
The future oceans—and what lives in them—are ours to choose.
Across the wide expanse of the oceans and their deep history, marine life is phenomenally tough. Marine species are the ultimate survivors: from microbes that conquered early Earth, to living fossils crawling from the Burgess Shale all the way to the Ocean City boardwalk. Some cling to boiling vents as the only food source in crushing depth and darkness. Others hew an existence from the bones of dead whales. Helpless anglerfish males comb the night for brides sustained only by faith and a gold nugget of yolk. Reef corals—just our current kind, never mind that there were others long ago— have survived five major extinctions and a quarter billion years of planetary perturbations.
Yet ocean life is also often fragile, living in highly specialized ways in highly specialized habitats. The secret of the extreme life of the sea is that it is extremely good at thriving in these difficult environments. And these successful species are often wonderfully adapted to a special niche—a particular way of life. On land such niches also occur, but they are often small—little fish living in skinny streams—so that they can only accommodate a few individuals of a specialized species. The oceans, however, are so big that even a relatively tiny niche can sprout whole ecosystems. Hydrothermal vents are a good example: they occupy a tiny, tiny part of the sea floor, and they each last only a few years. But the sea floor is so big that there are a lot of hydro-thermal vents, and marine life has adapted to skipping between them as the vents open and close.
Icefish survive temperatures that freeze blood. Swordfish employ eyeball heaters to accelerate their vision and catch prey. These intricate adaptations to ocean life rely on the ocean being so big that even a niche survival strategy can work. Once we see them this way, extreme marine species are like boutique businesses—extremely good at what they do and dependent on a thriving economy.
Unfortunately, these specialized successes are pressured from all sides by human activity. Every day we make life harder in the ocean. It doesn’t take much: a little more heat, a little more acidity, a hitch in the food supply, or an uptick in fishing. These changes have grown as the human economy has grown, and today the impact of more than 7 billion people presses down on the oceans. Humans affect ocean life all the way from the level of individual organisms to the functioning, and survival, of entire ecosystems.
If you had one foot in the coldest ocean habitat and one in the hottest, you’d straddle both the freezing polar sea and a seething hydrothermal stew. One foot would be at 30° F (–1° C) and the other at 250° F (121° C).1 Both places support a bright mélange of life, from icefish to Pompeii worms: life that thrives in its specialized niche.
In contrast to the wide temperature range of ocean life, global climate change projections suggest an increase in ocean temperature of 4–5.5° F 2–3° C).2 This may seem like a small number to us, because we are remarkably indifferent to climate. As big mammals living in air, with efficient heating and cooling mechanisms, we’ve got a warped perception of changes in heat. Our ancestors spread from tropical Africa to the edges of northern European glaciers in just thousands of years, warmed only by animal skins and open fires. Later they crossed the Bering Sea, spread to Texas, ambled across the Panama isthmus, took up residence in the Andes, and moved to the southern tip of South America.3 Along this path, we woke up to many different climates, but our big mammalian bodies helped us cope.
A human body uses about the same energy as a 100 watt light bulb to keep us warm or cool.4 When our body temperature drops, our metabolism speeds up a little to compensate.5 Calories burn faster and muscle activity generates more heat. And of course when outside temperatures rise, we sweat to dump the extra heat. Throughout these processes, the rest of our bodies operate at roughly normal temperature and speed. Our digestion, thinking, heart rate, and other metabolic functions are only slightly affected by exterior temperature.
But for cold-blooded creatures, environmental temperature is everything. An increase of about 22° F (10° C) in external temperature causes their metabolic rates to roughly double. The clock of life ticks twice as fast, more food is needed, and more waste products are produced. For perspective, a human heart doubles its rate after a single flight of stairs.6 Think of life at higher temperature as a perpetual workout.
This extra labor is difficult to sustain. San Francisco State University’s Jonathan Stillman tracked the heart rates of crabs in slowly heated water. As temperature increased from 61° to 93° F (16° to 34° C), heart rate climbed from 148 to 403 beats per minute—and finally crashed as the animals suffered fatal seizures.7 Stillman, no sadist, was investigating how close to this lethal temperature a typical crab lives. Surprisingly, he found that many crab species live pretty close to their temperature limits, inhabiting places where global warming’s projected 4–5.5° F (2–3° C) increase would prove fatal. For them, the projected temperature change over the next 100 years would not just bump up metabolism, it would push these crabs over a heart-rate tipping point on the hottest days of the year.
