Chapter 13
The Morphic Fields of Animal Societies
From the point of view of the hypothesis of formative causation, social groups are ordered by morphic fields, fields which embrace and contain the individual organisms within the social unit or holon. Social fields, like morphic fields at all levels of complexity, are shaped and stabilized by morphic resonance.
In this chapter I consider the organization of animal societies and in the following two chapters the organization of human societies and cultures. This discussion explores the role of morphic fields in the coordination of social behaviour and of morphic resonance in social and cultural inheritance.
Animal societies as organisms
Societies of termites, ants, wasps, and bees may contain thousands or even millions of individual insects. Some species build large and elaborate nests (Fig. 13.1) and exhibit a complex division of labour. They have often been compared to organisms or described as superorganisms.1

Figure 13.1 Mounds made by compass termites in Australia. Their broad sides (top) face east and west; their narrow sides (bottom) face exactly north and south. They thus expose a minimum surface area to sunlight in the middle of the day, avoiding excessive heating. (After von Frisch, 1975)
Not surprisingly, there has been a long-standing debate as to whether such societies really are new kinds of organisms at a level above that of individual animals, or whether they are complex aggregates that are fully explicable in terms of the properties and behaviour of the individuals that comprise them. Should they be thought of holistically as systems or organisms at a new level of complexity with irreducible properties? Or can they be understood in a reductionist manner as nothing but the sum of their parts and the interactions between them? The reductionist approach dominated institutional biology throughout the latter part of the twentieth century, but the superorganism concept is now in the ascendant again.
According to the hypothesis of formative causation, the organization of social systems depends on nested hierarchies of morphic fields, with the overall field of the society organizing the individual animals within it through their morphic fields, which in turn organize their component organs, and so on down to the cellular and subcellular levels.
A wide variety of colonial invertebrates consist of individuals so harmoniously linked together that at first sight they are easily mistaken for a straightforward single organism. This type of organization reaches its extreme in colonies of the order Siphonophora, which vaguely resemble jellyfish and live in the open ocean, where they use their stinging tentacles to capture fish and other small prey. One well-known example is the Portuguese man-of-war; another is Nanomia which consists of many specialized individual organisms (Fig. 13.2). At the top is an individual modified into a gas-filled float. Below it are organisms that act like little bellows, squirting out jets of water which propel the colony; by altering the shape of their openings they are able to alter the direction of the jets. Through their co-ordinated action the Nanomia colony is able to dart about vigorously, moving at any angle and in any plane, even executing loop-the-loop curves. Lower on the stem other organisms are specialized for the ingestion and digestion of nutrients for the rest of the colony. Long, branched tentacles arise from them and are used to capture prey. There are also bracts, consisting of inert, scale like organisms that fit over the stem and help protect it from physical damage. Finally, there are sexual organisms, which produce gametes which through fertilization can give rise to new colonies.
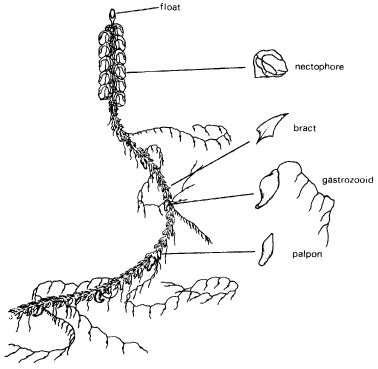
Figure 13.2 A colony of the siphonophoran Nanomia cara. The floats that buoy up the colony, the nectophores that propel it, the gastrozooids that capture and digest prey, and other member organisms such as the bracts are modified to such an extreme that they are comparable to organs of a single animal. (After G. O. Mackie, 1964, Royal Society Proceedings B159, 366–91)
These specialized individuals within the colony are like organs in an organism, and some are even connected together and co-ordinated through nerves. Such forms of life seem to be both colonies of individuals and composite organisms.2 Other colonial invertebrates, such as the corals, can likewise be regarded as both at the same time.
Societies of insects
The social insects, like colonial invertebrates, also exhibit a marked specialization of individuals. The queen is generally larger than the other members of the society and is specialized for egg laying, in some termite colonies producing up to 30,000 eggs a day.3 Among the sterile workers in many ant and termite societies there are distinct castes, including soldiers with formidable mouthparts. Even in bee societies, in which the workers look alike, there is a remarkable division of labour. In the hives of honeybees, some workers clean the cells and feed the queen and larvae, some construct and seal the cells of the honeycomb, some guard the hive, and others go out foraging. Any given worker can play all these roles, and typically does so sequentially, starting as a cleaner and ending as a forager.
