“Do you know, I always thought Unicorns were fabulous monsters, too? I never saw one alive before!”
“Well, now that we have seen each other,” said the Unicorn, “if you’ll believe in me, I’ll believe in you. Isn’t that a bargain?”
—Lewis Carroll,
Through the Looking Glass (1872)
CREATURE FEATURE was a staple of Saturday afternoon television when I was a kid. My best friend, Dave, was addicted to the show. He’d hole up in his basement with the curtains drawn, lights off, a baseball bat at his side, and all sorts of contraptions rigged to the door and windows in case one of the featured monsters happened to visit him in the middle of the show. He’d watch hours of Godzilla, Dracula, the Mummy, or worse. Dave would later recount the plot lines for us and speculate on the relative strengths and unique powers of all the beasts. Fueled by an active imagination, which was probably also stoked by the five-gallon tin of popcorn and the can of Betty Crocker frosting that were his standard refreshments, these creatures had become almost real to him.
Our fascination with and terror of monsters are universal and ancient. From Greek mythology to B movies, writers have imagined all sorts of giants, hybrids, and ghoulish creatures. I didn’t share Dave’s appetite for monster movies (or cans of frosting, for that matter), but monsters have played an important role in advances in embryology. One of the most successful approaches to figuring out how animal forms develop correctly has been the study of dramatic monsters with the wrong number of parts, or parts in the wrong places. Some of these forms were man-made creations, some the product of accidents and injuries during gestation, and others the result of rare mutational events in nature. The insights gained from study of these kinds of monsters have recently converged to reveal specific mechanisms underlying the assembly of all animal bodies and body parts.
I never bought into tales of the dead coming alive, humans transformed into bats or flies, gigantic skyscraper-scaling gorillas, creatures that were half human, half horse/goat/snake/fish/whatever, fire breathers, or invisible bodies. I put them all into the same category of dark fairy tales. I did the same with monsters with one central eye, but here is where I might have been too hasty to dismiss a creature.
While I was vaguely familiar with the mythology of Cyclops, I did not know that animals with one central eye were actually well-known to science. In fact, at one time in Utah, 5 to 7 percent of newborn sheep were afflicted with cyclopia, a lethal malformation in which they bear a single central eye, lack most nasal and jaw structures, and have incompletely developed brain hemispheres (figure 2.1). The formal term is holoprosencephaly, meaning single forebrain, and the key defects are that the forebrain and eyes fail to become separated into two symmetrical structures.
FIG. 2.1 Cyclopic lamb. Caused by exposure of mother at critical period to cyclopamine, a toxin produced by the plant Veratrum californicum. PHOTO COURTESY OF DR. LYNNE JAMES, POISONOUS PLANT RESEARCH CENTER, LOGAN, UTAH
The high frequency of cyclopia in sheep was eventually found to be associated with the presence of a plant, the lily Veratrum californicum, in the pastures in which their mothers grazed. Ingestion of this plant during a period of gestation (around the fourteenth day) was the critical factor. It turns out that the plant produces a chemical called cyclopamine that has a teratogenic (from the Greek teras, meaning monster) effect on the developing embryo.
Cyclopamine is just one of many known teratogens. Many other chemicals have adverse affects on development of embryos. The drug thalidomide, originally developed to treat nausea during pregnancy, is probably the most notorious, having caused thousands of birth defects in humans in the late 1950s and early 1960s. While these molecules have been known for decades, there was no progress in understanding how they acted until the recent convergence of embryology with molecular biology. These advances have stemmed from more specific experiments, especially through the manipulation of embryos and genes.
Over the past century, biologists have used scalpels, needles, tweezers, and all sorts of tools to chop, strangle, burn, blend, spin, and prick embryos to try to uncover some rules for building an animal. Pioneers in embryology relied entirely on physical methods to move and remove cells and then observe what unfolded in the embryo. From these crude tortures, some dramatic monsters were produced whose striking features revealed a few central principles governing the organization of developing animals.
