beds of Texas, had an even longer skull and thus was even larger than
Eryops.
By the Late Permian, the large terrestrial temnospondyls had retreated to a completely aquatic lifestyle, possibly due to competition from all the large predatory synapsids on land at the time. Temnospondyls managed to survive the worst mass extinction in Earth history at the end of the Permian (250 million years ago). They straggled on into the Triassic (250 to 200 million years ago), when they were common in the swamps and lake deposits of places like the Petrified Forest in Arizona. These last temnospondyls had weak legs that would not have supported them on land, flattened heads with eyes that looked upward only, and huge flat bodies that were adapted to living in shallow water and feeding on aquatic prey.
Figure 11.4
Early amphibians: (A) the temnospondyl Eryops; (B) reconstruction of the lepospondyl Diplocaulus; (C) the anthracosaur Seymouria. ([A and C] courtesy Wikimedia Commons; [B] courtesy Nobumichi Tamura)
The second group of extinct amphibians was the lepospondyls, which lived from the Early Carboniferous to the Early Permian, but only in Europe and North America. Most were smaller than the temnospondyls that lived alongside them and had long salamander-like bodies with tiny legs, suggesting that they were mainly aquatic. Some, such as the aistopods, lost their legs entirely and looked like aquatic snakes. Others, the microsaurs, were more lizard-like in body form, with deep skulls and strong limbs. The most famous of the lepospondyls is the strange-looking
Diplocaulus (see
figure 11.4B). Best known from the Early Permian red beds of Texas, it was one of the largest of the lepospondyls, reaching a length of 1 meter (3.3 feet), with a stocky salamander-like body. It had armor plating over most of its body and strong, wide jaws.
But it was the head of Diplocaulus that was truly bizarre. It was shaped like a boomerang, with a flattened skull from each side of which extended a large flattened “horn” and eye sockets that pointed straight up. The function of these odd “horns” is still controversial. Some have argued that they were used as a hydrofoil, allowing Diplocaulus to swim smoothly in an up-and-down motion with the boomerang head shape providing lift. But its body was relatively weakly built and did not have the robust bones needed to support strong swimming muscles. Others have suggested that the head shape would have made it difficult for a predator to eat Diplocaulus head first, since the “horns” would have made the head too wide to swallow, even for the largest Early Permian predators. The upward-pointing eyes suggest that Diplocaulus was more of an ambush predator that lay in the bottom of streams and ponds, and then lunged forward and upward to catch its prey with its strong jaws, possibly stunning it with a blow from its “horns.” The most likely hypothesis, however, is that the “horns” were analogous to the horns and antlers of antelope and deer. Males use their horns and antlers primarily as a display structure to advertise their strength and dominance while trying to find mates. That the growth of these “horns” can be traced through their younger stages and that there seem to have been both robust males and smaller-horned females appear to make this hypothesis most likely.
The third group of extinct amphibians is known as the “anthracosaurs,” a wastebasket group for all the more advanced amphibians that are on the lineage leading to reptiles (see
figure 11.4C). The Texas red beds are full of some amazing ones, including the 3-meter (10-foot) long, hippo-size herbivore
Diadectes, and the extremely reptile-like
Seymouria (named after Seymour, Texas, in the heart of the red beds).
FINDING THE “FROGAMANDER”
The giants of the mid-twentieth-century rush to the Texas red beds (such as Romer and Olson) are gone now, but their students continued to visit and collect important fossils. Some of the foremost successors were Robert Carroll of the Redpath Museum in Montreal (a student of Romer at Harvard), Robert Reisz (the first student of Carroll, now at the University of Toronto), the late Nicholas Hotton of the Smithsonian Institution (a student of Romer and Olson at Chicago), and the late Peter Vaughn (a student of Romer who trained many paleontologists during his career at UCLA, along with Olson). The current generation of paleontologists, intellectual grandchildren of Romer and Olson, have been making many important discoveries.
During an expedition to the Seymour area in 1994, undertaken by the Smithsonian and led by Hotton, the crew was working a locality nicknamed Don’s Dump Fish Quarry. They found many fossil fish and a number of amphibians, but there was no time to clean all the fossils and do a detailed study in the field. According to the story, Hotton recognized the importance of one particular fossil (found by Peter Krohler, a curatorial assistant at the Smithsonian) and kept it in his pocket with a slip of paper on which was written “Froggie.” But Hotton died in 1999 and never got the chance to study it or publish it.
