
What Is a Wolf?: Classification of North American Wolf Species, Subspecies, and Populations
By Matthew A. Cronin, PhD

Photo credit: Holly Kuchera/Shutterstock.com
Introduction
The previous chapter describes the problems with hybridization of wolves and how this influences their management. In this chapter I describe how and why the scientific community is so divided on wolf taxonomy.
In order to know how to best manage wildlife, it’s essential to know what you are managing. For most people that means taking out a field guide and comparing what you are seeing with what the book says.
The forerunners of today’s field guides date back at least to Herodatus, who compiled a book of beasts in the fifth century BC. His work was expanded on about two hundred years later by Aristotle in Historia Animalium, which stood as the definitive work in the field the publication of a second-century text by Physiologus, a pen name. In the first century AD, Pliny the Elder wrote Historia Naturalis, which marked the founding of the science of natural history.
In the twelfth and thirteenth centuries, science and religion markedly separated and heatedly sparred for control of the central values of culture. Catholic priests working with the blessings of Albert the Great, Saint Thomas of Aquinas, and others sought to bring together the collected wisdom about the animal kingdom into voluminous, handwritten books called bestiaries. Drawing upon earlier works, folk tales, hearsay, personal accounts, and scholarly interpretations, bestiaries become monumental tomes, handwritten in Latin and often richly illustrated. Works of art as much as science, bestiaries were largely penned from secondhand sources rather than personal field studies of the authors.
For the Christian monks of the Middle Ages, recording the kingdom of nature for posterity was also way to guide new generations away from pagan religious beliefs about animals, for wild creatures were often the gods of the old religion. Thus the monks who penned bestiaries often advanced interpretations of animal behavior that supported Christian teachings as much or more than fact. Animals that had especially offensive qualities were made agents of the devil. The crocodile stood for the sin of pride, the viper for lust, and the partridge for covetousness, along with other familiars of Lucifer including the monkey, the wolf, the fox, the ass, the goat, the dragon, and the whale. (The whale was included as toothed whales could attack boats and tear them apart.)
A breakthrough for science in interpreting the meanings of animals came in 1732 through the fertile mind of twenty-five year-old Carolus Linnaeus. In l735 Linnaeus published his thesis about categorizing animals according to their physical structure in Systema Naturae, forever influencing the nature of scientific thinking by a system of classification into kingdom, phylum, class, order, family, genus, species, and variety. In modern times “varieties” have been renamed “subspecies” for animals.
The wolf was first classified as a species (Latin name Canis lupus) by the Swedish naturalist Linnaeus in 1758. This included all of the wolves worldwide with a range that includes most of the Northern Hemisphere north of 30 degrees latitude, and south into India, Pakistan, the Middle East, and North Africa.
Since Linnaeus, classifications of wolves have included numerous subspecies, and recently new species and populations in North America. These classifications are important because species, subspecies, and populations (i.e., distinct population segments, or DPS) are all considered in the definition of “species” by the US Endangered Species Act (ESA).
The US Fish and Wildlife Service (FWS) and National Marine Fisheries Service (NMFS) implement the ESA and determine if a group qualifies as a species, subspecies, or population, and if it is threatened or endangered with extinction. The National Research Council has addressed taxonomic (i.e., classification) issues related to the ESA and notes that there is considerable debate among scientists as to what qualifies as a species, subspecies, or DPS.1
In this chapter, I review the current taxonomic status of wolves in North America beginning with a description of how scientists classify species, subspecies, and populations, and describe what has been happening on this topic, especially with regard to wolves. A key point I will emphasize is that designation of species, populations, and subspecies in particular, is not scientifically definitive.
Taxonomy
To understand wolf classification we must understand the science of biological classification, known as taxonomy. The basis of taxonomy is that organisms (plants and animals) are grouped according to phylogeny, which is simply the genealogy or ancestry of a group. In simple terms, we classify animals according to common ancestry, so two groups with recent common ancestry are classified together, and groups with more distant common ancestry are classified as different groups. This has been recognized since before Darwin.2 Groups in taxonomy are called taxa (singular taxon). The basic groups of taxonomy are kingdom, phylum, class, order, family, genus, and species. For example, wolves are in the animal kingdom, chordate phylum, mammal class, carnivore order, Canidae family, Canis genus, and lupus species. There are sub-levels of each of these levels as well, including subspecies, so a wolf subspecies, such as Mexican wolves, is further classified with a third Latin name (Canis lupus baileyi). For many plants, a category “variety” has been substituted for subspecies.
There are other methods of classifying animals besides using ancestry. For example, one might classify animals as aquatic (living in water) or terrestrial (living on land). In this case, aquatic fish and whales would be classified together, but separate from land mammals. However, we know that all mammals, including whales, share a more recent common ancestry than any mammal does with fish, so in biology, aquatic whales are classified as mammals along with terrestrial mammals. Another example is aquatic sea otters and river otters. They are classified as mustelids along with the terrestrial weasels, mink, fisher, marten, skunks, badger, and wolverine, and not with the aquatic, but distantly related, seals, sea lions, and walruses.
Another non-taxonomic classification is the designation of “ecotypes,” which are groups of populations of a species with similar characteristics due to local adaptations or environmental influence, not shared ancestry. Hence, ecotypes are not a part of taxonomic classification. An example is caribou (Rangifer tarandus), that include high arctic, mountain, forest, and barren-ground tundra ecotypes. These ecotypes have different characteristics in different ecological conditions but members of an ecotype do not necessarily share common ancestry and should not be classified as taxonomic groups.3
Wolves are now frequently classified as ecotypes, which sometimes confuses taxonomy.4
Species
A very important level of taxonomy is the species. Species are classified in two related, but subtly different, ways: by phylogeny (i.e., ancestry) and by reproductive isolation. There is a huge literature on what is known as the “species problem” in biology. The problem is that because closely related groups may interbreed, identification of species as distinct groups is not always possible. To identify species, it must be determined if two populations in different geographic areas share common ancestry and can interbreed. This is complicated because populations may interbreed, but the offspring may be inviable, infertile, or have reduced fitness. For example, horses (Equus caballus) and donkeys (Equus asinus) can interbreed but produce sterile mules as offspring. Horses and donkeys are considered different species because of this incompatibility. In some cases, wild populations have non-overlapping ranges and interbreeding is not testable. This leads to uncertain species designations. For example, if there are two wolf populations in different geographic areas that differ in size or coat color, should they be one species or two? The problem of uncertain species designations is succinctly described by natural historian J. C. Avise as: “In intermediate situations (and also in hybrid settings), educated nomenclatural judgments will remain necessary at species and subspecies levels.”5
Species is an important taxonomic level because species are groups that are separate and will remain so. If two populations are isolated and then reconnect and interbreed freely, then they are not species but simply populations of the same species. Different species concepts are used by scientists, and include the “biological species concept” (BSC) in which reproductive isolation defines species. BSC defines species as groups of interbreeding natural populations that are reproductively isolated from other such groups.6
The BSC is the most commonly used species concept. Another species concept is known as the “phylogenetic species concept” (PSC) and considers a species to be a group with an independent ancestry, without consideration of reproductive isolation. Under the PSC, groups that share a most recent common ancestor are considered members of the same species. The PSC is most useful for very distinct groups (e.g., wolves and foxes), and not for closely related groups (e.g., wolves and coyotes). Regardless of how they are designated, species are considered groups that are now, and will remain, separate. Clear identification of species definition is important with regard to wolves because of the designations of the eastern wolf and red wolf as species in a recent review by FWS biologists7 and subsequent reassessments.8
Subspecies
As noted above, subspecies are groups classified below the species level, and two definitions are commonly used. According to evolutionary biologist Ernst Mayr: “A subspecies is an aggregate of phenotypically similar populations of a species inhabiting a geographic subdivision of the range of the species and differing taxonomically from other populations of the species.”9
Another interpretation by Avise and Ball: “Subspecies are groups of actually or potentially interbreeding populations, phylogenetically distinguishable from, but reproductively compatible with other such groups. Importantly, the evidence for phylogenetic distinction must normally come from the concordant distributions of multiple, independent, genetically based traits.”10