Climate change is poised to do just this.8 Already ocean species are on the move, shifting to higher latitudes, just as terrestrial species have.9 But they also face a second man-made threat in the form of a corrosive acid.
Human beings pump 9.9 billion tons of carbon as CO2 gas into the atmosphere every year.10 Nearly 25% of that enormous amount dissolves into the ocean, where simple chemistry takes over and the CO2 becomes carbonic acid—the same chemical that lets children dissolve iron nails in glasses of soda.11
Across the planet, carbonic acid and ocean acidity12 have steadily risen for decades. Acidity in the world’s oceans increased 22% since the beginning of the twentieth century and shows no sign of dropping.13 The risk isn’t that marine creatures will melt into goo like the Wicked Witch of the West—they will suffer in subtler, but equally dramatic, ways.
Marine animals make their shells out of seawater, conjuring hard skeletons from clear fluid in an almost magic fashion. Accumulating calcium carbonate in tiny pockets of seawater surrounded by tissue, shelled organisms force the substance into crystals by maintaining a precise chemical imbalance. In this way, infinitesimal layers of new shell form along the edge of an existing shell. A coral polyp sits on top of its own skeleton, built layer by layer over hundreds of years.
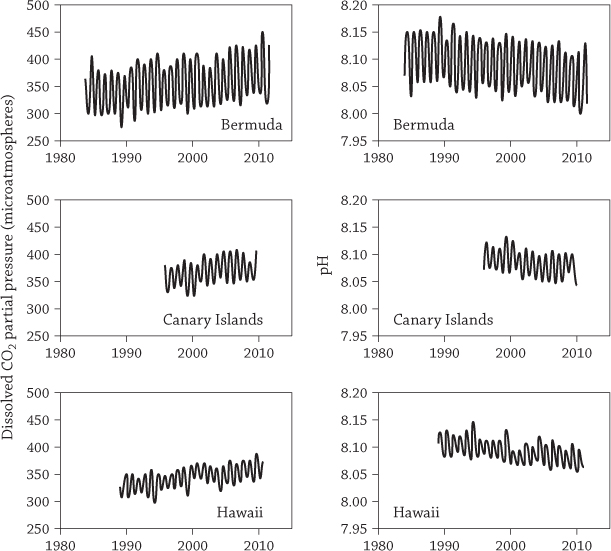
Increase in CO2 (left) and decrease in pH (right) at three places in the world’s oceans. Increased acidity results in decreased pH, so pH drops as CO2 increases.
But the chemical imbalance needed to make shell requires low acidity levels, much lower than normal seawater. It’s costly to reduce acidity for the purpose of shell-making, and the cost rises if the sea’s more acidic than normal. Think of boiling water on a stove: starting with cold water takes more energy and time to boil than does starting with warm water. And in the ocean, making shell starting with more acidic water takes more energy and time for the shell-making creature.
Rich Palmer, a marine biologist at the University of Alberta, estimated that a typical snail spends more food energy on its shell than on tissue growth or reproduction.14 Similarly, corals use 20–30% of their daily energy supply for skeletal construction.15 Increased acidity makes the process much harder, and over the next century the price of hard components (both shells and bones) may rise 30–50%.16 So, just as for high heat, acidity is a kind of metabolic tax.17 At a certain rate of taxation, the cost is too high, the metabolic bank is broken, and an organism cannot survive.
