There are various mechanisms that have been proposed and could potentially be at work in any one patient for monomorphic VTach to develop. These mechanisms include intraventricular reentry mechanisms, increased automaticity (both normal and abnormal), and late and early afterdepolarizations. By far, the most common one is intraventricular reentry. In this chapter, we will review how an intraventricular reentry circuit can be formed and how it can generate monomorphic VTach. (The others are beyond the scope of this text, and the reader is referred to an advanced textbook on electrophysiology.)
We first looked at reentry in Chapter 25, AV Nodal Reentry Tachycardia, when discussing AVNRT. At that time, we stated that there were some preexisting requirements needed in order to form a reentry circuit. These included:
The presence of an electrical circuit with at least two pathways.
The two pathways involved need to have different underlying properties: conduction time, refractoriness, and so on. This could be due to a structural difference in the pathways, ischemia, electrolyte abnormalities, or any other event that will temporarily or permanently alter conduction and refractory time.
There has to be an area of slowing in one of the circuits—just enough to allow the rest of the circuit to get over its refractory period.
Let’s look at each of these requirements individually and see how they can take form to create VTach.
1. The presence of an electrical circuit with at least two pathways
VTach is more commonly found in patients with coronary artery disease, myocardial infarctions (MIs), and congenital abnormalities. Any, and all, of these disorders can lead to the formation of various isolated areas of myocardium that can act as a pathway. For simplicity, let’s look at MIs.