These technical definitions simply mean that there must be genetically based traits that differ between populations for them to qualify as subspecies. However, the fact that subspecies are members of the same species and can interbreed results in subspecies that share genes and hence are not definitive groups. The scientific community recognizes the subjectivity of subspecies, as exemplified in Avise’s quote above and the following, first by Mayr, who stated that “This concept of the subspecies is fallacious. . . . The better the geographic variation of a species is known, the more difficult it becomes to delimit subspecies and the more obvious it becomes that many such delimitations are quite arbitrary.”11
In another view, scientist Paulo Vanzolini noted that “ . . . present applications of the subspecies concept are uneven, frequently undocumented, and lead to no improvement of either evolutionary theory or practical taxonomy.”12
Biologist Douglas Futuyma noted that there is so much variation among populations of most species that some combination of characters will distinguish each population from others and, consequently, there is no clear limit to the number of subspecies that can be recognized.13
Paul Ehrlich stated that “Widespread species thus can be divided into any number of different sets of ‘subspecies’ simply by selecting different characteristics on which to base them . . . as is the case with other species, geographic variation in human beings does not allow Homo sapiens to be divided into natural evolutionary units. That basic point . . . has subsequently been demonstrated in a variety of organisms . . . and use of the subspecies (or race) concept has essentially disappeared from the mainstream evolutionary literature.”14
Bart O’Gara observed: “Classification below the species level often has been subjective because there are no standard criteria for naming subspecies or populations . . . subspecies designations should be based on phylogenetic relationships, the same for species. In practice, they seldom are.”15
Robert Zink indicated that “Mitochondrial DNA sequence data reveal that 97 percent of . . . avian subspecies lack the population genetic structure indicative of a distinct evolutionary unit. . . . A massive reorganization of classifications is required so that the lowest ranks, be they species or subspecies, reflect evolutionary diversity. Until such reorganization is accomplished, the subspecies rank will continue to hinder progress in taxonomy, evolutionary studies and . . . conservation.”16
And biologists Haig et al. stated: “Among taxonomists, definitions of subspecies are a source of considerable disagreement . . . In an extensive literature review, we found no universally accepted subspecies definition within or across taxa . . . the scientific community has some level of comfort with the subjective nature of subspecies classification.”17
I provide these quotes to emphasize that the scientific community acknowledges that subspecies is not a rigorous scientific category and there are many different interpretations. However the subspecies concept can be useful to describe populations in geographic areas that differ from others, as long as the inherent subjectivity of the category is acknowledged. In addition, subspecies can be listed as “endangered species” under the ESA, so there is a practical need to deal with the category. Because subspecies are subjective, it is apparent that neither side in debates over subspecies designations is right or wrong in an absolute sense.18 However, one of the primary problems with the ESA is the designation of subspecies by the FWS, without adequate acknowledgement of the inherent subjectivity of the category. This has resulted in many management and legal controversies.
Populations
The problems with taxonomic uncertainty and the ESA become even more apparent at the population level, which is also included in the Endangered Species Act definition of species. Scientifically, a population is considered a group of interbreeding animals in a specific geographic area. It’s a general term and can be applied to a local area (e.g., the Yellowstone National Park population of wolves) or an entire continent (e.g., the North American population of wolves). However, the ESA includes in its species definition “distinct population segments” (DPS) which are defined as “populations that are discrete from other populations and significant to the species to which it belongs.”
There are no truly empirical criteria on what constitutes a discrete or significant population, so like subspecies, DPS designations are quite subjective.
Hybrids
As the previous chapter has described, another category that is frequently encountered in wolf taxonomy is hybrids. Hybrids are the offspring of parents of different breeds, varieties, subspecies, or species. It’s a general term, but the criterion for designation of a hybrid is that the parents are of different named groups. For example, a mule is an interspecies hybrid of horse and donkey parents. Wolf researchers have begun to consider both interspecies and intra-species hybrids. For example, interspecies wolf-dog and wolf-coyote hybrids and intra-species (inter-subspecies) Mexican wolf-northern wolf hybrids are current topics of research.20 Because subspecies are indefinite as discussed above, hybrids of two subspecies will likewise be indefinite. A supporter of a subspecies designation will consider mixed offspring to be hybrids, while someone who doesn’t support a subspecies designation will not consider mixed offspring to be hybrids, and a third group will simply consider them the products of dispersal and interbreeding among populations. Such differing perspectives can lead to political controversies regarding a litany of entities for the ESA.
Genetics, DNA, and Taxonomic Inflation
Traditionally, species and subspecies have been identified with morphology (i.e., anatomy), including body size, skull and bone proportions, coat color, or other characteristics. During the last half of the twentieth century, genetic technology (i.e., biotechnology) advanced to where DNA could be compared between animals. This is highly technical work, but the basic idea is that similarity of DNA reflects recent common ancestry, and hence can shed light on taxonomic relationships.
There are many technical approaches for using DNA to compare species, subspecies, and populations. One approach is to compare the proportions of gene variants (called “alleles”) in populations. For example, if there are alleles for a gene controlling black and white coat color in wolf populations, the proportion of black and white alleles in each population can be compared and the level of interbreeding between the populations can be estimated. Another approach involves comparing actual DNA molecules between groups. In this approach, the number of differences (i.e., mutations) in the DNA sequence (which is composed of a series of four molecules called nucleotides or base pairs, designated A, G, C, and T) can be counted, and the relationship of the DNA sequences determined. This can then be used to infer the relationship, and even the time of divergence, of the groups. This is straightforward for very divergent groups (for example, dogs and cats) but for related groups it is complicated because they often share the same DNA sequences. Both of these approaches have been used in wolf taxonomy.
The use of genetics and DNA over the last fifty years has allowed better resolution of variation in wildlife populations. However, it has also been accompanied by a decided tendency to split groups into more species and subspecies. This has been termed “taxonomic inflation” and its impetus is often to elevate the status of groups for conservation.22 For example, there has been an increase in elevating subspecies of wildlife to species status to enhance their chances of legal protection with laws such as the ESA.23 The designation of the eastern gray wolf as a distinct species, Canis lyacon described below, is an example of taxonomic inflation that is relevant to the ESA status of wolves in North America. Taxonomic inflation reflects the current scientific tendency of extensive splitting of animals into species, subspecies, and populations. I believe this tendency springs from three primary factors: the recent availability of DNA technology that allows detailed comparison of populations, peoples’ inherent tendency to classify animals into discrete groups, and the current focus on conservation and the use of the ESA to protect wildlife. This has led to acceptance of many new species, subspecies, and DPS without full acknowledgement of the scientific uncertainty of such designations, and has allowed science to be influenced by political and legal maneuvering.24
Wolf Species and Subspecies
Several scientific papers on wolf species and subspecies taxonomy have been published since the first edition of The Real Wolf. These studies have used new DNA technologies developed in dogs that allow assessment of large amounts of data covering the entire genome. The dog genome contains approximately 2.5 billion DNA base pairs (also known as nucleotide pairs) which is comparable in size to the genomes of other mammals, including humans. The dog is used as a model for genetic diseases, the genetics of behavior, and in pharmaceutical research.25
Analysis of the entire genome allows identification of differences in DNA sequences called single nucleotide polymorphisms (SNP). This technology is now used extensively in human medical genetics and dog and livestock breeding programs. Because of the close relationship of dogs and wolves this technology can be applied to study of wolf genetics. Here I will briefly review the status of wolf species and subspecies in light of these new data.
Wolf Species
In addition to wolves, the canid family (called the Canidae) also includes foxes, coyotes, jackals, dingoes, African wild dogs, and domestic dogs (Canis familiaris). There are wild canids on every continent except Antarctica. Both wolves and jackals were thought to be the progenitors of domestic dogs, but genetics has now shown that dogs were domesticated exclusively from wolves in Europe and Asia within the last fifteen thousand years or so. Recently, Fan and colleagues analyzed relationships of wolves and dogs with a large data set of thirty-four canine genome sequences that indicate that the dog was domesticated in Eurasia from ancestral wolves.26 The same research team previously also have found extensive admixture between dogs and wolves, with up to 25 percent of Eurasian wolf genomes showing signs of dog ancestry.27
Fossils suggest that the genus Canis arose in North America more than three million years ago during the Pliocene Epoch. They spread to Eurasia and Africa and differentiated into jackals: side-striped jackal (Canis adustus), golden jackal (Canis aureus), black-backed jackal (Canis mesomelas), gray wolves (Canis lupus), and the Ethiopian wolf (Canis simensis). Another idea is that gray wolves originated in Africa and then spread to the northern hemisphere. Coyotes (Canis latrans) developed from the original Canis and have only occurred in North America.