The members of insect societies communicate with each other through the exchange of food, by means of various chemical substances, by touch, and in a variety of other ways, the most remarkable being the famous waggle dance of honeybees by means of which returned foragers indicate to others where food can be found.4
These societies have striking self-organizing properties. Honeybees, for example, maintain the temperature of their nest with remarkable constancy; from spring to autumn the interior temperature is almost always between 34.5 and 35.5 degrees centigrade. This degree of control is facilitated by the way they seal up all crevices and gaps, leaving only a single entrance hole. The output of heat from the insects themselves is the source of warmth, and in response to cold weather they huddle together in clusters. In warm weather the workers cool the hive by fanning with their wings; and if this does not suffice, some of them collect water, carry it into the nest, and distribute it on the brood cells; others spread the water into a film, from which evaporation is rapid, while a third group fans the moist air away from the brood cells and out of the nest.5
On the present hypothesis, over and above the behavioural fields of individual insects are the morphic fields of the society as a whole, which co-ordinate the activities of the individuals. These fields are spatially extended and embrace the entire colony; the individuals live within them. It is through these supra-individual fields that the colony comes into being and maintains its structure and organization, in spite of the continual turnover of individual workers, whose lifespan is generally much shorter than that of the colony as a whole. The self-organizing properties of these social fields are what enable the colonies to adjust to accidents, damage, and environmental fluctuations and to repair their nests.
The need for some such concept can be illustrated by considering the way in which termites construct their nests. The African fungus-growing termite Macrotermes natalensis forms vast colonies that last for years. Some colonies contain more than 2 million insects.
The nest develops from a small underground chamber made by the royal couple, and can grow to a height of ten feet above the ground. At the base of the mound is the nest proper, with the royal cell in its centre. In its many chambers, connected by numerous passages, are masses of finely chewed wood, on which the termites cultivate the fungus they eat. Above this there is a large air space enclosed by the outer casing of the mound, on the outside of which are ridges or buttresses. Channels as thick as a man’s arm radiate into many small ducts within the buttresses. The air in the fungus chambers is heated by the fermentation process and by the termites themselves; this hot air rises and is forced into the duct system of the ridges, the walls of which are so porous that they allow gas exchange: carbon dioxide escapes and oxygen penetrates from the outside. From these ‘lungs,’ the cooled and regenerated air now flows back down another system of wide ducts into the cellar, whence it returns to the nest.6
These structures are built by workers from pellets of soil glued together with excrement or saliva. But how do they know where to put these materials? In the words of Edward O. Wilson:
It is all but impossible to conceive how one colony member can oversee more than a minute fraction of the work or envision in its entirety the plan of such a finished product. Some of these nests require many worker lifetimes to complete, and each new addition must somehow be brought into a proper relationship with the previous parts. The existence of such nests leads inevitably to the conclusion that the workers interact in a very orderly and predictable manner. But how can the workers communicate so effectively over such long periods of time? Also, who has the blueprint of the nest?7
As the insects build the nest, they respond to the structure that has already been completed, as is they know what needs doing. For example, in the building of arches, workers first construct columns, and then if another column is being built sufficiently close by, they bend the column towards the other one (Fig. 13.3) until the tilted growing ends of the two columns meet.8 No one knows how they do this. They cannot see the other column: they are blind. There is no evidence that they run back and forth at the base of the columns measuring the distance. Moreover, ‘it is improbable that in the midst of all the confused scampering in the vicinity, they can recognize distinct sounds from the column by conduction through the substrate’.9 By a process of elimination, it is generally assumed that they must be able to locate the other column by smelling it.10
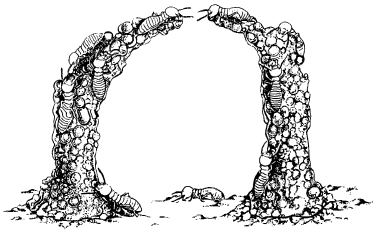
Figure 13.3 The construction of an arch by workers of the termite species Macrotermes natalensis. Each column is built up from pellets of soil and excrement, carried by the insects in their mandibles. When the column reaches a certain height, the termites, which are blind, begin to extend it at an angle towards a neighbouring column. (From Animal Architecture by Karl von Frisch, illustrated by Turid Holldobler. Copyright © 1974 by Turid Holldobler. Reprinted by permission of Harcourt Brace Jovanovich, Inc.)
