
![]() Groups of plant cells are organized into meristematic, vascular, dermal, and ground tissues.
Groups of plant cells are organized into meristematic, vascular, dermal, and ground tissues.
![]() Meristematic tissue is composed of undifferentiated cells that can become any type of cell in the plant.
Meristematic tissue is composed of undifferentiated cells that can become any type of cell in the plant.
![]() Vascular tissue is composed of the xylem and the phloem.
Vascular tissue is composed of the xylem and the phloem.
![]() The xylem transports water and nutrient ions up from the roots.
The xylem transports water and nutrient ions up from the roots.
![]() The phloem transports water and the sugars and other materials produced by a plant both upward and downward.
The phloem transports water and the sugars and other materials produced by a plant both upward and downward.
![]() The dermal tissue provides protection to the outer surfaces of a plant.
The dermal tissue provides protection to the outer surfaces of a plant.
![]() Root hairs are specialized dermal cells that extend into the soil and are the main points of nutrient uptake.
Root hairs are specialized dermal cells that extend into the soil and are the main points of nutrient uptake.
![]() Ground tissue makes up the bulk of the plant body.
Ground tissue makes up the bulk of the plant body.
![]() Leaves are plant organs that specialize in photosynthesis, which is the production of sugars from carbon dioxide, water, and the energy captured from sunlight.
Leaves are plant organs that specialize in photosynthesis, which is the production of sugars from carbon dioxide, water, and the energy captured from sunlight.
![]() Stomata are leaf pores that let in carbon dioxide and let out water vapor.
Stomata are leaf pores that let in carbon dioxide and let out water vapor.
![]() The evaporation of water from the leaf surface, in combination with water’s unique chemical properties, is what draws water into the vascular system.
The evaporation of water from the leaf surface, in combination with water’s unique chemical properties, is what draws water into the vascular system.
![]() Plants form symbiotic relationships with specialized nitrogen-fixing bacteria and mycorrhizal fungi, and these partnerships are key for the uptake of nutrients by plants.
Plants form symbiotic relationships with specialized nitrogen-fixing bacteria and mycorrhizal fungi, and these partnerships are key for the uptake of nutrients by plants.
JUST ONE LOOK at plants and you can tell that they’re made up of lots of different kinds of cells, not just the typical one I have been describing. Although plant cells may start out basically the same, at some point in their lives they differentiate and specialize in order to carry out the various functions required to enable the plant to grow and stay alive long enough to reproduce. It is these functions that allow scientists to organize plant cells into four types of tissue: meristematic, ground, vascular, and dermal tissues.
The appropriate place to start is with the meristematic tissue because it is the source of all new cells for plant growth and expansion. Meristematic cells are small and undifferentiated and have the ability to divide, a trait most plant cells lose at maturity. These cells have very large nuclei, small vacuoles (if any at all), and very thin cell walls. They are closely packed together. Meristematic cells are found only in the areas of the plant where growth actually takes place: at the tips of stems aboveground and just above the tips of roots underground. These are known as the shoot apical meristems and the root apical meristems. The development of these primary meristem cells adds to the height of the plant and the length of the roots, but not to the girth. Increases in diameter come from secondary or lateral meristem cells that surround trunks and limbs.
The cytoplasm of a meristematic cell is undeveloped. There are only precursors to plastids, for example, so the cell can be made into one containing chloroplasts or not. It just takes the right signal and enzymes go into action. Meristematic cells have very small vacuoles but lots of protoplasm filling them. This protoplasm is a stew of plant cell building blocks. Once a signal comes in, it provides the elements for construction. It’s like having a lumber and hardware store right there on the building site during the construction of a home.
One tenet of the Cell Theory is that cells multiply by dividing. The word meristem comes from a Greek word meaning “division.” Once a meristematic cell has differentiated, however, it loses the ability to divide. So, meristematic cells start out undifferentiated, and as division produces new cells the older meristematic cells are pushed outward or upward. At this point, a very complex signaling system begins the process that results in differentiation and specialization of the new cells. Thus, the meristematic cell transforms into one that is then part of another tissue group.
