one
The Natural History of Pine Trees
Pines, genus Pinus, are native to the northern hemisphere – North America, Eurasia and down through China into Southeast Asia. The British have a limited view of them, unless they have had the cause or curiosity to observe many species. Scots pine is most familiar to them: a handsome tree with short, dark foliage, small cones and orange bark on its upper branches. They might gaze, perhaps disparagingly, across blocks of plantation conifers, unaware that they are looking at lodgepole pine (P. contorta), native to North America, pass shelter belts of P. nigra, commonly known as either Austrian or Corsican pine, or admire ornamental specimens of Japanese black pine (P. thunbergii) in gardens. None of these gives the faintest hint of the variability of which the genus is capable, or of its range or habitats. In addition there are many subspecies, variants, cultivars (forms deliberately selected by gardeners) and hybrids, naturally or artificially produced.
The clearest point that distinguishes pines from their close relatives such as spruce and fir in the family Pinaceae is the needles, or leaves, ‘long and small like Threds; hard, durable, and constantly green; pointed or prickly [sharp] at their Tops; surrounded at the Bottom with a membraneous Sheath’, as the French apothecary Pierre Pomet reported in the late seventeenth century.1 These emerge from dwarf shoots, which in turn grow from the larger, easily visible, long, woody branches of the tree. Firs (genus Abies) and spruces (genus Picea) are most commonly confused with pines; these two genera both have single needles all over the growing branches. Those of firs leave circular scars when pulled off, differentiating them from spruces, which leave a small, curved peg at the base, making the twigs rough.

The papery sheath (actually another form of leaf ) binds the leaves together in a fascicle; inside this the leaves fit tightly together at the base, where a cross-section shows them like wedges dividing a pie. The number of needles per bundle is important for initial identification: depending on the species they grow in twos, threes or fives, or up to eights in some Mexican pines – although there are atypical species: parry pinyon (P. quadrifolia) with four leaves in most bundles, and single-leaf pinyon (P. monophylla) with only one leaf per bundle but still with a fascicle sheath, and therefore a true pine. The fascicle sheath, too, is an important identifier; in some species it is shed and in others it remains on the tree.

The drooping needles of the Mexican weeping pine (P. patula).
All details are important to the botanist, but this is not a book about botany. One thing remarkable to the observer of a group of pine species – for instance, in a pinetum – is the variety in colour, texture and especially length that the needles display. Some, especially in species from cold or arid environments, such as Rocky Mountain bristlecone pine (P. aristata), or jack pine (P. banksiana of Canadian boreal forests), are only 2–3 cm long. Needles may leave the branch in short, dense, stubby masses (the aptly named foxtail pine, P. balfouriana), or grow in tufts at the ends of branches (Korean pine, P. koraiensis). Individual needles might twist (Bishop pine, P. muricata, from southwestern North America; Chinese red pine, P. tabuliformis) or grow long and filamentlike, up to 25 cm in Mexican egg-cone pine (P. oocarpa, in Spanish piño de colorado), droop gracefully in bunches over the cones (chir pine, P. roxburghii of the Himalayas) or weep (Mexican weeping pine, piño triste, P. patula). Some bear a Christmas-card frosting of white resin flecks (the bristlecone pines), curve in a graceful arc (Japanese white pine, P. parviflora), or show other distinctive features such as a layer of white wax on the inner surface (Martinez pinyon, P. maximartinezii, in Spanish maxipiñon). Colour, in many pines, is deep forest green, but the range includes a handsome blue-green (sugar pine, P. lambertiana), dull grey-green (gray pine, P. sabiniana) and yellow-green (sand pine, P. clausa).
One species, Krempf’s pine (P. krempfii) of the Vietnamese forests, has scimitar-like needles 3–7 cm long and about ½ cm wide. These puzzled botanists so much on its discovery that they were uncertain how to classify it and suggested it be given its own genus, or at least subgenus – Ducampopinus. It is now considered likely to be a true pine.2
Although pines are colloquially called evergreens, the needles are actually shed. This happens every two to three years in most species, but foxtail and bristlecone pines retain their needles for up to seventeen years. John Muir (1836–1914), writer and champion of the natural environment of the Yosemite Valley, said that the foxtail pine was given its name by ‘the miners on account of its long bushy tassels. It is by far the most picturesque of all pines.’3 The habit of retaining needles gives foxtail pine branches their characteristic tufted appearance.
The cone, the fruiting body, consists of scales arranged in a spiral around an axis. It was also described by Pomet, who said that it begins
by a Button, which arrives to be a large scaly Apple, almost round, or pyramidal, of a reddish Colour: These Scales which form it are hard, woody, thicker commonly at the Point, or Top, than at the Bottom [that is, where they join the central stem]; hollow lengthwise with two Cavities, each of which contains a hard Shell, or oblong Nut, cover’d, or edg’d with a thin, light, reddish Rind.4
The term ‘strobilus’, plural ‘strobili’, derived from a word that means ‘turning’, was adopted as the overall botanical term for the immature cones containing the reproductive structures of pine trees (and other conifers). Each tree bears both male and female strobili in spring. The male strobili, also known as pollen cones, are sometimes called ‘catkins’ or ‘flowers’, although neither male nor female strobili are technically flowers.