In short, very little is actually understood about the way in which the termites construct these prodigious structures. Moreover, the conventional idea that instinctive abilities are somehow ‘programmed’ or ‘hard-wired’ in the nervous system might lead us to expect that termites that build such complex nests have larger and more complex nervous systems than species that build much simpler nests. In fact they do not.11
The hypothesis of formative causation provides an alternative approach, and suggests that the structures of the nests are organized by morphic fields embracing the nests as wholes, with a nested hierarchy of fields within them for the various elements of the overall structure. These fields are not inside the individual termites; rather the individual insects are inside the social fields.
If this is so, the organizing activity of the fields would extend beyond the range over which individual insects can communicate with each other by smell or by mechanical means. This idea is experimentally testable, and indeed there is already suggestive evidence that some such effects occur.
Over 80 years ago, the South African naturalist Eugene Marais made a series of observations on the way in which the workers of the species Eutermes repaired large breaches that he made in their mounds. The workers started rebuilding from all sides and the new parts all joined correctly, even though workers on different sides of the breach did not come into contact with each other and could not see each other, being blind.
Marais then carried out a simple but remarkable experiment. He took a large steel plate several feet wider and higher than the termitary and drove it right through the centre of the breach in such a way that it divided the mound and indeed the entire termitary into two separate parts.
The builders on one side of the breach knew nothing of those on the other side. In spite of this the termites built a similar arch or tower on each side of the plate. When eventually the plate was withdrawn, the two halves matched perfectly after the dividing cut had been repaired. We cannot escape the ultimate conclusion that somewhere there exists a preconceived plan which the termites merely execute.12
The same thing happened when the steel plate was driven in first and then a breach was made on either side of it.
Apparently this fascinating experiment has never been repeated; it would obviously be well worth doing so, preferably using material that would provide better acoustic insulation than a steel plate.
Marais thought that the queen was like the ‘brain’ of the colony and was somehow connected with the entire colony directly, over and above the chemical and other influences that were physically carried to other members of the colony by the workers who tended her. On the present hypothesis, such a linkage could be thought of in terms of the extended morphic field embracing both the queen and all the other members of the colony. Marais claimed to have demonstrated the existence of such non-material connections by means of simple experiments, such as the following:
While the termites are carrying out their work of restoration on either side of the steel plate, dig a furrow enabling you to reach the queen’s cell, disturbing the nest as little as possible. Expose the queen and destroy her. Immediately the whole community ceases work on either side of the plate.13
Again, Marias observations have not been followed up, but clearly it would be of great interest to know how immediate this effect is. An effect through the morphic field of the colony could be immediate, whereas an effect that depended only on normal sensory communication could not. In such experiments it would not be necessary to kill the queen; removing her from the colony would probably suffice.
Deborah Gordon and her colleagues from Stanford University made some intriguing observations on harvester ants in the Arizona desert that suggest another line of research. In order to study how ants switched tasks within the colony, Gordon’s team collected ants so they could mark them with paint, before releasing them and studying which tasks they performed on subsequent days. To collect them, they used an aspirator, a tiny vacuum device that quickly sucked the ants into a tube and deposited them in a vial. ‘Collecting foragers was easy; foragers didn’t seem to react at all when their fellow foragers suddenly disappeared from the trail. We were just another predator, like the horned lizard that stands beside the trail sucking up ants while the surviving foragers keep walking by, apparently heedless.’14 Collecting midden workers was easy too, and so was collecting nest maintenance workers, as long as the aspirator was operated properly. If any air flowed out of it, the gust of smells sometimes sent the maintenance workers running back to take cover in the entrance to the nest.