During this process an unbelievable number of proteins, many in the form of very specialized enzymes, flow into the cell. Hormones and other plant-made compounds, such as auxins that give plants directional growth, all activate processes that result in a cell’s differentiation. The transport proteins in the plasmalemma of meristematic cells are very active, as there is a tremendous amount of activity in the cytoplasm. The large nucleus responds to chemical signals that come though the plasmalemma and begins to direct the construction of ribosomes to process amino acid chains into proteins. The vacuole grows to hold nutrients and to regulate the pH of the cell. It also sequesters the growing amount of waste produced. Energy is provided by the mitochondria, and the brand new transit system of microtubules and actin fibers is put to use and enlarged.
During the period of cellular differentiation, there is a tremendous amount of development. RNA copies the DNA in the nucleus of the cell. Messenger RNA, using instructions copied from the DNA blueprints, constructs all manner of proteins in ribosomes attached to the endoplasmic reticulum as well as floating free in the cytosol. These proteins are transferred out of the endoplasmic reticulum or the cytosol (depending on where the ribosomes are located) to the Golgi apparatus for final packaging. Then the proteins move to the places in the cell where they are needed or to the plasmalemma to serve as a transport protein or to be transported to another part of the plant and combined with some other molecule to make a new compound.
Meristematic cells are the completely undifferentiated cells in a plant. Upon receipt of a signal, a meristematic cell develops into a specialized cell with a specific function, losing its ability to develop into any other kind of cell. In the aboveground parts of a plant (shoot apical meristem), new cells are constantly formed just below the differentiating ones. Those in roots are located above differentiating cells (root apical meristem).
At the top of this photo is a small group of slowly dividing meristematic cells located in the central zone of the meristem summit in apical meristem tissue. Cells of this zone function as stem cells and are essential for meristem maintenance. A short distance away (middle), the meristematic cells begin to differentiate. In this instance they can be seen developing into leaf epidermal cells that contain stomata (bottom). Had a different signal been sent to the undifferentiated meristematic cells, they would have taken on a different structure and served a totally different function—all still broccoli, but pretty amazing nonetheless.
The tip of a small broccoli flower bud with actively dividing apical meristematic tissue along the leading edges of the bud
Just what kind of cell a meristematic cell becomes depends on what the plant needs as a whole. All plant cells are connected by their plasma membranes and countless tiny plasmodesmata. One part of the plant can signal a need, and that signal is transmitted to one of the two types of meristematic tissues so cells of that type can be created.
Primary meristematic cells are responsible for producing cells that result in primary growth. This is growth that causes a plant to get taller. For the most part, growth doesn’t happen by adding cells, but rather by cells elongating. Primary cells are the cells that elongate.
Primary growth occurs at the apical meristems, which are located just above the root cap and in shoot tips. Hormones signal apical meristem tissue in the stem to produce cells in only one direction so the stem will grow upward. Root meristem tissue, on the other hand, grows in two directions, downward and outward.
Secondary growth is the growth that increases a plant’s diameter. This is what produces growth rings in woody plants. Secondary growth happens in lateral meristem tissue, which receives hormonal signals to differentiate. Not all plants undergo secondary growth. Monocots (maize, grasses), for example, lack lateral meristematic cells.
Undifferentiated truly means undifferentiated. For example, a shoot meristem cell can be moved to a growing tip where it will differentiate into a bud, leaf, or flower cell. In fact, each meristematic cell is capable of cloning the entire plant. All of the ground, vascular, and dermal tissues come from meristematic tissue—that is, all of the plant’s organs: its leaves, stems, and roots.
Ground tissue provides most of the mass of a plant as well as its support. These cells also serve as sites for photosynthesis, food storage, protection, and regeneration after injury. There are three types of ground tissue cells, again grouped by function: parenchyma, collenchyma, and schlerenchyma.
Parenchyma cells are responsible for metabolic functions. They have thin primary walls and large vacuoles. In fact, of all the cells in the plant, these are most like the typical cell described in chapter 1. Although parenchyma cells retain the ability to divide, they don’t use it very often. This trait allows a plant to regenerate parts of itself when damaged or when it is under extreme stress. Parenchyma cells are not fully specialized cells, and they retain the ability to become more specialized. They are capable of growing, and they function in photosynthesis or storage. There are lots of parenchyma cells in leaves and roots. Fruit flesh is made up of parenchyma cells, and potatoes and other tubers are full of them.