Male pollen cones on an umbrella pine (P. pinea), releasing a cloud of pollen to the wind.
Pollen cones cluster around the bases of growing shoots and are relatively small, up to about 15 mm long. They are usually creamy-yellow or tan in colour, but some species bear red, purple or pink pollen cones. Pomet described these cones as being ‘of several membraneous Foldings . . . fill’d with nothing but a light Dust’.5 This is pollen, produced in enormous quantities. Two minute air-filled bladders support each individual grain. Pines rely on the wind to distribute pollen and it can be carried vast distances. Near forests, it can be very noticeable: the early nineteenth-century North American botanist Stephen Elliott (1771–1830) wrote of loblolly pine (P. taeda) that:
This species, (as I believe of all the real pines) . . . bears aments of sterile flowers [that is, pollen cones] in clusters at the summit of the branches . . . the flowers, when mature, discharge so much pollen, that [the] surface of stagnant pools appears to be almost covered with this ‘yellow dust’ . . . In the streets of Charleston, after heavy storms, I have seen small ponds margined with the pollen which had been born by the winds across adjacent rivers.6
The female cones, Pomet’s ‘buttons’, are technically known as mega sporangiate strobili. Initially small, they grow singly on the branch tips or around the bases of terminal buds (the buds at the ends of branches that progress the growth of the tree in the subsequent growing season). Oval or egg-shaped and a couple of centimetres long, they stand upright into the wind. Their spiral structure is clearly visible, the scales recurving slightly, fresh and pliable, thin and delicately beaded with resin drops along their curved edges. The colours vary according to species and are often vivid – pink, deep red, magenta or purple.
Female cones contain two types of scale – ovuliferous ones, which bear the seeds, and bracts, modified leaves, supporting them. Cut a young cone in half lengthways and the ovules appear as pearl-like, rounded shapes clustered along the vertical axis. Wind-blown pollen drifts between the scales, where it is trapped by resin. A grain develops a pollen tube penetrating the ovule wall at a point where it faces the axis of the cone, and cells inside both the pollen grain and the ovules divide once. The scales close up, their edges sealed by resin, and the cells become dormant until the growing season of the following year, when the ovule develops two egg cells.7 Likewise, two sperm cells develop in the pollen tube, and fertilization occurs. The seeds develop, two per scale, cupped in dark hollows, their embryonic root systems facing the axis.
|
Branch of Scots pine tree (P. sylvestris) showing new season’s growth and immature female cone. |

Cones take two to three years to mature, drooping from the branches as they grow. Immature cones may be a brilliant magentapurple, bright viridian or pale whitish-green depending on the species; their surfaces are shiny and textured with a complex origami of oval or diamond-shaped scale edges. As they reach maturity, they become the more familiar tan or chestnut-brown colour. Their common name, ‘cone’, derives from Latin and describes their overall shape, pointed at the top, curved below, the angle of a quadrant. On the exterior overlapping scales spiral from base to apex. Pick off the woody scales one by one and the arrangement around the central axis becomes apparent.
All pine cones grow to the same general plan, but they show remarkable variation in size, form and texture. Two overall pine-cone forms catch the attention. One is elongated and narrow, with slightly flexible, leathery scales and an elegant, slim curve springing from the peduncle that attaches the whole to the branch (associated with a major subgenus known as Strobus). The other (more typical of subgenus Pinus) is more compact, egg- or oval-shaped, with hard and woody scales on a shorter, thicker stem. Differences are apparent in details of the scales. In the slender, flexible cones, the exposed part of each scale is long from top to bottom, a deep scallop terminating in a small bump. In the more compact, woody form, the long axis of the scales runs around the cone, the exposed edges forming a network of close-packed diamonds, each with a ridge running like a backbone from side to side. In the centre of each is a more or less pronounced bump, often bearing a spike or prickle. The name for this bump – on the terminal edge of the scale in one type of cone, dorsal in the other – is an ‘umbo’ (Latin for the central boss of a shield). In some species it forms a woody pyramid. The shape and size of a cone, the appearance of the umbo and the presence or absence of a prickle are all important for identifying different species.
The huge, spiked cone of the gray pine (P. sabiniana), native to California. |
|

On some cones the prickly umbo is so obvious that it is incorporated into the name. Bristlecone pine cones, both immature and mature, literally bristle. The Table Mountain pine of the eastern USA gained its Latin epithet, pungens, meaning ‘piercing’, partly because of its prickly leaves, but also because the cone bears sharp, upcurved spikes. Shortleaf pine (P. echinata) has immature female cones that bristle like hedgehogs or sea urchins, and P. muricata, the Latin name of bishop pine, recalls the spiky murex snail.
Pine cones range widely in size, from jack pine, whose tiny cones are only 2–3 cm long, to sugar pine of Oregon and the Californian Sierra Nevada, with cones up to 50 cm long. Weight, too, ranges from a few grams for the smallest cones up to that of the ‘big cone’ pine, Coulter pine (P. coulteri), whose dense, rounded, oval, massive woody cones weigh up to 2 kg; Torrey pines (P. torreyana) also have massive cones, and in both species the scales carry prominent sharp hooks.
Several pine species, such as knobcone pine (P. attenuata), have cones that are noticeably lopsided. Jack pine cones are ‘remarkable for curving to one side, which gives them the appearance of small horns’, said John Claudius Loudon in the 1840s.8 The asymmetrical cones of Monterey pine (P. radiata) have scales markedly larger on one side than on the other, and their off-set stems make the cones lean at an oblique angle when fallen. Species vary, too, in terms of how the cones open. In some this is immediately on ripening, when their seeds shed to the wind, the cones falling to the ground afterwards; but others have cones that remain attached to the tree, disintegrating scale by scale. ‘Serotinous’ cones remain closed on their parent trees, their scales sealed by resin, until the heat of a forest fire melts this and they burst, shedding seed over the fire-cleared ground. The most extraordinary example of this occurs in knobcone pine, whose closed cones hang in clusters on the tree for years, while the wood of the trunk and branches slowly expands beneath and around them, gradually engulfing their forms. The woody, pointed and slightly curved cones resemble in shape nothing so much as the abdomen of a wasp, giving the trees a surreal appearance, as if groups of giant insects were clinging at intervals to their limbs. Other species rely on external agents, especially birds, to distribute their seeds; these include whitebark pine (P. albicaulis), whose cones do not open even in fires.