Collecting patrollers was completely different. Even the most careful aspirating of only a few patrollers, well apart from each other, could cause the whole colony to shut down for the day. At first, Gordon thought that this must be because of the change in pattern of interaction between the patrollers with each other and with other ants. But it happened too quickly:
When some patrollers outside the nest disappeared, the rest of the patrollers sometimes headed back into the nest immediately, within seconds – long before there was time for anyone to go back into the nest and assess the rate at which patrollers were returning. This happened when we put the aspirator right over the patrollers … so that there would be little opportunity for a cloud of alarm pheromone to escape. The patrollers still reacted, often at a distance that seemed too great for any pheromone to travel so quickly.15
Gordon then explored the possibility that this reaction depended on the rate at which patrollers met each other and touched antennae. But simulations showed that the variation in intervals between interactions was so large that ‘it would be hard for an ant to reliably detect a fall in the rate of interactions. So I don’t know how to account for the reaction of the patrollers.’16
These dramatic responses to the removal of patrollers occurred mainly in the morning, soon after the nests’ daily activity began and when the patrollers were circling around the nest. Later in the day, when the patrollers had moved further away, it was sometimes possible to remove patrollers without having much effect on the activity of the rest of the colony.
Probably the most decisive experiments to distinguish between the field approach to animal societies and the conventional approach could be done with ants and termites.17
Schools of fish
In vertebrates, the coordination of individuals within a group is sometimes so close that it is almost impossible not to think of them as a kind of composite organism. Many species of fish form schools or shoals, and at a distance a fish school resembles a large organism. Its members, numbering up to several million, swim in tight formations, wheeling and reversing in near unison. The fish do not follow particular leaders. When the school turns to the right or left, individuals formerly on the flank assume the lead.18
Schools exhibit characteristic patterns of behaviour, particularly in response to potential predators. When under attack, a school may respond by leaving a gaping hole around the predator (Fig. 13.4). More often the school splits in half and the two halves turn outwards, eventually swimming back around the predator and rejoining. This is known as the fountain effect, and leaves the predator ahead of the school. Each time the predator turns, the same thing happens.

Figure 13.4 The formation of an empty space within a school of fish around a predator. (From The Oxford Companion to Animal Behaviour, edited by D. McFarland, © Oxford University Press 1981. Reproduced by permission.)
The most spectacular of the schools’ defences is the ‘flash expansion,’ so called because on film it looks like a bomb bursting as each fish simultaneously darts away from the centre of the school as the group is attacked. The entire expansion may occur in as little as one-fiftieth of a second, and the fish may accelerate to a speed of ten to twenty body lengths per second within that time. Yet they do not collide. ‘Not only does each fish know in advance where it will swim if attacked, but it must also know where each of its neighbours will swim.’19
One of the most striking aspects of a school of fish is its polarization, whereby the members of the school swim in parallel. An analysis of the velocities of particular fish and the movement of the school as a whole has shown that ‘the school has no leader: that is, speed and heading are not closely related to those of any other single fish … Thus, in a sense, the entire school is the leader and an individual is a follower.’20
How schooling behaviour is co-ordinated remains a mystery. Vision no doubt plays a part; but some species continue to swim in schools at night. Moreover, in experiments in which fish were blinded by being fitted with opaque contact lenses, they were still capable of joining a school of normal fish and maintaining their position indefinitely within the school. Perhaps they judged the position of their neighbours by special pressure-sensitive organs, known as the lateral lines, which run along their length. This idea was tested by severing the lateral lines. Such fish still schooled normally.21
Even if the means by which they are aware of each other’s positions were understood, this would still not account for such rapid co-ordinated responses as the flash expansion: a fish could not sense in advance where its neighbours were going to move. However, if the school is organized by a morphic field that embraces all the fish within it, the properties of this field could underlie the behaviour of the school as a whole and help account for the co-ordinated behaviour of the individual fish within it.
Flocks of birds
Flocks of birds, like schools of fish, show such a remarkable coordination of their individual members that they too have often been compared to a single organism. The naturalist Edmund Selous, for example, wrote of the movement of a vast flock of starlings as follows:
Each mass of them turned, wheeled, reversed the order of their flight, changed in one shimmer from brown to grey, from dark to light, as though all the individuals composing them had been component parts of an individual organism.22
Selous also observed the way in which flocks of peewits, gulls, and other birds took off all at once, often for no apparent external reason.
