Chapter 19. Investigating Cell Division
Equipment and Materials
You’ll need the following items to complete this lab session. (The standard kit for this book, available from www.thehomescientist.com, includes the items listed in the first group.)
Materials from Kit
None
Materials You Provide
Microscope (oil-immersion if available)
Prepared slide: plant mitosis (onion tip)
Prepared slide: animal mitosis (optional)
Background
The cell division cycle (or cell cycle) is a fundamental biological process. To understand genetics and reproduction, you must first understand cell division. Prokaryotes use simple binary fission, during which one cell splits in two to form two identical cells. The cell cycle in eukaryotes, which we investigate in this lab session, is considerably more complex.
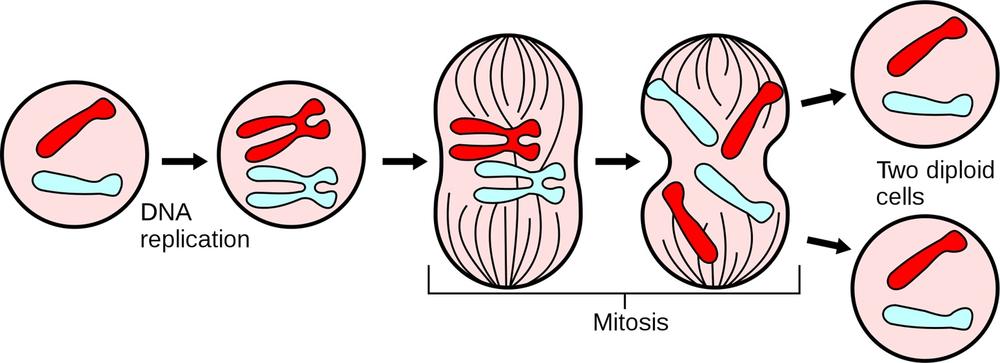
The eukaryotic cell cycle encompasses three major phases. During interphase, shown on the left in Figure 19-1, the cell grows, accumulates the nutrients needed for the next phase, and replicates its DNA. Interphase typically occupies about 90% of the cell cycle. During mitosis, shown in the center in Figure 19-1, the cell nucleus divides, separating its chromosomes into two identical sets and forming two separate nuclei from these chromosomes. Mitosis typically occupies about 10% of the cell cycle. During cytokinesis, the cell splits into two distinct daughter cells. The cycle then continues as these daughter cells individually enter interphase and continue to divide.
Note
Although cytokinesis is often thought of as the final step in the mitotic stage of the cell cycle, it is actually a separate and distinct stage of the cell cycle, occurring after the final stage of mitosis (telophase) and before the cell returns to interphase.
Interphase comprises three phases. During G1 Phase or Gap One Phase, the cell grows and carries on its routine activities. During S Phase, the cell duplicates its chromosomes, which as the cell enters this phase are dispersed and invisible with a standard microscope. Initially, each chromosome consists of only one chromatid. After DNA synthesis, each chromosome comprises two genetically identical sister chromatids, attached at the centromere. During G2 Phase or Gap Two Phase, the cell prepares itself for mitosis.
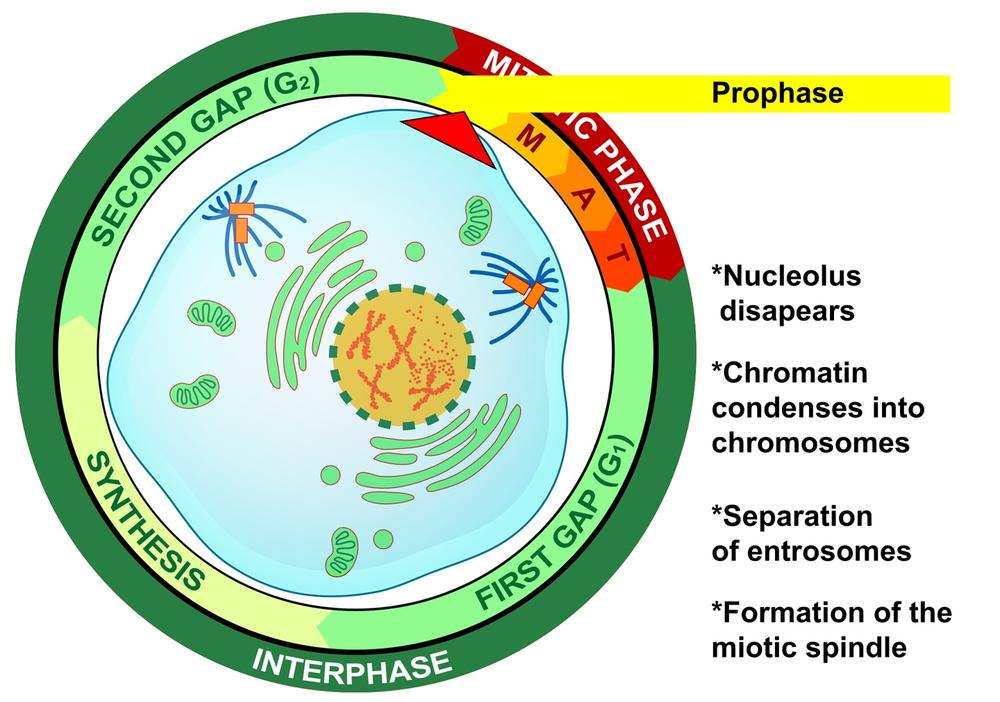
The exact sequence and timing of mitosis steps varies between cell types, but mitosis is conventionally divided into four or five phases. During the first phase, called prophase, the cell exiting the interphase second gap (G2) stage enters prophase and begins the mitotic sequence.
In early prophase, the chromatin condenses (coils) into chromosomes, which become visible with a standard microscope. As prophase continues, the two centrosomes (in animal cells, each made up of two centrioles) separate to opposite sides of the nucleus, the microtubules begin to lengthen and form the mitotic spindle, and the nucleolus disappears. Figure 19-2 is a graphic illustration of prophase, with the centrosomes shown in orange and the microtubules in blue. Figure 19-3 is an image of an actual cell in prophase.