Gray wolves are thought to have originated about five hundred thousand years ago and migrated from Eurasia to North America during the Pleistocene epoch. This was possible because the sea level dropped during glacial periods exposing the Bering land bridge between Asia and Alaska. It has been suggested that gray wolves entered North America at least three different times, with the new migrants mixing with or displacing the existing populations.
There are different views of wolf species in North America. In one view, there is one species of wolf in North America, and some geographic areas contain subspecies. In another, wolves have been classified as two species: the gray wolf that ranges across most of the continent, and the red wolf (Canis rufus) that inhabits the southeastern United States.28
Some taxonomists consider the red wolf a subspecies of gray wolf (Canis lupus rufus) and not a full species. Red wolves were extirpated in the wild, and now exist only in captive populations and an introduced population on an island in North Carolina. A third view developed amongst taxonomists recently identifies a third species that has been designated as the eastern wolf (Canis lycaon) in the Canadian provinces of southern Quebec and Ontario and in the northeastern United States as far west as Minnesota.29
The eastern wolf is also considered a subspecies of gray wolf (Canis lupus lycaon) (i.e., not a full species) by some taxonomists.30
The species designations of the eastern wolf and red wolf are based on their apparent common ancestry with coyotes in North America, separate from gray wolves in Eurasia.31
The assessment is uncertain because of the extirpation of wolves from much of their North American range and recent interbreeding between gray wolves, the proposed eastern wolf, and coyotes, as well as interbreeding between red wolves and coyotes. This greatly complicates the genetic patterns of living populations. Also, the species designations were based on a limited number of genes and an uncertain number of samples from pure red wolves and eastern wolves. In my opinion, this makes the species designation of an eastern wolf premature and unwarranted. A recent study of a large genetic data set described below supports this view and that there is only one species of wolf in North America, the gray wolf Canis lupus.32
The eastern wolf and red wolf species designations are based on the theory that these two groups and coyotes all evolved in North America from a common ancestor. It is proposed that gray wolves evolved independently in Eurasia and then entered North America where they came into contact with the coyote, eastern wolf, and red wolf. The fact that these four groups (gray wolf, eastern wolf, red wolf, and coyote) interbreed makes the historical reconstruction with DNA data very complex and speculative. Gray wolves and coyotes don’t typically interbreed where they co-occur, but they may in areas where coyotes expanded their range, and where wolves are rare (like the northeast United States and southeast Canada). In these areas it is believed that coyotes have hybridized with wolves (red, gray, and eastern) within the last hundred years, making the genetic composition of each group mixed to varying degrees.
It’s important to note that until recently the red wolf and eastern wolf species designations were based on limited data. The primary supporting data is from skull measurements and short sequences of mitochondrial DNA (mtDNA) and Y-chromosome DNA. These data are not in agreement with an extensive DNA assessment (forty-eight thousand single nucleotide polymorphisms, called SNP)33 that show that the eastern wolf is genetically more similar to gray wolves than coyotes, and does not support the eastern wolf as a distinct species. However, a reanalysis of the data suggested that the eastern wolf may still be a legitimate species.34
The analyses of two studies are consistent in showing that red wolves are more similar to coyotes than to gray wolves. Because the genetic patterns of both red wolves and eastern wolves reflect recent interbreeding between various groups of wolves and coyotes, the taxonomy of the groups was still unclear following these studies.35, 36
More recent studies provide additional insights. A study by Rutledge and colleagues37 using genomic data for 127,235 SNP found the eastern wolf is a distinct genetic group, and supported the recognition of three species of Canis in North America (gray wolf, eastern wolf, and coyote). This view considers the red wolf to be the same species as the eastern wolf. Rutledge et al. support recognition of the eastern wolf as a separate species, but note that they have hybridized with gray wolves and coyotes in some areas.
In contrast, vonHoldt and colleagues used twenty-eight genome sequences to derive 5,424,934 SNP with which to compare wolves and coyotes.38 These extensive data showed that the red wolf and eastern wolf are hybrid populations of the gray wolf and coyote. This study showed what many have thought all along, that there is one species of wolf in North America (the gray wolf) and they have interbred with coyotes in some areas. My research involving 123,801 SNP also showed that coyotes in the northeast United States (Maine and Connecticut) have a degree of wolf ancestry, probably from hybridization.39
Further, vonHoldt and colleagues claim their data make removing the wolf from the ESA list invalid, and that the hybrid populations have unique genetics of wolves worthy of conservation and ecological processes (i.e., selection) that can be allowed to restore historical patterns of genetic variation.
The lack of consensus of the species status of wolves in North America indicates there is a need for a taxonomic assessment with full consideration of species concepts and definitions, additional genetic data, and the study of fossils. In regards to the ESA, there is a need to either determine consensus about taxonomy or clarify the law regarding the definition of species. In addition, comparisons should be made with other species groups for which DNA data are not concordant with species designations.40
This includes white-tailed deer (Odocoileus virginianus), mule deer (Odocoileus hemionus hemionus), black-tailed deer (Odocoileus hemionus sitkensis), brown bears (Ursus arctos), and polar bears (Ursus maritimus). In the case of the deer, mtDNA is more similar between white-tailed deer and mule deer (different species) than between mule deer and black-tailed deer (the same species). In the case of the bears, mtDNA is more similar between polar bears and one population of brown bears in southeast Alaska (different species) than between the southeast Alaska brown bears and other brown bears (the same species). It is well known among scientists that mtDNA relationships may not reflect species relationships.41
In both of these cases, other DNA data shows concordance with the species designations (i.e., white-tailed deer are different from mule deer and black-tailed deer, and all brown bears are different from polar bears). Comparisons of wolf genetics with others such as these will provide insights for wolf taxonomy.
Wolf Subspecies
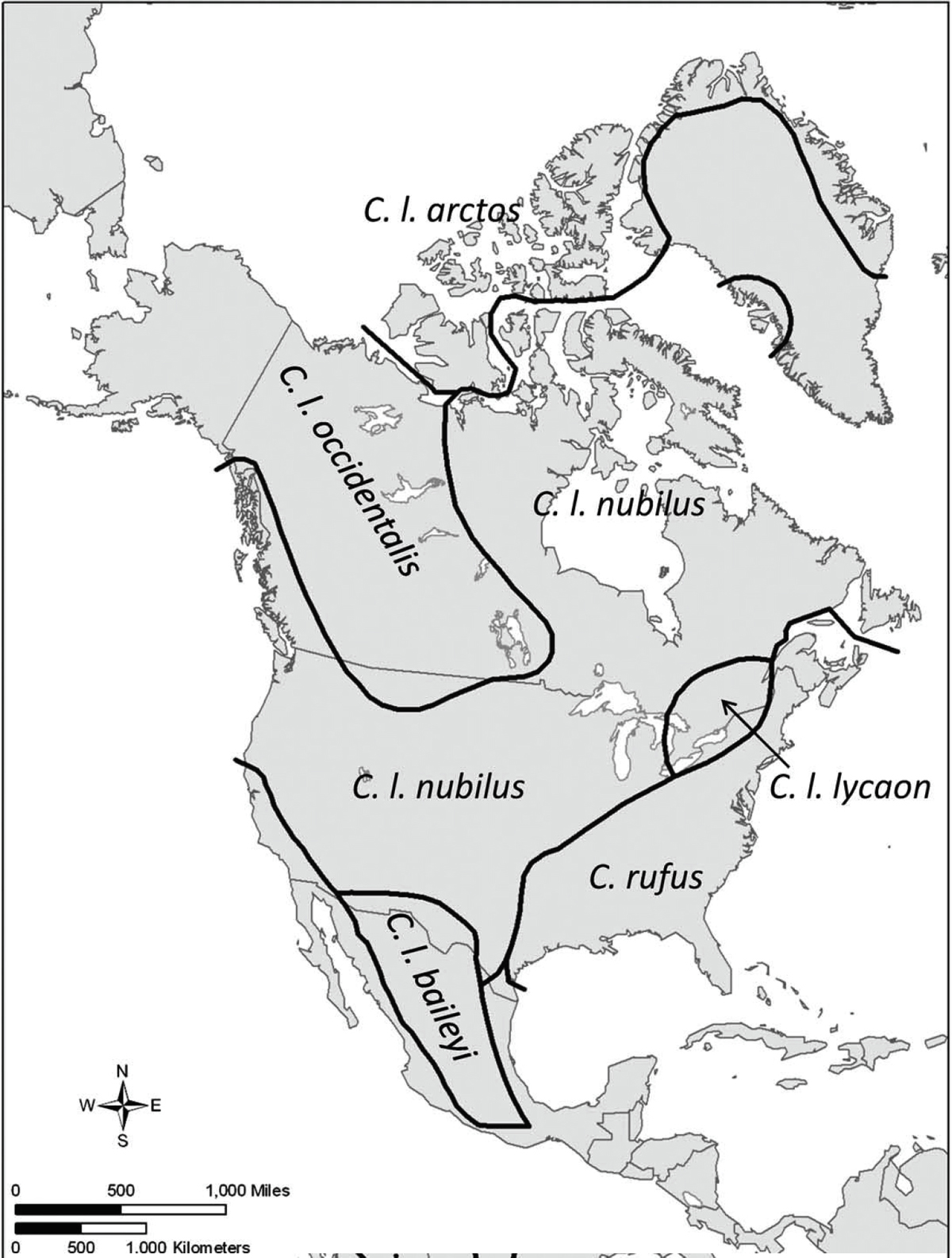
Many subspecies of wolves were recognized during the 1900s based on morphology, particularly skull measurements. The primary classification recognized twenty-three subspecies of gray wolves in North America,42 although up to twenty-seven subspecies were recognized.43 This was later reduced to five subspecies including the Mexican wolf (Canis lupus baileyi), the northern timber wolf (Canis lupus occidentalis), the plains wolf (Canis lupus nubilus), the eastern wolf (Canis lupus lycaon), and the Arctic wolf (Canis lupus arctos) whose ranges are shown in Figure 1. The red wolf was designated a different species in these analyses.