In a more acidic future, many shell-building species are likely to suffer. Hundreds of experiments have tested CO2 resistance in marine animals. Most show negative effects, especially for shell-builders, though the result isn’t universal.18 By the end of the current century, the predicted build up of heat and acidity will make it much more difficult for many species to thrive.19
Some of those species directly impact human welfare: oyster farms across the U.S. West Coast face reduced hauls stemming from ocean acidification. Like a young college graduate, the fate of an oyster depends sensitively on the climate in which it was spawned. For humans, entering the work force during a recession can cripple earnings for a whole career.20 Likewise, oysters spawned into acidic seas face clouded long-term prospects. Batches of eggs yield fewer viable larvae when released into water with a high CO2 and higher acidity. What’s more, those larvae don’t grow as quickly—their present weakness can persist, like human poverty, into the next generation.21
Ocean warming and acidification harm species in very different ways, but ultimately, their damage can be distilled to hard metabolic currency: calories flowing in from food, and calories burned by metabolism, growth, and reproduction. Acidification and overheating are, in their own ways, taxes on organisms’ metabolic income. Every calorie spent on higher metabolism to deal with heat or acidification is one not spent on growth or reproduction. The tax rate may be low today (though some species are already pinched), but it climbs with every year. By the end of this century, it will have become truly onerous for many species across much of the world.
Of course, the most successful species—the ones with the highest incomes—can most easily pay the tax. Some reef-building corals can ramp up their feeding and stave off acid’s burn.22 Having the right stress-resistant genes allows a coral to live in warmer water or a sea urchin to grow in low pH.23 Yet even these lucky species pay a price; every year of extra heat and acidity erodes their metabolic reserves. In the end, even the richest species will feel the metabolic tax of climate change.
Warm water expands a little bit, and when a lot of water expands a little bit, the ocean rises. This kind of thermal expansion is thought to be the major cause of the sea level rise, with important contributions from glacier melt and other human-made causes.24 As the ocean continues to warm, it will continue to expand, glaciers will continue to melt, and sea level will continue to rise. Even without the contribution from a catastrophic collapse of ice sheets into the sea, which could bump up sea level very quickly, the future of the ocean is that there will be more of it.
Average sea levels have risen 7 inches in the past 50 years, and the best science predicts a rise of 2.5–7.0 feet over the next 100 years.25 For low-lying coral atolls, such as Funafuti in the atoll nation of Tuvalu, in the central equatorial Pacific, such a rise will nearly inundate the country—the highest point of Funafuti is 9 feet above sea level. The rise in the western Pacific and Indian Ocean is likely to be even greater.26 This rising tide floods all shores, not just coral reefs. Salt marshes, like the ones that protect much of the U.S. east coast from wave damage, grow upward by only a quarter to a half inch a year and will be flooded if sea level rises as currently projected.27
Flooding is the most important consequence of sea level rise, not just because it damages habitats and alters the way many coastal species will live, but also because it promises huge damage to our human economy. Ten percent of all humans live in coastal zones, and that fraction is expected to rise.28 Increased tidal height and storms have already created urban devastation in New York City, costing $60 billion to repair. New Yorkers are proposing to spend $20 billion more to try to guard the city’s core against future catastrophes without even being certain that the proposed changes will work.
Around the world, the economics is similar—the cost of rising tides is the cost of coastal devastation by waves and storms, devastation that is often guarded against—now—by living marine life. Coral reefs catch the waves offshore and buffer erosion on the coast. Living corals absorb more wave energy than a dead, smooth pavement of former reef.29 But when the tide and waves are high, or the corals are dead, the usually placid shores of a tropical lagoon can become a cauldron of churned up land and sand. Man-groves protect other tropical shores. Places that had intact mangrove forests showed less damage from the 2004 tsunami in Southeast Asia than did areas where communities had cut down their mangroves.30 Coastal marshes, oyster reefs, and sea-grass beds play this kind of hidden buffering role on temperate shorelines.
Seawalls built to hold out the ocean might seem like a good solution to this problem, but they are expensive: $2 million dollars a mile in some cases.31 But suppose there was a seawall that would grow itself, and would also grow as sea level increased, so that it provided continuous protection for the coast? A healthy, fast-growing coral reef or marsh can do that at about 3–4 feet a century. And it will provide this service for free across hundreds or thousands of miles of coast. All that these free seawalls need is for their ecosystems to be kept healthy and for the coastal zones to be managed so that they can grow.
Maintaining those healthy ecosystems in a changing climate presents some major challenges. But adding to those challenges is a new layer of ocean problems—when the species in the sea, and the way they interact with one another, are fundamentally altered.
Once you understand a bit about the amazing life of the sea—from the smallest microbes to the largest whales—you can better see its incredibly interconnected nature. As in all closed systems, even gigantic ones like the world’s oceans, several effects can combine into a feedback loop and unleash even bigger consequences. Oceans will not only be warmer and more acidic; their ecosystems will also operate under fundamentally different rules.