Anatomy of a pine cone showing the scales with the hollows which hold the seeds in an illustration to Joseph Pitton de Tournefort’s Corollarium Institutionum rei herbariæ . . . (1703).
These differences lead to variability in seeds. Inside a ripe, unopened cone the seeds lie in pairs, cupped in the hollows of the scales. Each is enclosed in a thin, hard dark brown shell. Trees that rely on wind dispersion have small seeds, each with a single, parchment-like wing. Relative area of seed to wing varies according to the species; one of the smallest, that of Japanese red pine (P. densiflora), is a tiny, thickened point on a calligraphic sweep, a graceful exclamation mark of a seed. The general shape and plan are repeated in varying size and detail across species, but the seeds are never longer than about 3 cm in total. Some species have a detachable wing that grips the seed coat from either side between pincer-like extensions. The wing catches the wind as the seeds fall out of the cone and the seeds ‘“helicopter” to the ground, languidly rotating in still air, beating against rocks and tree trunks in a stiff breeze, and possibly being dispersed over miles’.9
|
Clark’s nutcracker in an illustration by J. J. Audubon for his Birds of America (1840–44). |

Other pine seeds are large and appear wingless; the wing is actually present but reduced to the pincer-like arms. The trees they come from are often called stone pines, a name that referred originally to Italian stone pine (the reasons for the name are unknown, but may relate to the hardness of the seeds). They include Siberian pine (P. sibirica), dwarf Siberian pine (P. pumila), Korean pine (P. koraiensis), Swiss stone pine and various pinyon pines of the American southwest. Mexican pinyon (P. cembroides) was named after the arolla or Swiss stone pine (P. cembra), partly because of resemblances between the seeds. These pines have adopted a seed-distribution strategy that requires other organisms. Birds are the most important vectors, although other animals help – bears in North America and Siberia, squirrels and small rodents more generally.
The relationship between birds and pines has been most carefully studied in the mountains of Nevada, where whitebark pine has coevolved with Clark’s nutcracker (Nucifraga columbiana) in a relationship that benefits both tree and bird.10 These large grey birds remove ripe cones from the trees, clasp them between their feet and extract the seeds, storing up to about 90 in a special pouch under their tongues. Then they fly away and hide the seeds elsewhere, often several kilometres away. One study found that a flock of nutcrackers stored about a ton of seeds in one autumn. Inevitably, some are never retrieved and grow into trees.
Other bird species distribute pine seeds, notably pinyon jays (Gymnorhinus cyanocephalus), raucous birds that inhabit the pine woodlands of the American southwest. In late summer they prise green but ripe cones off the branches. They pick them apart, store the seeds in their throats and hide them for food in winter and the spring breeding season. The birds seem to discriminate between dark brown seed coats (likely to contain a good nut) and ones that are probably empty, which tend to have tan-coloured coats. Pinyon jays rely on pine nuts to feed nesting females and young, harvesting until all the seeds have either been collected or have fallen. Poor autumn crops seem to lead to a delay in breeding until new green cones appear on the trees the following summer. Researchers suggest that the visual trigger of the cones stimulates testis development in male pinyon jays.
Apparently, those brilliant emerald seed cones with their sparkling beads of resin are an aphrodisiac for the garrulous blue crow of the woodlands, persuading him that while late breeding may be poor breeding, it is better than no breeding at all.11
Inside their shells, pine nuts are small (even the largest, those of single-leaf pinyon, are only a little over a centimetre long), with an apparently insignificant aperture at one end – so small that one might assume that it had been caused by abrasion in shelled nuts. When the nut is divided carefully in half it can be seen that this is not so: it contains a minute, fragile, rod-like structure running from the hole through the length. At the rounded end, the rod has a tiny whorl of palest, delicate green, the whole being a pine tree in waiting, with a slender embryonic root and closely furled cotyledons.
Given the right circumstances, a pine seed on the ground surface finds sufficient warmth and moisture to enable the outer shell to split. The narrow end quickly develops a root, penetrating the soil. This helps to haul the other end upright, delivering water and nutrients to the growing plant and the cotyledons, which deepen in colour, expand and unfurl above ground. There are between five and 24 cotyledons, depending on the species. They are soon replaced by primary, then secondary leaves. This stage in the existence of some pines, especially longleaf pine (P. palustris), is known to botanists and foresters as the ‘grass stage’. It lasts several years and is followed by a sudden growth spurt. Other species simply grow steadily. Young pines need light to grow well, and are crowded out by more shade-tolerant species.
Pine-root systems often develop in conjunction with mycorrhizal structures, fungi that colonize their outer surfaces with a pale coat. This, despite a sinister appearance, is a symbiotic relationship from which the fungi take nutrients, and which benefits the tree by facilitating the uptake of minerals, helping pines colonize poor soils.12
All sapling pines follow the same general growth pattern. A pseudo whorl (actually a very tight spiral) of secondary branches develops around the base of the leader. The tree expands rapidly over the space of a few years, the leader reaching ever upwards and secondary branches expanding sideways, giving the stereotypical pointed conifer shape. It produces two different types of shoot. One type comprises the minute dwarf shoots that support leaf bundles; the other, much more obvious, the large, branching shoots that progress the growth of the tree. Pines grow in increments. Every year the growing tip divides to produce a leading shoot surrounded by a further spiral of branches. These new, initially upright stems – needles closely packed parallel to the direction of growth, with a pale, slightly waxy appearance – are known colloquially as candles. Growth is seasonal. In northern climates it takes place in spring, while in more tropical conditions it is dictated by wet and dry seasons. If planted in uniformly warm and humid tropical conditions, pines sometimes display a condition known as ‘foxtailing’, in which a tremendously long, ever-growing leader develops.13
Scots pine seedling (P. sylvestris) emerging from the seed coat. |
|

