It is the mystery and beauty of organic form
that sets the problem for us.
—Ross Harrison, embryologist (1913)
THE AMAZING VARIETY of animal forms does not end with those on land or in the sea. Belowground, buried in as little as a few inches of sand or up to several thousand feet of rock, is the story of 600 million years of animal history—from the enigmatic forms of early animals in Canadian shale, to the enormous bodies of dinosaurs in the buttes and valleys of the American West, and the teeth and skull fragments of our bipedal ancestors in the Rift Valley of east Africa. And some of what lies below the ground can be quite surprising given what breathes just above.
I learned this firsthand only recently in, of all places, Florida, a favorite destination for vacationers and retirees seeking sun, entertainment, and relaxation. It is a land of palm trees, soft sandy beaches, graceful pelicans and ospreys, gentle manatees and dolphins, and Homo sapiens in plaid pants … but also of six-foot-long armadillos, tusked mastodons, sixty-foot-long sharks, camels, rhinoceroses, jaguars, and saber-toothed cats?
FIG. 1.1 Fossils from a Florida riverbed. Mammal bone, turtle shell fragments, and shark teeth abound. Note the variety of shapes and sizes. The largest tooth is from the enormous shark Charcharadon megalodon. FOSSILS COLLECTED AND PHOTOGRAPHED BY PATRICK CARROLL
Yes, indeed. Well, it depends on where you look.
Journey inland to a river cutting through the sandy soil and a shovel of gravel from the riverbed might contain teeth from any of ten species of shark, from the intricately serrated and curved snaggletooth, to the absolutely terrifying six-inch flesh rippers of the long extinct behemoth Charcharadon megalodon (figure 1.1). In the same gravel there will also be bones of Florida’s geologically recent past—of tapirs, sloths, camels, horses, glyptodonts, mastodons, dugongs, and other species now vanished.
The diversity of living and fossil animal forms in just this one locale frames the two central mysteries at hand: How are individual forms made? And, how have so many different forms evolved?
At first, the variety of animal forms may seem overwhelming. But there are some general, long established trends in animal design that we can make sense of. In this chapter, we’ll search for some generalities in animal architecture and evolution that will help us reduce this mind-boggling diversity to some basic themes.
A basic theme of animal design becomes obvious when one tries to figure out just what bone or tooth one has found in that shovelful of Florida river gravel. The challenge of the game is both to match the fossil to a species, and also to determine where in the animal it belonged. Why is this so hard? This is one demonstration of a basic fact of animal design. Related animals, such as vertebrates, are made up of very similar parts.
Now say, with a bit of expert help, one is able to figure out that a piece of bone is from a dugong (an extinct sea cow). But if it is a rib, which rib? Or, if a toe bone is from an extinct horse, which toe is it? From individual bones, it is really difficult to tell. Why this is the case punctuates a second basic fact of animal design—that individual animals are made up of numbers of the same kinds of parts, like building blocks.
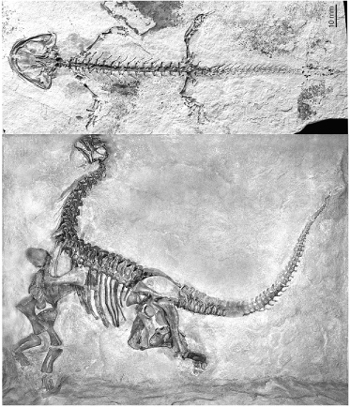
FIG. 1.2 The modular architecture of vertebrates. Top, a Jurassic salamander about 10 centimeters in length. Bottom, Camarasaurus, a Jurassic sauropod dinosaur, almost 19 feet in length. SALAMANDER COURTESY OF NEIL SHUBIN, UNIVERSITY OF CHICAGO; CAMARASAURUS COURTESY OF CARNEGIE MUSEUM OF NATURAL HISTORY, ALL RIGHTS RESERVED
Some of these parts can be small, such as an individual toe bone, others gigantic, like the backbones (vertebrae) of some vertebrates. These basic elements are ancient and their proportions maintained over vast differences in body size. Both enormous sauropod dinosaurs and small, delicate salamanders from the Jurassic age (over 150 million years ago) display the same repeating modular architecture of the vertebrate body plan (figure 1.2).