Five years later, a group of younger scientists retrieved the unstudied specimen from the collections and spent countless hours finishing the preparation on it to completely expose the fossil (which was only partly visible when Hotton had it). Finally, in 2008, Hotton’s “Froggie” was described and published, 14 years after it was found. The authors of the paper included Jason S. Anderson of the University of Calgary (a student of both Carroll and Reisz), plus Robert Reisz, Stuart Sumida of California State University, San Bernardino (a student of Vaughn), and Nadia Fröbisch of the Museum für Naturkunde in Berlin (a student of Carroll). They named it Gerobatrachus hottoni (Hotton’s ancient frog), although the press labeled it the “Frogamander” as it spread the news of the discovery.
The specimen itself is a nearly complete skeleton only 11 centimeters (4.3 inches) long, found lying on its back with some of the hip region, tail, and shoulder bones missing (
figure 11.5A). What first catches your eye when you see the fossil is the combination of a salamander-like body with a broad flat frog-like snout (hence the nickname “Frogamander”). It has many other anatomical features of the skull and skeleton typical of frogs, especially the large eardrum. Most important, its teeth are attached to the jaw on tiny pedestals with a distinct base (
pedicellate teeth), a feature unique to the living amphibians and just a few other extinct amphibians.
Figure 11.5
Gerobatrachus hottoni: (A) the only fossil; (B) reconstruction of its appearance in life. ([A] courtesy Diane Scott and Jason Anderson; [B] courtesy Nobumichi Tamura)
Fossils that do not fit into modern groups, but are squarely between them, are true transitional fossils, sometimes called (improperly) “missing links.”
Gerobatrachus is the perfect transitional fossil linking frogs and salamanders. The oldest known salamander is
Karaurus sharovi, from the Late Jurassic (about 150 million years old) of Kazakhstan. The oldest known frog is
Triadobatrachus massinoti, from the Early Triassic (240 million years old) of Madagascar (
figure 11.6; see
figure 11.3).
Triadobatrachus looks similar to living frogs, with its broad snout and long webbed feet, except that it had a long trunk region with 14 vertebrae in its spinal column; all modern frogs have shorter trunks with four to nine vertebrae. It still had a short tail that was not lost, even in adults, unlike any living frog. Its hind legs were larger than those of any salamander, but nowhere near the large muscular legs of all modern frogs, so
Triadobatrachus could swim strongly but not jump. All these features, and many others, make
Triadobatrachus the perfect transitional fossil between modern frogs and more primitive forms like
Gerobatrachus, the “Frogamander.”
At 290 million years old, Gerobatrachus is much older than any member of the frog or the salamander lineage, and it is so primitive in its features that it cannot be called either a frog or a salamander. It contributes to the evidence that frogs and salamanders were not created as separate “kinds” but evolved from common ancestors, one of which could have been Gerobatrachus.
Figure 11.6
Reconstruction of the primitive Triassic frog Triadobatrachus. (Courtesy Nobumichi Tamura)
SEE IT FOR YOURSELF!
The “Frogamander” is not on display at any museum, as far as I know. However, large fossils of the Permian amphibians of Texas, including
Eryops and
Diplocaulus, can be seen at the American Museum of Natural History, New York; Denver Museum of Nature and Science; Field Museum of Natural History, Chicago; Museum of Comparative Zoology, Harvard University, Cambridge, Massachusetts; National Museum of Natural History, Smithsonian Institution, Washington, D.C.; and Sam Noble Oklahoma Museum of Natural History, University of Oklahoma, Norman.
FOR FURTHER READING
Anderson, Jason S., Robert R. Reisz, Diane Scott, Nadia B. Fröbisch, and Stuart S. Sumida. “A Stem Batrachian from the Early Permian of Texas and the Origin of Frogs and Salamanders.” Nature, May 22, 2008, 515–518.
Bolt, John R. “Dissorophid Relationships and Ontogeny, and the Origin of the Lissamphibia.” Journal of Paleontology 51 (1977): 235–249.
Carroll, Robert. The Rise of Amphibians: 365 Million Years of Evolution. Baltimore: Johns Hopkins University Press, 2009.
Clack, Jennifer A. Gaining Ground: The Origin and Early Evolution of Tetrapods. Bloomington: Indiana University Press, 2002.