The leader grows rapidly upwards for a few centimetres, then stops and forms a terminal winter bud. This contains a series of other structures – buds for pollen cones, and for dwarf shoots. Lateral buds sometimes contain shoots or seed cones, in which case they will not grow further. Once the shoots have expanded to their full length within a given season, the tree will not grow any more until the following year.
From the start, the seedling pine lays down harder tissues that strengthen its core. Initially these are composed of the starch polymer cellulose, but after about fourteen days more complex woody tissue, lignin, starts to form. It is produced in the spring growing period. Cells divide from the cambium, the layer of tissue one cell thick that covers the entire plant from the smallest root tip to the end of the newest growing shoot. On the inside they produce xylem, which carries water and nutrients from the root. Through the length of this run tracheids, elongated canals of former xylem cells, which conduct water upwards through the tree; these form about 90 per cent of the wood.14 Their walls contain pits that communicate horizontally, allowing water movement across as well as vertically. Collectively they make up the sapwood, which is usually a pale creamy colour. As the tree grows, those in the centre gradually clog with minerals and acquire a deeper, often reddish colour, becoming heartwood. Physiologically inactive, this forms the strong core of the tree and is more resistant to decay than sapwood. New wood is produced early in the growing season, when hormones from bud growth stimulate the cambium to divide, producing new cells for both the inner bark and the xylem; early-season new-wood cells are wider and paler in colour than later, narrower, more lignified ones. Together the two form an annual ring.
In addition, pines have resin canals. Resin is not sap (which is a solution of nutrients in water). Resin is secreted by special cells, has a protective function and permeates all parts of the tree: needles, cones, branches, heartwood and the stumps of cut or dead trees. Resin is also visible on growing needles and immature female cones. The trunk is especially rich in resin, producing cells that surround vertical and horizontal canals running through it.15 When a tree is damaged in some way – if a branch is broken, a hole is made by a wood-boring insect or a cut is made in the bark – the cells that line the resin canals become active. They may be remote from the site of the damage, but produce a flow of resin that ejects foreign matter and seals and protects the wound.
Resin is a factor in one of the most notable things about a pine forest: the smell. Although fragrant and unforgettable, this is difficult to describe, perhaps because it is more one-dimensional than perfumes associated with flowers. It is also more ephemeral. Nicholas Mirov, a botanist and chemist who spent his life studying pines, called it ‘primeval’ and explained that the essential oils of pine that emanate from the needles lack fixatives, the substances which give many perfumes stability. Pine resin ‘consists of three kinds of substances: volatile oil, commonly called turpentine; the non-volatile part called rosin; and the high-boiling ingredients’.16 The latter hold back the volatile oils, making them longer lasting. The volatile oil composition differs between species, giving each one a unique perfume. Compounds collectively known as terpenes are responsible for the smells. Some trees have simple mixtures of alpha and beta pinenes, which give the characteristic smell of turpentine, but many have unusual and distinctive chemistry. Lodgepole pines from the Rocky Mountains contain phellandrene, which smells like grass, and Italian stone pines contain limonene, also a component of citrus smells. Jeffrey pine (P. jeffreyi) resin contains no terpenes at all, but is composed of various aldehydes dissolved in heptane; it has a distinctive sweet smell described variously as resembling that of violets, vanilla or pineapple. Other species have their own distinctive but indescribable fragrances. This is one of the more inscrutable aspects of pines, and the reason for the existence of perfume in pine oil and resin remains unknown.
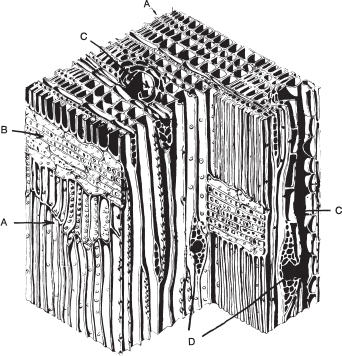
Diagram of the structure of pinewood. Vertical resin canals are shown at C, and horizontal ones at D.

George Inness, Georgia Pines, 1890, oil on wood. Pine trees in the warm climate of Georgia, U.S.: an image which conveys the claustrophobic quietness of some pine forests.
Outside the cambium the outer cells die and become compacted as bark, a protective layer against wounds and fires. As the tree increases in girth, the outer layers break up in rifts and polygons. In older trees the layers at lower levels can become several centimetres thick. Bark patterns vary from species to species. In younger trees the difference is not especially noticeable, although whether the bark is smooth or rough is an aid to identifying whether trees are placed by botanists in subgenus Strobus or Pinus. In older trees, patterns become quite distinct. The most striking and ornamental are seen in the Chinese species lacebark pine (P. bungeana), whose Chinese name means ‘white bark pine’, much esteemed in northern and central China. Lacebark pines shed bark in plates, giving irregular, grey-green and chalky-white patches that are very attractive. Cinnamon-coloured bark is a distinguishing feature of slash pine (P. elliottii); mature Scots pines have a distinctive glowing orange bark on their upper branches and Austrian pine (P. nigra) derives its Latin name, ‘black’, from the dark-coloured bark that contrasts with that of Scots pine. Whitebark pine has, as its name suggests, grey-white bark when young. Many species develop typical patterns: irregular, chequerboard like blocks (western white pine, P. monticola); plates (longleaf pine, whose orangebrown bark breaks up into small plates divided by deep, dark grey furrows); roughly rectangular, vertically oriented plates divided by deep dark furrows (ponderosa pine, P. ponderosa); or irregular, dark greyish-brown ridges separated by reddish-brown furrows (Monterey pine).
Eventually a tree achieves full height, as allowed by an interplay of genetics and environment. This is a subtle and highly variable element of the natural history of pines, which is partially responsible for the confusion that surrounded – and still to some extent surrounds – the classification of different species. Lodgepole pine is a notoriously variable species, partly because of the wide range of habitats it occupies. As a ‘shore pine’, it occurs as shrubby, twisted trees (as suggested by the Latin name) of relatively small stature, rarely reaching 30 m in height down the coast of the Pacific northwest of America. Inland, from the Yukon down through the Cascades and into the northwestern part of the USA, the trees grow tall and straight in stereotypical conifer spires of up to 50 m in height. Continuing inland, further south in the Cascade Mountains of Oregon and the Sierra Nevada, lodgepole pines (often known as tamarack or tamrack in this area) are generally trees of straight growth and slender trunk growing up to 40 m tall. The species was described by Mary Curry Tressider as
often a most untidy tree; the dead branches clasping the lower portion of the trunk, the bark stripped and hanging, and the lower limbs drooping mournfully toward the ground combine to give it a most desolate appearance.17
The tallest pine species is sugar pine. ‘No-one forgets their first meeting with a sugar pine,’ said John Muir. On discovering it in 1826, botanist David Douglas wrote:
and lest I should never see my friends to tell them verbally of this most beautiful and immensely large tree, I now state the dimensions of the largest one I could find that was blown down by the wind: Three feet from the ground, 57 feet 9 inches in circumference; 134 feet from the ground, 17 feet 5 inches; extreme length, 215 feet.18
Sugar pines have been heavily logged, and none of such broad dimensions survives, but botanists reckon the height to range from 40 to 60 m (130–97 ft) exceptionally up to 85 m (280 ft).
On reaching their full height, pines lose their conical shape and spread or broaden, their crowns becoming less dense, oval or rounded in shape, and their lower branches shedding to give less regular silhouettes. Pliny the Younger described the ash cloud from the eruption of Vesuvius in AD 79 as looking like a pine tree, ‘for it rose high up into the sky on what one can describe as a very long trunk, and it then spread onto what looked like branches’.19 He had in mind a multi-branched structure with a spreading crown, typical of mature pines, especially the Italian stone pine with its umbrella-like crown. In outline, the gray pine of the southwestern USA looks a little like an oak, but the foliage and cones distinguish it clearly from these. Sugar pines shed their lower limbs to give a tall, bare trunk, and have always captured the imagination of naturalists for grandeur and form:

Mottled colours on the bark of lacebark pine.
Completely disregarding the conventional spire-like forms of their associates, Sugar Pines toss out horizontally from the massive trunks a few, huge, irregularly-shaped, and lengthened branches.20
Chinese red pine takes its Latin epithet, tabuliformis, from the flattopped shape the crown develops on maturity, and Japanese black pine develops a picturesque, irregular, windswept form. The twisted and irregular forms of pines and the notion of endurance and longevity have an ancient connection in China, where the trees are a feature of many landscapes, notably precipitous mountain scenery. Much more recently, in Western culture, the age-contorted, pale, weathered spires with a few dark living branches of the aged bristlecone pines of the southwestern USA have became icons of endurance.
Few pines form shrubs, although the European mountain pine (P. mugo) of the Alps and Carpathians has a low, open, multi-branched form. Other pines, frequently stunted by the environment as well as genetics, are the partially dwarfed scrub Jersey, or Virginia pine (P. virginiana), which grows on poor soils on the eastern seaboard of the USA, and the dwarf Siberian pine, which grows in mat-like bushes and withstands the freezing winters of the Siberian tundra. Whitebark pines grow up to the timber-line in the Sierra Nevada of the southwestern USA, where ‘they are more like carpets than shrubs or trees . . . beaten down by the heavy blankets of snow and clipped smooth by the furious gales of winter’.21 Sometimes the German word krummholz (krumm, bent, twisted, and holz, wood) is used to describe particularly bent mountain pines that have become stunted and deformed by exposure to tough environments.
Fire is a natural phenomenon in conifer forests, an integral part of the life cycle of pines. It is the death of many individual trees, but one that allows natural regeneration to flourish. The trees have adapted to it, achieving a dynamic equilibrium that allows them to dominate forests in which non-Pinus tree species would otherwise succeed them, crowding out the light from newly germinated pine seedlings. Lowintensity grass fires hug the ground, burning grasses and small shrubs, and doing relatively little damage to trees, but high-intensity fires, driven by wind, are hot and sweep through the crowns of mature trees, burning everything in their path and usually killing them. With their wood full of resin, pines are naturally flammable. They are drought tolerant and often grow in climates that have hot, dry periods of weather during which thunderstorms occur. Before humans learned how to make fire, the conflagrations in pine woods must have been due to natural phenomena such as lightning strikes. Man, accidentally or otherwise, must have vastly increased the occurrence of fire.
Evolution has given pines strategies against fire. The ‘grass stage’ in young longleaf pine is a defence against frequent low-intensity fires shortly after germination. The dense tuft of leaves protects the seedling for several years, while below ground a strong root system develops rapidly, acting as a food store if the topknot of leaves is burned off. Once the root is sufficiently developed, the tree grows quickly through the stage when it is vulnerable to a size large enough to resist low-intensity fires. Pitch pine (P. rigida) and Virginia pine exhibit a rarer strategy among pines: resprouting branches from burned trees. Red pine (P. resinosa) has a curious characteristic, probably related to fire resistance, in that roots of these trees in dense stands connect to those around them. If individual trees are cut, their stumps can remain alive, their root systems fed by those of the surviving trees.
A different strategy, used by trees with serotinous cones that are sealed until the heat of an intense crown fire warms them, is one in which the adult trees perish but young ones replace them. Although this is catastrophic for the individual trees, seeds are scattered onto ground newly cleared and fertilized by burning, ready to start the cycle again. Knobcone pine provides the most extreme example of this phenomenon. Bearing cones while still a sapling of a few years old, these and all subsequent cones remain on the tree until the heat of a fire opens them. Bishop pine, Monterey pine and lodgepole pine are also examples of these ‘fire-dependent’ species. Some pines – lodgepole pine, Monterey pine, jack pine – retain dead branches on their lower trunks that provide fuel for potential fires, making them especially vulnerable to high-intensity crown fires. Typically, the trees occur in single-aged stands, cohorts generated by fire.22

The twisted, wind-polished wood of a dead bristlecone pine (Balfourianae spp.).