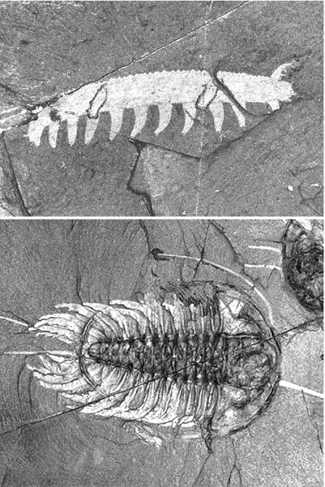
The theme of modular design is by no means limited to vertebrates. The famous fossils of the Burgess Shale, some of the first large, complex animals that populated the Cambrian seas more than 500 million years ago, display all sorts of variations on modular body plans (figure 1.3), as do their living descendants today.
The attraction to these fossils is manifold. Certainly, there is a sense of awe and wonder in seeing and touching extinct beasts that lived in worlds that have long since vanished. But we are also drawn to their form. The fossils demonstrate evolution’s pervasive use of repeating parts and modular architecture in forging animal designs.
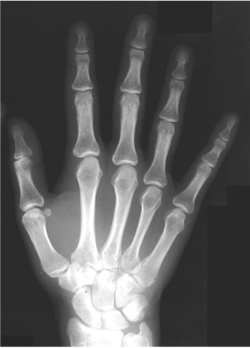
Individual body parts also reflect this theme of modular design. Our limbs, for example, are of similar modular design, each limb built of several pieces (thigh, calf, ankle; upper arm, forearm, wrist) and the hands and feet bear five similar digits (figure 1.4). The modular architecture of the limbs of four-legged vertebrates is also an ancient design, plainly evident in the Jurassic fossils.
Sometimes, the modular design of a structure may not at first be apparent. The complex patterns on a butterfly wing may appear chaotic, but on closer inspection one can see that the overall pattern is also built of repeating motifs. The underside of the blue Morpho butterfly has repeating patterns of lines, chevrons, and spots where each of the individual elements are separated by the wing veins (figure 1.5). This shows that each division of the wing enclosed by veins is a unit. The overall pattern is a product of repeating these modular units, each with some variations on the size or shape of the lines, chevrons, or spots within.
The repeating patterns on body structures extend down to very fine details, some almost out of range of the naked eye. Such beautiful butterfly wing patterns are actually built from tiny scales on the wing. Each scale is a projection of single cells that are assembled in many rows in the wing. Each scale has its own specific color, like the brushstrokes on a pointillist painting that when combined in a field of thousands and millions of scales, create the overall pattern we admire. The body patterns of fish, snakes, and lizards are also composed of scales (different from those of butterflies) arrayed in a regular geometric pattern. The reflective or refractive properties of scales depend on even finer cellular microanatomy that determines the wavelengths of light that are absorbed or reflected (figure 1.6).
FIG. 1.3 The modular architecture of Cambrian animals. The lobopodian Ayshaeia pedunculata (above) and trilobite Olenoides serratus (below) display repetitively organized, modular body forms. PHOTOS COURTESY OF CHIP CLARK, SMITHSONIAN INSTITUTION
FIG. 1.4 The modular design of a human hand. The finger bones revealed by an × ray display a serially reiterated architecture. COURTESY OF JAMIE CARROLL
FIG. 1.5 Serially repeating designs within butterfly wings are shown in the underside of this Morpho butterfly. Each wing is made up of serially reiterated subunits demarcated by two veins and the wing edge. Each subunit contains variations of the same elements—eyespots, bands, and chevrons. BUTTERFLY GIFT OF NIPAM PATEL, PHOTO BY JAMIE CARROLL
From just these few descriptions, we can begin to appreciate the immense task of development—to build large, complex animals beginning with only a tiny single cell. There are millions of details, and the details count. A small shift in an early process would have a cascade of later effects. What processes can assemble both a massive dinosaur and paint the delicate details of a spot on a butterfly?
Given such enormous differences in scale, and such great variety in animal forms, it would seem that the details of development would present what molecular biologist Gunther Stent described only twenty years ago as “a near infinitude of particulars which have to be sorted out case by case.” But biologists have been surprised and delighted to find there are generalities we can make about form and, fortunately, these generalities also extend far more deeply than outward appearances, into the genetic machinery of development. I’ll start with the outward similarities here and work my way down to the genes that do the job in the next two chapters.