Collenchyma cells form bundles and strands (the strings in celery, for example) and help support plants. Because they lack a secondary (cell) wall and lignin, collenchyma strands are flexible and the plants they support are as well. They can gently bend in the wind, as it were.

Parenchyma cell
Schlerenchyma cells develop a lignified secondary wall and cannot elongate. These cells serve as support and provide some physical protection to the plant. In fact, most are no longer living. They die when they reach maturity, leaving their lignin shells. Sclereids, a special kind of schlerenchyma cell, make up the hard coatings on seeds and hard nut shells, as well as the skin of a pear, which is why it is harder and tastes different from the skin of apples.
The vascular tissue consists of two types of cells that transport materials in a plant. The xylem carries water and nutrient ions in it, and the phloem transports sugars and organic molecules synthesized inside the plant. Almost no cell is further than a few neighboring cells away from the vascular system.
Phloem and xylem cells are appropriately known as vessels, as they hold water. They run throughout the plant, side by side but separated by the vascular cambium, a wall of lateral meristematic cells that provide lateral growth. As the vascular cambium cells differentiate and mature, they develop into phloem and xylem cells.

This scanning electron micrograph of a leaf vein in cross section shows the outer epidermal layer (with spiky projections known as trichomes), followed by the cortex and then large vascular bundles, which contain the phloem (nearest the cortex) and xylem.
 Xylem The xylem tissue plays a huge role in the story of how plants eat. The xylem is a system of specialized cells, connected end to end, that transports water along with mineral nutrients dissolved in it. This oneway system carries water upward to the leaves of the plant, where xylem tissue is contained in veins. These are placed so efficiently that no single plant cell is further than about 0.5 millimeter (0.02 inch) away from one and so can be readily reached by water traveling from the xylem system and then through cells using the symplastic, apoplastic, and direct pathways. Water in the xylem only flows upward from the roots to the leaves. The connections between xylem cells are much wider than the cell-to-cell plasmodesma, thus providing much less resistance to the movement of water.
Xylem The xylem tissue plays a huge role in the story of how plants eat. The xylem is a system of specialized cells, connected end to end, that transports water along with mineral nutrients dissolved in it. This oneway system carries water upward to the leaves of the plant, where xylem tissue is contained in veins. These are placed so efficiently that no single plant cell is further than about 0.5 millimeter (0.02 inch) away from one and so can be readily reached by water traveling from the xylem system and then through cells using the symplastic, apoplastic, and direct pathways. Water in the xylem only flows upward from the roots to the leaves. The connections between xylem cells are much wider than the cell-to-cell plasmodesma, thus providing much less resistance to the movement of water.
Vascular tissue consists of xylem and phloem cells.
The xylem’s pipes are very elongated cells and are either tracheids or vessel elements that are joined together. Tracheids are the dominant of the two types in gymnosperms, including conifers, cycads, and ginkgoes, all softwoods. The end walls of tracheids have pits, thin areas through which water can flow. These cells are tapered and overlap, and water flows from one tracheid cell to the next. They are about the width of a human hair.
Vessel elements, on the other hand, dominate in hardwoods. The ends of these xylem cells are perforated with actual holes. In addition, vessel elements are actually connected, not just overlapping, so there is even less resistance to the movement of water than through tracheid pits. The vessel elements are about six times wider than tracheids, measuring about 500 microns. When linked together, they form vessels that can be a whopping 10 cm long, about the length of your forefinger.
Both tracheids and vessel elements have walls strengthened with lignin, the material that makes wood. This provides support, preventing xylem tubes from collapsing in on themselves. Xylem cells die at maturity, but the lignin allows their shapes and functionality to remain after their living contents disappear. The lignin also helps to waterproof the tubes to form a continuous, unidirectional plumbing system that moves water and its contents upward from the plant roots through the stems and branches to the leaves.
Scanning electron micrograph of a conductive vessel element (sieve tube) in wood
Water Transport in the Xylem Water transport via xylem is one of the wonders of nature. For starters, it all occurs without the expenditure of any energy by the plant. Imagine how much power would be required to pump water up to the top of a redwood tree. Yet, that tree can bring water to its leaves without using its valuable energy reserves. We would not have tall trees if energy were required to deliver water through them.