Marine environments are powerful biological machines, capable of staggering levels of productivity. Human fishing fleets remove about 88 million tons of fish and shellfish annually, which sounds like a big number until you realize the ocean’s population of marine microbes produce this amount of biomass in about an hour (see Chapter 3). Left to pile up, this bounty would swiftly bury us all in sludge. It doesn’t—because equally massive systems exist to consume that biomass. The burgeoning biomass is held in check by the normal ecology of the sea.
Microbes and single-celled algae fuel a worldwide food web that eventually produces sardines, whales, turtles, tunas, sharks, and every other extant marine creature. Human beings won’t destroy that productivity—microbial life is so old and powerful, we simply can’t—but for the first time, we can direct it. Humans are so numerous, so technologically advanced, and so widely polluting that our species can finally alter the planet’s biggest habitat and its most abundant citizens, the microbes.
The effects pile up, quickly and literally: an unebbing tide of bacteria, algae, and other weeds all freed from the ocean’s normal checks and balances. That unleashed and uncontrolled productivity is an ironic bloom of living things, but it wrecks the way the ocean naturally channels energy from species to species in the great chain of life. The combination is a fearsome feedback loop we call a Productivity Bomb. It has two primary fuses: one involving the most advanced marine life, the other the least advanced.
Decades of relative peace and prosperity following World War II led to an odd and undeclared global conflict: the War on Fish, where the expanding global population devoured everything the oceans could provide.32 Humans won the war, naturally, but our victory was somewhat Pyrrhic. Rampant overfishing has reduced populations of fish across the world’s oceans, from the tropics to the polar seas. We are simply taking more than the ocean can replenish— something we never realized was possible. This shift is unique in the planet’s natural history. Never before could a single predator have such a massive global impact.
In the Philippines, off an island called Bohol known for its rolling “Chocolate Hills,” sits an unusual coral formation: double barrier reefs. On a calm evening near the edge of the inner reef, Steve Palumbi gets ready for a night dive. The dive is led by Amanda Vincent of Project Seahorse, and seahorses are precisely what they’re hunting. Both are dressed in wetsuits with tanks and regulators. Masks, flashlights, steel knives, watches, and depth meters round out their expensive equipment. The Filipino boatman, piloting his tiny banca, wears a fraying pair of swimming trunks along with goggles carved from a coconut husk and two disks of Coke-bottle glass. A kerosene lantern casts wan light from the bow. With soft splashes, each of the three divers goes over the side. Amanda leads Steve off to seek her favorite seahorses. The boatman takes off alone, frog-kicking down toward the reef, his bare feet a stark contrast to Steve and Amanda’s flippers. He’s got a family to feed and his own seahorses to catch—for sale rather than for science.
It takes Amanda and Steve 30 minutes to find just one seahorse, 45 feet below the boat. Elegant and enigmatic, it grips a coral branch with a delicate prehensile tail. It eyes the divers warily. A larger figure looms out of the dark water, and Steve swings his flashlight around to reflect off coke-bottle goggles. The boatman has doubled back in his hunt. With a wave he turns away, leaving them and the rare little animal in peace.
The scientists surface after an hour, cold and tired, unable to find a second specimen. The pilot ferries them back to shore, but once their gear is unloaded, he heads back out to sea. For the next 8 hours, he’ll swim shivering through the dark water, diving again and again, finally returning near dawn with his own seahorse. It’s only one animal, the length of your long finger, but he’s able to sell it for 25 cents. It’s enough to buy a few cups of rice. His wife and children will eat for another day. When the Sun goes down again, he’ll return to hunt on the reef.
There were once plenty of seahorses in these waters; the men who fished them led easier lives. Only a few hours of work would yield a bagful of wriggling little flute-nosed fish. But as generations of fishermen took freely from the reef and the Philippine population exploded, seahorses became truly rare on the reefs.
