1 Effects of Regularities in the Presence of a Single Stimulus
After reading this chapter, you should be able to:
- Summarize the available functional knowledge about different effects of noncontingent stimulus presentations.
- Describe the mental process theories of Sokolov, Bradley, and the opponent-process theories of Solomon.
Introductory Task
Sometimes the repeated presentation of a stimulus can lead to changes in reactions to that stimulus. Can you think of three reactions or behaviors that can change as a result of repeated stimulus presentations? Also, think about possible explanations of those effects: how can repeated stimulus presentations result in such effects?
1.1 Functional Knowledge
In this section we provide an overview of what is currently known about the conditions under which regularities in the presence of a single stimulus (i.e., noncontingent stimulus presentations) lead to changes in behavior. Most of the research we discuss concerns a phenomenon known as habituation, which is one of several effects of noncontingent stimulus presentations. It differs from those other effects with regard to the nature of the change in the behavior (i.e., a decrease in the intensity of a response). For example, a loud bang will elicit a strong startle response the first time you hear it. The second time you hear the bang, the reaction will be less strong, the third time even less so, and so on. This effect (the decrease in intensity of the original reaction as a result of the repeated presentations of a stimulus) is called habituation (see http://www.youtube.com/watch?v=Kfu0FAAu-10). Note that habituation is an effect and not a mental process, so it is pointless and circular to say that the intensity of a reaction decreases because habituation occurs. Although this form of learning was first described thousands of years ago, a systematic study was carried out only in the middle of the last century (see Thompson, 2009, for a historical overview). After a period of lesser interest in this phenomenon, there has been an increase in interest in recent years, mostly outside of traditional learning psychology, and especially in neuroscience and biology (but see De Paepe, Williams, & Crombez, 2019, and Rankin et al., 2009, for recent reviews of behavioral research). Below, we provide a brief overview of what is known about the way effects of noncontingent stimulus presentations are moderated by the type of stimulus that is used (section 1.1.1), the type of behavior that is observed (section 1.1.2), the organism that is studied (section 1.1.3), the broader context in which the stimulus is presented (section 1.1.4), and the way the stimulus is presented (section 1.1.5). Taken together, this constitutes a summary of the available functional knowledge about the effects of noncontingent stimulus presentations on behavior.

Figure 1.1
A baby rat becoming acquainted with human contact.
1.1.1 The Nature of the Stimuli
Although habituation effects have been found with a wide variety of stimuli, these effects occur more quickly for some stimuli than for others. Consider studies on the habituation of skin conductance responses. In response to the first presentation of a particular image (e.g., the picture of a cute puppy), there is a slight increase in the extent to which electricity is conducted across the skin (as measured by, for instance, electrodes attached to the hand of a person). This increase is referred to as the skin conductance response. When that image is presented a second time it will again result in a skin conductance response, but the magnitude of the response is smaller, and even smaller the third time, and so on (see figure 1.2). Interestingly, the rate of habituation (i.e., the extent to which the skin conductance response weakens as the result of the repeated presentation of the image) depends on the content of the image. Specifically, habituation of skin conductance is slower for affective (positive and negative) images (e.g., a picture of a cute puppy or a snarling dog) than for neutral images (e.g., a picture of a table; see Bradley, 2009, for an overview).
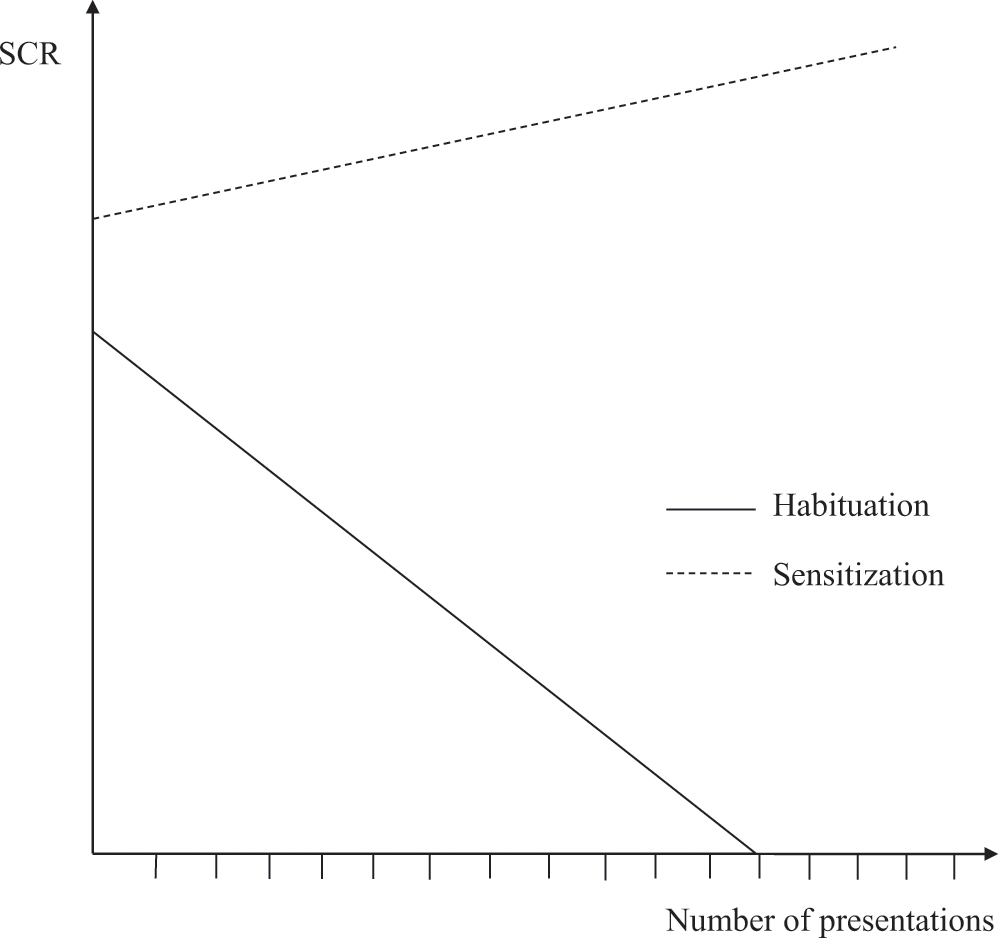
A fictitious example of habituation (a decrease in the strength of skin conductance responses as the number of stimulus presentations increases) and sensitization (an increase in the strength of skin conductance responses as the number of stimulus presentations increases).