Chambers and colleagues reviewed these five gray wolf subspecies with morphological and genetic data.45 They found the existing data support designation of the Mexican wolf, northern timber wolf, and plains wolf subspecies. They also found that the data for the Arctic wolf is not definitive, and that the eastern wolf is a full species, not a subspecies. This work was prior to the recent recognition of the eastern wolf as a gray wolf-coyote hybrid by vonHoldt and colleagues, so their recognition of the eastern wolf as a species may change.46
The primary support of the three subspecies of gray wolf recognized by Chambers and colleagues. The Mexican wolf, northern timber wolf, and plains wolf include the skull measurements of Goldman47 and Nowak48 and genetic data showing limited interbreeding among them. The data indicate a degree of differentiation of the proposed subspecies, but the assessment is complex because data come from different studies with different sets of samples. Also, the subspecies range boundaries are uncertain so there was likely interbreeding across the original ranges of the different subspecies. The near extinction of the plains wolf and Mexican wolf subspecies make attempts to characterize their genetic makeup particularly problematic because the living populations from which samples can be obtained may not represent the original genetic patterns. North American wolf subspecies in general are questionable and have been described as arbitrary, typological, and an intergrading series of populations.49
Mexican Wolf
The case of the Mexican wolf (C. l. baileyi) warrants special discussion because it is listed as an endangered subspecies under the ESA.
My data50 and that of vonHoldt and associates51 show SNP differentiation of Mexican wolves from other North American wolves. However, living and historic samples show that Mexican wolves share mtDNA haplotypes with wolves in other areas and with coyotes and the living Mexican wolves came from only seven founders of a captive breeding population.52 In addition, the living Mexican wolves may have included dog ancestry.53 This finding is supported by a large genome study that showed 1 percent to 3 percent of the Mexican wolf genome has dog ancestry.54
I’ve pointed out that these factors indicate that designation of a Mexican wolf subspecies is questionable in my wolf genetic study published in 2015.50 However, a study conducted by Fredrickson et al.56 disagreed and claimed the Mexican wolf is a legitimate subspecies, despite additional arguments to the contrary.57 Fan and colleagues also showed Mexican wolves as “a divergent form of gray wolf” compared with gray wolves from Yellowstone National Park (which originated with transplants from Alberta and British Columbia, Canada) and claimed their data contradicted my suggestion58 that the Mexican wolf was not a distinct subspecies.59
In claiming subspecies status of the Mexican wolf, Fan and Fredrickson60 missed the primary point of my colleagues and I made as indicated in the quotations about subspecies above61 that the subspecies category is subjective. They also do not consider the importance of the point we made that living Mexican wolves are derived from only seven founding animals that are not representative of the native population of wolves in Mexico and the southwest United States.62
Other recent studies are relevant to Mexican wolf classification. Hendricks analyzed mtDNA and four autosome SNP loci in wolves in the southwest United States and Mexico including an historical sample (collected in 1922) in San Bernardino, California, and mtDNA in the Pacific Northwest United States.63 The San Bernardino sample was deemed to have Mexican wolf ancestry based on the genetic data. They also modeled Mexican wolf habitat and potential distribution. The findings state that these data support expanding the historical range of the Mexican wolf subspecies and Pacific Northwest coast ecotype for recolonizing wolf populations.
However, this assessment has the potential problem of promoting typological thinking,64 which is inherent in subspecies designations.65 Typological thinking in biology assumes a morphological “type” for a species or subspecies, which is in conflict with “population thinking,” and in which variation among individuals, not a fixed type, is the actual state of natural populations. In this case, an mtDNA haplotype and SNP alleles found in Mexican wolves were found in the San Bernardino wolf so it was considered of Mexican wolf ancestry.66
I think it is more appropriate to apply population thinking to these cases and consider the dynamic nature of populations’ ranges expanding and contracting with dispersal and interbreeding among areas rather than associate a typological subspecies name with genetic markers.67
It should be obvious that there are many varying opinions on classification. Ecologists Wayne and Schaffer, and their associates, for example, discuss the issue of hybrids and ESA protection and provide a decision-tree framework for evaluating hybrid protection, including the processes that produced hybrids and the ecological impacts of hybrids.68 They suggest that hybrid populations may have value and warrant protection. As examples, they discuss the Mexican wolf, red wolf, and eastern wolf, noting that Mexican wolves had historic admixture (i.e., hybridized) with wolves to the north and that such hybrids should be protected.
At this point, it’s necessary to raise a basic question regarding subspecies in this case: If the Mexican wolf is a genetically distinct subspecies as Fan and Fredrickson maintain,69 but historically mixed with wolves to the north, how can it be distinct?70
The Mexican wolf has been variously termed a subspecies, a population, an ecotype, an evolutionarily significant unit (ESU), a genetically distinct subspecies, and “the most endangered and . . . genetically distinct wolf subspecies in the New World,” in the wolf genetics papers cited in this chapter, such as Hendricks and colleagues,71 the Mexican wolf also had a “large zone of intergradation” with the subspecies to the north (C. l. nubilus),72 and historic admixture with other subspecies, according to several researchers such as Vila, Frederickson, Hendricks, and Wayne and Schaffer, and their associates maintain.73 How can the Mexican wolf be genetically distinct and also mix and intergrade with other wolves? Wolves in areas of admixture will not be as genetically distinct from the neighboring subspecies as are wolves in the center of the range. This apparent contradiction shows how typological subspecies confuse understanding of the reality of populations that are dynamic in space and time.
This issue doesn’t need to be so complicated. A simple solution for conservation and wildlife management is to not use subspecies, but to refer to wolves and other wildlife as populations with geographic information as descriptors. This approach has been advocated by a number of researchers.74 For example, Mexican wolves can be referred to as the population of wolves (Canis lupus) in Mexico and the southwest US management objectives can then be applied to the population without the uncertainties of subspecies.
Southeast Alaska Wolf
The subspecies classification proposed by Goldman and Hall include another subspecies of gray wolf in southeast Alaska and coastal British Columbia, the Alexander Archipelago wolf (Canis lupus ligoni).75 However, both Nowak and Chambers and their respective colleagues76 consider these wolves to be part of the plains wolf subspecies (Canis lupus nubilus). Figure 1 shows the range of the plains wolf subspecies including the coastal region of British Columbia and southeast Alaska. Indeed, wolves from this area share DNA genotypes with wolves from the late 1800s from Kansas and Nebraska (sampled from museum specimens). This group of wolves is thought to have colonized from the western United States northward to coastal British Columbia and southeast Alaska following the melting of the continental glaciers within the last fifteen thousand years.