Water’s journey into the plant starts at its roots. By now you should be aware that water can move into a root cell and travel for a bit in the cell wall via the apoplastic pathway. Once it hits the Casparian barrier, however, water has to move through the cell wall and cell membrane (the plasmalemma), probably via an aquaporin, thus entering the cell and the symplastic pathway. Once inside the plant cell, water moves from cell to cell and makes it way to the xylem tissue located toward the center of the root.
This raises two questions. First, why does the water enter the plant? Molecules of water enter the plant root because of root pressure. There is usually a higher concentration of nutrients than water inside root cells. As a result of osmosis, water outside the roots moves across the root cell membranes to dilute these concentrations inside the cells.
The second question is why water inside a root moves toward the xylem system. Here, there are three forces at play: transpiration (the evaporation of water from the surface of the plant), water cohesion, and water adhesion. To understand transpiration, one needs to consider the leaves of the plant. The stomata are leaf pores that open during the day to let in the carbon dioxide needed to make sugars. At the base of these openings are the ends of xylem tubes. Water in the xylem evaporates out into the atmosphere when the stomata are open. Some leaves also have trichomes, tiny hair-like structures that help draw the water that evaporates out, up, and away from the leaf body where there is more air circulating, speeding the evaporative process.
As we learned in the chapter on basic chemistry, water molecules are polar, with one side having a negative charge and the other a positive charge. This causes water molecules to stick together—that is, to show cohesion. If you lined up water molecules one after another the entire line could be pulled like a rope because of this attractive force. This means that when one water molecule leaves the stomata and evaporates away, it pulls up the molecule behind it, and this happens all the way down the chain. This creates a negative pressure at the bottom of the column, which pulls new water molecules into the system. Water in the roots is literally pulled up through the xylem straw. Once inside the xylem, the water molecules and ions dissolved in it can travel at up to 30 meters (100 feet) per hour.
Ah, but there is more help. The force of adhesion causes water molecules to stick to the surface of their container, in this case the xylem tubes. Adhesion is why a concave meniscus forms when water is held in a glass tube or straw. The water on the edges is higher than the water in the middle of the surface, and because all the water molecules stick to each other there is a tension created. The tension is enough, in combination with water cohesion, to fill up a redwood-sized xylem system.
At night, the stomata receive signals to shut and transpiration ceases. This is the time when plants respire; they utilize the water and nutrients brought into the system by transpiration during the day. Although water is no longer moving upward through the xylem, cohesion and adhesion hold the water column in place.
High magnification image of a lower leaf surface showing stomata and some fungi attached to the leaf
Phloem The phloem is the other half of a plant’s vascular tissue. Its function is to transport the sugars made during photosynthesis—along with the amino acids and other organic compounds synthesized by cells, even RNA—to where they are needed. The phloem system consists of only living cells and operates bidirectionally, though not in both directions at the same time.
The phloem distributes sap from a source, such as a chloroplast in a leaf cell or a storage cell in a root, to a sink, with cells in low supply. During summer, the source is usually in the leaves. Sap flows through the phloem to the roots when things are stored for the winter. In the spring, these stored nutrients move from the roots up into the aboveground parts of the plant, only on this trip the sap travels in the xylem. This is the source of maple and birch syrups and is why trees are tapped only in the spring.
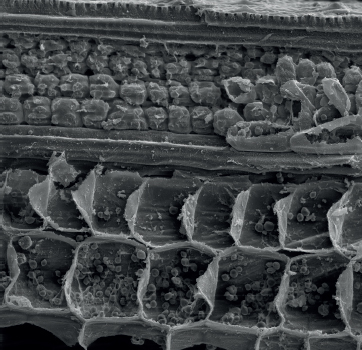
This scanning electron micrograph shows the vascular (bottom layer) and cortex cells of a stem. Starch granules are visible inside vascular cells, and chloroplasts are visible inside cortex cells.
Phloem cells are specialized and elongated cells that lack lignin. These sieve elements are connected in vertical stacks to form sieve tubes, which are separated by sieve plates, perforated structures. The holes of a sieve plate are lined with callose, a polysaccharide. As they mature, sieve element cells lose their nuclei, vacuoles, and ribosomes, but they retain plastids, endoplasmic reticulum, and mitochondria. At maturity, sieve element cells are alive in a way that xylem cells are not.