The biological relevance of stimuli also seems to be important. Evans and Hammond (1983a, 1983b) found that rats habituated as quickly to the distress call of a conspecific (arguably, a biological significant stimulus) as to a control stimulus. However, habituation of the response to the distress call was less long-lasting and more context-specific than habituation of the response to the control stimulus. This effect of the biological significance of stimuli on habituation could have important evolutionary benefits.
With certain very important or intense stimuli (e.g., a painful shock or loud noise), sensitization can occur. Sensitization is an effect in the opposite direction from habituation; that is, it is an increase in the intensity of a reaction as a result of repeated stimulus presentations (see figure 1.2). Habituation and sensitization are therefore two possible effects of noncontingent stimulus presentations. As a real-life example, consider people who move to a house near a busy road. Whereas some people get used to the noise (i.e., habituation), other people become more and more annoyed by the sound of cars (i.e., sensitization). Whether sensitization or habituation occurs depends on various factors (see Groves & Thompson, 1970, for a review). Influential theories of addiction attribute a very important role to sensitization (e.g., Robinson & Berridge, 1993, 2008). For example, it has been established in nonhuman animals that the repeated administration of drugs leads to an intensification of certain motor reactions to the drug. This finding led to the idea that repeated use of a drug results in an increase in the urge to use the drug (more wanting), even if you experience less pleasure in using the drug (less liking; but see Tibboel, De Houwer, & Van Bockstaele, 2015, for a critical discussion of the concepts “wanting” and “liking”).
Finally, we note that the effects of noncontingent stimulus presentations are not limited to the stimulus that is presented in a noncontingent way. Habituation responses can be generalized from one stimulus to another: if one repeatedly presents a certain stimulus (e.g., a loud tone with a frequency of 1000 Hz), the reaction to other stimuli will also be influenced if these other stimuli resemble the stimulus that was repeatedly presented (e.g., a loud tone with a frequency of 900 Hz; Rankin et al., 2009).
1.1.2 The Nature of the Observed Behavior
Noncontingent stimulus presentations can have an influence on different aspects of different types of behavior, ranging from very simple to very complex behaviors. In the nineteenth century, Fechner (1876) wondered how the aesthetic appreciation of certain stimuli (e.g., paintings, musical works) changes when they are repeatedly presented. He formulated the following law: due to constant repetition, what is originally considered pleasant will first become more pleasant but ultimately unpleasant. This constitutes the first reference to what later became known as the mere exposure effect, which typically refers to the observation that novel, neutral stimuli become positive as a result of the repeated presentation of that stimulus (Moreland & Topolinski, 2010; Zajonc, 1968). Also in the nineteenth century, Peckham and Peckham (1887) published an article titled “Some Observations on the Mental Powers of Spiders.” It describes how the researchers repeatedly presented the sound of a tuning fork next to the web of a spider. The first time, this triggered a pronounced response from the spider, which moved on its web. This response diminished with each additional presentation. These old studies not only illustrate the range of behaviors that can be influenced by noncontingent stimulus presentations, they also show that interest in this form of learning has long existed (see Thompson, 2009, for a historical overview).
Finally, the effects of noncontingent stimulus presentations are not limited to the initial reactions elicited by these stimuli; they also involve reactions that come about only after those presentations. For example, a change in behavior due to a regularity in the presence of two stimuli (classical conditioning; e.g., the impact of bell-food pairings on salivation) can be reduced by first presenting one of these two stimuli by itself (e.g., repeatedly presenting the bell on its own before the bell-food pairings; see section 2.2.2).