A flight of ox-birds [dunlins] – some 150 to 200 I should say – went down on a mudflat here. After a little time, and when I had the glasses full on them, taking in the whole line, they rose all together instantaneously, without any visible extraneous cause. Soon they came down again, and now a swan flies across the line, not more than a foot or two above them. If all went up now, who could be surprised? But this was not the case. A certain number did, when the swan cut the line, but only a few inches from the ground, and settled again almost at once while the rest stood where they were. Some minutes later, under no discernible provocation, all rose in flight again on the instant.23
Selous studied the behaviour of such flocks over a period of thirty years, and became convinced that it would admit of no normal sensory explanation: ‘I ask how, without some process of thought transference so rapid as to amount practically to simultaneous collective thinking, are these things to be explained?’24
The banking movements of large flocks of dunlins have been studied by taking slow-motion films and examining the way in which the movement of the flock was initiated. The movement was not exactly simultaneous, but rather started either from a single individual or from two or three birds together. The initiation of a change of direction could occur anywhere within the flock, and manoeuvres always propagated through the flock as a wave radiating from the initiation site. These waves moved very rapidly, taking on average 15 milliseconds (15 thousandths of a second) to pass from neighbour to neighbour.
In the laboratory, captive dunlins were tested to find out how rapidly they could react to a sudden stimulus. The average startle reaction time to a sudden light flash was 38 milliseconds. This means that it is very unlikely that they can bank in response to what their neighbours do, since this banking response occurs quicker than the measured reaction time. When the films were examined very carefully, it turned out that at the very beginning of the flock’s movement the neighbours of the initiating bird reacted more slowly: on average the immediate neighbours took 67 milliseconds to react.
Wayne Potts, who carried out this study, proposed what he called the chorus-line hypothesis to explain it, based on experiments carried out in the 1950s with human chorus lines. The dancers rehearsed particular manoeuvres, which were then initiated by a particular person without warning. The rate at which the manoeuvre propagated along the line was estimated from films, and was on average 107 milliseconds from person to person, nearly twice as fast as an average human visual reaction time of 194 milliseconds. Potts suggested that the individuals saw the approaching manoeuvre wave and estimated its arrival time in advance. He thought his findings with dunlins supported this hypothesis: the slower reaction time of the immediate neighbours of the initiating birds occurred because they could not see an advancing wave, for this wave had not yet developed.25
At first sight, Potts’ chorus-line hypothesis appears to provide a straightforward mechanistic explanation for the banking phenomenon, removing the need for more mysterious factors such as thought transference or morphic fields. But does it? It is worth examining Potts’s hypothesis in detail because it is the most plausible conventional explanation for the way in which such flocks behave as wholes.
First of all, because these waves can propagate in any direction through the flock, Potts assumed that the birds sensed and noticed such waves almost immediately, even if they were coming from directly behind them. This required them to have practically continuous 360-degree visual attention, which is not plausible.
However, for the purpose of argument let us assume with Potts that this was the case. A second problem now appears. The birds did not respond to the approaching manoeuvre wave in a non-specific reflex manner like the startle reaction to a sudden flash of light. They changed their pattern of flight in a precise way; the angle, speed, and duration of their turning were precisely integrated with those of the rest of the flock; densely packed as they were, none of the birds collided with each other. Although there were a limited number of flock manoeuvres, such as banking, they were not stereotyped in their quantitative details. They were more flexible than a well-rehearsed chorus line going through standard routines. Not only would the birds have to sense an advancing wave, they would have to know exactly how they should turn.26 On the present hypothesis, the coordination of their movements depends on the morphic field of the flock.
There have been few studies as detailed as that of Potts. The most impressive concerned huge flocks of starlings, containing up to 2,600 birds, flying above Rome railway station. Starlings habitually gather in the evening and fly together over the roost for up to twenty minutes in spectacular aerial displays. They form sharp-bordered, strongly cohesive flocks. A team of scientists, including engineers and physicists, set out to measure the way in which the birds moved by setting up two cameras at right angles to each other, and analysed the digital films using a computer programme that could track individual birds. This was a complex technical challenge, and none of their films lasted more than eight seconds because of limited storage capacity of their recording devices. Also, because the cameras were fixed, they could only film starlings in a limited range of movement, when they were flying almost horizontally. They could not observe other aspects of flock behaviour, such as the response of the flock to an attack by predators.