FIG. 1.6 Repetition on a fine scale. The scales of a butterfly wing are like the strokes of a pointillist painting, each stroke being a single scale of a particular color; collectively they form geometric pattern elements. PHOTO BY STEVE PADDOCK
The modular and repetitive aspects of animal design reflect an order to animal forms. Anatomists have long appreciated that no matter how diverse their outward appearance, animal bodies and their parts are constructed along some perceivable themes. More than a century ago, some of these themes were formally defined by the English biologist William Bateson. His perspective turns out to be a very helpful framework for thinking about the logic of animal design and understanding how variations on basic themes evolve.
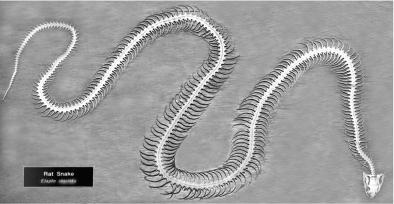
Bateson recognized that many large animals were constructed of repeated parts, and many body parts themselves were constructed of repeated units. In considering particular groups of animals, it appeared that some of the most obvious difference between members of a group were in the number and kind of repeated structures. For example, while all vertebrates have a modularly constructed backbone made up of individual vertebrae, different vertebrates possess different numbers and kinds of vertebrae. The number of vertebrae from head to tail differs greatly, from fewer than a dozen in frogs, to thirty-three in humans, and up to a few hundred in a snake (figure 1.7). There are also different kinds of vertebrae such as cervical (neck), thoracic, lumbar, sacral, and caudal (tail) vertebrae. The main differences between these types in any one animal are their size and shape and the presence or absence of structures attached to them, such as ribs. There is great diversity in the number of each type in different vertebrates.
A similar pattern applies to arthropod form and diversity. Arthropod bodies are made up of repeating segments, which in the trunk (behind the head) may vary from about eleven segments in insects to dozens in centipedes and millipedes. Groups of segments are distinguished from one another (e.g., the thoracic and abdominal segments) by their size and shape and especially by the appendages that project from them (e.g., the thoracic segments in insects each bear a pair of legs while the abdominal segments do not).
These two groups of animals have successfully exploited every environment on earth (water, land, and air) and are the most complex animals in terms of anatomy and behavior. Both groups are constructed of repeated assemblages of similar parts. Is there a connection between modularity of design and the success in evolutionary diversification? I certainly think so. The challenge for biologists has been to figure out how these animals are built, beginning from just a single cell, and how all sorts of variations on a body design evolve. The modular construction of vertebrates and arthropods, and their variation in the number and kinds of modules, are important clues to the processes involved.
FIG. 1.7 Snake skeleton. Hundreds of repeating vertebrae and ribs make up the snake body form. COURTESY OF DR. KURT SLADKY, UNIVERSITY OF WISCONSIN
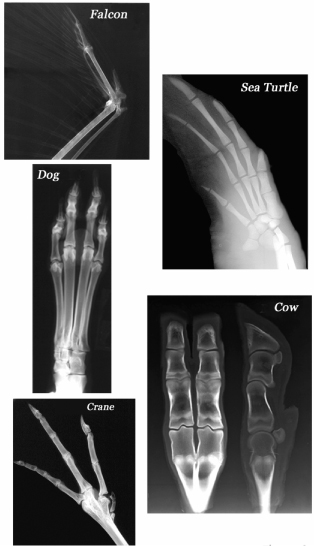
Body parts that are often modular and constructed of similar units often differ between species largely in number and kind. The limbs of four-legged vertebrates (tetrapods) usually bear one to five digits. We recognize five distinct types of digits on our own hands (thumb, forefinger, etc.) and feet. The similarities among digits are obvious, the differences largely a matter of size and shape. The tetrapod limb has been adapted to many functions in a great variety of designs, and the basic five-digit design has persisted for more than 350 million years, although digit number has evolved extensively such that anywhere from one to five digits may be present (for example, camels have two toes, rhinos have three, etc.). The variations on the tetrapod theme are spectacular, as a sample of × rays among vertebrates highlights (figure 1.8). Interestingly, closely related animals can differ widely; some groups have evolved many species that differ from one another in digit number.