Although many types of behaviors can be influenced by noncontingent stimulus presentations, laboratory research often focuses on changes in one type of response: the orientation response (OR). Interestingly, it was Pavlov (1927) who described this response, when he noticed how difficult it was to demonstrate classical conditioning to guests visiting his laboratory. After all, the dogs oriented their attention to those new people instead of the sounds that accompanied the food. The OR is now considered to consist of a set of different reactions that all seem to be aimed at the investigation of new and potentially important stimuli in the environment (which is why the OR is sometimes also called the investigatory reaction or what-is-it response). These components include orienting the head toward the stimulus, an increase in skin conductance, a decrease in heart rate, and neuronal responses such as certain changes in electrical activity in the brain (as measured by an electroencephalogram, or EEG). Importantly, research has shown that repeated stimulus presentations can have different effects on different components of the OR. For example, the decrease in heart rate seems to be the effect that most quickly disappears as a result of the repeated presentation of the trigger stimulus. On the other hand, there is almost no habituation of certain changes in EEG (see Barry, 2006, and Bradley, 2009, for an overview). Hence, there is growing doubt about the usefulness of viewing the OR as one unitary response (see Barry, 2009).
Habituation studies also reveal intriguing interactions between the nature of the stimulus and the nature of the response (Bradley, 2009). As we mentioned earlier, habituation of the skin conductance response is faster for neutral images than for affective images. However, habituation of changes in heart rate is as fast for neutral as for affective images. Such findings suggest that the different components of the OR are determined by different aspects of the environment.
Finally, the effect of repeated stimulus presentations can be opposite for reactions compared to counterreactions. Certain stimuli (e.g., drugs) elicit not only a reaction (e.g., a “high”) but also a counterreaction (e.g., a hangover). This also applies to certain activities (e.g., jogging can lead to discomfort initially, and later to a buzz). This sequence of observable reactions and counterreactions is called the dynamics of affect because it refers to the dynamic development of affective reactions over time. However, research has shown that the dynamic changes as the result of repeatedly presenting a stimulus. The initial reaction to a stimulus will become increasingly weaker, while the counterreaction will become increasingly stronger as a result of the repeated presentations of a stimulus. For example, a drug user will feel less of a “high” the more often she takes a certain fixed amount of the drug. She will therefore have to use increasingly larger quantities to reach the same state of high. The counterreaction (withdrawal symptoms) when using a certain amount of the drug, however, becomes stronger with repeated use. We see something similar in activities where the reaction is unpleasant and the counterreaction pleasant. Take the example of jogging. Initially, one has strong negative experiences during jogging and few positive experiences afterward. However, as people jog more often, the negative reactions during jogging become weaker and the positive reactions after jogging become stronger. Some seasoned joggers seem addicted to the buzz they experience after jogging. More generally, it seems that repeated presentations have opposite effects on reactions and counterreactions: reactions become weaker and counterreactions become stronger with the repeated experience of stimuli. In section 1.2.2 we discuss the opponent-process theory of Solomon, which provides an explanation for this intriguing phenomenon.
1.1.3 Properties of the Organism
Effects of noncontingent stimulus presentations have been demonstrated in humans (see Bonetti & Turatto, 2019, for a recent example), nonhuman animals (e.g., spiders), and even single-celled organisms and plants (box 1.2). Nevertheless, certain characteristics of the organism do moderate the effects of noncontingent stimulus presentations; that is, they determine whether and how those presentations influence behavior. The fact that biological significance moderates habituation (see section 1.1.1) shows that the type of organism involved is important: whether a stimulus is biologically significant (and thus how quickly habituation takes place for that stimulus) varies from species to species (e.g., distress calls by rats are relevant mainly to rats).
1.1.4 The Impact of the Broader Context
There is very little research on the influence of the broader context on the effects of noncontingent stimulus presentations. One such study was carried out by Iacono and Lykken (1983). They repeatedly presented a loud tone (110 dB) and examined the extent to which this led to a decrease in the skin conductance response that was triggered by the tone. Importantly, they also manipulated the instructions given to the subjects. Certain subjects were instructed to ignore the tones as much as possible. Others were asked to count the tones and see if each tone was the same. Habituation was more pronounced in the first group. The focus away from stimuli thus seems to accelerate habituation.
Box 1.1 Habituation as a Research Tool
Both nonhuman animals and young human babies cannot say what they see, hear, and feel. Yet we can form an idea of their subjective experience by using habituation as a research tool. After all, if habituation is different for different stimuli, then it seems likely that the organism can perceive the difference between these two stimuli. For example, a baby will look at figure 1.3B for a longer period than figure 1.3A, and the habituation of this fixation time will occur more quickly for figure 1.3A than for figure 1.3B (see Colombo & Mitchell, 2009, for an overview of research into habituation in children).

An example of stimuli used with babies in order to study habituation of the fixation time (based on the description provided by Bashinski, Werner, & Rudy, 1985, p. 585).