A detailed mathematical analysis of the data showed that the movements of individual birds were coordinated with those of their neighbours up to seven birds away. What mattered was the number of neighbours, rather than their distance from any particular bird. This means that flocks of high and low density showed similar patterns of behaviour.27 Another important finding was that when the flock turned, the individual birds all changed direction within the flock, rather than wheeling in parallel paths. For example, in a 90-degree left turn, birds at the front of the flock ended up at the right of it, while those on the left ended up at the front.28
The more that flocks are studied, the more remarkable their behaviour seems. In a further series of observations on flocks of starlings near the centre of Rome published in the Proceedings of the National Academy of Sciences in 2010, it turned out that every bird in the flock was influenced by every other bird, however large the flock. In technical language, the birds were said to show ‘scale-free correlations’, meaning that ‘the behavioral change of one individual is influenced by all the other individuals in the group … The effective perception range of each individual is as large as the entire group … making the group respond as one.’ The authors of this study concluded, ‘How starlings achieve such a strong correlation remains a mystery to us.’ They suggested that their observations supported the metaphor of a collective mind.29
Computer models
There have been many attempts to simulate flock behaviour on computers. Craig Reynolds developed one of the best known, called ‘boids’, in the 1980s.30 This model is individual-based, that is to say it starts from individual boids. These boids are programmed to behave according to three simple rules:
- Steer to avoid being too close to neighbours.
- Steer towards the average direction that neighbours are heading in.
- Steer to move towards the average position of neighbours.
By following these rules, a collection of boids on the computer screen behaves rather like a flock. This simulated ‘emergent behaviour’ seems to show that the behaviour of the flock as a whole is a product of individuals interacting with their neighbours according to simple rules, with no need for any mysterious organizing principles. But while this may be true of the two-dimensional computer model, it bears little relation to the behaviour of real, three-dimensional flocks of birds. Reynolds developed the boids program starting not from data about real birds, but rather from a kind of computer programming concerned with ‘artificial life,’ typically involving two-dimensional models in which neighbouring units ‘interact’ according to simple rules. Special-effects wizards have used these kinds of programs to create animations of flocks or herds in films like The Lion King and Batman Returns.
Computer models of the boids type are useful for producing animations, but they are biologically naïve. As discussed above, flock behaviour does not depend simply on neighbour-to-neighbour interactions. Attempts to make mathematical models are now more sophisticated. One class of model combines principles of magnetism and hydrodynamics. In these models, each bird is like a magnetic domain within an iron magnet. Neighbours influence each other through their fields, and the flock as a whole has a field, like a magnet. But unlike a magnet, the flock is in motion, polarized in a particular direction, and by introducing the principles of fluid flow, it is possible to make models of some of the features of bird flocks, fish school and animal herds in motion.31
Other models consider what happens if a few members of the flock or school of fish have a preferred direction in which they want to go, for example based on knowledge of some feature in the environment not shared by other members of the flock. In one of these models, a minority of individuals were endowed with a strong preference, while most members of the flock were not. By moving in the same preferred direction, the motivated individuals caused the entire model flock to move with them. But if some of the motivated minority wanted to move in one direction, and others in another, the outcome depended on a kind of majority vote. If the numbers of birds motivated to go in different directions were approximately equal, the flock split in two. But if one direction had a majority in its favour, the flock moved in that direction, despite the minority that wanted to go another way.32
These models made it clear that groups are unlikely to remain cohesive and allow the spread of information if individuals respond only to others very close to themselves: ‘As sensory range is increased, a response to a greater number of neighbours increases cohesion and allows effective long-range transfer of directional information.’33 But although models based on some birds having strong preferences may explain some aspects of flock movement, they cannot explain them all, especially when the birds are not moving in search of food or any other directional goal, but simply flying together as starlings do before roosting.
Interesting as these models are, they are just a beginning of trying to understand behaviour of groups of animals that move in a coordinated way. Schools and flocks have distinct edges, and these models based on the behaviour of neighbours do not explain the edge effects. Moreover these models do not explain how flocks can be denser or rarefied, or what causes the movement of a flock as a whole when there seems no particular reason for it to move in one direction or another, or how a flock responds in complex and rapid way to attacks by predatory birds.
Morphic fields provide another way of thinking about the group as a whole and its relation to the fields of individual animals and their responses to their neighbours. The ‘bottom-up’ approach starting from individual animals and their neighbours needs to be complemented by a ‘top-down’ model of the field of the group as a whole, a field that has its own properties, inherited by morphic resonance from similar groups in the past, and from the past of the group itself.
Just as many species of fish form schools and birds flocks, so many mammalian species form herds or packs. These groups also move in a co-ordinated way, sometimes at great speed, and the animals do not collide. Here too the organization of the group can be thought of in terms of morphic fields.34 Again, this is not an alternative to communication between members of the herd through senses such as sight, sound, and smell. Morphic fields do not supplant the need for sensory communication; rather they provide the structured context within which the animals’ communications and responses occur.