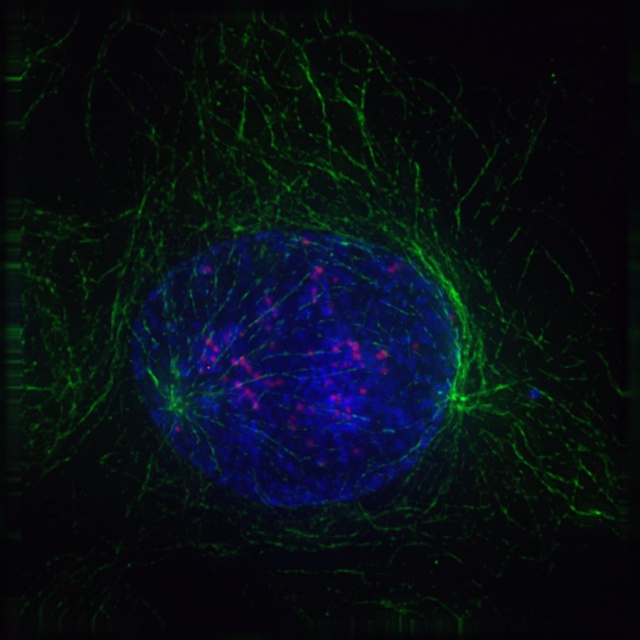
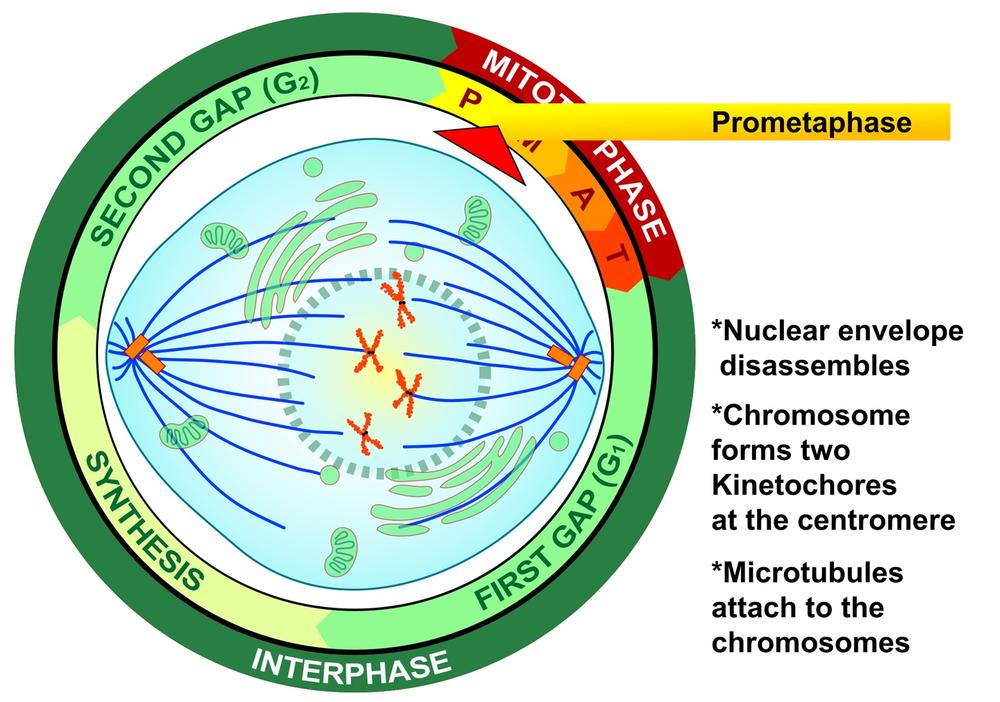
During the second phase of mitosis, called prometaphase, the nuclear membrane disintegrates, chromosomes form kinetochores at the centromere, and the microtubules that make up the mitotic spindle attach to the chromosomes. Figure 19-4 is a graphic illustration of prometaphase. Figure 19-5 is an image of an actual cell in prometaphase, with the mitotic spindle in green, the condensed chromosomes as a cloud of blue, and the kinetochores and centromeres in magenta.
Note
Some texts describe mitosis as occurring in four phases: prophase, metaphase, anaphase, and telophase. Other texts place an additional phase, prometaphase, between prophase and metaphase. Prometaphase incorporates what four-phase texts include in late prophase and early metaphase. The reality is that the divisions between phases are somewhat arbitrary, and it doesn’t really matter if you think of mitosis as having four or five phases.
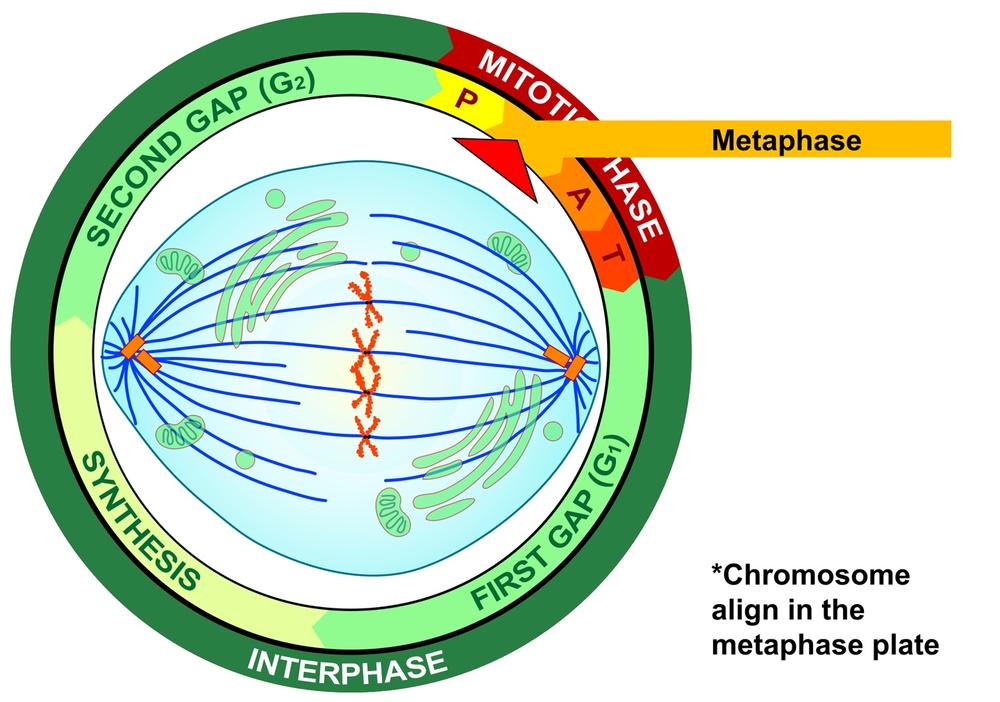
During the third phase of mitosis, called metaphase, the centromeres of the condensed and coiled chromosomes align in an imaginary plane called the metaphase plate (also called the equatorial plate) located at the equator of the cell, with the telomeres (chromosome ends) drifting away from the equator toward the centrosomes at the poles. This even alignment physically organizes the cell in preparation for later division into two daughter cells. At this point, and before entering the next mitotic phase, the cell does a literal “self-test” to ensure that every chromosome is properly aligned at the metaphase plate and that every kinetochore is properly attached to the mitotic spindle. Figure 19-6 is a graphic illustration of metaphase.