Recently, habituation of neuronal responses (as measured by fMRI) has been used to investigate the functioning of brain parts. For example, imagine that a drawing of a bicycle is presented repeatedly. During a subsequent test phase, the presentation of that drawing is followed by the presentation of either the same drawing or a drawing that differs in a single respect (e.g., the bicycle is slightly larger or a different color). If in a certain part of the brain there is a different reaction to the altered image than to the identical image, this would indicate that this part of the brain is involved in the processing of the deviating characteristic (e.g., size or color; see Kumaran & Maguire, 2009, for an overview).
However, many questions remain unanswered about this class of moderators. For example, researchers have not yet investigated whether habituation is also influenced by the presence of other tasks. We know that the original response to a stimulus will usually decrease in intensity if the stimulus is repeatedly presented. But what happens if test subjects have to perform another task while the stimuli are presented (e.g., count backwards from one thousand)? Does habituation become stronger or weaker? Further research is needed to answer this and other such questions.
1.1.5 Characteristics of Noncontingent Stimulus Presentations
When one repeatedly presents a stimulus, different aspects of those presentations can be varied. In what follows we discuss those aspects of the presentations that have an influence on habituation (a decrease in the intensity of the reaction as a result of repeated stimulus presentations) or sensitization (an increase in the intensity of the reaction as a result of repeated stimulus presentations; see Rankin et al., 2009, for a more complete overview).
1.1.5.1 The nature of the noncontingent stimulus presentation What appears to be particularly important is the frequency with which the stimulus is presented. With each presentation of the stimulus, the intensity of the reaction gradually decreases to a certain minimal level. It is therefore not the case that only the first presentation has an effect on behavior.
1.1.5.2 Changes in the nature of the noncontingent stimulus presentations The effect of repeatedly presenting a stimulus (e.g., habituation) can be nullified by presenting another stimulus unexpectedly. This nullification of the habituation effect is called dishabituation (see http://www.youtube.com/watch?v=4x-2WoyXPSM; see also box 1.2). Suppose that an intense light is repeatedly presented. The response to the light will decrease across repeated presentations of the light (i.e., habituation as an effect). If, afterward, a loud bang is unexpectedly presented, and later the light is presented again during a test phase, then the reaction to the light will be more intense than the reaction to the previous presentation of the light. Dishabituation is an important phenomenon because it shows that the original decrease in the intensity of the reaction is not due to a change in the behavioral repertoire (i.e., the collection of all responses that an organism can make at a certain point in time; see box 1.2).
Box 1.2 Habituation in Single-Celled Organisms and Plants
It is often thought that only organisms with a central nervous system are able to learn (but see Burgos, 2018, for a conceptual analysis, and Adelman, 2018, for a review of the available evidence). This misconception is reinforced by a definition of learning as a mental process responsible for storing information (because it seems natural to assume that living organisms need brains to store information), and even more so by the idea that learning is accomplished through the formation of connections between neurons in the brain (because by definition, you would need a system of neurons to learn). One important advantage of our functional definition of learning is that it does not make any a priori assumption about mediating mechanisms; it requires only an organism whose behavior might change as the result of regularities in the environment. Hence, our definition of learning is perfectly compatible with the idea that organisms without a central nervous system can learn, too. It is therefore interesting to see more and more evidence for learning in organisms that do not have a central nervous system (such as plants) and even organisms that consist of only one cell.
Take the example of habituation in slime mould, a creature that consists of just one enormous cell (Boisseau, Vogel, & Dussutour, 2016; see also https://www.youtube.com/watch?v=2UxGrde1NDA for an interesting TED talk about this organism). Slime mould can move via pulsating movements, and in this way it actively searches for food in its surrounding environment. In recent studies on the habituation of slime mould, researchers have made use of the finding that mould does not like to move across quinine (a bitter but harmless substance). In figure 1.4 the right panel (EXP) shows that the mould initially does not move over the quinine, but after repeated exposure to the quinine (images with the letter Q) it eventually moves over the quinine. This is an example of habituation: an original avoidance reaction to quinine (hesitation to move over this substance) disappears after repeated exposure to that substance.
Habituation also has been observed in plants. Consider Mimosa pudica, the “shy” plant. This particular plant closes its leaves whenever they are stimulated (see figure 1.5). In a study by Gagliano, Renton, Depczynski, and Mancuso (2014), this reaction was repeatedly elicited by dropping the plant (which is in a pot) from a certain height. After a while, the response disappeared: the plant did not close its leaves after it had fallen.
The fact that habituation can be found in single-celled creatures and plants shows that learning is not always dependent on a central nervous system. Partly on the basis of these findings, it has become clear that learning is not always due to the formation of connections between neurons; it can also be achieved by chemical processes within one cell, including changes in genetic material (i.e., genetic learning; see box 0.1). Even a single phenomenon such a habituation could in principle be mediated by different mechanisms in different species or even within one species.
These examples of habituation in single-celled creatures and plants allow us to further clarify the definition of learning in general and habituation in particular. For example, it is possible that the disappearance of the leaf-folding response of the Mimosa plant is a consequence of exhaustion. In other words, it is quite possible that the plant has only enough energy to close its leaves two or three times.