Myocardial infarctions are typically caused by obstruction to flow in the larger arteries supplying blood and nutrients to the heart. Sometimes, the larger arteries communicate with each other through smaller “tributaries” and the blood will continue to flow to an area even if the main artery supplying that area is blocked. This type of cross-circulation is known as collateral circulation.
In addition to the type of collateral circulation we just mentioned, there is another type that we will call micro-collateral circulation. This type of collateral circulation occurs because the exact area of perfusion by any one artery is sometimes “fuzzy” along the edges of its territory, and small areas along the outskirts are covered by the adjoining arteries.
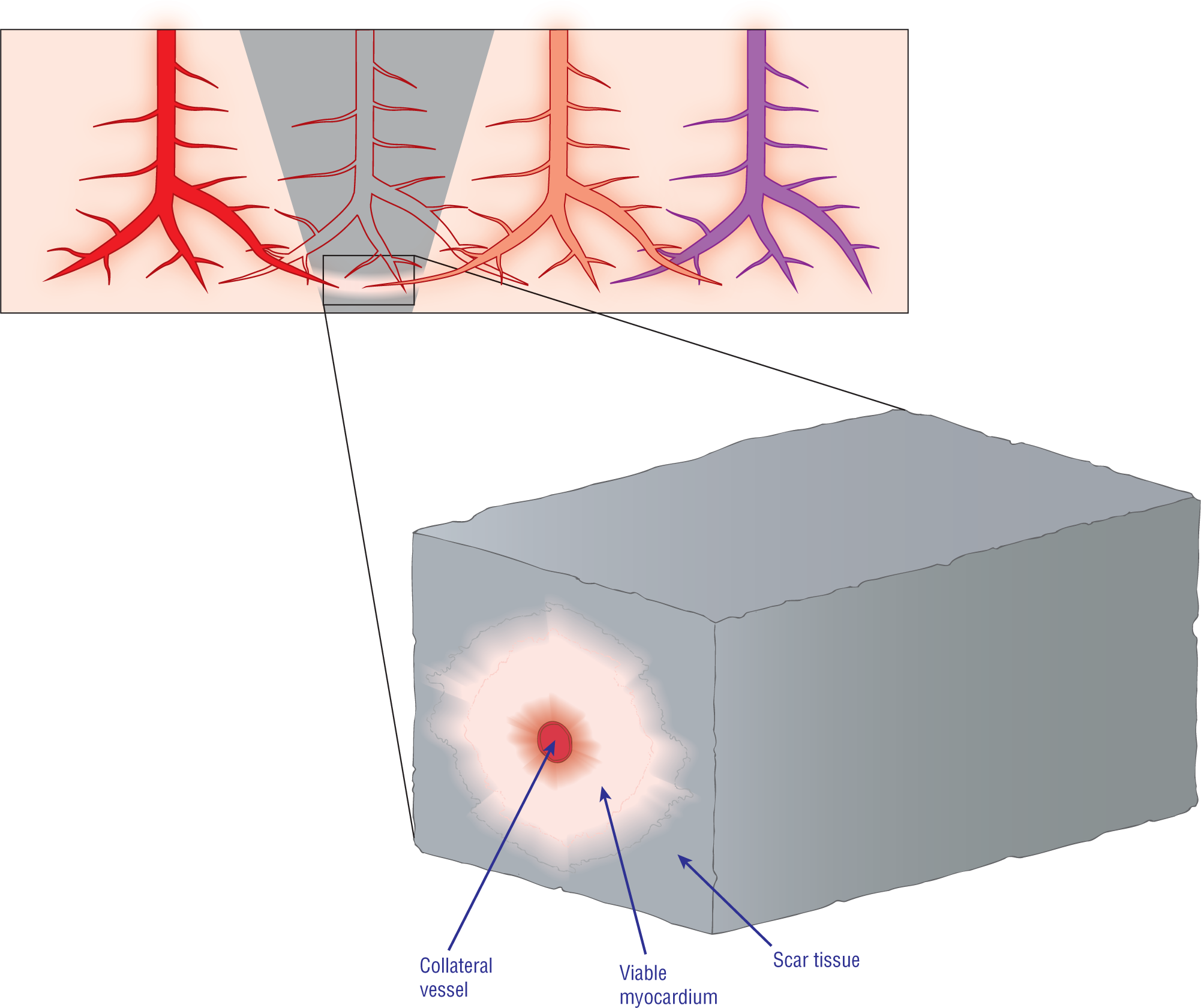
During a myocardial infarction (MI), the cells die and are eventually replaced by nonconductive fibrous scar tissue. Micro-collateral circulation will sometimes keep small islands or “bridges” of cells viable, despite the infarct that is occurring around them (see pink bridge of cells that are following the path of the collateral artery at the bottom of the gray wedge in Figure 32-3). These “bridges” of viable cells can continue to conduct the depolarization wave normally and function just like normal tissue. These “bridges,” therefore, can create the pathways that are required for the formation of a ventricular reentry circuit.