My recent study assessed genome variation of dogs, wolves, and coyotes in several parts of North America, including southeast Alaska.77 We determined genotypes of 305 wolves for 123,801 SNP loci, and found varying levels of differentiation, including low differentiation of wolves in interior Alaska, British Columbia, and the northern US Rocky Mountains. There is considerable differentiation of wolves in Southeast Alaska from wolves in other areas, but wolves in Southeast Alaska are not a genetically homogeneous group and there are comparable levels of genetic differentiation among areas within Southeast Alaska and between Southeast Alaska and other geographic areas. Considering these data, and the analyses of Chambers and associates, we suggested the wolves in southeast Alaska do not warrant subspecies designation.78
Figure 1. Ranges of the North American gray wolf (Canis lupus) and red wolf (Canis rufus) recognized by Nowak (1995, 2002). The subspecies are Mexican wolf (Canis lupus baileyi), northern timber wolf (Canis lupus occidentalis), plains wolf (Canis lupus nubilus), eastern wolf (Canis lupus lycaon), and Arctic wolf (Canis lupus arctos). The eastern wolf is recognized as a species (Canis lycaon) by some authors. This is Figure 3 in Chambers’s paper, which is a publication of the US Fish and Wildlife Service and includes the following statement: All material appearing in North American Fauna is in the public domain and may be reproduced or copied without permission unless specifically noted with the copyright symbol ©.
Weckworth and associates, however, disagree.79 They did not explicitly state that they support a subspecies designation, but they say that recognizing the wolves as a subspecies or a set of genetically distinct populations is important for conservation. My team and I responded, noting the subjectivity of subspecies and the need to manage populations regardless of terminology.80
The wolf in southeast Alaska is important because the USFWS considered the ligoni subspecies for listing as an endangered species under the ESA. Acknowledging the lack of consensus in the scientific literature, USFWS noted that ligoni may or may not be a subspecies, but considered ligoni to be a subspecies for their ESA decision. However, USFWS did not list it because they found that the wolf in southeast Alaska is neither threatened nor endangered with extinction.81 The finding by Chambers and associates (who are USFWS biologists) that ligoni is not a legitimate subspecies, and the finding that it is a legitimate subspecies in the ESA ruling, is scientific inconsistency within the USFWS, however Chambers and associates’ opinion was prior to the ruling of the ESA.82
Another interesting facet of the subspecies designations is the introduction of wolves from the range of the northern timber wolf subspecies (Canis lupus occidentalis) into the northern US Rocky Mountains including Yellowstone National Park, which is within the original range of the Great Plains wolf (Canis lupus nubilus). These wolves were introduced (and listed as a distinct population segment under the ESA), apparently without concern for the subspecies of origin. This situation may well become more complex if the wolves in the northern US Rockies expand southward and come into contact and interbreed with the Mexican wolf subspecies in New Mexico and Arizona. There are already some reports of wolves in Colorado that are from the Northern Rockies population of gray wolves.
The northern timber wolf subspecies that has been introduced into the Northern Rockies is a large wolf from western Canada and Alaska. Skull and body sizes indicate that the northern timber wolf is larger than the Great Plains wolf. For example, plains wolf males weigh ninety-five to ninety-nine pounds, and northern wolf males weigh eighty-five to 115 pounds, occasionally reaching 130 pounds. A male northern wolf caught in Alaska in 1939 weighed 175 pounds.
The data showing the northern timber wolf subspecies is generally larger than the Great Plains wolf subspecies is consistent with reports that the introduced wolves in the northern US Rockies are considerably larger than the native Great Plains wolves. In Yellowstone National Park, one male wolf has been caught that weighed at 143 pounds with a full stomach and another weighed 147 pounds with an empty stomach. However, it is important to recognize that we do not know if the size differences of wolves in these areas is due to heritable (i.e., genetic) or environmental (e.g., good nutrition) causes.
In summary, these observations indicate that wolf subspecies are uncertain. A simple solution for conservation and management is to designate populations by geographic area instead of typological and subjective subspecies.83 This has practical applications for several ESA cases in addition to the wolf. Recent research indicates that several subspecies listed under the ESA are not supported by genetic data, including the coastal California gnatcatcher (Polioptila californica californica),84 the southwestern willow flycatcher (Empidonax traillii extimus),85 the Preble’s meadow jumping mouse (Zapus hudsonius preblei),86 the wood bison (Bison bison athabascae),87 and the wolf subspecies we’ve already discussed. This indicates there is unwarranted taxonomic inflation of wildlife subspecies designations similar to unwarranted species designations.88 For wildlife management the traditional use of geographic populations as management units is more appropriate than subspecies.
Ecotypes
Schweitzer and associates assessed genome variation in gray wolves to identify genes under selection and local adaptation in six geographic regions (called ecotypes).89 Their analysis identified potential local adaptation in different populations related to seasonality of precipitation and vegetation. They further found that Arctic and High Arctic wolf ecotypes have several genes under selection, and discuss their conservation value in light of climate change.90 This is a rigorous study, and is similar to studies of genes underlying performance traits in livestock. A key point, however, is that studying adaptive variation due to selection is not the basis of taxonomy and my focus in this chapter. The “ecotypes” reflect local adaptation (or environmental influence) and not necessarily ancestry. This can result in what is called homoplasy (or convergence) in taxonomy, in which populations share traits due to common selection pressure, not ancestry. This is a highly technical subject, but important to consider when classifying animals.
As noted above, “population” is a general term, and wolf populations can be designated at any scale. For our discussion, the population category used in the ESA, distinct population segment (DPS), is relevant. A population must be discrete and significant to be considered a DPS. Discreteness can be simple geographic separation or a combination of factors, including genetic differentiation. Significance reflects the importance of the population to the “taxon to which it belongs” which can be the entire species or a subspecies. Significance is usually a subjective judgment by USFWS or NMFS. Despite Congress’s order that DPS be used “sparingly” under the ESA, the USFWS and NMFS has listed many DPS including wolf populations.91
In the case of wolves, the introduced population in the northern US Rocky Mountains (Montana, Wyoming, and Idaho) and the population in the western Great Lakes region (Minnesota, Wisconsin, and Michigan) were each listed as a DPS under the ESA. The USFWS hopes to remove the gray wolf from the list of endangered species in the contiguous United States as it has already exceeded population numbers originally established to determine its establishment; however, there has been a continual series of courtroom battles about this.