Because each sieve element is missing a nucleus (and other important organelles), it functions by pairing up with a companion cell that carries out the functions of its missing organelles. Sugar molecules to be transported by the phloem system are dissolved in water and flow into the companion cell, where they are collected and then actively loaded into the sieve tubes. Energy is required to move the sugar into the system.
The phloem depends on hydrostatic pressure, not transpiration, to move the water that contains sugars and synthesized molecules. As sieve tubes are filled with sugar molecules, the concentration goes up inside the tubes. Osmosis causes some of the water molecules in adjacent xylem vessels to flow in to dilute it. There is only so much room for the water moving into the cell, which eventually starts to expand. The positive pressure created inside the phloem pushes the sap through the system.
At the same time, at the other end of the phloem system there is an area of low concentration of ions, the sink, which will receive the phloem sap. Ions flow into the sink through plasmodesmata, to lower the concentration of sap at the source. This relieves some of the pressure in the system, which also causes sap flow toward the sink.
Stems While not part of the vascular tissue, stems evolved to protect it as much as to provide support for the leaves, fruits, and flowers. Specialized cells on the outside of stems protect the insides and act as waterproofing. The stem also has cells that act like Styrofoam packing around the xylem and phloem to protect the vascular tissue.
The dermal tissue forms the skin of the plant, keeping in water and protecting the plant from external injury. There are two parts to the dermal tissue: epidermis and periderm. All plants have epidermal cells, whereas only woody plants produce a periderm. Both are composed of tightly packed cells that help provide protection.
The epidermis is usually just a single layer of cells that covers plant surfaces. This tissue is found on flowers, leaves, stems, roots, and even seeds. In general, epidermal cells lack chloroplasts and are transparent.
Epidermal cells produce cutin, a waxy substance that helps form a cuticle that covers all aboveground surfaces of the plant and prevents water loss. The cuticle is usually thicker on top of leaves, where there is more exposure to sunlight, than on the bottom of leaves.
Epidermal cells include the stomata, which contain guard cells that open to allow carbon dioxide to diffuse into the leaves and water vapor and oxygen out. In addition, trichomes, tiny leaf hair cells that help in transpiration, are part of the epidermal tissue. Some trichomes possess glands that produce anti-grazing substances that protect plants from herbivores. Others act as foils, decreasing the flow of air over the surface of the plant and slowing the loss of water from it.
As some plants age, a periderm is formed from lateral meristematic cells. This replaces the epidermis of stems and roots and is more commonly called bark. The periderm not only provides protection, but also allows airflow into the plant. Cork, dead cells full of suberin, is made from periderm.
Root Hairs Specialized epidermal cells on roots, known as root hairs, are extremely important to the uptake of nutrients. Root hairs dramatically increase the surface area of the root that is exposed to soil. Root hairs form because epidermal cells along the outer surface of a root have the ability to change from their normal shape and placement on the surface of a root into an extension of the root that grows out into the soil. Each root hair is a single cell, and these cells can grow to amazing lengths of up to 1500 microns (0.06 inch) and can be as thick as 15 microns (0.0006 inch).
Only a small zone located just below a growing root tip contains meristematic cells coded to become root hair cells. These cells live only from a few days to a couple of weeks, depending on the plant and conditions. Root hairs are constantly being replaced as the growing tip moves into new territory. Still, an astonishing number of these cells can grow at one time. Some plants produce tens of thousands of them in an area no bigger than an inch or two around. For example, studies have shown that a single rye plant can have miles of root hairs growing in 2 cubic feet (0.06 cubic meters) of soil.
Only some of the root epidermal cells form root hairs, because those that do secrete chemicals that prevent their neighboring cells from becoming root hairs, too. If this didn’t happen, plants would have huge tangled roots, made up of ever-expanding epidermal cells.
Root hairs act as the eyes of a plant root. Calcium, one of the essential plant nutrients, moves into the root hair through the plasmalemma. The presence of calcium is necessary to complete the process that causes root hair cells to grow downward. As long as there is calcium coming into the root hair from the soil, the root hair cell elongates in the same direction. When an obstacle, say a small rock, is encountered, then that surface of the root hair cell can no longer take up calcium. Elongation in that direction stops, but it starts in another area of the cell where calcium does come through the membrane. Once the obstacle has been passed by the root hair, it reorients itself and travels in the original direction. Consider that this goes on all over those miles of hairs. Calcium shortages have obvious consequences here.