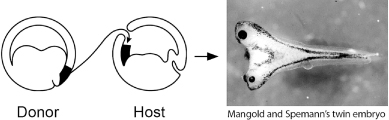
Foremost among these pioneers was Hans Spemann, the first, and for a span of more than sixty years, the only embryologist to win the Nobel Prize (but as we will see embryologists have lately been catching up). One of the very first illuminating experiments he carried out was to test whether the first two cells of a newt embryo had similar or different properties. Spemann used a fine baby’s hair, taken from his own daughter, to tie off and separate individual halves of the embryo. The cells on each side of the knot gave rise to normal newt tadpoles, demonstrating that the two halves of early amphibian embryos could give rise to two entirely identical animals.
When Spemann divided the egg differently by tying it perpendicular to the furrow between the two cells of the embryo, he obtained a dramatically different result. Only one side made a normal tadpole, while the other made a disorganized mess of belly tissue. This eventually led to the recognition that a region of the embryo, called the dorsal lip of the blastopore, was critical for the organization of the embryo. If this region of the embryo was removed, the embryo formed a blob of tissue lacking structures that normally form on the top (dorsal) side of the animal. More spectacularly, if this dorsal lip region was transplanted to the presumptive belly region of another developing embryo, it organized a second embryonic axis and two embryos formed that were joined together (figure 2.2)! Spemann dubbed this region “the organizer” because he deduced that it organized the dorsal part of the embryo into neural structures and could initiate the development of another embryonic axis.
The spectacular effects of the Spemann organizer revealed that one way order is brought about in development is by interactions between one part of an embryo and other parts. Some other organizers with dramatic properties have been discovered that show that this principle works on many scales in development: across the whole embryo, within individual body parts, and right down to intricate details of patterns. Let’s look at two more organizers that illustrate these dramatic activities.
FIG. 2.2 Induction of a second axis and embryo in a frog tadpole. Transplantation of “organizer” tissue to a different site induces the formation of a conjoined embryo. PHOTO COURTESY OF HIROKI KURODA AND EDDY DE ROBERTIS, UCLA
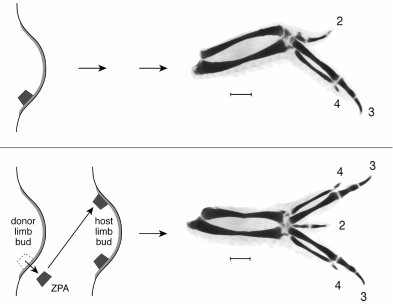
The formation of limbs has long held the fascination of embryologists. Beginning from just a small bud on the flank of the embryo early in development, the limb takes shape in many stages. In a three-day-old chicken embryo, the bud is initially only about 1 millimeter long and 1 millimeter wide but this will grow more than a thousand-fold by the time the chick hatches. In the intervening period, this tiny pad of tissue will grow outward, lengthen, and develop bone, cartilage, muscles, tendons, digits, and feathers in a beautiful display of the coordinated processes of development. Perhaps most striking is the orderly formation of cartilage elements (which will later be replaced with bone). The cartilage is formed around condensations of cells and laid down in order from the shoulder to the wrist and then to the digits. The whole process can be seen with special stains (figure 2.3). The order of events in limb development and the polarity of the digits suggests that, like the embryo, there must be some sort of system that cues cells as to what they will become.
Several decades ago, John Saunders, another pioneer embryologist, discovered an organizer of polarity in the chicken embryo wing bud. A chicken wing normally has three digits, which we can identify due to their size and shape as digits 2, 3, and 4 (in order from the front to the back of the wing; digits 1 and 5 do not form in the wing). When Saunders transplanted a chunk of tissue from the posterior part of the growing limb bud (near where digit 4 would originate) to an anterior position (where digit 2 would normally appear), a wing with extra digits formed. The digits were a mirror image duplication of the normal pattern—that is, instead of the 2 3 4 sequence of wing digits, a pattern of 4 3 2 3 4 formed (figure 2.4). The wing’s mirror image polarity suggested that cells in the posterior zone of the limb bud organized the polarity of the digit sequence (4, 3, 2), such that when moved elsewhere, the 4, 3, 2, sequence was induced in a new place.