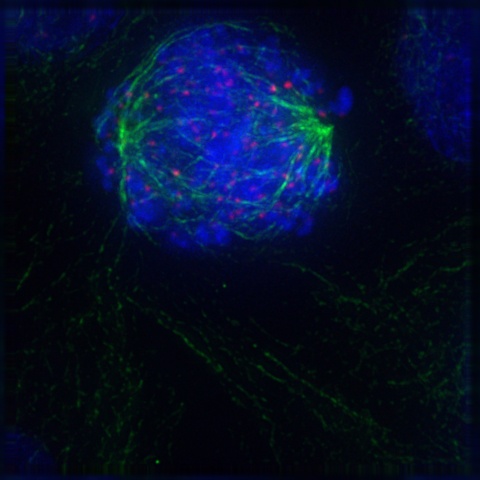
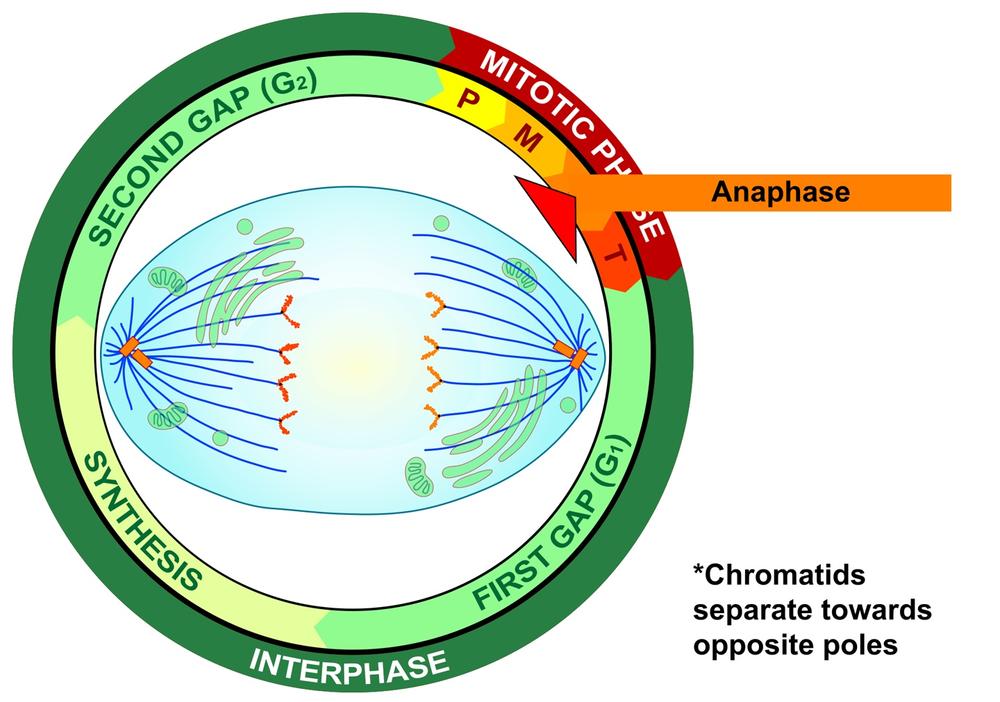
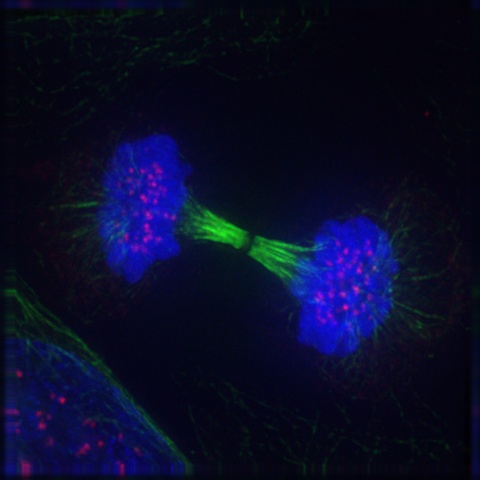
If and only if the cell passes its metaphase self-test, it enters the fourth stage of mitosis, called anaphase. During anaphase, the microtubules that make up the mitotic spindle attach to the centromere of each chromosome and literally pull the sister chromatids of each chromosome apart toward the centrosomes at each pole, with the telomeres of each chromosome pointing toward the equatorial plate. Those microtubules that are not attached to kinetochores act to stretch the cell lengthwise into an oval form. Figure 19-7 is a graphic illustration of anaphase. Figure 19-8 is an image of an actual cell in anaphase.
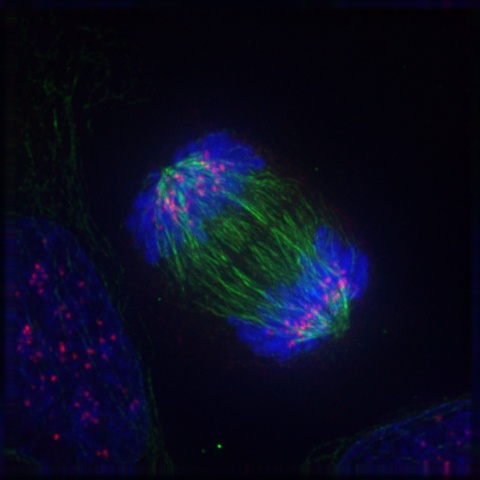
The final phase of mitosis, telophase, begins when anaphase is complete. Essentially, telophase is the “clean-up” phase. The cell continues to elongate. Chromosomes uncoil and disperse as chromatin. The mitotic spindle breaks down and microtubules disintegrate. Two daughter nuclei form, one on either side of the ovular cell, with their nuclear membranes forming from the fragments of the original nuclear membrane. As the new nuclear membranes form around each chromatid pair, new nucleoli form. In animal cells, a contractile ring of fibers called a cleavage furrow develops. which during cytokinesis closes to pinch off the binucleate cell into two individual daughter cells. In plant cells, cell wall materials coalesce into a structure called the cell plate, which develops into a full cell wall, separating the two nuclei. Figure 19-9 is a graphic illustration of telophase. Figure 19-10 is an image of an actual cell in telophase.