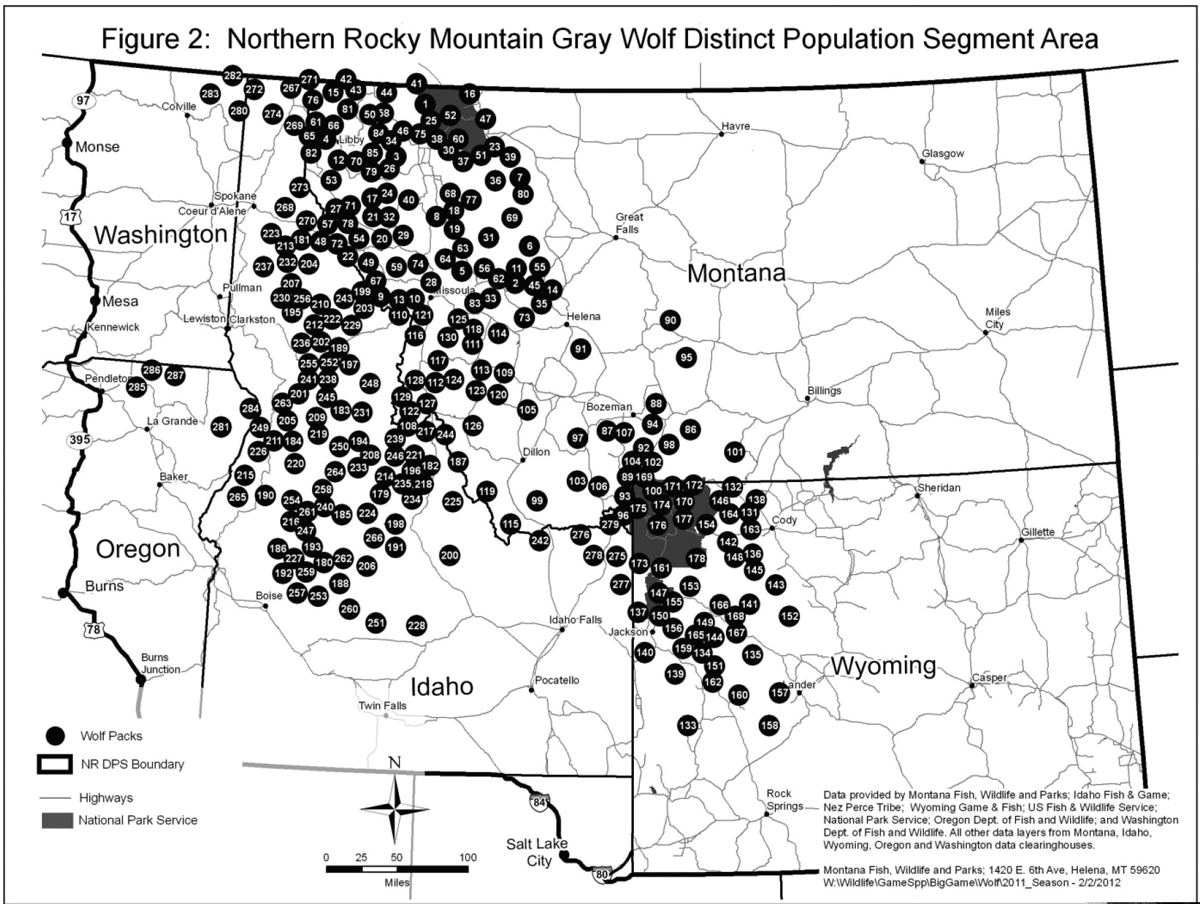
Endangered Species Act protections for gray wolves in Montana, Wyoming, and Idaho were removed in 2011 (Figure 2). The wolves have spread into Washington, Oregon, California, Utah, and Colorado (Figure 3).
In September 2014, the Federal District Court for Washington, DC, vacated the delisting of wolves in Wyoming under the ESA. Therefore, these wolves were again listed as a nonessential experimental population in all of Wyoming. However, on December 19, 2014, following two court orders, the USFWS reinstated regulatory protections under the ESA for the gray wolf in Wyoming and the western Great Lakes on February 20, 2015.
On November 9, 2015, the Oregon Fish and Wildlife Commission delisted wolves from protection under the Oregon Endangered Species Act.
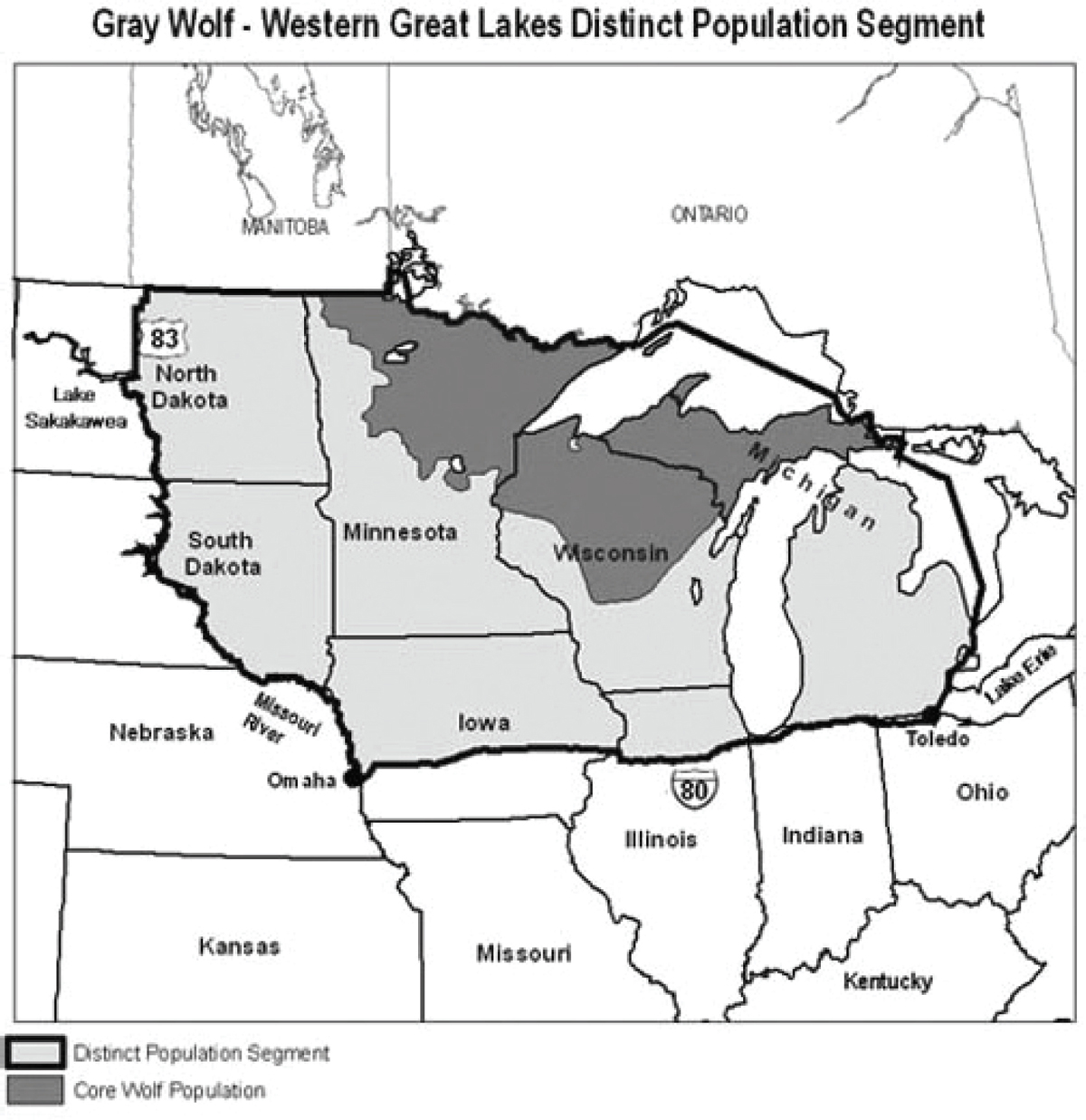
Wolves in Minnesota, Wisconsin, and Michigan—the Great Lakes population—have now spread into North and South Dakota, Iowa, Illinois, Nebraska, and Ontario. Wolves were delisted in the Great Lakes States in 2012. However, as a result of a federal court decision, wolves in the western Great Lakes area (including Michigan, Minnesota, and Wisconsin) were relisted under the Endangered Species Act, effective December 19, 2014.
This review indicates that the current state of North American wolf taxonomy (and wolf ESA listings) is very confusing and not definitive. However, the management of wolves in the United States, first under the federal ESA and then by States after ESA delisting, can be influenced by taxonomic assessments. Both populations in the northern US Rocky Mountains and western Great Lakes region have been managed with considerable population increases. However, both populations are of questionable taxonomic status. As noted above, the wolves in the northern US Rocky Mountains were transplants of the northern timber wolf subspecies into the historic range of the plains wolf subspecies.
Figure 2. Wolf distribution in the northern US Rocky Mountains. From the US Fish and Wildlife Service website (15 August 2013), http://www.fws.gov/mountain-prairie/species/mammals/wolf/20120828MapPackDistributionFigure%202.jpg.
Figure 3. Wolf distribution in the Great Lakes States. From the US Fish and Wildlife Service website (15 August 2013), http://www.fws.gov/midwest/wolf/delisting/WGLDPS.html.
The wolves in the western Great Lakes states are within the range of the plains wolf subspecies92 but are also considered the eastern wolf species and there has been mixing of wolves and coyotes in this area.93 There are also inconsistencies in which subspecies are considered important in some cases (e.g., the Mexican wolf), but subspecies designations are ignored in others (e.g., introducing the non-native northern timber wolf subspecies into the northern US Rocky Mountains).