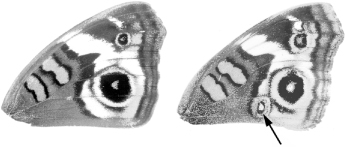
The realms of influence of Spemann’s organizer and the zone of polarizing activity (ZPA) in the chick limb are pretty large, affecting the development of the entire embryo or a large body part. But organizers have been found that act in finer scales. In 1980, Fred Nijhout of Duke University showed that the eyespot patterns on butterfly wings were also induced by an organizer. When Nijhout killed a tiny patch of cells that would form the center of the eyespot, no eyespot formed. More interesting, he found that when this small group of cells was isolated from the developing butterfly wing in the first day of the chrysalis stage and transplanted to a site elsewhere on the wing, a new eyespot now appeared (figure 2.5). Only cells at the future center of the eye-spot had this property. Nijhout dubbed the eyespot organizer the “focus.”
FIG. 2.3 Limb formation in a chicken. The wing and leg buds grow dramatically over several days of embryonic development. The laying down of cartilage, which precedes bone deposition, is visualized with a special stain and proceeds in order from upper limb parts to the digits. Note that the details of wing and leg anatomy differ in detail. PHOTOS COURTESY OF JOSEPH J. LANCMAN AND JOHN FALLON, DEPARTMENT OF ANATOMY, UNIVERSITY OF WISCONSIN
FIG. 2.4 Induction of polydactyly in a chicken. Transplantation of the zone of polarizing activity in the developing wing bud from a posterior site to a new anterior position induces extra digits with opposite polarity to the normal digit pattern. PHOTOS COURTESY OF JOSEPH J. LANCMAN AND JOHN FALLON, DEPARTMENT OF ANATOMY, UNIVERSITY OF WISCONSIN
All organizers share the property of influencing the formation of pattern, or morphogenesis, in tissues or cells. The basic interpretation of their special activity is that the cells of organizers produce substances that can influence the development of other cells. Such substances have been dubbed morphogens. The effects of organizers depend upon their distance from target cells: cells nearby are most affected whereas cells farther away in the newt embryo, limb bud, or butterfly wing are not (or less) affected. It has long been thought that morphogens produced in one site diffuse outward and form concentration gradients from their source. The idea then is that cells surrounding the source respond according to the amount of morphogen they experience. For example, in the chicken wing bud, cells close to the ZPA develop the posterior type of digit (digit 4), and those farther away progressively develop more anterior type of digits (digits 3 and 2 respectively). In the butterfly eyespot, the concentric rings of differently colored scales are thought to reflect different responses to different levels of the focal morphogen at different distances from the focal source.
FIG. 2.5 Induction of eyespots in a butterfly. Transplantation of cells at the center of a developing eyespot to other locations in the developing wing induces an eyespot in those locations. PHOTO COURTESY OF DR. H. FREDERIK NIJHOUT, FROM HIS THE DEVELOPMENT AND EVOLUTION OF BUTTERFLY WING PATTERNS, USED BY PERMISSION OF SMITHSONIAN INSTITUTION PRESS
The morphogens responsible for the activity of organizers were some of the most sought “Holy Grails” of embryology. The major difficulty that retarded further advances was that the organizer activity was a property of collections of cells. Any cell makes thousands of substances and it was always possible that more than one substance was responsible for organizer activity. While transplantation was a powerful tool, embryologists needed some way to find morphogens in the complex soup of cell biochemistry. They would wait decades.