Tales of local overfishing tug at the heartstrings because of the immediate impact on fishermen and their families. In the past, the fishing industry would respond with technology. For thousands of years, people fished from wooden craft propelled by wind or their own oars. They cast hemp lines and hurled harpoons. Modern fishing boats are battle tanks compared to these chariots. Satellite navigation, high-precision sonar, diesel winches, and deep freezers make these vessels technological marvels. If there are fish to be had anywhere on the globe, industrial fishing fleets will find and take them. But for all their sophistication, these high-tech armadas often don’t take more fish than the simpler fleets of the past. How could this be?
Ruth Thurstan and Callum Roberts chronicled the history of one fishery in Scotland: the Firth of Clyde, a region of deep submerged canyons in the Irish Sea. Fed by the North Atlantic’s nutrient-rich waters, it consistently yielded big hauls of herring, cod, and haddock through the early nineteenth century. As time wore on, bottom-trawling ships with steam engines replaced the old wooden craft. Bit by bit the fleet was modernized. Scouring the Firth’s deep chasms improved fishing yields, but only temporarily. Thurstan and Roberts found that before long, despite these better ships, the catches dipped far below their previous levels.33
Callum Roberts cast a wider net with his research and found the same patterns repeated throughout Great Britain. Each time the trawling fleets took a technological leap, the catch would rise but then rapidly decline. Today in British waters, for all our technological achievements, it’s twenty times harder to catch a fish than it was 200 years ago.34 The fish are no smarter or more evasive—they’re just less common.
And when a previously common fish is stripped from the sea, there are consequences for other species. For their predators, because food will now be scarce. And for their prey, because there are now fewer checks on the growth of the prey population. The result is an ocean ecosystem out of balance.
The fish-eating public prefers to eat oceanic predators. We have decided they taste better: tuna, salmon, marlin, grouper, rockfish, orange roughy, halibut, and flounder all are fearsome predators in their natural environments. And when we eat these fish, we change the ecology of the oceans, because we are lopping off the top of the food chain. Daniel Pauly and his colleagues called this fishing down the food web.35 Like blowing the tops off mountains in search of coal, this approach does massive collateral damage.
No one appreciates the unintended consequences of overfishing like scallop fishers from the Carolina coast. For generations they harvested bay scallops (Argopecten irradians), scooping millions of pounds from muddy sea-grass beds.36 As the twentieth century wore on, the scallop numbers started to fall. Down and down they slid, finally hitting rock bottom in 1994, when the entire season’s harvest netted just 150 pounds.37 The fishery collapsed, but not because of overharvesting the scallops themselves. In this case, the culprit was a salty soup brewed halfway around the world.
Shark-fin soup is a traditional Chinese delicacy that signals status and prestige. As China’s economy grew over the past few decades, many millions of people suddenly could afford to serve shark-fin soup at their children’s weddings and other important events. To feed this fad, the world-wide hunt for shark fins exploded over the past decades, and eventually came to the Carolina coast.
Most sharks are not really very extreme—their most extreme features are their teeth and evolutionary past (see Chapter 2)—but they are prodigious eaters. By the time they’re full grown, they’ve consumed countless times their weight in lesser seafood. Off the placid Carolina coast, sharks make regular meals of cow-nosed rays (Rhinoptera bonasus), chomping them like leathery pancakes—that is, they used to. With the shark population depleted by international fishing fleets, the gentle cow-nosed rays were freed from most predation. The rays frolicked through this new undersea utopia, reproducing in vast numbers and merrily devouring their own favorite prey: bay scallops. Thick shells were ground to broken china in the rays’ flat powerful jaws, and one scallop bed after another was depleted. Fishermen found themselves desperately competing against the rays.
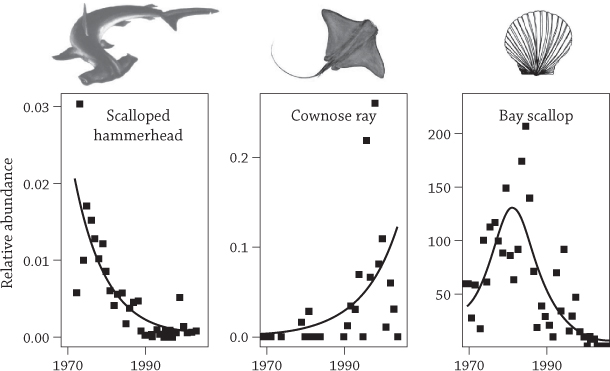
The decline in coastal sharks (left) was followed by an increase in their prey, cow-nosed rays (middle), and a steep decline in bay scallops (right). Redrawn from Myers, R. A., J. Baum, T. Shepherd, S. Powers, and C. Peterson. 2007. “Cascading effects of the loss of apex predatory sharks from a coastal ocean.” Science 315:1846–1850.