Figure 1.4
Habituation in slime mould (retrieved from Boisseau et al., 2016; figure 5)


Figure 1.5
Leaf-folding response of the Mimosa plant. The image above shows the leafs when open; the opposite image shows the leafs when closed. Transition from the open to closed state occurs when the leaves are stimulated, for instance by touching or shaking them.
In that case, it seems pointless to say that the plant has learned something when we find that it no longer closes its leaves on the fourth presentation of a stimulus. The plant simply is no longer able to perform the behavior, and at that moment, the closing of the leaves no longer belongs to the behavioral repertoire of the plant (i.e., the collection of all behaviors that the plant can make). Imagine, for example, that someone repeatedly eats poisonous food and then dies. There can be no doubt that the behavior of the person changes when he or she dies, but this can hardly be called learning because someone who is dead is no longer able to emit any behavior; the repertoire of possible behaviors has been reduced to zero. A distinction can therefore be made between learning on the one hand and changes in the behavioral repertoire on the other hand. More specifically, changes in behavior that are due to changes in the behavioral repertoire could be excluded from the realm of learning.1
Fortunately, it is relatively easy to exclude the possibility that there is a change in the behavioral repertoire: one simply has to demonstrate that the organism is still capable of performing the old behavior. Let us return to the example of habituation in the Mimosa plant. After the leaf-folding response has disappeared as a result of the plant repeatedly falling, you can elicit the same response by stimulating the plant in another way—for example, by shaking the plant back and forth (Gagliano et al., 2014). This shows that the leaf-folding response is still part of the plant’s behavioral repertoire and that the disappearance of this response is thus a real change in behavior due to the repeated stimulation of the plant. In other words, by showing that habituation does not generalize to other stimuli (e.g., the shaking of the plant), one can conclude with more certainty that habituation has occurred. Changes in the behavioral repertoire can also be excluded by demonstrating that dishabituation occurs. Dishabituation refers to the finding that a habituated response to a first stimulus is restored after a second stimulus is presented. Take our previous example of habituation in the slime mould. On the right-hand side of figure 1.4 you see that after a successful habituation of the avoidance response during the first six days, a neutral stimulus (A) is presented on the seventh and eighth days. This stimulus, too, is not avoided. On Day 9 the quinine is presented again and a renewal of the avoidance response occurs. This restoration of the original response as a result of presenting a different stimulus is an example of dishabituation (see section 1.1.5.2). Based on this observation, we can conclude that the avoidance response still belongs to the behavioral repertoire of the slime mould, which supports the hypothesis that the disappearance of the avoidance response on Days 1–6 is an example of habituation.
In short, research into habituation in plants and single-celled beings is interesting not only because it shows that habituation (and therefore learning) is a very general phenomenon that occurs in all kinds of organisms, but also because it provides new insights into the biological basis of learning and allows us to further refine the definition of learning.
Epstein, Temple, Roemmich, and Bouton (2009) developed a very nice analysis of eating behavior in terms of habituation and dishabituation. You can see the eating of a certain food (e.g., potatoes) as repeated presentations of that food, where every bite is a new presentation. As a result of these repeated presentations, the pleasure elicited by the food decreases (habituation) and you stop eating after a while. However, if you take a bite of another food (e.g., white cabbage) and then a bite of the first food (potatoes), then the first food will once again elicit more pleasure and satisfaction (dishabituation). This analysis provides a functional explanation for why people eat varied meals with many different types of food. It also explains why there is always room for a dessert, because the dessert is a completely different type of food than the starter and main course. There will therefore be little generalization of the habituation: the habituation that has occurred in relation to the food you have eaten during the starter or main course will have little or no effect on the pleasure that the eating of the dessert elicits. You eat more of a dessert than a kind of food that is similar to the starter or main course. Finally, it has also been established that there is a relation between habituation to food and obesity. The speed of habituation to food in the lab was a predictor of an increase in body weight in the following year (Epstein, Robinson, Roemmich, & Marusewski, 2011).
1.1.5.3 Temporal aspects of the noncontingent stimulus presentation Temporal aspects of the stimulus presentation seem to strongly influence the course of the habituation. The stimulus can be repeatedly presented in short succession (short interstimulus interval) or repeatedly with longer intervals. In the first case, one speaks of massed practice; in the second case, of distributed practice. In case of massed practice, habituation is faster (i.e., fewer presentations are needed to eliminate the response) but is less sustainable (i.e., the reaction will quickly reemerge as the result of the mere passage of time), whereas with distributed practice, habituation is relatively slower but more sustainable (see Rankin et al., 2009).
1.2 Process Theories
There is no overarching theory that provides an adequate explanation for all effects of noncontingent stimulus presentations. That is why the following section is limited to a discussion of two sets of theories, each of which attempts to explain a subset of effects of noncontingent stimulus presentations. First, we discuss the theories of Sokolov (1975) and Bradley (2009) on habituation of the orientation reflex (OR). Then we discuss Solomon’s theory about the effect of repeated stimulus presentations on reactions and counterreactions (i.e., on the dynamics of affect).