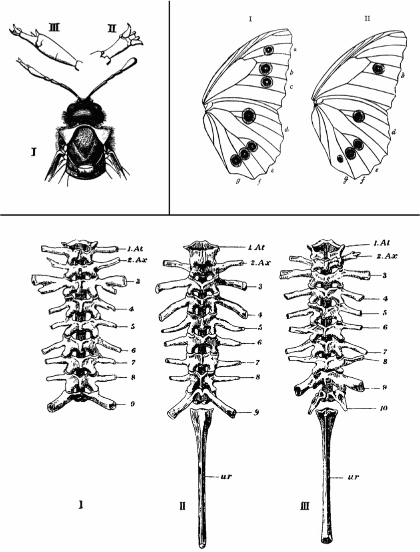
The animals created by Spemann, Saunders, and Nijhout were man-made monsters with duplicated axes or extra digits or altered wing spots. But these sorts of abnormalities were not unknown in nature. In fact, Bateson, in his 1894 treatise Material for the Study of Variation, catalogued and described a whole parade of “monsters” from across the animal kingdom that displayed extra, missing, or altered parts. Bateson culled from museums, collectors, and anatomy departments across Europe a menagerie of oddities including: a sawfly and a bumblebee with legs in place of their left antennae; crayfish with extra oviducts; butterflies with missing or extra eyespots; frogs with extra vertebrae or transformations of vertebral types; and much more (figure 2.6).
Bateson divided these abnormalities into two basic types: those in which the number of repeated parts was altered and those in which one body part was transformed into the likeness of another. He called the latter variants homeotic (from the Greek homeos meaning same or similar), and this will be a very important term to remember. Bateson’s motivation for collecting these oddities was to show that leaps in morphology can occur in nature and thus could be the basis of evolutionary change. I have to say right off the bat that, as intuitive and appealing as Bateson’s reasoning may first appear, biologists generally believe with very good reason that the notion of evolution making such large leaps in a single bound is very, very unlikely. The fact that such variations arise does not mean that these are founders of new types or species. Rather, from what we now know these monsters are almost certainly misfits that will be swept away by the power of natural selection with no chance of passing on their traits. This notion of “hopeful monsters” giving rise to new forms in one single bound has been very difficult to dispel, particularly in the popular scientific press (the BBC even produced a program with this title a few years ago, despite my pleas with the producer that it was a discredited idea). It is a seductive notion, but without any merit. In the course of this book we shall see that there is no support for, nor any need to invoke, hopeful monsters as agents of evolution.
Perhaps the most obvious limitation presented by Bateson’s catalog is that most examples presented defects in only one of a pair of structures. While provocative, these one-of-a-kind museum pieces were rare finds and their causes were unknown. It was important to understand, for example, whether such forms were heritable or might be the result of physical damage to the embryo while it was forming (and thus would not be inherited). It did turn out that Bateson’s kinds of monsters were informative, not so much for telling us about the true cause of evolution, but for insights into development that bear on evolution. As foretold in one of my favorite essays by the late Stephen Jay Gould, one that in fact influenced a change in direction early in my scientific training, Bateson’s monsters were “helpful” scientifically, but hopeless as individuals.
FIG. 2.6 Some of Bateson’s monsters. Upper left, a homeotic transformation in a sawfly where one antenna is transformed into a leg. Upper right, eyespots missing from a butterfly wing. Lower panels, variation in vertebrae and their projections in a frog. FROM W. BATESON, MATERIALS FOR THE STUDY OF VARIATION (1894)
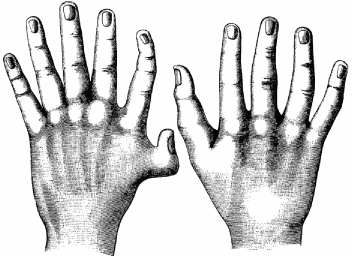
Bateson’s collection of variations also included cases of humans bearing extra ribs, men with one or a pair of extra nipples, a spectacular case of eight fingers arranged in mirror image symmetry on a left hand, and individuals with extra digits on one or both hands (figure 2.7). The latter conditions, termed polydactyly, are actually not that rare, occurring about 5 to 17 times per 10,000 live births.
There is quite a range of degrees of polydactyly, from the appearance of just an extra flap or bud of skin on the side of the pinkie or thumb to duplications of the nail, individual bones, or entire digits. Extra digits may be separate or fused to other digits; the latter condition is known as synpolydactyly. In some cases, the condition is fully bilateral on both hands and feet (figure 2.8).