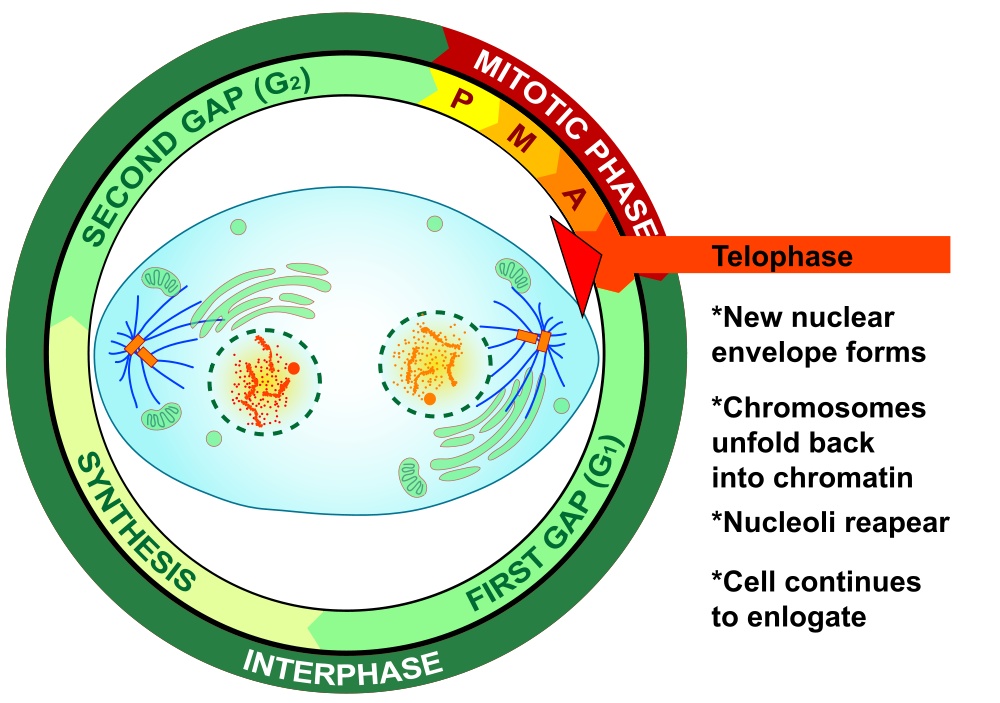
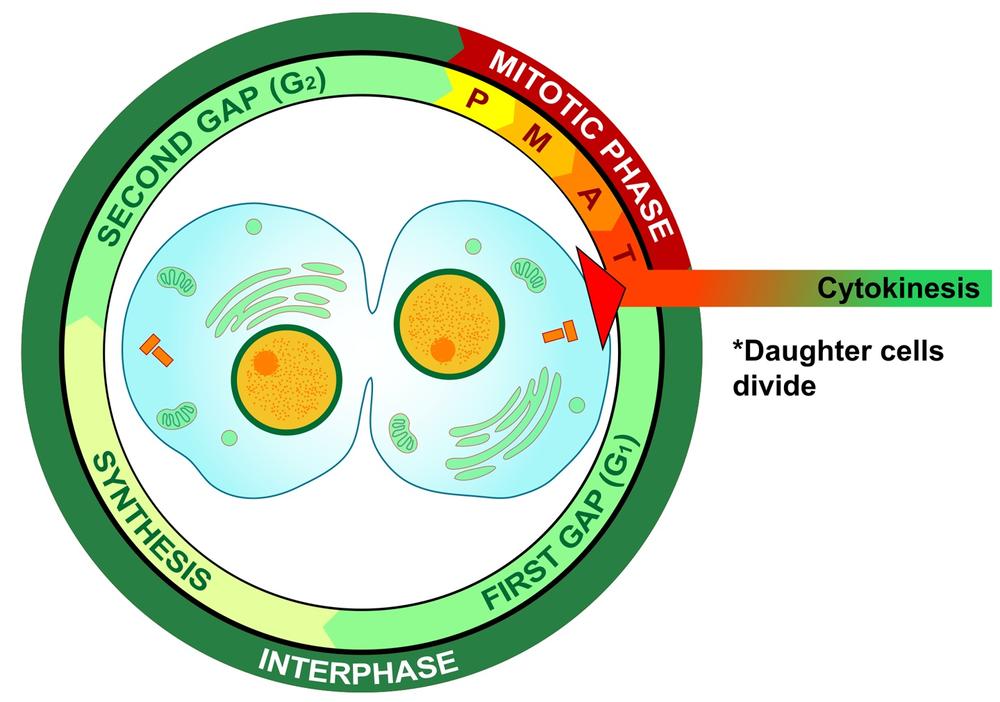
Finally, cytokinesis, shown graphically in Figure 19-11, occurs when the cell that has undergone mitosis splits into two new individual daughter cells, each of which then enters interphase to start the cycle anew.