Figure 32-3 Microscopic view of an area of infarct (gray wedge) showing circulation from three different arteries. One artery, the colorless one, is no longer perfusing an area of myocardium, giving rise to the infarct. The two surrounding arteries (the red and pink ones) have some small blood vessels penetrating the infarcted area. This redundancy of arterial supply is known as collateral circulation and is a protective mechanism for the heart. Oftentimes, the collateral circulation will keep small regions or cords of cells alive. These cords can continue to transmit the electrical depolarization wave. These pathways are completely surrounded by nonconducting scar tissue, which acts like an insulation layer.
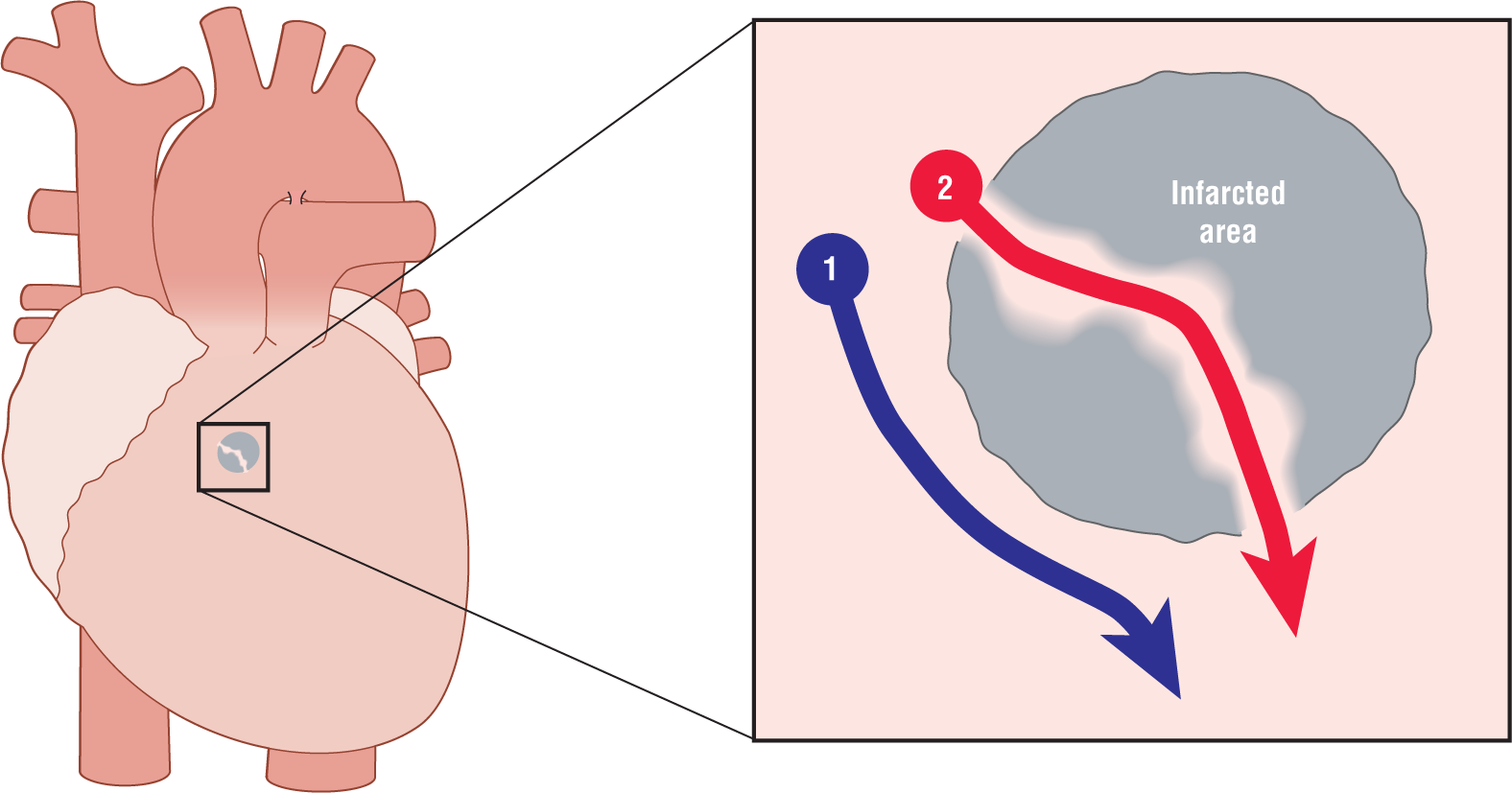
Figure 32-4 shows how two pathways can be present around an area of infarcted tissue in the ventricle. The red pathway, labeled with the number 2, is formed when collateral circulation has caused some sparing of viable cells in the areas shown. These “tubes” can conduct the depolarization wave through the center of the dead tissue or scar tissue that is formed when an MI has healed and form the pathways for a potential reentry circuit.
Figure 32-4 An area of infarct with some isolated islands and cords of viable cells forming an electrical pathway through the scar tissue (see red pathway 2). The blue pathway 1 is the normal route of depolarization around the infarct.
2. The two pathways must have different properties
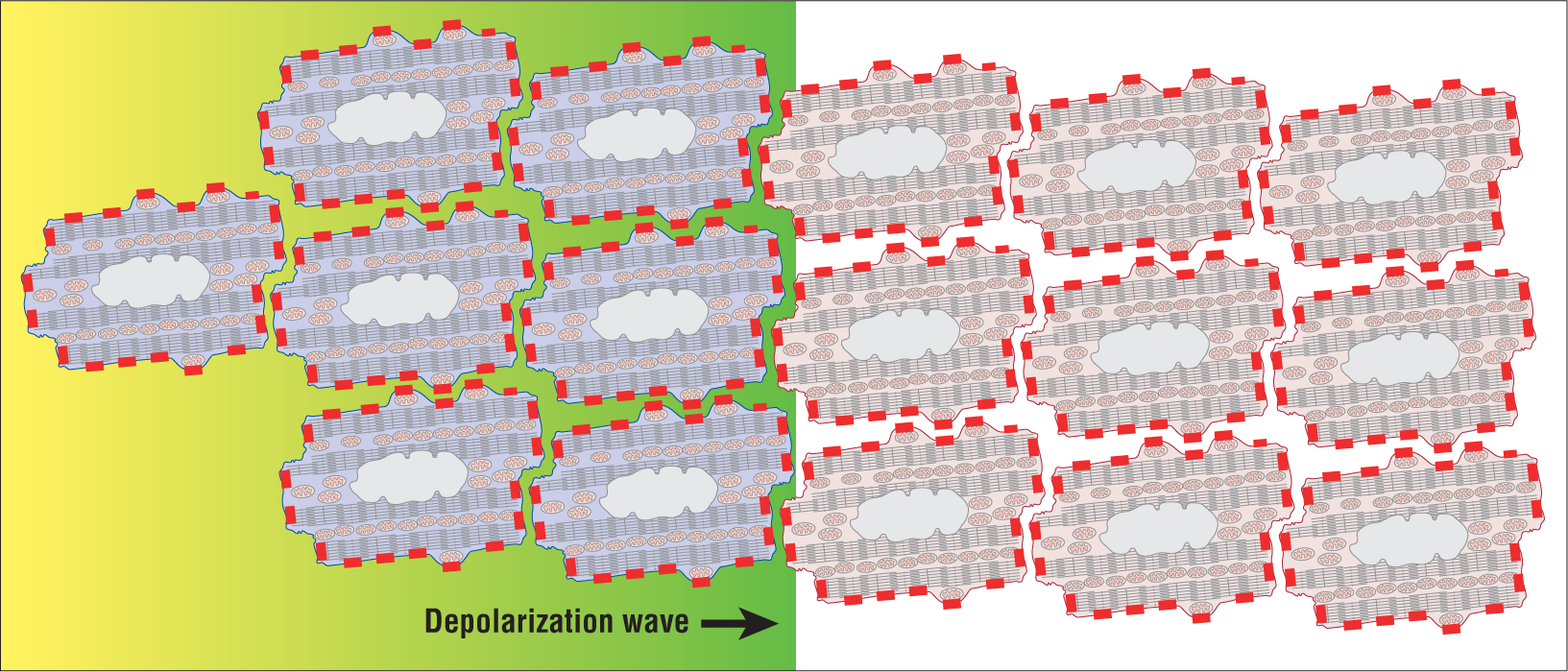