FIG. 1.8 Diversity of vertebrate limb forms. All vertebrate limbs are variations on a common design in which the number, size, and shape of elements (such as digits) differ. COURTESY OF DR. KURT SLADKY, UNIVERSITY OF WISCONSIN; SEA TURTLE COURTESY OF DR. CRAIG HARMS, NORTH CAROLINA STATE UNIVERSITY
When comparing body parts between species, it is important to know whether one is comparing the same body part that might have changed in different ways, or one is comparing parts in a series, where the one-to-one relationship may be obscured. For example, the forelimbs of salamanders, sauropods, mice, and our arms are all homologs. This means they are the same structure modified in different ways in each species. They are derived from a common ancestral forelimb. Hindlimbs, our legs and the hind legs of four-legged vertebrates, are also homologs. With respect to each other, forelimbs and hindlimbs are serial homologs, structures that arose as a repeated series and have become differentiated to varying degrees in different animals. Vertebrae and their associated structures (ribs); tetrapod forelimbs and hindlimbs; digits; teeth; the mouthparts, antennae, and walking legs of arthropods; and the fore- and hindwings of insects are also serial homologs.
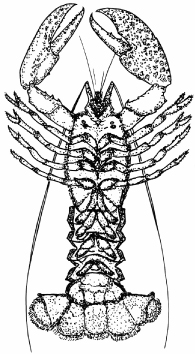
Changes in the number and kind of serial homologs are a principal theme in animal evolution. Let me drill this home with a couple of more examples of familiar structures. If you are a seafood lover, chances are you have dissected a lobster. While dismembering it, perhaps you might have noted the modular design and admired the great variety of body appendages (figure 1.9). There are several aspects to lobster construction that reflect the general themes of modularity and serial homology. First, the body is organized into a head (with the eyes and mouthparts), a thorax (with walking legs), and a long tail (yum!). Second, different sections of the body possess numbers of specific appendages (antennae, claws, walking legs, swimmerets). And third, each jointed appendage is itself segmented, and different kinds of appendages have different numbers of segments overall (compare a claw with a walking leg). If you were feeling adventuresome and dissected an insect or a crab, you’d see some general similarities in body organization, segmentation, and appendages but, again, differences in the number and kind of serially homologous structures.
FIG. 1.9 The diversity of the serially repeated appendages of a lobster. The antennae, claws, walking legs, swimmerets, and tail structures are all modifications of a common limb design. DRAWING BY JAMIE CARROLL
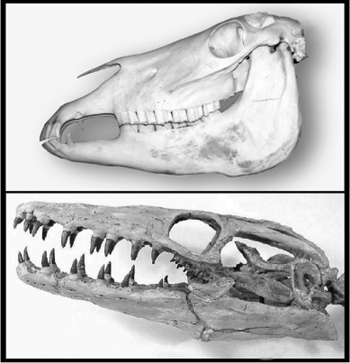
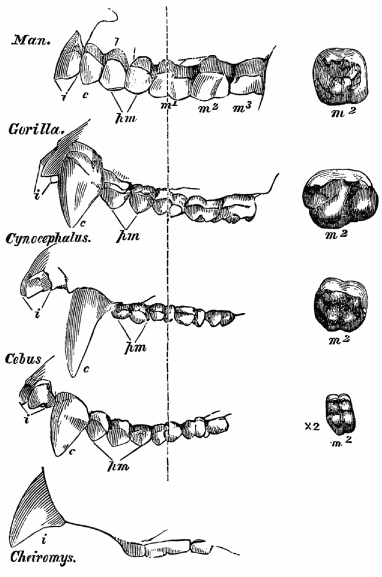
A second example of serially homologous parts would be the teeth you used to mince and crush that lobster. Our jaws host a variety of teeth (canines, premolars, incisors, molars, etc.). Again, one of the obvious differences among all sorts of vertebrates are the number and kind of teeth. Primitive reptiles, like great marine forms, had a mouth full of mostly similar teeth, but later species evolved different kinds of teeth, adapted for biting, tearing, and compacting food. The differences in dental hardware reflect differences in diet, with carnivores bearing incisors and canines and grazers bearing mostly molars (figure 1.10). We differ from our primate relatives in our dentition (figure 1.11). You may be aware that teeth make hardy fossils and such finds have played a major role in deciphering the identity and lifestyle of our ancestors.