Scanning electron micrograph of root hairs developing on a radish during seed germination. These cells release a hormone that prevents neighboring cells from developing into root hairs.
Root hairs are tremendously important, but their development is discouraged by the presence of too many soil nutrients. Producing root hairs takes energy and nutrients, and a plant only does this when it is necessary to get more nutrients. If there are enough nutrients, why expend the energy?
The key function of leaves is to provide the platform for photosynthesis. They evolved to best expose as many chloroplasts to sunlight as possible. Leaves also house the stomata, the exchange organs that allow carbon dioxide to move into a plant and water vapor and oxygen to move out.
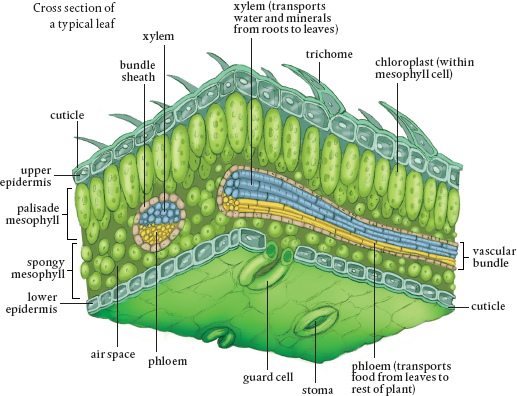
A typical leaf, again assuming there is a typical one, comprises the leaf blade or lamina; the petiole, the stalk that attaches the leaf to the stem; and the leaf axial, a small structure where the petiole attaches to the stem. The xylem and phloem tubes run through the petiole into the lamina. Many plants are identified based on the shape and size of their petioles, which can be circular in diameter, winged, triangular, or flattened.
Leaves are coated on both sides with epidermal cells. Some also have trichomes, hair-like structures, on their upper and lower surfaces. Most leaves are coated on the outside with a waxy substance that forms a cuticle that keeps in water and provides protection. Many metabolic compounds are released into epidermal cells to protect the plant, and these cells are responsible for the regulation of water loss and gas exchange with the atmosphere.

This leaf shows a vein pattern such that no cell is more than three away from a vein.
The chloroplasts reside in between the two layers of epidermal cells, in the mesophyll. The mesophyll is also full of parenchyma and collenchyma cells as well as xylem tubes that provide the water necessary for photosynthesis. The mesophyll also has lots of phloem structures to take away the sugars produced in chloroplasts. When large enough, this network of vascular tissue becomes veins, which are usually sheathed in parenchyma cells and supported by collenchyma cells. The type of leaf venation of a plant is used to describe the plant.
Leaf epidermal cells contain the guard cells that regulate the opening and closing of the stomata. When water rushes into the guard cells, they fill up, swell, and bend on one side and shrink on the other. This results in a hole, the stomatal opening. When guard cells lose water, they return to their flattened shape and the stomata close. Potassium, one of the essential plant nutrients, is key in the operation of stomata.
One function of the epidermis of a leaf is to keep water in when the stomata are closed. However, at night when there is excess water in the soil compared to inside the root cells, a process known as guttation occurs. This is the exudation of liquid water from leaf tips. The water flows into the plant and creates enough pressure to force water into the leaves, where it escapes through hydathodes, tiny holes along leaf tips.
Let’s end this brief botany lesson with a look at roots. Of course, there isn’t a typical root. Some are fibrous and branched; these fibrous roots usually grow near the surface and are laid out to absorb the maximum amount of water quickly when it rains. Others form the familiar carrot-like taproot that can reach deeper water. Still, there are certain characteristics applicable to most garden plants that make a general model of a root possible.
New cells are produced in the root apical meristem and are pushed out toward the root tip, where they differentiate into columella cells, rectangular cells that line up in columns. These get squashed as new cells are produced above them, and this compaction makes room for new cells. Columella cells form the root cap that protects the apical meristem while the root moves through the soil.
The root cap cells contain statoliths, specialized organelles that appear to help in the detection of gravity and are partially responsible for the downward direction of root growth. These organelles are heavy and sink to the bottom of the cell. In essence, they act as weights and help roots sense gravity. This causes a series of chemical signals to pass through the cell membrane, resulting in increased production of the growth hormone auxin in the area.