1.2.1 The Model of Sokolov and the Model of Bradley
The model of Sokolov (1960, 1963, 1975) is concerned with the habituation of the OR. Following numerous tests, Sokolov came to formulate a kind of discrepancy model. The basic idea in his model is that each organism constantly builds up a model of the environment in which it is located. When a stimulus is subsequently administered that is not part of this model, an OR is triggered, and at the same time, this new stimulus is included in the model. After repeated presentations of the same stimulus in the same context, there is no longer a discrepancy between the input and the stimulus representation, and as a result, the OR mechanism will be inhibited (see figure 1.6).

The model of Sokolov. The input (1) activates the OR mechanism (by which an OR is elicited) and is gradually included in the model of the environment. As the difference between the environment and the model becomes smaller, the inhibitory activity is triggered via (2), which blocks the impact of the stimulus on the OR mechanism. If, after habituation, the stimulus is no longer presented, then the OR mechanism is triggered by the difference between the actual environment and the model of the environment (3), as a result of which the absence of the stimulus will elicit an OR.
However, as soon as there is any change in the stimulus, the OR will be retriggered. In other words, as soon as something changes, this is immediately recorded. From this perspective, it is also understandable that an OR can also occur when an expected stimulus does not occur. An everyday example is the ticking of a clock. Over time, the ticking of a clock is not consciously attended to, but one usually becomes alert as soon as the ticking stops.
Although Sokolov’s theory was groundbreaking, it also had shortcomings. First of all, doubts have arisen about the finding that the unexpected absence of a stimulus can elicit an OR. At best, this phenomenon seems to be limited to certain reactions such as skin conductance (see Thompson, 2009, for a historical overview). Because this finding is the raison d’être of the Sokolov model (it is a crucial finding that is generally regarded as the origin of the idea that organisms build a model of the environment; Barry, 2009), these doubts have led to doubts about the model itself.
Box 1.3 Sokolov and Predictive Coding Models
The Sokolov model can be considered a precursor to predictive coding models that are currently very popular in cognitive psychology and neuroscience (e.g., Clark, 2013; Friston, 2009). The idea of predictive coding is also based on the assumption that organisms build a model of their environment and, on the basis of this model, make predictions about what should happen in the world. When there is a mismatch between a prediction and the actual situation in the environment, this will lead to an adjustment of the model. Predictive coding models have further crystallized this basic idea and applied it to various aspects of perception, thinking, and behavior (e.g., Clark, 2013). However, it is remarkable to see that in the extensive literature on predictive coding, there is seldom or never a reference to the work of Sokolov (see Bridgeman, 2013, for an exception). This is a good example of how ideas in science are sometimes reinvented without making the link with relevant earlier work. It is not surprising that the same idea can arise at different places or moments in time, but it is regrettable that often there is too little effort made to check whether similar ideas have already been formulated and, above all, what can be learned from earlier research. After all, one can learn a lot from both the successes and the problems of previous research. We therefore endorse Pavlov’s statement, “If you want new ideas, read old books.”2
Furthermore, it has become clear that the heuristic value of the model is limited. For example, based on Sokolov’s theory, one cannot explain why habituation is influenced by the nature of the stimuli or the observed behavior. For example, we mentioned earlier that habituation of the skin conductance response is faster for neutral stimuli than for affective stimuli. Habituation of heart rate deceleration (i.e., the slowing down of the heartbeat upon presentation of a salient stimulus), on the other hand, appears to be as fast for affective as for neutral stimuli. Bradley (2009) has proposed an alternative theory for such findings (see also Bernstein, 1981; Gati & Ben-Shakhar, 1990). It is based on the assumption that every stimulus that is motivationally relevant will elicit an OR. New, unexpected stimuli are motivationally relevant because new stimuli can indicate a danger or an opportunity. Other stimuli can be motivationally relevant because people learn that they are important for certain goals—biological goals such as survival and reproduction as well as cognitive goals such as tasks that people have in a given context. In other words, stimuli can elicit an OR, either on the basis of the extent to which they are new (novelty) or on the basis of their motivational meaning (significance; Gati & Ben-Shakhar, 1990). New neutral stimuli will therefore initially trigger an OR weaker than that triggered by new affective stimuli. The neutral stimuli elicit an OR only on the basis of their novelty, whereas affective stimuli elicit an OR on the basis of both novelty and significance. Repeatedly presenting a stimulus very quickly makes the novelty disappear, but only very slowly changes the meaning (and therefore significance) of a stimulus. As a result, the repeated presentation of a neutral stimulus will lead to the rapid disappearance of the OR in relation to that stimulus (because the OR is elicited on the basis of novelty only, and novelty disappears very quickly), whereas the repeated presentation of affective stimuli leads to the slow disappearance of the OR (because the OR is elicited by novelty and significance, and novelty quickly disappears but significance disappears very slowly). Bradley’s model can therefore better explain the influence of the nature of the stimuli on the habituation of the OR by assuming that different elements of a stimulus can lead to an OR and that these different elements are influenced in a different way by repeatedly presenting a stimulus.