previous: The resins contained in living pine trees explode into fire, creating flames hundreds of metres in height.
For animals and humans caught in fires, the story is one of fear and uncertainty. John McPhee, writing about the pine barrens (vast areas of poor soil that support little but pine and an understorey of grass and a few shrubs) of New Jersey, notes the ‘V’-shape a typical forest fire describes on the ground as it progresses through a forest. The point of the ‘V’ represents the head fire, which leads lateral fires that spread out from each side. The line occupied by a fire may be 3 m or hundreds of metres wide, and the lateral fires may suddenly gain the fuel and energy to become head fires in their own right. Unpredictable in direction, intensity and width, forest fires are terrifying – the only safe place is on ground already burned.23 Fire moves swiftly and threatens life and property; not surprisingly, the thought of a natural but unpredictable fire regime is unpopular with both foresters, for whom trees are a crop, and those who have chosen to make their homes in forests, for work or because they love the environment. The Canadian author and naturalist Chris Czajkowski gave a dramatic description of a forest fire that threatened her home on the remote Chilcotin Plateau of British Columbia:
Behind us a huge black wall of smoke had erupted. It had towered above us and the leading edge was beginning to curl over the cabin . . . most alarming was the vivid orange colour at the bottom of the smoke wall . . . the flames must either have been huge or a good deal closer than we had been told. The whole of the north ridge was haloed in orange. The water on the lake . . . was a dull orange brown, the trees a sickly dull green. White ash started to fall.24

A cluster of pointed cones on a knobcone pine (P. attenuata).
Sometimes human communities learned to use fire to create a particular environment in pine woods. In eastern North America some fires were ‘managed’ by indigenous American tribes who periodic ally burned the undergrowth in the forests. They may have done this as often as twice a year, burning grass and forest litter, creating a habitat in which grasses for game animals such as deer and elk flourished, and making an environment across which they could hunt more easily. Regular, frequent fires reduced debris and burned at low intensities. Manipulation of fire over a long time created open stands of mature trees in some areas, such as the longleaf pine forests of North Carolina. Early European settlers in North America followed native patterns of fire management, but later arrivals had a view that did not accommodate such a dynamic natural environment, and fire became something to be suppressed.25
In the absence of fire, pines tend to occupy sites on which other tree species are unable to grow – poor, rocky soils in the Great Basin of Colorado, serpentine soils on Cuba and in the Balkans, or sand dunes in Les Landes in southwest France. Opportunists and survivors, they flourish in a wide range of often unpromising circumstances: ‘They . . . affect the cold, high and rocky grounds’, wrote John Evelyn (1620–1706) in his book Sylva, or a Discourse of Forest Trees (first published in 1664).26 ‘The debris of granitic rock may be considered as the universal soil of the pine and fir tribe, and a dry subsoil an essential condition for their prosperity’, was the opinion of John Claudius Loudon (1783–1843), although he admitted that ‘they will grow on all soils whatever, that are not surcharged with water.’27 Although in popular literature pine trees are associated with northern, cold, gale-infested, rock-bound mountainsides, they grow happily in more fertile soils and gentle climates. Their problem is that faster-growing plants crowd them out.
The only naturally occurring southern-hemisphere pine trees are in a stand of Sumatran pine (P. merkusii) found in the Barisian Mountains of Sumatra, at two degrees south of the Equator, but a surprising number of species grow in warm northern-hemisphere climates. Sumatran pine and Caribbean pine (P. caribaea) – the latter grows on low-lying flats on islands such as the Bahamas and the Turks and Caicos – are both essentially tropical species, but several others rank as subtropical and grow in Southeast Asia, Central America and the Caribbean. In Cuba, Cuban red pine (P. tropicalis) grows in grasslands, or in stands in the Pinar del Rio province, the wet and dry seasons showing in the wood as paired light and dark rings. Hispaniolan pine (P. occidentalis) forms a forest in Haiti. Southeast Asian examples of subtropical pines include Vietnamese white pine (P. dalatensis) and Taiwan red pine (P. taiwanensis) from Taiwan and southern China. However, pines disdain humid tropical environments, in which they will grow, but not reproduce. Apart from such environments, and the most frozen of arctic wastes and arid of deserts, they have a wide distribution in the northern hemisphere.
The opportunist nature of pines is visible in open ground, whether it has been cleared naturally by forest fires or gales, or by man and then abandoned (‘old field pine’ is a common name for loblolly pine because of its invasive tendencies). Once seeded in such places they grow quickly – quicker than competing trees – and establish dominance. This can also be a weakness, for mature pines shade out their own light-loving seedlings. Eastern white pine (P. strobus) seeds on newly cleared ground, then outlives hardwood species that grow up with it; shade-tolerant species succeed them, and the white pines continue growing, towering over a forest in which their own seedlings can no longer germinate.

Pine trunks, stripped by fire of needles and branches, look like stitches holding the fabric of a snowy Yellowstone landscape together.