The next condition that must be met to form a reentry circuit is that the two pathways must have different properties. How does this fit into the pathways formed above? In the normal ventricle, any one myocardial cell is surrounded by other cells. As such, there are hundreds of connection points along the membranes where communication can take place between the cells (Figure 32-5). These connection points are called gap junctions. The larger the number of gap junctions present between two cells, the faster that the electrical depolarization wave is passed between them. This type of communication continues throughout the myocardium of the entire ventricle.
Figure 32-5 A depolarization wave as it is spread by direct cell-to-cell transmission. The transmission occurs between cells through biochemical communication at the gap junctions (red rectangles on cell membranes). Note the number of actual connections between cells.
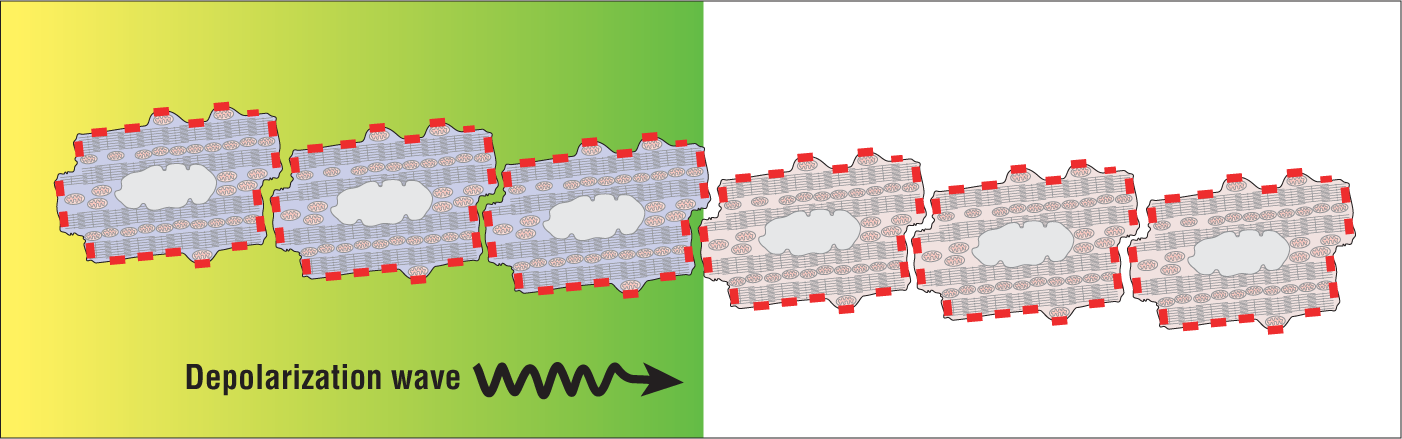
In general, the small viable groups and “tubes” of cells formed due to microcirculation are thin. Thinner tubes have fewer connections between cells than large three-dimensional groups of cells (Figure 32-6). In fact, these tubes can be so thin that only a few gap junctions at the end of cells may actually be in contact with each other. As you can imagine, conduction of the depolarization wave along these pathways is slower than through the regular, noninfarcted ventricle. This slower cellular communication creates a difference in the individual conduction rates between the two pathways. Let’s move on.
Figure 32-6 A depolarization wave as it is spread by direct cell-to-cell transmission. Note the small number of gap junctions (red rectangles) that are in contact with each other as compared with the cells in Figure 32-5. The low level of communication leads to slower conduction times.