Bradley (2009) also accounts for the fact that the nature of the observed behavior has an influence on habituation, by assuming that different components of the OR are determined to varying degrees by novelty and significance. The delay in heart rate is determined mainly by novelty, whereas skin conductance is determined mainly by significance. Through repeated stimulus presentations, the novelty decreases and therefore the strength of those OR components that are determined by novelty also decreases. Habituation therefore quickly occurs for those components. However, the repeated presentation of a stimulus adds little to the significance of the stimulus and thus also little to those components of the OR that are determined by it. Habituation is therefore slower for those components (see also Barry, 2006).
The interaction between the influence of the nature of the stimulus and the influence of the nature of the observed behavior is explained as follows: habituation of a component of the OR that is determined by novelty only (e.g., the delay of heartbeat) will occur equally quickly for neutral stimuli and affective stimuli. After all, only novelty counts for such components, and both types of stimuli are initially new or unexpected. Habituation of components that are determined also by significance, however, will show a slower habituation for affective stimuli than for neutral stimuli, because the significance of the affective stimuli disappears slowly. The mental process theory of Bradley (2009) is therefore able to explain more functional knowledge than the original theory of Sokolov (1975) can, and in that sense is a better theory. It does this by making additional assumptions about mental processes, namely (1) that the OR is determined not only by the novelty but also by the significance of a stimulus (where novelty and significance are hypothetical mental constructs), (2) that repeatedly presenting a stimulus has a different effect on novelty than on significance, and (3) that different components of the OR are influenced differently by novelty and significance.
1.2.2 The Opponent-Process Model of Solomon
Whereas the theories of Sokolov and Bradley try to explain habituation of the OR to all possible stimuli, Solomon (Solomon & Corbit, 1973, 1974; Solomon, 1980) formulated an opponent-process theory that is aimed specifically at habituation of reactions and sensitization of counterreactions (i.e., on changes in the dynamics of affect). Examples from daily life (e.g., use of drugs, jogging) suggest that repeatedly presenting a stimulus results in the weakening of the reaction and the strengthening of the counterreaction. Solomon studied the dynamics of affect in the laboratory by repeatedly administering electric shocks to dogs. He observed the heartbeat frequency of the dogs during the presentation of the shock (the reaction) and after stopping the shock (the counterreaction). Figure 1.7 shows what happens when the shock (lasting ten seconds) is administered (left figure) and when the shock ceases (right figure).

Heart rate during (left) and after (right) shocking dogs that had never experienced a shock before, measured in milliamperes, where 8 ma is more intense than 4 ma (retrieved from Solomon, 1980, figure 1).
The left graph shows a clear acceleration of the heartbeat, which is greater for more intense shocks. Note that while the shock is still being delivered, a slight decrease in heart rate is already visible. The right graph shows what happens immediately after the shock has stopped. Heart rate does not immediately return to the baseline (the heart rate level before the shock was presented); there is a clear delay before returning to the baseline.
These observations of the dynamics of affect are based on the first administration of such shocks. But what is the effect of repeated electric shocks on the dynamics of affect (i.e., on the occurrence of the reaction and counterreaction)? The answer can be found in figure 1.8. The results come from what Solomon calls veteran laboratory dogs (i.e., dogs that have often been exposed to electric shocks). On the left, we see the heartbeat during the shock—there is no acceleration at all (compare with the left side of figure 1.7). On the right is the heartbeat after the shock. We see that the delay in heart rate is much more pronounced than after the first administration (compare with the right side of figure 1.7).

Heart rate during (left) and after (right) the shock in dogs that have already been exposed many times (retrieved from Solomon, 1980, figure 3).
According to Solomon, these findings reflect a fundamental principle with regard to the reactions and counterreactions of humans and animals to emotionally charged stimuli. Initially, emotional stimuli provoke strong primary reactions that are positively or negatively valenced. The cessation of the stimulus brings the organism into an opposite state. As the result of the repeated presentation of the emotional stimulus, the primary reactions reduce in intensity (habituation) but the opponent state (the counterreaction) becomes even stronger (sensitization).
To explain these changes in dynamics of affect, Solomon (1980) developed the opponent-process theory (see Koob & Le Moal, 2008, for a variant of the model). Every emotional stimulus evokes a primary process that Solomon conveniently calls an a-process. This a-process is constant as long as the stimulus lasts. The a-process also does not change as a result of the repeated experience of the stimulus. It is determined only by the presence and intensity of the stimulus (the more intense the stimulus, the stronger the a-process). The organism reacts to the a-process with an opposite process, the b-process. In other words, the b-process is evoked by the a-process and its intensity depends on the intensity of the a-process. Initially, it takes some time for the b-process to get going, it is rather weak, and it lasts only a little while. After repeated presentations of the emotional stimulus, the b-process gets a shorter latency time (it is thus provoked more quickly), grows in strength, and lasts longer after the stimulation stops.