A number of colleagues and I contend that for wildlife management it is most important to recognize that the wolves in the northern US Rocky Mountain and western Great Lakes regions fill the ecological role of a large predator, regardless of opinions regarding species, subspecies, and DPS. Extensive effort and resources can be wasted debating wolf species and subspecies status and saving “pure” species or subspecies, instead of focusing on practical wildlife management.94
It’s perhaps most important that taxonomy should be practiced as a science by taxonomists and not by wildlife biologists, and the inherent uncertainty associated with subspecies, DPS, and some species designations should be openly and widely acknowledged. In particular, the USFWS should not favor one taxonomic opinion on species, subspecies, or DPS designation, and ignore equally valid differing opinions. Considering all of these points, my conclusion is that wolf management and wildlife management in general, including implementation of the ESA, should focus on populations in specific geographic areas, without being shackled by unresolvable debate over species, subspecies, and DPS taxonomy.
As my colleagues and I stated in our genetic study in 2015: “For wildlife management, the traditional use of geographic populations as management units . . . is more appropriate than subspecies. The scientific rigor of population genetics, systematics, and taxonomy, and their application to management and conservation, would be enhanced by implementation of this practice.”95
Endnotes:
1. NRC (National Research Council). 1995. Science and the Endangered Species Act. National Research Council, Committee on Scientific Issues in the Endangered Species Act. National Academy Press, Washington, D.C., USA.
2. Darwin, C. 1859. On the Origin of Species. Murray, London.
3. Cronin, M.A. 2006. “A Proposal to eliminate redundant terminology for intra-species groups.” Wildlife Society Bulletin 34:237–241. And Cronin, M. A., M. D. MacNeil, and J. C. Patton. 2005. “Variation in mitochondrial DNA and microsatellite DNA in caribou (Rangifer tarandus) in North America.” Journal of Mammalogy 86:495–505.
4. Cronin, M.A. and L.D. Mech. 2009. “Problems with the claim of ecotype and taxon status of the wolf in the Great Lakes region.” Molecular Ecology 18:4991–4993.
5. Avise, J.C. 2000. Phylogeography, the History and Formation of Species. Harvard University Press, Cambridge, Massachusetts, USA.
6. Mayr, 1970, Populations, Species, and Evolution. Belknap Press, Harvard University Press, Cambridge, MA, USA. And Mayr, E. 1982. The Growth of Biological Thought. Cambridge MA: Harvard University Press.
7. Chambers et al. 2012
8. Rutledge et al. 2015, vonHoldt et al. 2016
9. Mayr, E. 1970.OP CIT
10. Avise, J.C., and R.M. Ball Jr. 1990. “Principles of genealogical concordance in species concepts and biological taxonomy.” Oxford Survey of Evolutionary Biology 7:45–67.
11. Mayr (1970), OP. CIT
12. Vanzolini, P. E. 1992. Third world Museums and Biodiversity. Pages 185–198 in N. Eldredge, editor. Systematics, ecology, and the biodiversity crisis. Columbia University Press, New York, New York, USA.
13. Futuyma, D. J. 1986. Evolutionary Biology. Sinauer Associates, Sunderland, Massachusetts, USA.
14. Ehrlich, P. R. 2000. Human Natures. Island Press, Shearwater Books, Washington, D.C., USA.
15. O’Gara, B.W. 2002. “Taxonomy” Chapter 1, pages 3–65 in: North American Elk Ecology and Management. Compiled and edited by D. E. Toweill and J. W. Thomas. Smithsonian Institution Press, Washington D.C. and London.
16. Zink, R.M. 2004. The role of subspecies in obscuring avian biological diversity and misleading conservation policy. Proceeding of the Royal Society of London B. 271:561–564.
17. Haig, S.M., E.A. Beever, S.M. Chambers, H.M. Draheim, B.D. Dugger, S. Dunham, E. Elliott-Smith, J.B. Fontaine, D.C. Kesler, B.J. Knaus, I.F. Lopes, P. Loschi, T.D. Mullins, and L.M. Sheffield. 2006. “Taxonomic considerations in listing subspecies under the US Endangered Species Act.” Conservation Biology 20:1584–1594.
18. Cronin, M.A. 2007. “The Preble’s meadow jumping mouse: subjective subspecies, advocacy and management.” Animal Conservation 10:159–161.
19. Cronin 2006, OP. CIT.
20. vonHoldt, B.M., J.A. Cahill, Z. Fan, I. Gronau, J. Robinson, J.P. Pollinger, B. Shaprio, J. Wall, and R.K. Wayne. 2016. “Whole-genome sequence analysis shows that two endemic species of North American wolf are admixtures of the coyote and gray wolf.” Science Advances 2:doi: 10.1126/sciadv.1501714. (July 27, 2016).
21. Wayne, R.K. and H.B.Schaffer. 2016. “Hybridization and endangered species protection in the molecular era.” Molecular Ecology 25:2680–2689.
22. Zachos, R.E., M. Apollonio, E.V. Bärmann, M. Festa-Bianchet, U. Göhlich, J.C. Habel, E. Haring, L. Kruckenhauser, S. Lovari, A.D. McDevitt, C. Pertoldi, G.E. Rossner, M.R. Sanchez-Villagra, M. Scandura, and F. Suchentrunck. 2013. “Species inflation and taxonomic artefacts-A critical comment on recent trends in mammalian classification.” Mammalian Biology 78:1–6.
23. Marris, E. 2007. “The species and the specious.” Nature. 446:250- 253. and The Economist. 2007. Hail Linnaeus. Conservationists and polar bears should heed the lessons of economics.
24. Crandall, K.A. 2006. “Advocacy dressed up as scientific critique.” Animal Conservation 9:250–251. and Cronin 2007, OP. CIT.
25. https://www.genome.gov/12511476/2004-advisory-dog-genome-assembled/.
26. Fan, Z., P. Silva, I. Gronau, S. Wang, A. Serres Armero, R. M. Schweizer, O. Ramirez, J. Pollinger, M. Galaverni, D. Ortega Del-Vecchyo, L. Du, W. Zhang, Z. Zhang, J. Xing, C. Vilà, T. Marques-Bonet, R. Godinho, B. Yue, and R. K. Wayne. 2016. “Worldwide patterns of genomic variation and admixture in gray wolves.” Genome Research 26:163–173.
27. Ibid.
28. Nowak, R.M. 1995. “Another look at wolf taxonomy.” Pages 375–397 in Carbyn LN, Fritts SH, Seip DR, editors. Proceedings of the second North American symposium on wolves. Edmonton, Alberta: Canadian Circumpolar Institute, University of Alberta.
29. Chambers, S.M, S.R. Fain, B. Fazio, and M. Amaral. 2012. “An account of the taxonomy of North American wolves from morphological and genetic analyses.” North American Fauna 77:1–67. doi:10.3996/nafa.77.0001
30. Nowak 1995. OP CIT
31. reviewed by Chambers et al. 2012. OP CIT
32. vonHoldt et al. 2016. OP CIT.
33. Chambers et al. 2012. OP CIT
34. Rutledge, L.Y., P. J. Wilson, C. F.C. Klütsch, B. R. Patterson, and B.N. White. 2012. “Conservation genomics in perspective: A holistic approach to understanding Canis evolution in North America.” Biological Conservation 155:186–192.
35. Chambers et al. 2012. OP CIT
36. vonHoldt, B.M., J.P. Pollinger, D.A. Earl, J.C. Knowles, A.R. Boyko, H. Parker, E. Geffen, M. Pilot, W. Jedrzejewski, B. Jedrzejewski, V. Sidorovich, C. Creco, R. Ettore, M. Musiani, R. Kays, C.D. Bustamante, E.A. Ostrander, J. Novembre, and R.K. Wayne. 2011. “A genome-wide perspective on the evolutionary history of enigmatic wold-like canids.” Genome Research 21:1294–1305.
37. Rutledge, L.Y., S. Devillard, J.Q. Boone, P.A. Hohenlohe, and B.N. White. 2015. RAD sequencing and genomic simulations resolve hybrid origins within North American Canis. Biology Letters 11:20150303. http://dx.doi.org/10.1098/rsbl.2015.0303.