The precipitous granite peaks of Huangshan, hung with pines, scenery admired and painted for centuries by the Chinese.
The pine tree as a survivor can be seen in high altitudes and high latitudes. Pines can tolerate cold and dryness on mountain ranges up to the treeline, or close to the Arctic Circle, where they grow as singlespecies forests, or in company with other conifers such as spruce or larch, or deciduous trees such as birch. The most extreme examples of survival are the Great Basin bristlecone pine (P. longaeva) and the closely related Rocky Mountain bristlecone pines and foxtail pines which live for thousands of years in the thin atmosphere and arid, rocky soils of high uplands in the southwestern USA. The oldest living single plant on earth is a pine tree, a Great Basin bristlecone pine known as Methuselah, whose precise location is known only to a few botanists and foresters. About 4,500 years old,28 this tree was already venerable by the start of the Christian Era. In contrast, Monterey pines generally live for about a century.
The survival of a species is sometimes a matter of geography and luck. Monterey pine has an extremely limited natural range on a small area of the coast of California and three islands just offshore. This zone is bounded on the north and east by a climate that is too cold for the tree to survive in winter, and on the south by one that is too dry. It is an area prone to sea fog, which provides moisture, and ‘It seems . . . reasonable to suggest that it is only a fortuitous accident of oceanography and coastal geomorphology giving rise to local sea fogs that have saved the species’ that allowed Monterey pine to survive in this dry climate zone.29
Those who study pine trees recognize natural ‘pine regions’ in which pine flora is distinctive and specific in combination with particular trees and shrubs. Their boundaries depend on geology, topography and climate. North American pine regions run down either side of the continent. In eastern Canada and the USA they form a broad mass, spreading inland from the Atlantic coast across lowlands to the Great Lakes and the Mississippi plains, forests of great economic significance. The species gradually change depending on climate and soil, the most northerly being jack pine, graduating southwards through eastern white pine, red pine, pitch pine, Virginia pine, longleaf pine, loblolly pine, shortleaf pine and slash pine. Pine barrens are a feature of the poor soils of New Jersey, Virginia and North Carolina.
On the west coast, mountain ranges run north to south parallel to the ocean, providing habitats from sea level to about 3,500 m. Here a diversity of pine species grow in intricate mosaics, oriented roughly in north–south bands, the cold-tolerant species occurring at increasingly high altitudes as one proceeds further south, until they eventually meet conditions too warm or too arid. In the boreal forests of Canada, lodgepole pine holds sway. Further south the range of species increases to give some of the richest conifer flora in the world, including ponderosa pine and sugar pine; inland, the dry semi-deserts of Nevada and adjoining states are home to bristlecone and pinyon pines. These pine regions do not overlap, except in northern Canada, where lodgepole pine and jack pine occur together.
The east–west divide in regions relating to pine flora continues into Central America. The western one extends down the mountains of the Sierra Madre Occidental. The eastern one fringes the Gulf coast of the southern U.S. and Mexico, encompassing both the mountains of Central America and some Caribbean islands. These two southern pine regions overlap in a species-rich confusion in southern Mexico and Guatemala, where the Sierra Madre Occidental and the Sierra Madre Oriental converge.

Mildred Bryant Brooks, The Pine Trees of Monterey, 1935, etching.

Frederick E. Church, Above the Clouds at Sunrise, 1849, oil on canvas. A North American pine forest in a romantic, hyperreal representation, a world away from the activities of the loggers and lumber companies who were harvesting trees for the transatlantic timber trade.
Pine regions to the east of the Atlantic display a quite different pattern. In contrast to the richness and diversity of North American pine flora, Europe north of the Alps is a region of pine poverty in which the only native pine of importance is Scots pine. It has an enormous range, the largest of any pine species, from Scotland in the west to the Russian Pacific coast in the east. East of the Urals, two other pine species occupy huge areas – Siberian dwarf pine and Siberian stone pine. Korean pine overlaps the borders of several countries in the far east of Asia.
China and Southeast Asia are rich in pine species, but cultural and linguistic barriers make their study difficult for Western botanists. The complexities of Chinese geography, with a topography that includes broken mountain ranges, deserts and river plains, and climates ranging from cold continental to subtropical, also makes studying the distribution of pines in the region a difficult task. The country has about 22 pine species, some of which (such as lacebark pine) are both ornamental and relatively rare. The barriers that make the study of East Asia difficult are reinforced in Southeast Asia, where the details and distribution of subtropical species such as Khasi pine (P. kesiya) and Krempf’s pine can be difficult to elucidate. Chinese and Southeast Asian pines are of great interest to specialists because the area is one in which new species are probably evolving. Japan has six common species, including Japanese black pine and Japanese red pine, both of which are valued as ornamental trees. The Himalayas, perhaps surprisingly in view of the association between mountains and pine trees, have few pine species, but these include the important chilgoza (P. geradiana), chir pine and Himalayan white pine (P. wallichiana).
The Mediterranean basin forms another pine region. Here, a long history of human interaction with the ecosystem in this area has obscured the natural ranges of pine species, but they are an important component of the vegetation and landscape.30 About a dozen species are native, depending on how the region is defined, and exactly which trees are accepted as species. Canary Island pine (P. canariensis) is considered among these. Both Iberia and the eastern Mediterranean have been suggested as areas of origin for Italian stone pine, and the western Mediterranean for maritime pine (P. pinaster). One of the most widespread species of this region is Aleppo pine (P. halepensis), the only pine to occur widely in North Africa. It divides into two genetic groups – eastern and western – with debates about how the distributions of these relate to the past and present.31 Other species, such as Austrian pine, are associated with mountains and have sometimes been subdivided so that, for instance, Corsican pine was in the past considered a separate species, Pinus laricio. The clearance and abandonment of land, fire and deliberate planting of pines for wood or pitch have all played their part in the complex history of pines in this region.

Ivan Shishkin, Morning in a Pine Forest, 1889, oil on canvas.