Here is the great chain of life at work: sharks eat rays, rays eat scallops, scallops feed us. Ecologists call this a trophic cascade: a natural chain reaction of collapsing and exploding populations. But when the cascade is altered, it can leave the sea fundamentally unbalanced. When the food chain is frayed or fractured, life accumulates just below the fissure. Kill the sharks, and their prey run amok. The backup of life at the break in a food chain is like cars backed up at an accident: the flow of traffic comes to a halt. But the delivery of cars along the highway doesn’t stop, and so a traffic jam ensues.
Imagine sitting in that traffic jam, because of an accident up ahead. As 5 o’clock comes around, thousands of people get off work and hit the roads. The extra cars make the extant clog much worse. The same thing can happen in a food chain if discarded fertilizer kicks its smallest species into overdrive.
The Mississippi River drains 41% of the central United States, including the country’s agricultural breadbasket.38 Mountains of nitrogen, phosphorous, and potassium are slathered on American fields every year.39 At least 3 billion pounds of discarded fertilizer wash down the Mississippi each season, most of the product flowing into the Gulf of Mexico.40 That fertilizer is great for plants when it is on the farm fields, but it’s also a microbial bonanza. The annual runoff creates ideal conditions for the single-celled algae that live in the warm, calm waters near the Mississippi’s mouth. In days they can reproduce to make enormous blooms, forming dense surface slicks that color the water for miles.
The pattern is global. In 2010, runoff from French pig farms and other agriculture produced a bolus of green algae off the Brittany coast. Countless tons of thick green filaments washed onto the beaches and rotted in the Sun. The decaying piles were so foul that they incapacitated a horse and rider on the beach, killing the horse and necessitating rescue for the rider.41 For the past decade these algae have regularly plagued the shore—the pig farms have never ceased operation—and more than a few animals have succumbed to the fumes.
Red tides, brown tides, and yellow tides have grown alarmingly common across the globe—all named for the colored algae that produce them and churn out noxious toxins. Shellfish and other grazers eat the algae, collecting the poison in their own bodies and potentially passing it to human beings.42 In 2008 alone, more than 400 harmful algal blooms were recorded worldwide.43
A heavy tide of nutrients doesn’t have to be a bad thing. In Antarctica, nutrient-rich upwellings from the deep sea feed seasonal megablooms of algae. Krill and copepods teem where the algae bloom, reproducing at planktonic rates and powering an ecosystem that’s come to rely on such bounty. In March 2012, researchers from the Australian government’s Antarctic Division noticed a floating bloom of algae 120 miles long.44 Likely made from the single-celled species Phaeocystis, the bloom bound up billions of cells in a gooey mucus matrix.45 Algae are a natural part of the ecosystem, and the polar food web consumed the bloom in mere weeks. In this case, the productivity explosion was blunted by transferring it from the bottom of the food chain to every other level, dispersing the excess energy and keeping the system balanced. In the absence of human intervention to sever the food chain or fuel further algal growth, there was no traffic jam.
The Gulf of Mexico has no comparable system left intact. Algae grazers like shrimp and oysters are depleted by overfishing, so the Gulf struggles with fertilizer runoff. The extra productivity takes too long to filter up the food chain. Algae die faster than they’re eaten, and then voracious bacteria move in to consume them—a second bloom following the first, rapidly consuming oxygen. When the oxygen is gone, they shift their metabolic engines into an oxygen-free mode and keep on eating. The bacteria live without oxygen quite well, but they acidify and suffocate the sea for thousands of square miles around.
Overfishing and nutrient pollution react with each other, creating much worse problems than either might alone. So far, we have examined examples that carefully document each separately, but we have also seen them act together. We have seen the Productivity Bomb’s results firsthand.