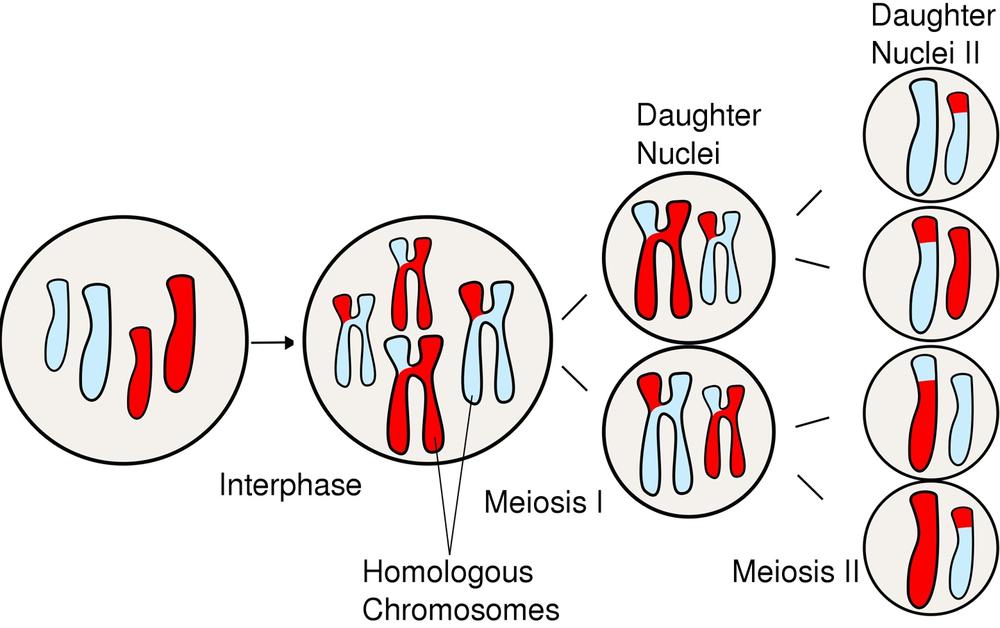
Meiosis is a specialized type of cell division that occurs in eukaryotes to produce the gametes (sperm cells and egg cells, produced by all animals and some protists) or spores (in all plants and fungi and some protists) required for sexual reproduction. Because of the similarity of names, students sometimes confuse mitosis and meiosis. Although there are similarities in the mechanisms and the names of the phases, the two processes are distinctly different in several respects:
Mitosis is the asexual reproduction of diploid cells; meiosis is the production of haploid cells (gametes or spores) in preparation for sexual reproduction.
Mitosis is a part of the cell cycle, a circular process with no beginning or end. New cells produced at the end of the cell cycle restart the cycle to produce still more new cells. Meiosis is a process with a starting point and an ending point. In effect, mitosis runs in a circle, while meiosis runs irreversibly in a straight line from point A to point B.
Mitosis produces daughter cells that are genetically identical to the mother cell, with no reshuffling of genetic material. In meiosis, the chromosomes recombine, shuffling the genes to produce different genetic combinations in each gamete.
Mitosis produces two genetically identical diploid cells; meiosis produces four genetically distinct haploid cells.
Meiosis starts with one diploid cell that contains two copies of each chromosome, one from each parent of the organism. (For example, your cells contain 23 pairs of chromosomes, with one chromosome of each of the 23 pairs originating from your mother and one from your father.) Meiosis produces four haploid cells (gametes), each of which has one copy of each chromosome. Each of those chromosomes is a unique mixture of the paternal and maternal DNA from the organism. This mixing of genetic material ensures that offspring are genetically distinct from both parents, thereby ensuring diversity in any population that reproduces sexually.
Chromosomes in ordinary eukaryotic cells occur in pairs. The nuclei of such cells are referred to as diploid (2N). The two chromosomes of each pair are called homologous chromosomes. Each of those chromosomes has the same genes at the same positions, called loci (singular locus), although each of the homologous genes may have different alleles (different variants of the same gene) at the corresponding homologous loci. Chromosomes in specialized reproductive cells (gametes and spores) are unpaired, and the nuclei of such cells are called haploid (1N).
In preparation for meiosis, a cell undergoes a meiotic interphase stage that is similar to the mitotic G1, S, and G2 stages. The cell’s chromosomes are duplicated via DNA replication, producing the homologous chromosomes in paternal and maternal versions, each of which comprises two exact copies, called sister chromatids, attached at the centromere.
Like the cell cycle (including mitosis), meiosis is a seamless and continuous process, but is for convenience normally treated as occurring in phases. The names of those phases are the same as those of the cell cycle—prophase, metaphase, anaphase, telophase, and cytokinesis—and those phases are similar to the corresponding cell-cycle phases in mechanism and appearance. The major difference is that while the cell cycle involves only one division, meiosis involves two divisions, called Meiosis First Division (or Meiosis I) and Meiosis Second Division (or Meiosis II). During Meiosis I, the parent cell divides into two diploid daughter cells. During Meiosis II, the two diploid daughter cells divide into four haploid daughter cells The meiotic phases are as follows:
Prophase I – homologous chromosomes condense and form pairs, with crossing over.
Metaphase I – the microtubular structure forms, with the microtubules attaching to the chromosomes.
Anaphase I – homologous pairs of chromosomes separate and the cell elongates.
Telophase I – one set of paired chromosomes completes its migration to each pole.
Cytokinesis I – the binucleate cell splits into two daughter cells, with each daughter cell receiving one of each pair of homologous chromosomes.
Prophase II – the meiotic spindle forms between the two centrioles.
Metaphase II – chromosomes align along the microtubules of the spindle.
Anaphase II – sister chromosomes separate.
Telophase II – chromatids arrive at each pole.
Cytokinesis II – cell division is complete, with four daughter cells resulting, each with half the number of chromosomes of the parent cell.
Figure 19-12 graphically illustrates the major events in meiosis.
In this lab session, we’ll observe the cell cycle by examining prepared slides that are stained to show the stages of mitosis in plant and animal cells. Although the stages of meiosis can also be observed by examining prepared slides, there’s little point to doing so because the physical appearance of the various meiotic phases is nearly impossible to discriminate microscopically from the corresponding mitotic phases.