Root cap cells help build the mucilage that lubricates the root and the soil, making it easier to penetrate and increasing the contact between the soil and the root. Eventually, the outer cells of root tips are sloughed off and replaced by new root cells produced in the meristematic tissue. These sloughed off cells supplement the plant’s exudates (sugars and amino acids, mostly) that attract microbes necessary for nutrient absorption to the rhizosphere. The mixture of microbes living in and digesting this sloughed-off cellular material generates wastes that contain essential plant nutrients, particularly nitrogen.
Mucilage has a great influence on the uptake of nutrients, particularly metal nutrients, into plants. Phosphorus, zinc, iron, and magnesium all diffuse into the mucilage gel and from the gel into the roots. This gel is particularly apt at oozing into soil particle crevices and pores, thus increasing contact with the surfaces where these metals are located. Mucilage contains acids that dissolve phosphorus, which can then diffuse through it to the root. In addition, chemical reactions that occur in the mucilage and the gel’s pH result in the uptake of metals. Plant exudates become part of the mixture, produced, at least in part, in direct response to the need for some nutrient or a need to turn off a nutrient supply, which requires a change in the mixture of microbes. In any case, the presence of mucilage contributes to the rhizosphere chemical factory. Some plants take up various metals in greater quantities than others due to the makeup of the root mucilage mixture.
Cells on the upper side of the root’s apical meristem move upward. As they do, they form a zone of elongation where, as the name suggests, cells grow by elongating. These cells are little biohydraulic pumps working away to force roots through soil. Their empty vacuoles fill with water as they grow, creating hydraulic pressure inside the cell and making the root tip move downward. Just above this area is a zone of maturation, where meristematic root cells differentiate into their mature form and functions and become part of one of the other plant tissues, the dermal, ground, or vascular tissues.
THE CROSS SECTION OF A TYPICAL ROOT
The root cap protects the root apical meristematic cells. These cells have the ability to differentiate into all of the structures shown in the diagrams: from root hairs, the entry points for nutrients, to the vascular system that transports water up into the plant (xylem) and sugars and other compounds throughout it (phloem). In fact, if you moved a root meristematic cell into a shoot apical position, it would generate into a tip-related part of the plant and not a root-related one.
The elongation of cells in the root provides some of the hydraulic pressure that drives the root forward through soil. Mature parts of the root usually stop producing root hairs. Cells of the root cap are constantly being replaced along with other root cells, and those that are sloughed off add to the carbon that attracts bacteria and fungi to the rhizosphere, the area immediately surrounding roots.
The endodermis is the dividing point between the apoplastic and symplastic pathways for water. It contains the Casparian strip, a layer of cells filled with the waxy substance suberin, which stops the movement of water through the cell walls so that it must travel the symplastic pathway. Because cell membranes are much more selective than porous cell walls, this allows the plant to better control what it takes in.
Different kinds of plants have different vascular bundles, and their arrangement is a distinguishing factor be tween monocot and dicot plants. All of this specialization starts with the undifferentiated meristematic cells.
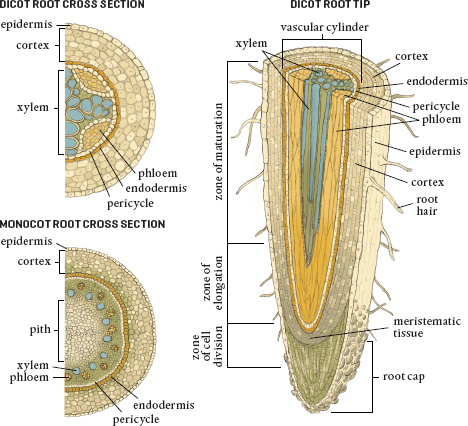
The major parts of a typical root from various perspectives
If we were to look at a cross section of a root, at the very outer edge we would find the epidermis, or outer skin. This layer of cells covers the entire root except for the area immediately around the root cap. The portion of the epidermis just below the zone of elongation is where root meristematic cells specialize and become root hairs.
The next layer in is the cortex, which is composed of lots and lots of parenchyma cells that store food, especially in the form of starch. On the inside of the cortex, toward the center of the root, is a layer of cells known as the endodermis. These cells are squished together so tightly that they lack any space between them. This is the location of the Casparian strip, the barrier that prevents water (or anything else, for that matter) from traveling unregulated farther into the plant.