In the Soviet Union’s dying days, the Black Sea faced the Bomb’s full fury. In this small inland sea, porpoises had been hunted heavily for decades, and were only at about 10% of their historical numbers, so the small fish that were their normal prey flourished.46 The abundant fish ate the zooplankton, small crustaceans that in turn normally graze on single-celled algae (phytoplankton) floating in the water. Too few porpoises led to many small fish, and too few zooplankton, and too many phytoplankton: ecologists consider this a classic tropic cascade. The first fuse of the Productivity Bomb—disrupted food chains—was lit.
Then, industrial farming along the rivers feeding into the Black Sea dumped a huge load of fertilizer into it, triggering massive blooms of floating algae. Massive reds tides plagued the Black Sea, creating dead zones that killed bottom-dwelling invertebrates and many fish.47 The whole environment skewed toward algae, copepods, and anchovies until the little silver fish found themselves targeted by overactive fishing fleets. With anchovies gone, there was nothing left but algae and copepods.48
The copepod bonanza might have helped decrease the algal blooms. Enter a new species from elsewhere in the oceans—a comb jellyfish introduced into the Black Sea from the vast ballast tanks of intercontinental cargo ships.49 The jellyfish ate copepods aplenty, and in the depleted ecological setting of the Black Sea, there was nothing to hold back a jellyfish explosion. So they formed vast drifts in the cold water. It turns out the jellyfish swallowed juvenile anchovies too, preventing any fisheries recovery. This entire system, massively perturbed, had no way to correct itself. The result was leagues of barren water where once many fisheries had thrived. Barren, but for 3 tons of jellyfish in every square mile of ocean surface.50
The collapse of the Soviet Union cut off the flow of discarded fertilizer down the rivers leading to the Black Sea, and overfishing of anchovies ceased. The invasive jellies began to be devoured by a second invasive jellyfish that ate the first jellyfish. The Black Sea ecosystem gradually began to regain its balance, the grinding chain of destruction painfully rewinding. These changes were not simple: the repair bill included major shifts in agriculture, fishing, and porpoise protection. The first successes of all these efforts have begun to restore a functioning ecosystem in the Black Sea and show that the Productivity Bomb can be reversed.
Tropical corals have long been half animal and half plant. This internal diversity lets them thrive when many creatures would starve. The coral animal eats as its ancestors did: sweeping its tentacles through the water to capture tiny creatures. Its protein diet is full of the phosphorous and nitrogen that plants need to grow, and so the animal passes these nutrients to algae that live placidly inside its own cells. The algae take the phosphorous and nitrogen and combine them with abundant sunlight to manufacture carbohydrate food. The interior algae transport these sugars to the coral host as rent payment.
It is a clever arrangement; the algae couldn’t thrive in most tropical waters, because these oceans tend to be nutrient poor. So, the corals capture prey and use them for algae fertilizer. The corals cannot catch enough food to grow sturdy skeletons by themselves, and in turn they rely on the algae to drive their growth. Together they form a partnership that for hundreds of millions of years has survived under conditions that would starve each separately. What’s more, the reefs they built shelter thousands of other species.
But this arrangement is deceptively fragile. Dump sewage and farm runoff into the water, consume most of the plant-eating fish, and watch a Productivity Bomb detonate. Fleshy green seaweeds—abundantly available once there is enough discarded fertilizer in the water—begin to grow quickly where they couldn’t before. And where the herbivores have been removed by fishing, the algae can grow unchecked. Terry Hughes first saw this in Jamaica in the 1980s: millennia-old staghorn reefs and cities of mounding corals were carpeted and killed by weedy algae. Their survival hung by a thread.
That thread was a sea urchin named Diadema, a baseball-sized animal with venomous black spines the length of chopsticks. Its needle-sharp spines made it a bad food for fishermen, and they left it alone. But those fishermen did not leave the fish alone, and for decades the reefs of Jamaica had been stripped of algae-eating fish. Yet the coral reefs remained healthy and growing. The reason was that Diadema came out of its cracks and crevices at night and scoured the reef, eating all the algae they could find.
But then the thread broke. The Diadema were caught up in a massive disease outbreak that swept the Caribbean in a single year, killing millions or even billions of sea urchins. The last herbivores were gone, and no safeguards remained to defuse the Productivity Bomb. Algae swept over the reef, smothering corals until only a fraction of the original reef remained. Today, over-fishing continues. The urchins have not fully recovered. And neither have the corals, relegated to a few thriving reefs, the rest scattered across their former domain in broken bone yards.