3. An area of slowing in one of the circuits—just enough to allow the rest of the circuit to get over its refractory period
Now, let’s begin to put it all together. Figure 32-7 shows an area of infarct with a viable “bridge” of myocardial cells, which acts as a second pathway. Due to the low amount of communication in the “bridge,” the depolarization wave is spread much more slowly than outside the infarcted zone. This slowing allows enough time to pass so that the refractory period of the noninfarct myocardium is sufficiently repolarized to accept a new impulse. This, however, does not usually occur because the impulse enters both ends of the “bridge,” essentially nullifying the two wavefronts.
Figure 32-7 A depolarization wave (see green rectangle or wavefront) enters the intra-infarct pathway and is also normally spread around the infarct zone. The transmission through the intra-infarct “bridge” of viable tissue is conducted much slower than through the normal extra-infarct myocardium. Under normal circumstances, however, a reentry circuit is not formed because the impulses enter both ends of the bridge. The two wavefronts then meet inside the tube and cancel each other out.
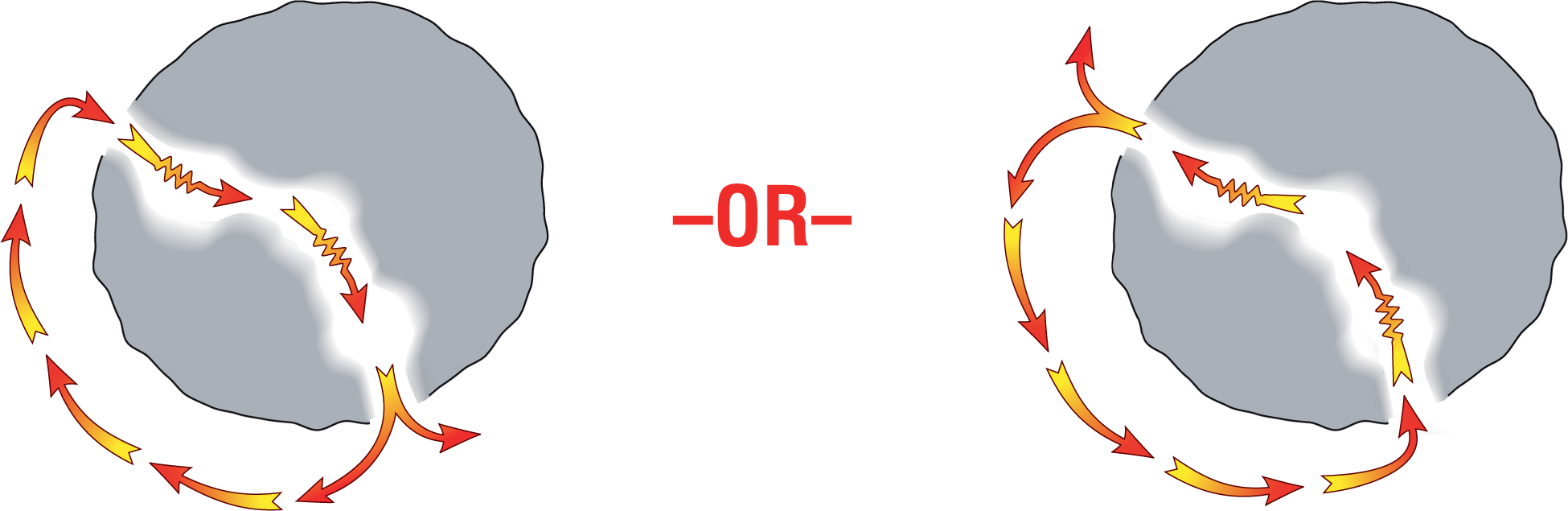
Under ideal arrhythmogenic circumstances, however, a reentry circuit can easily be started by a premature ventricular contraction (PVC). By mechanisms very similar to the one we saw when we examined AVNRT, a PVC can arrive earlier than expected and be blocked from traveling down the refractory pathway. Since conduction will not occur through that refractory pathway, the conduction must proceed exclusively through the other nonrefractory pathway. As we saw before in AVNRT, by the time that the impulse reaches the other end of the nonrefractory pathway, the original refractory pathway will be ready and willing to receive the impulse. This sets up the reentry circuit, which will continue to self-propagate and loop from this point on (Figure 32-8).
Figure 32-8 The two possible routes for a reentry using the pathway indicated.
Note that the direction of the circuit will depend on which pathway was initially refractory. The orientation of the resulting depolarization waves as they travel through the myocardium in potentially opposite directions will give rise to vectors, which can lead to morphologically different complexes on the ECG.