On the inside of the endodermis is the stele, the central part of our model root. The outer boundary of the stele is the pericycle, a thin layer of cells that retain the ability to divide. These form lateral (branch or side) roots, which are quite different from root hairs. They grow through the cortex and the epidermis and out into the soil, often developing root hairs themselves. Pericycle cells form new cells that help thicken the root.
At the center of the root, inside the pericycle, are the vascular tissues. Here the xylem system begins to transport the water that enters the root hairs upward though the plant. The phloem also starts here and brings nutrients down from the leaves to the roots for use or for storage, or it carries stored materials back up into the aboveground parts of the plant.
All gardeners appreciate what special organisms plants are, although many never fully appreciate how their parts work. At this point, you should be able to contemplate water and nutrients entering the root hairs of a plant as a result of diffusion, and then moving through the layers of the root and entering the xylem, where these materials move up through the stem and petioles and into the leaves. Similarly, you can imagine the phloem in the root as it moves stored sugars and organic molecules from the storage cells in the root up into the leaves or as it carries these materials down from the leaves where they were produced.
Don’t get hung up on all the names used to describe the parts of a plant. Just try and revel for a moment (in three dimensions) over the elegant organization of a plant. Hopefully, I have given you something to contemplate during some of the quieter moments out in the garden.
![]() Apical meristematic tissue is located just below the tips of shoots and just above the root caps. These cells can mature to become any type of cell in the entire plant and are responsible for increasing plant height and length. Lateral meristem cells add diameter to plants.
Apical meristematic tissue is located just below the tips of shoots and just above the root caps. These cells can mature to become any type of cell in the entire plant and are responsible for increasing plant height and length. Lateral meristem cells add diameter to plants.
![]() Ground tissue provides most of the bulk of a plant as well as its support. These cells also serve as sites for photosynthesis, food storage, protection, and regeneration after injury.
Ground tissue provides most of the bulk of a plant as well as its support. These cells also serve as sites for photosynthesis, food storage, protection, and regeneration after injury.
![]() Vascular tissue is made up of xylem and phloem vessels that transport water and nutrients.
Vascular tissue is made up of xylem and phloem vessels that transport water and nutrients.
![]() The xylem carries water and dissolved nutrients upward from the roots. Transpiration, cohesion, and adhesion allow water to move through the xylem without the plant expending any energy.
The xylem carries water and dissolved nutrients upward from the roots. Transpiration, cohesion, and adhesion allow water to move through the xylem without the plant expending any energy.
![]() Phloem vessels move sap (water and materials produced by the plant) both upward to the leaves and downward to the roots.
Phloem vessels move sap (water and materials produced by the plant) both upward to the leaves and downward to the roots.
![]() Dermal tissue forms the skin of the plant, which keeps in water and helps to protect the plant from external injury.
Dermal tissue forms the skin of the plant, which keeps in water and helps to protect the plant from external injury.
![]() Stomata are pores in the epidermis that open to allow carbon dioxide to diffuse into the leaves and water vapor and oxygen out.
Stomata are pores in the epidermis that open to allow carbon dioxide to diffuse into the leaves and water vapor and oxygen out.
![]() The key function of leaves is to provide the platform for photosynthesis. Chloroplasts reside in cells between the two layers of epidermal cells that cover each leaf.
The key function of leaves is to provide the platform for photosynthesis. Chloroplasts reside in cells between the two layers of epidermal cells that cover each leaf.
![]() Root hairs are extremely important to the uptake of nutrients. Each is a single epidermal cell that can grow to an amazing length.
Root hairs are extremely important to the uptake of nutrients. Each is a single epidermal cell that can grow to an amazing length.
![]() Roots take in water and nutrient ions with the aid of microbes that live in the soil.
Roots take in water and nutrient ions with the aid of microbes that live in the soil.
![]() Mucilage is the mixture of sloughed off root tip cells, exudates from root tips, and microbial populations and by-products. It acts as a lubricant for roots growing into soil and influences the uptake of nutrients, particularly metal ions.
Mucilage is the mixture of sloughed off root tip cells, exudates from root tips, and microbial populations and by-products. It acts as a lubricant for roots growing into soil and influences the uptake of nutrients, particularly metal ions.