Jeremy Jackson, a saber-rattling marine ecologist (yes, such people exist) with long kinky orange hair pulled back in a ponytail, has long examined ocean problems through an unconventional lens. He has traveled the world, pointing out the oceans’ fundamental collapse—the steady march through ever more devastating Productivity Bombs. Trained as a paleontologist, Jeremy helped create the field of historical ecology: comparing modern ecosystems to those of the past.51 His “Reefs since Columbus” research was unveiled in 1996 and ever since, he’s pointed to the demise of coral reefs as a genuine historical shift.52 He and his colleagues call our present course “The slippery slope to slime,” a wonderfully alliterative phrase describing the ocean’s epochal shift from large fish and coastal gardens to a stew of bacteria, jellyfish, and tar-like algae.53 The oceans of the future will be extremely productive—maybe even more so than today, in terms of raw biomass—but they’ll leave much of the human race without the food we are used to, and perhaps with oceans devoted to microbes. The Productivity Bomb’s most common physical product is slime.
Imagine an ocean seething with microbes, its red-brown surface weedy with algae. Vast dead zones and tides of neurotoxic water hamstring high-order productivity, wiping out the top half of the food chain while its base fills the world with toxic sludge. Waves crash on the beach, kicking up drifts of sticky green foam that cling to the sand. The clean salt air turns sickly toxic with the stench of decay. The ocean’s delicate species have perished during the extremely hot days of summer or have expired in a bankruptcy of expensive skeletons.
Even the highly successful species in the ocean will find their comfortable niches altered by climate change, and like a pensioner on a fixed income, will not forever be able to absorb the metabolic tax of a warmer, more acidic ocean. The massive bolus of discarded fertilizer we put into the ocean, and the surgical precision with which we can cut out the very species that can quell the resulting ecological revolution, selects for a certain set of organisms that has always been in the oceans but hasn’t been our favorite part.
We leave an ocean legacy of small, simple, soft, deep, blind, and weedy species. Curtains of jellyfish, lawns of microbes, a mud pile of worms, and perhaps a few lonely anglerfish in the deep sea. The oceans of the future will be alive—nothing will ever truly kill them—but what remains won’t be the life we currently know.
The oceans are in a state of emergency, suffering from more ailments than science has remedies to offer.54 If the policy “medicine” of reduced CO2, better land management, fishing restraint, and protected areas isn’t swiftly administered, the oceans will experience truly dire crises at ever-increasing rates.
All species will feel the pinch of changing temperature and acidity, from the depths of the sea where atmospheric CO2 will eventually percolate, to the melting polar ice caps. Maybe hydrothermal vent shrimp won’t notice much, bathed as they are in boiling poison. Maybe the broad abyssal plains will still flicker with bioluminescence. But great Antarctic whales will suffer when krill have to live without sea ice. The coldest species will suffer when the temperature rises a few degrees. The hottest corals, already living near their heat ceilings, will break through these ceilings. The shallowest intertidal species will bake at low tide and drown under storm surges. The oldest fish in the ocean may watch their descendants live progressively shorter, weaker lives. Even the deepest-ocean organisms—probably the least environmentally flexible species in the ocean—may find their homes awash in high acidity or altered currents.
Of course, in a few million years, conditions will improve. Nature is good at balancing itself out, after all. Previous massive alternations in the oceans have smoothed out, over geologic ages. And in that amount of time, the same time it has taken the human species to diverge from our ancestors, invent tools, and lift our eyes to wonder at the world around us, the planet and its diversity might recover.
Which is to say, over the long term the oceans don’t need saving. People need saving. People will need to live through the next hundreds or thousands of years when the oceans are no longer the pantry of the world, no longer safe to swim in or sail across, toxic and wracked by ever-stronger storms. Hundreds of millions of people now live directly off the sea, and billions more benefit indirectly from marine life. Human society cannot simply wait out a centuries-long global Productivity Bomb or wait millennia for a return to “normal” ocean productivity. The fate of the oceans has become our fate too, and we out of easy ways to ensure that the future of the oceans is secure.