Importantly, the a- and b-processes are not directly observable; they are hypothetical constructs that researchers have come up with in the hope of explaining learning effects (in this case, the influence of repeated stimulus presentations on reactions and counterreactions). Solomon assumes that the observable emotional state of the organism is the result of the sum of the states of the a- and b-processes (see figure 1.9). Note that the a- and b-processes have opposite directions. Hence, the b-process can counteract the effect that the a-process has on behavior.

Schematic depiction of the opponent-process theory. Panel A shows what happens when the stimulus is presented for the first few times; panel B, what happens after repeated presentations. The top graphs depict the course of the manifest affective response, which is seen as resulting from the underlying opponent processes a and b (retrieved from Solomon, 1980, figure 7).
Another important idea that was added to the opponent-process model is that the b-process can be provoked not only by the a-process but also by stimuli that repeatedly co-occur with the b-process (see Schull, 1979, for a detailed explanation). The importance of this assumption is best illustrated in the context of drug abuse (see Drummond, Tiffany, Glautier, & Remington, 1995, and Siegel, 1989, 2008 for overviews). Researchers have found that the desire for a drug (i.e., drug craving) and the risk of relapse depend on context. More specifically, the desire for a drug and the risk of relapse increase in contexts in which a person often used drugs in the past. Suppose someone smokes a lot at work, but not at home. This person, if he or she stops smoking, will experience a stronger desire (craving) to smoke at work than at home. Often, drug users who successfully kick a heroin habit in a clinic will relapse if they are in situations similar to those in which they formerly used drugs. In the opponent-process theory, these context effects have been explained by assuming that stimuli that co-occur with drug use can trigger the b-process. This triggering of the b-process causes a “craving,” which could lead to using the drug in order to satisfy the “craving.” Not only external stimuli such as certain rooms but also internal stimuli such as feelings can be associated with the b-process. For persons who used drugs when they felt depressed or anxious, there will be a stronger craving for the drug when feelings of depression or anxiety return. Also, the context dependency of drug tolerance can be understood from this perspective (e.g., Remington, Roberts, & Steven, 1997). In situations where a person uses drugs regularly, the b-process is enhanced by environmental stimuli. This compensates for the direct effects of the drug, including dangerous physiological effects (e.g., increased blood pressure as part of a-process). If the same person uses the same amount in another situation, the b-process will be weaker (less tolerance), and this can lead to an overdose.
These insights are also important for designing treatments (see Drummond et al., 1995). For example, abstaining from drugs while being in a clinic may result in a reduction of the b-process (“cold turkey”), but it will not change the ability of external stimuli to trigger the b-process. Even after all withdrawal symptoms are gone, returning to situations that co-occurred with drug use in the past will trigger the b-process and hence increase the chance of relapse. A better approach is to expose the patient during the withdrawal to stimuli that were related to drug use. However, as we will see in the next chapter, such extinction procedures seem to have only a temporary effect (see the phenomenon of spontaneous recovery). Perhaps the best thing is to also teach the patient how he or she can cope with “craving” without returning to drug use.
The idea that the b-process can be elicited by stimuli that frequently accompany the process has proven very influential (e.g., Ramsay & Woods, 2014; Siegel, 2008). However, it raises questions about one of the key assumptions of the opponent-process model—namely, the idea that repeated presentations of a stimulus will lead to the strengthening of the b-process. Whenever a drug is presented, there will also be other stimuli that are present (e.g., the room in which the drug is used). The more you administer a drug, the more other stimuli will co-occur with the b-process, and thus the more the b-process can be enhanced by the presence of those other stimuli. It is therefore perhaps unnecessary to assume more than that the b-process strengthens as a result of the repeated administration of a drug. According to this view, changes in reactions and counterreactions with the use of drugs are not an example of noncontingent stimulus presentation effects (i.e., it is not the repeated use of the drug itself that is responsible for the change in the reactions and counterreactions), but rather an example of classical conditioning (the change in the reaction and counterreaction to the drug caused by stimuli that repeatedly occur with the drug). This is a good illustration of the fact that learning can never be observed directly. When one says that a certain type of learning has occurred (e.g., an effect of noncontingent stimulus presentations), this implies only a hypothesis about the cause of a change in behavior. Often, a similar change in behavior can be the result of several regularities in the area. The various hypotheses (types of learning) can then be distinguished by very carefully and separately manipulating the various regularities in the environment.
Notes
1. As pointed out by a reviewer, a related phenomenon can occur at the sensory level. For instance, if an organism is no longer able to sense certain stimuli (because of exhaustion or structural damage to sensory organs), then responses to the presence of those stimuli will also change. Again, one could argue that such changes in behavior are not instances of learning because they are not due to regularities in the presence of events. That is, like changes in the behavioral repertoire, changes in sensory abilities provide a potential alternative explanation for changes in behavior.
2. The first author of this book (Jan De Houwer) learned this statement from his mentor Paul Eelen, who had it written in large letters on the door of his office.