The organization of animal societies
All animal societies by their very nature involve patterns of organization and coordination. Social animals respond and relate to each other within the context or framework of these structures.
In many species the social structure is relatively simple and may be only temporary – for instance, when males and females come together during the reproductive period and cooperate in mating and providing for their young, then go their separate ways again to lead a more or less solitary life. At the other end of the spectrum are a wide variety of complex and enduring social structures such as those of bees and chimpanzees. Ethologists have described many patterns of social organization, for example dominance hierarchies, as in the pecking order of chickens, and complex co-operative activities, such as the hunting behaviour of a pack of wolves.35
Everyone agrees that the patterns of organization of such societies are to a large degree inherited. The conventional assumption is that the social order somehow ‘emerges’ from the genetically-programmed interactions between the individual animals. But this merely restates the problem in different words. How do patterns of social organization emerge? On the present hypothesis, they depend on morphic fields, and these fields are stabilized by morphic resonance from similar past societies: the patterns of social organization are not inherited in the genes.
The concept of such social morphic fields is not an alternative to the many known forms of interaction and communication between animals within the group. A physical or chemical signal, gesture, or call from one animal to another has a meaning only within a context – indeed, anything that is meaningful or significant is only so by virtue of its relationship to other things. Morphic fields are what underlie such patterns of relationship and meaning.
Likewise, morphic fields are not alternatives to physiological influences on behaviour such as hormones. An increased amount of sex hormones in the blood-stream of birds, for example, can result in their entering a breeding phase. But the characteristic behaviour of birds in courtship, nest-building, incubating the eggs, and rearing their young is not explained by the chemistry of the sex hormones: different species have very different patterns of behaviour but the same hormones. Rather, such hormones bring about specific physiological and biochemical changes within the birds, which tune them in to the morphic fields of breeding behaviour, including the social fields that organize the complementary activities of the males and females.
Cultural inheritance
Just as an individual animal behaves in a way that is characteristic of its species and within this framework shows various habits and peculiarities of its own, so one animal society may resemble others in the same species, but at the same time has its own customs, habits, or traditions. Many of these behaviour patterns are related to the particular territory or environment that the group inhabits and are adopted by new members of the group, especially by young animals growing up within it. There is in effect a kind of social memory. Migratory animals provide spectacular examples of such traditional behaviour. Herds of reindeer, for instance, follow traditional migration pathways and return annually to the same calving grounds; various species of ducks, geese, and swans migrate in flocks of mixed ages along traditional routes year after year; and some of the breeding grounds of colonial birds have been used for centuries.36
Such patterns of group behaviour have an autonomy that depends on the group itself, they are not individually inherited. Young animals from one group raised within another generally adopt the patterns of the group within which they grow up. These habits or traditions develop through the participation of many individual members over many generations. They are forms of cultural inheritance.
There is general agreement among biologists that cultural inheritance cannot be explained genetically, but depends instead on another kind of transmission that takes place within the context of the group.
Perhaps the simplest forms of cultural inheritance can be explained in terms of individual imitation. For instance, in species of birds that learn their songs by listening to those of nearby adults, local dialects often develop. But in so far as the cultural traditions depend on the behaviour of the group as a whole, the tradition is transmitted by the group itself.
From the point of view of the hypothesis of formative causation, the forms of social behaviour are shaped by the morphic fields of the group. By morphic resonance these fields are influenced by the behaviour under similar circumstances of all similar groups in the past, right back through the history of the species. But because a given group in general resembles itself in the past more closely than it resembles other groups, it will be most strongly and specifically influenced by morphic resonance from its own past patterns of activity. This self-resonance is the means by which the traditions of the group are transmitted. The morphic fields of the group contain a kind of group memory.
In this chapter, we have seen how the idea of social morphic fields enables animal societies to be conceived of as social morphic units, or social holons, or superorganisms, and how this idea provides a way of understanding the coordination of the behaviour of individual organisms within the social unit: the colony, school, flock, herd, pack, group, or pair. The inheritance of these social fields takes place by morphic resonance from previous similar social units; and self-resonance from a group’s own past stabilizes its particular characteristics and traditions. I now consider how these principles apply to human societies and cultures.