Procedure IV-3-1: Observing Mitosis
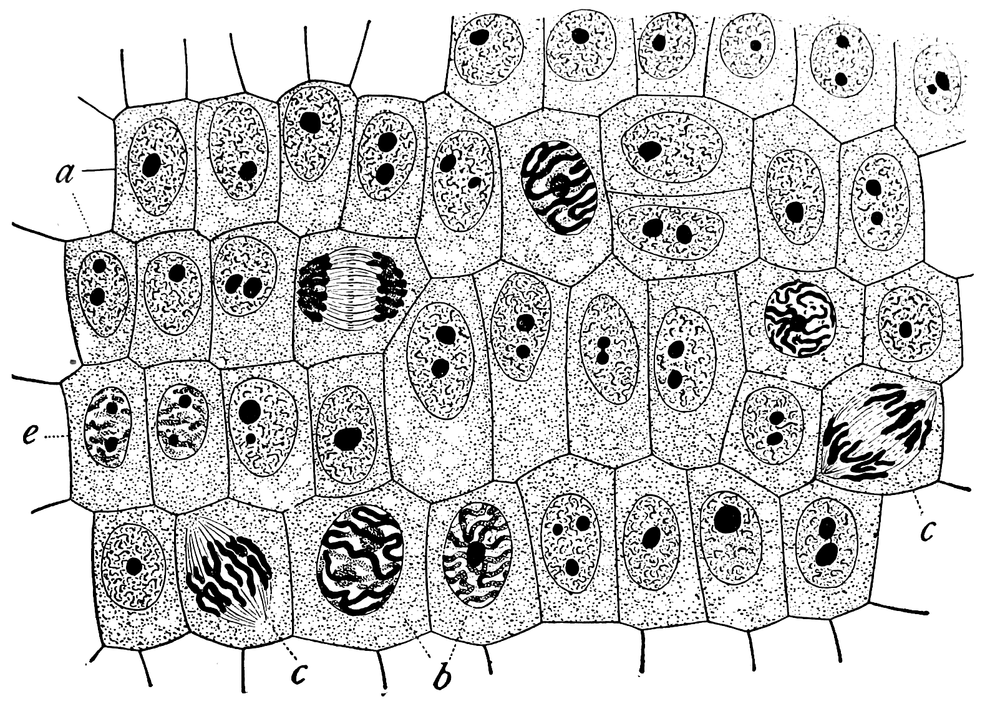
Students are often surprised that mitotic phases can actually be seen and discriminated with a microscope, but the staining protocols for doing so have been in use since the 19th century. Figure 19-13 is a sketch that appeared in the 1900 book The Cell in Development and Inheritance by Edmund B. Wilson. The original description for this sketch read, “General view of cells in the growing root-tip of the onion, from a longitudinal section, enlarged 800 diameters. a. non-dividing cells, with chromatin-network and deeply stained nucleoli; b. nuclei preparing for division (spireme-stage); c. dividing cells showing mitotic figures; e. pair of daughter-cells shortly after division.” Comparing Wilson’s sketch with the graphics and images in the preceding section, it’s clear that mitotic phases were well understood (albeit under different names) more than 100 years ago.
The problem with observing mitosis is that the objects you are viewing are both tiny and three-dimensional. That means you’ll need to use high magnification (400X or 1,000X), and that you’ll spend a lot of time tweaking the fine focus to accommodate the very shallow depth of focus at high magnification.
As you perform this procedure, record your observations in your lab notebook. Identify the features that should be present in each mitotic phase and verify their presence. Sketch the significant features you observe that are pertinent to each mitotic phase.
In many plants, including Allium (onion), many or most of the cells undergo mitosis. If a specimen is properly sectioned and stained to show mitosis (usually with toluidine blue or Schiff’s reagent), the various stages of mitosis are clearly visible in different cells in the section.
Obtain a prepared slide of an Allium root tip in longitudinal section, stained to show mitosis.