The evolutionary trends in the number and kinds of repeated structures are so pronounced that the paleontologist Samuel Williston declared in 1914, “it is [also] a law in evolution that the parts in an organism tend toward reduction in number, with the fewer parts greatly specialized in function.” Williston was studying ancient marine reptiles. He noted that in the course of evolution, earlier groups tended to have large numbers of similar serially reiterated parts, but that later groups exhibited reduced numbers and specialized forms of these structures. Furthermore, the specialized pattern rarely reverted to the more general form. One interesting case is that when digits first evolved in tetrapods, there were as many as eight digits per foot. But among these eight, there were no more than five types, which eventually reduced to five digits that were specialized, or further reduced, in later species. Laws in biology are few, and those dared to be articulated are almost certain to be broken by some organisms. Yet Williston’s Law is a useful observation that seems to pertain to trends in more than just the ancient marine reptiles he was writing about. The trend appears to be that once expanded in number, serial homologs became specialized in function and reduced in number. The specialization of vertebral, tooth, and digit morphology in vertebrates, and of legs and wings in arthropods, was in fact generally accompanied by a reduction in the number of these repeated structures. Williston and Bateson appear to have captured some simple truths about animal design and evolution, allowing us to boil down the vast history and variety of some of the largest and most diverse groups into some generalities.
FIG. 1.10 Teeth in a primitive vertebrate. In mosasaurs (bottom), all teeth appear mostly similar, whereas later vertebrates (top; here a horse) had teeth of distinct types. RECONSTRUCTION OF PLATECARPUS PLAIFRONS COURTESY OF MIKE EVERHART, OCEANS OF KANSAS PALEONTOLOGY
FIG. 1.11 The diversity of primate dental hardware. Primates differ in the number and shape of canine, premolar, and molar teeth. FROM T. H. HUXLEY, MAN’S PLACE IN NATURE (1863)
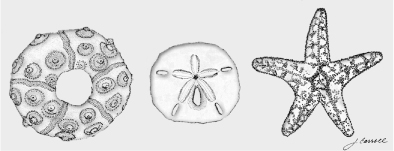
In addition to the repetition of modular parts, animal bodies and body parts usually display two additional features—symmetry and polarity. Most familiar animals are bilaterally symmetrical in that they have matching right and left sides with a central axis of symmetry running down the middle of the long axis of the body. This design also imposes a front/rear orientation to animals and has enabled the evolution of many efficient modes of locomotion. Some animals exhibit other symmetries, such as the pentaradial (five-fold) echinoderms, a group including sea urchins, sand dollars, and a spectacular variety of other species (figure 1.12). The axes of symmetry in an animal are clues to how the animal is built.
So, too, is the polarity of an animal and its parts. In most animals there are three axes of polarity: head to tail, top to bottom (back and front in ourselves, since we stand up), and near to far from the body (in reference to structures that project from the main body—such as a limb whose parts are organized perpendicular to the main body). Individual structures also have polarity. Think of the hand, which has three axes oriented by the thumb to pinkie, back to palm, and wrist to fingertip directions.
Modularity, symmetry, and polarity are nearly universal features of animal design, certainly of larger, more complex animals such as butterflies and zebras. These features and the evolutionary trends noted by Williston and Bateson suggest that there is order and logic to animal architectures. They suggest that underneath the great variety of animal forms, there are some general “rules” to be discovered about how animals are built and evolve.
FIG. 1.12 Other symmetrical animal forms. Echinoderms such as sea urchins (left), sand dollars (center), and starfish (right) are radially symmetrical. DRAWING BY JAMIE CARROLL
In the course of this book, I will focus on four main questions:
1. What are some of the major “rules” for generating animal form?
2. How is the species-specific information for building a particular animal encoded?
3. How does diversity evolve?
4. What explains large-scale trends in evolution, such as the change in number and function of repeated parts?
Where do we look for these rules and instructions? In DNA. In the entire complement of DNA of a species (the genome), there exists the information for building that animal. The instructions for making five fingers, or two eyespots, or six legs, or black and white stripes are somehow encoded in the genomes of the species that bear these traits. Does this mean there are genes for fingers, genes for spots, genes for stripes, etc.? I will focus on how anatomy is encoded in the genome in the first part of the book.
I’ll tackle evolutionary diversity in the second part of the book. Somehow, different species with three versus four fingers, two versus seven eyespots, six versus eight legs, or that are all black or all white, must have different instructions encoded in their DNA. Evolution of form is ultimately then a question of genetics. But in order to understand how genes sculpt all of this breathtaking animal beauty, we will first look to monsters for some critical clues.
Poster for the movie The Cyclops (1956). B & H PRODUCTIONS, INC. ALL RIGHTS RESERVED