Shi Tao, View of Huangshan, 1670, ink-painted album leaf. Pine trees are a feature of the work of the Buddhist monk and artist Shi Tao, who was deeply impressed by the scenery of Huangshan.
The distribution of pine trees over the northern hemisphere is the product of millions of years of evolution. The fossil record is fragmentary, but the Pinaceae originated in the Upper Jurassic (c. 150 million years ago).32 Unambiguous pine fossils first appear in the early Cretaceous (c. 130 million years ago); the earliest, found in Belgium, is named Pinus belgica. On the other side of the world, Borneo, too, is rich in pine fossils. To state the changing distribution of pines with reference to present-day geography is futile because of the movements of landmasses over aeons of time. Some broad patterns can, however, be attributed to the distant past. A general division between pine species into ones with distinctly northern associations and those that belong to subtropical or tropical areas is probably due to the early Tertiary, a period of the Earth’s history when the climate was significantly warmer and wetter than at present. During this time, pine trees appear to have retreated from latitudes that have a presentday temperate climate to (cooler) northern ones, or to upland areas in the tropics, spreading back when the climate cooled, but leaving an inheritance of division between warm-climate and temperate pines that exists to the present day.33
The absence of pine species across the Great Plains of North America is attributed to the existence of a shallow sea running north–south during the late Cretaceous, resulting in distinctly different pine flora to the east and west of this divide. In Eurasia, the north coast of another ancient sea, the Tethys Sea, which once stretched from the edge of an embryonic mountain range (now the Himalayas) to somewhere off the present-day coast of North Africa, allowed the spread of geographically well-separated but related populations – Canary Island pine and chir pine (native to northern India).34 Ancient patterns of pine migration influenced by climate change during the Holocene ice ages are probably responsible for a pattern of pine distribution down the Malay peninsula onto Sumatra and into the Philippines, the trees moving southwards in the face of advancing glaciers as the climate cooled, and spreading back northwards once more as the climate warmed, colonizing new territory in the mountainous north of the island of Luzon.
The poverty of pine flora in northern Europe is also attributed to glaciation. Fossil evidence suggests that formerly a larger range of pine species was found in Europe north of the Alps, but glaciation pushed the trees southwards to meet a major barrier in the form of the Alps and mountains extending eastwards. Only Scots pine, a cold-tolerant species, survived this, probably in refugia (pockets of land with conditions locally clement enough for the trees to survive and reproduce). Once the ice retreated about 10,000 years ago, the pines spread northwards again.35 Arid cold was probably the factor that also limited the number of pine species found in eastern Eurasia.
Mary Anne Turner after Adam Pijnacker, Rocky Landscape, c. 1835, print. |
|

North America, with coastal plains and broken mountain ranges on the east side, and chains of high mountains running north–south on the west coast, provided much more scope for pine species to migrate north or south as the climate cooled and warmed again. The mountains on the west side of the continent were produced by tectonic activity that began 80 million years ago and still continues, providing varying habitats according to latitude and altitude. The fossil record of pine trees in Mexico is fragmentary, probably because this area was, and remains, seismically very active. It has also led to some of the most interesting and problematic aspects of the natural history of pine trees: Mexican pine species are highly variable, and botanists have always found them difficult to categorize.
European interaction with Mexico began with the conquistadors, following Columbus’s discovery of North America. They found the countryside of ‘New Spain’ challenging: ‘From Socochina we marched over a high mountain, through a pass, to Texutla’, wrote Bernal Díaz in 1519, and

Scots pines (P. sylvestris) at Rothiemurchus, Scotland, today. The pine, birch and juniper woodland has regenerated to a point where it appears natural and little sign of the early 19th century logging which took place here survives.
arrived into a rugged and wild mountain district . . . the first night we had excessive cold . . . and the wind so keen, which blew across the snow mountains, that we shook again with the frost; indeed, no one can wonder at this, for we had come so suddenly from the hot climate of Cuba, the town of Vera Cruz, and the neighbouring coast, into a cold country.36
As the Spanish discovered, steep mountains characterize southern Mexico – the product of geologically recent and continuing volcanicity. The Sierra Madre Oriental and Sierra Madre Occidental meet in a jumble of ridges and valleys and microclimates, with a rich genetic inheritance of pine species. The Spaniards were unaware of it, but they were moving through a region that would later be described, for its pine flora, as ‘a natural forest genetics laboratory for foresters, botanists, taxonomers and geneticists . . . [a] quite remarkable and infinitely precious natural experiment’.37
The botanical world did not take much interest in Mexican pine flora until the mid-nineteenth century. ‘In the year 1857 there was published in the City of Mexico a “Catalogue des Graines de Conifères Mexicains,” by B. Roezl & Cie., in which were described eighty-two new species of Pinus’, wrote George Russell Shaw (1848–1937), an American architect fascinated by the natural history of pines.38 Benedikt Roezl (1824–1885) was a native of Czechoslovakia and the son of a gardener. He went to Mexico looking for novel species to send back to European horticulturalists, and described over 82 species of pine he considered hitherto unknown. Only one of these was an ‘accidental hit’, still accepted by botanists as Lawson’s pine (P. lawsoni).39 Roezl’s catalogue confused thoughts about Mexican pines for decades. Enthusiastic and certainly unaware of a phenomenon elucidated by later botanists, Roezl
unintentionally made . . . an extremely important discovery; in the Mexican highlands is located a secondary center of evolution and speciation in the genus Pinus: hence the never-ending and utterly confusing variability of Mexican – Central American pines.40
Shaw, in his Pines of Mexico, wrote:
|
Resin flows from an immature cone of Mexican white pine (P. ayacahuite). |

A cursory examination of the specimens of Mexican Pines preserved in herbaria suggests a large number of species; the varied altitudes and climates of Mexico point to a like conclusion. Each excursion among the living trees, however, increased my suspicion that the numerous forms represent not many species but several varieties of new species.41
Much twentieth-century work on Mexican pines is inaccessible to the non-speaker of Spanish; a significant amount was carried out by Maximo Martinez (1888–1964) and Dr Jerzy Rzedowski (b. 1926), both of whom have trees named after them. The broken mountain ranges of Mexico and Guatemala give scope for rapid changes in altitude, rainfall and temperature: ‘myriad microclimates’ as Perry described them,42 and the pine forests of these countries contain mixtures of several species growing in relatively close proximity. Due to the topography, climate and vegetation, ‘Evolution in these pines is not something that only happened a million years ago, it happens every year.’43 Left to hybridize and grow under natural circumstances, the pine trees of Mexico would no doubt continue to evolve with their environment, but the area is under pressure from population growth, and threatened by exploitation for firewood, construction wood and pine nuts, and by general environmental degradation. Genetic diversity in so useful a tree is a low priority for an impoverished and growing population.