Humans can fare quite well with extra digits. There are famous cases of polydactylism in history, including Anne Boleyn, wife of Henry VIII, who apparently had an extra nail on one hand. It is also reported that King Charles VIII of France and Winston Churchill may have had extra digits. Antonio Alfonseca, a relief pitcher for the 2003 baseball World Series Champion Florida Marlins, has six digits on both hands and feet. The extra finger does not affect his pitching grip, so it does not affect his success on the mound. It does appear, however, to offer some psychological advantage as opponents often refer to batting against Alfonseca as “facing the six-fingered man.”
FIG. 2.7 Polydactyly of a human hand. FROM W. BATESON, MATERIALS FOR THE STUDY OF VARIATION (1894)
Polydactyly is often heritable and pedigrees of polydactylous families are well-known. Indeed, it is reported that in a region of Turkey near Ephesus called Altiparmak, some families have taken the last name Altiparmak, meaning six-fingered ones.
Polydactyly is known widely throughout vertebrates, especially in cats, mice, and chickens. It is striking that similar digit patterns can occur in different animals, including humans, and they can be induced by experimental manipulations or inherited. This suggested that there could be some mechanisms in common for generating extra human fingers and chicken digits. But no progress was made into the mechanisms underlying digit number and pattern until advances were made in understanding some spectacular mutants in animals that have no fingers or toes—the humble fruit fly.
FIG. 2.8 Polydactyly on both hands and feet. This patient has six fingers on each hand and seven toes on each foot. PHOTOS COURTESY OF DR. ROBERT HILL, MRC HUMAN GENETICS UNIT, EDINBURGH, SCOTLAND; FROM PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCE, USA99 (2002): 7548
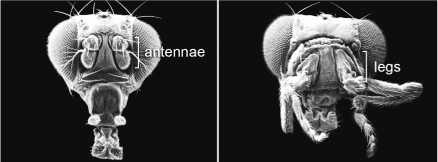
In order to make further progress into what monsters could teach us about the rules of development, a continual supply of abnormal types was needed, monsters that would breed true in the laboratory such that their offspring and subsequent generations would exhibit the same characteristics. In 1915, geneticist Calvin Bridges obtained the first true breeding homeotic mutant in the fruit fly Drosophila melanogaster, which was then just beginning to become a leading species for genetic investigations. Bridges isolated a spontaneous mutation that caused the tiny hindwings of the fruit fly to resemble the large forewings. He dubbed this mutant bithorax. Subsequently, several more homeotic mutants were identified in Drosophila. For example, a rather spectacular mutant Antennapedia causes the development of legs in place of the antennae on the head (figure 2.9).
It is remarkable how these homeotic mutants can so completely transform one structure into another. It is not that development is stunted or fails, but that the fate of an entire structure is altered, such that a part is put in the wrong place or the wrong number of parts form. Crucially, the transformation is of one serial homolog into the likeness of another (antenna to leg, hindwing to forewing). They are also so intriguing because each transformation is due to a mutation at a single gene. In Drosophila, only a small number of “homeotic” genes give homeotic forms when they are mutated, indicating that a small number of “master” genes govern the differentiation of serially homologous body parts in the fly.
FIG. 2.9 Homeotic mutant fruit fly. Left, a normal fly head with antennae; right, an Antennapedia mutant fly in which the antennae are transformed into legs. PHOTOS COURTESY OF DR. RUDY TURNER, INDIANA UNIVERSITY
The spectacular effects of homeotic mutants inspired what would become a revolution in embryology, and then another in evolutionary biology. But in order to appreciate their meaning and the insights they have to offer, we have to dig deeper to understand how these master genes work. How can one gene affect one whole structure and not another? What do the genes encode that have such large effects on animal bodies? Perhaps your first response is: “A fruit fly? Why should I get excited about a fruit fly?” The answers to all of these questions unfold from understanding more about DNA and how genes work, and along the way we’ll learn some surprising discoveries about the makeup of different animal genomes.
Whimsical representation of Monod’s famous quip. COURTESY OF DR. SIMON SILVER, UNIVERSITY OF ILLINOIS–CHICAGO