Scan the slide at low magnification to identify an area where mitosis is occurring (or was, at the time the slide was made). That area is one in which cell nuclei have distinctly differing appearances, which are visible even at 40X. Some will appear small, compact, and roughly circular, while others will appear broken or fuzzy.
Center that area in the field of view and change to high-dry (400X) magnification. Note the distinctly different appearance of many of the cell nuclei in the field of view. Do a count of the cells visible in the field of view, judging which mitotic phase each of the cells is in.
Here’s what you’ll actually see. Figure 19-14 through Figure 19-18 show the same 400X field of view of a longitudinal section of an onion root tip stained with toluidine blue, with several stages of mitosis visible. The visible differences among these images are caused by very minor changes in fine focus, one or two units on our 100-unit fine-focus knob. Even a thin section has depth, and the extremely small depth of focus at 400X means you’ll need to tweak your fine focus constantly to bring different parts of the same nucleus into focus.
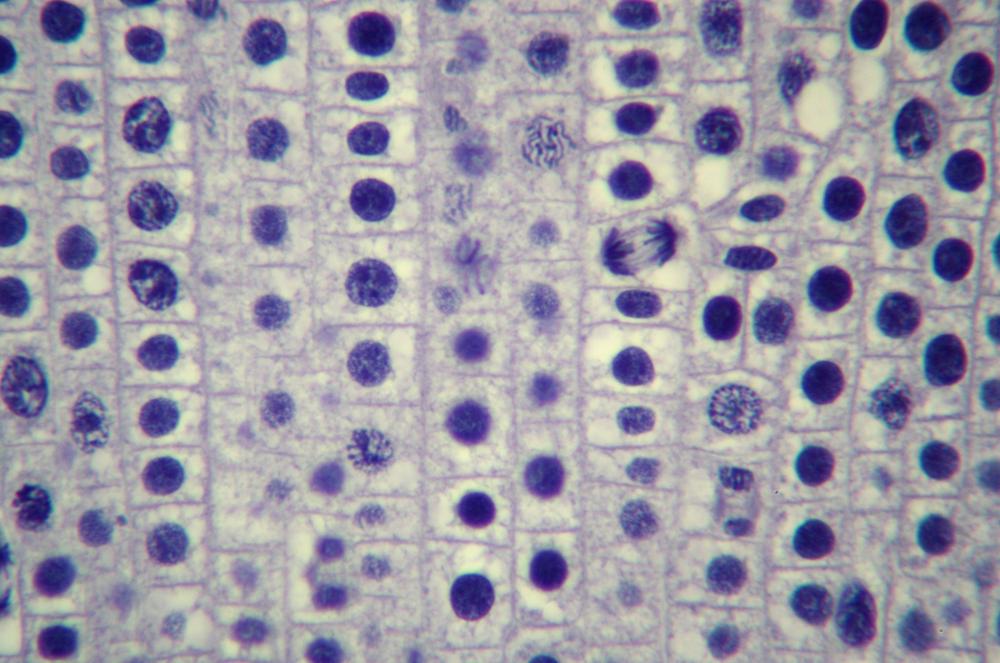
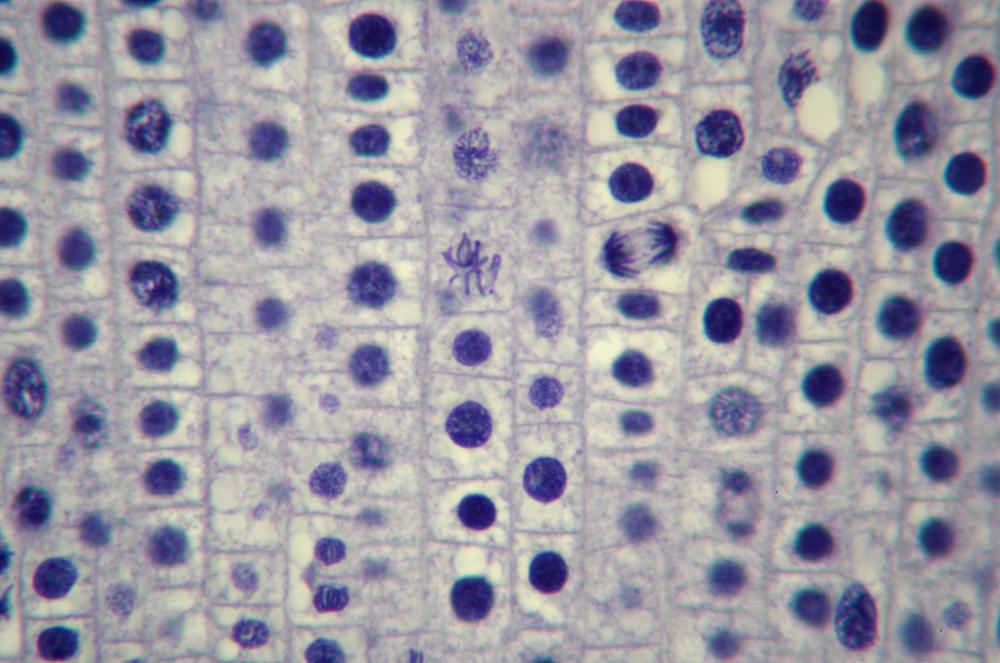
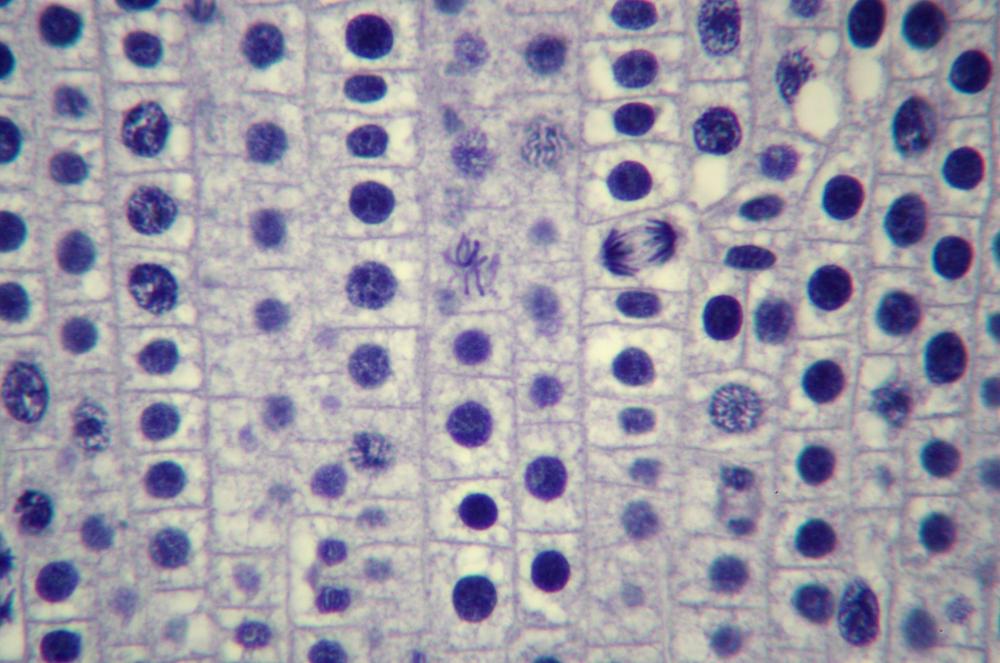
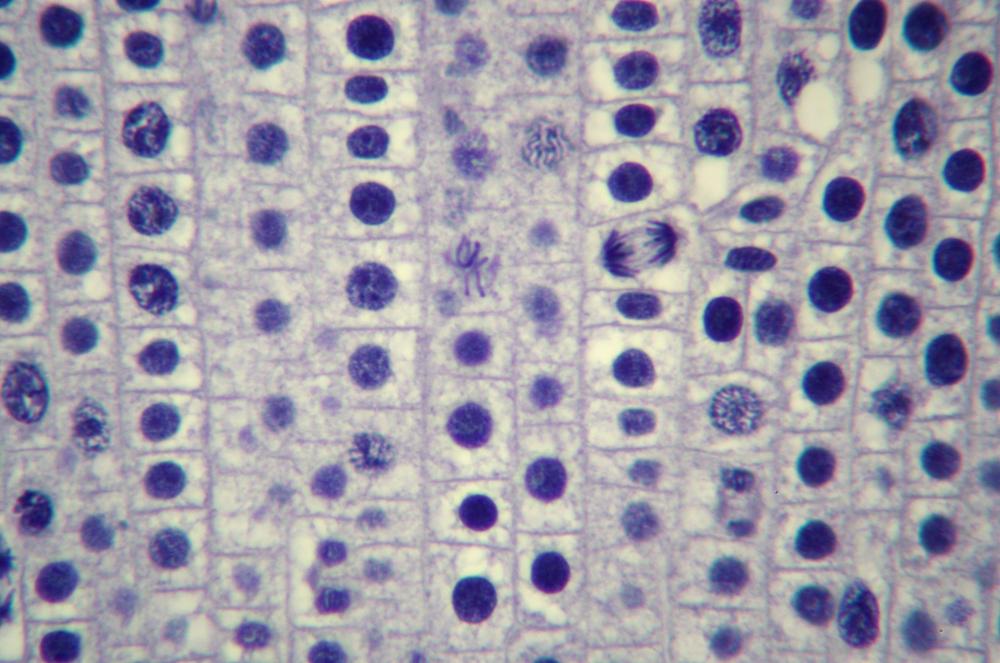
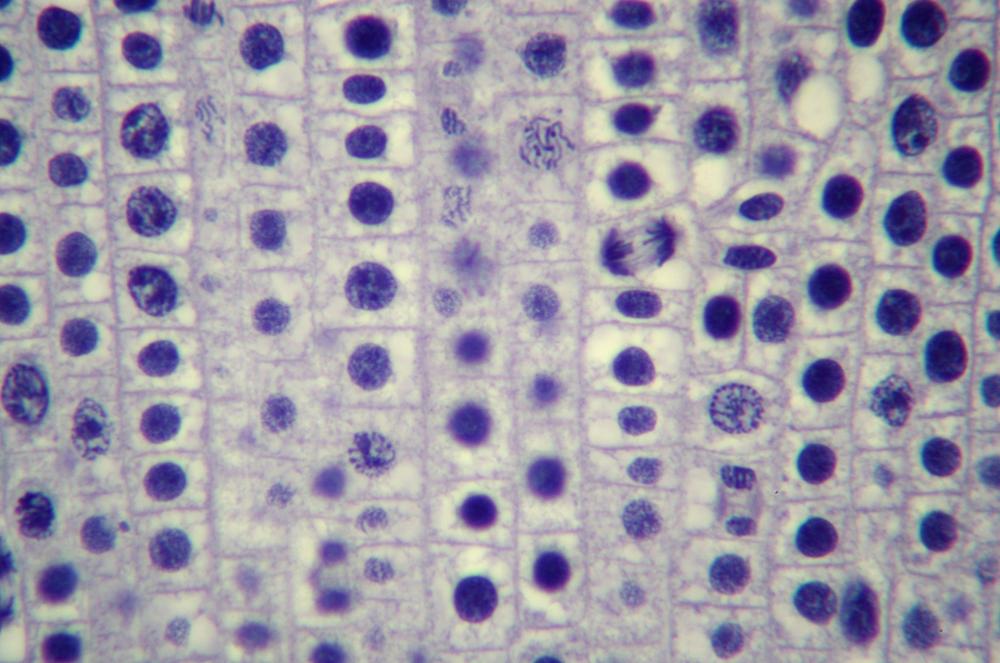
Locate a nucleus that is in any of the five stages of mitosis or one that is in cytokinesis. (You needn’t observe nuclei “in order”; for example, if the nucleus nearest the center of the field in in telophase, work with telophase first rather than searching out a nucleus in prophase.)
Observe that nucleus at 400X, tweaking the fine focus as necessary to bring all features into sharp focus sequentially. Identify as many as possible of the features that should be present in that stage. If possible, count the chromosomes. Make a sketch of that phase.
If your microscope has an oil-immersion objective, center the nucleus in question carefully and change to 1,000X magnification to see if any more detail is visible.
Note