38. vonHoldt et al. 2016. OP CIT
39. Cronin, M.A., A. Cánovas, A. Islas-Trejo, D.L. Bannasch, A.M. Oberbauer, and J.F. Medrano. 2015a. “Single nucleotide polymorphism (SNP) variation of wolves (Canis lupus) in Southeast Alaska and comparison with wolves, dogs, and coyotes in North America.” The Journal of Heredity. 106:26–36.
40. Cronin, M.A. 1993. Mitochondrial DNA in wildlife taxonomy and conservation biology: cautionary notes. Wildlife Society Bulletin 21:339–348.
41. Cronin 1993. OP CIT (Cronin, 2006. OP CIT Zink 2004).
42. Goldman, E.A. 1944. Classification of wolves: part II. Pages 389–636 in Young SP, Goldman EA, editors. “The wolves of North America. Washington, D.C.: The American Wildlife Institute.” And Hall, E.R. 1981. The Mammals of North America, 2nd. Ed. John Wiley and Sons, New York.
43. Table 2 of Chambers et al. 2012. OP CIT.
44. Nowak 1995, 2002. OP CIT
45. Chambers et. al. 2012 OP CIT
46. vonHolt et. al. 2016 OP CIT
47. Goldman 1944 OP CIT
48. Nowak, 1995, 2002 OP CIT
49. Wayne, R.K. and C. Vilá. 2003. “Molecular genetic studies of wolves.” In: Mech LD, Boitani L, editors. Wolves: Behavior, Ecology, and Conservation. Chicago: University of Chicago Press. p. 218–238
50. Cronin et al. 2015a OP CIT
51. vonHoldt et al. 2011 OP CIT
52. Leonard, J.A., C. Villa, and R.K. Wayne. 2005. “Legacy lost: genetic variability and population size of extirpated US grey wolves (Canis lupus).” Molecular Ecology 14:9–17.; and Hailer, F. and J.A. Leonard. 2008. “Hybridization among three native North American Canis species in a region of natural sympatry.” PLOS ONE. 3(10), e333. doi: 10.1371/\ journal.pone.0003333.
53. García-Moreno, J., M.D. Matocq, R.S. Roy, E. Geffen, and R.K. Wayne. 1996. “Relationships and genetic purity of the endangered Mexican wolf based on analysis of microsatellite loci.” Conservation Biology 10:376–389. And Hedrick, P.W., P.S. Miller, E. Geffen, and R.K. Wayne. 1997. “Genetic evaluation of the three captive Mexican wolf lineages.” Zoo Biology 16:47–69.
54. Fan et al. 2016. OP CIT
55. Cronin et al. 2015a OP CIT
56. Fredrickson, R., P. Hedrick, R.K. Wayne, B.M. vonHoldt, and M. Phillips. 2015. “Mexican wolves are a valid subspecies and an appropriate conservation target.” Journal of Heredity 106: 415–416.
57. Cronin, M.A., A. Cánovas, A. Islas-Trejo, D.L. Bannasch, A.M. Oberbauer, and J.F. Medrano. 2015b. “Wolf Subspecies: Reply to Weckworth et al. and Fredrickson et al.” The Journal of Heredity 106:417–419.
58. Cronin et al. 2015a OP CIT
59. Fan et. al. 2016 OP CIT
60. Fredrickson et al. (2015. OP CIT
61. Cronin et al. 2015a, 2015b, OP CIT
62. Ibid
63. Hendricks, S.A., P.C. Charruau, J.P. Pollinger, R. Callas, P.J. Figura, and R.K. Wayne. 2015. “Polyphyletic ancestry of historic gray wolves inhabiting US Pacific states.” Conservation Genetics 16:759–764. And Hendricks, S.A., P.R. Sesink Clee, R.J. Harrigan, J.P. Pollinger, A.H. Freedman, R. Callas, P.J. Figura, and R.K. Wayne. 2016. “Re-defining historical geographic range in species with sparse records: Implications for the Mexican wolf reintroduction program.” Biological Conservation 194:48–57.
64. Mayr 1982, OP CIT
65. Patten, M.A. and K.F. Campbell. 2000. “Typological thinking and the conservation of subspecies: the case of the San Clemente Island loggerhead shrike.” Diversity and Distributions 6:177–188.
66. Hendricks et al. 2015, 2016. OP CIT
67. Cronin and Mech 2009. OP CIT
68. Wayne and Schaffer 2016 OP CIT
69. Fan et al. 2016. OP CIT and Fredrickson et al 2015. OP CIT,
70. Hendricks et al. 2016. OP CIT, (Wayne and Schaffer 2016), OP CIT
71. Hendricks et al. 2016. OP CIT
72. Leonard et al. 2005. OP CIT
73. Vilá, C, I.R. Amorim, J.A. Leonard, D. Posada, J. Castroviejo, F. Petrucci-Fonseca, K.A. Crandall, H. Ellegren, and R.K. Wayne. 1999. “Mitochondrial DNA phylogeography and population history of the gray wolf Canis lupus.” Molecular Ecology 8:2089–2103. And Fredrickson et al. 2015, Hendricks et al. 2016, Wayne and Schaffer 2016). OP CIT
74. Wilson and Brown 1953; Cronin 2006; Cronin and Mech 2009). OP CIT
75. Goldman 1944, and Hall 1981. OP CIT
76. Nowak 1995, 2002, and Chambers et al. 2012. OP CIT
77. Cronin et al 2015a OP CIT
78. Chambers et al. 2012. OP CIT
79. Weckworth, B, N. Dawson, S. Talbot, and J. Cook. 2015. “Genetic distinctiveness of Alexander Archipelago wolves (Canis lupus ligoni), reply to Cronin et al. (2015).” Journal of Heredity 106:412–414.
80. Cronin et. al. (2015b) OP CIT
81. FWS. 2016. Endangered and Threatened Wildlife and Plants; 12-Month Finding on a Petition to List the Alexander Archipelago Wolf as an Endangered or Threatened Species. Fish and Wildlife Service. Federal Register Vol. 81, No. 3 Wednesday, January 6, 2016 Proposed Rules pages 435–458.
82. Chambers et al. 2012
83. Wilson and Brown 1953.
84. Zink, R.M., J.G. Groth, H. Vazquez-Miranda, and G.F. Barrowclough, 2013. “Phylogeography of the California gnatcatcher (Polioptila californica) using multilocus DNA sequences and ecological niche modeling: Implications for conservation.” The Auk 130:449–458.
85. Zink, R.M. 2015. “Genetics, morphology, and ecological niche modeling do not support the subspecies status of the endangered the Southwestern Willow Flycatcher (Empidonax traillii extimus).” The Condor Ornithological Applications 117:76–86.
86. Malaney, J.L.and J.A. Cook. 2013. “Using biogeographical history to inform conservation: the case of the Preble’s meadow jumping mouse.” Molecular Ecology 22:6000–6017.
87. Cronin MA, MacNeil MD, Vu N, Leesburg V, Blackburn H, Derr J. 2013. “Genetic variation and differentiation of extant bison subspecies and comparison with cattle breeds and subspecies.” Journal of Heredity. 104:500–509.
88. Zachos et al. 2013.
89. Schweizer, R.M., B.M. vonHoldt, R. Harrigan, J.C. Knowles, M. Musiani, D. Coltman, J. Novembre, and R.K. Wayne. 2016a. “Genetic subdivision and candidate genes under selection in North American gray wolves.” Molecular Ecology 25:380–402. doi: 10.1111/ mec.13364. And Schweizer, R.M., J. Robinson, R. Harrigan, P. Silva, M. Galverni, Marco Musiani, R.E. Green, J. Novembre, and R. K. Wayne. 2016b. “Targeted capture and resequencing of 1040 genes reveal environmentally driven functional variation in grey wolves.” Molecular Ecology 25:357–379.
90. Schweitzer et al. (2016b) OP CIT.
91. (Cronin 2006), OP CIT.
92. Nowak 1995. OP CIT and Nowak, R.M. 2002. “The original status of wolves in eastern North America.” Southeastern Naturalist 1:95–130.
93. Chambers et al. 2012. OP CIT.
94. Cronin 1993, 2007, Cronin and Mech 2009, OP CIT; and Ramey, R.R. 2012. “On the origin of specious species.” In: Institutions and Incentives in Regulatory Science. J. Johnston (ed.), Lexington Books.
95. Cronin et al. 2015a.
{kind=link}