You don’t have to dance with the girl you brung. For example, the first nucleus you pick may turn out to be a damaged telophase. Just move along to the next nucleus, whether or not it’s telophase. Keep scanning for “good” examples of each phase until you’ve critically examined and sketched at least one nucleus in each phase.
Repeat steps 4 through 6 for each of the other stages of mitosis in plants.
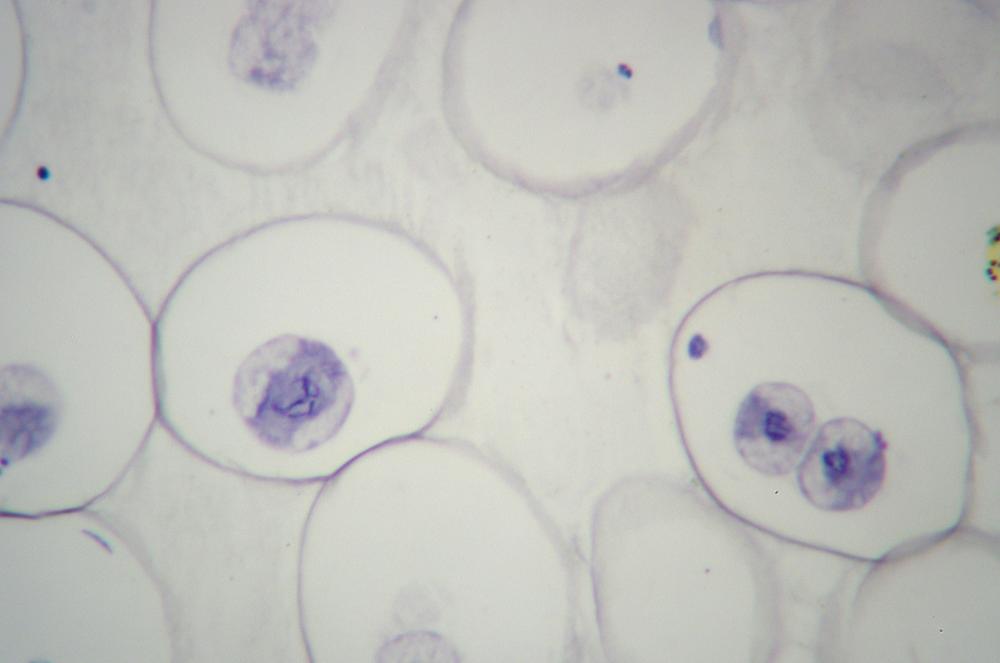
Obtain a prepared slide of an animal cell section (such as the traditional whitefish blastula or Ascaris), stained to show mitosis. Figure 19-19 shows mitosis in a roundworm (Ascaris sp.) egg at 400X.
Repeat steps 2 through 7 to observe all of the stages of mitosis as well as cytokinesis.
Review Questions
Q1: Does mitosis occur throughout the onion root tip or in only one specific part of it? How do you know?
Q2: When you counted the onion root tip cells, did you find that one or more phases occurred more frequently in your field of view than others? Did you expect this result? Why or why not?
Q3: Why do some of the onion root tip cells have little or no chromatin visible?
Q4: Were you able to observe more detail at 1,000X than was visible at 400X?
Q5: The diploid number of chromosomes in Allium is 16. Did your observations of a cell in anaphase confirm this number? If not, propose an explanation.
Q6: Although mitosis has been widely studied since the 19th century, the actual number of human chromosomes (46, in 23 pairs) was not definitively determined until 1956. Propose an explanation.
Q7: What structures are present in animal mitosis that are not present in plant mitosis?
Q8: What, if any, difference is there in the duration of mitosis in plants versus animals? How can you tell?
Q9: How does the division of daughter cells differ between animal mitosis and plant mitosis?
Q10: In metaphase, what is the chromatin material that aligns on the equatorial plane for mitosis, meiosis I, and meiosis II?
Q11: In anaphase, what is the chromatin material that aligns on the equatorial plane for mitosis, meiosis I, and meiosis II?