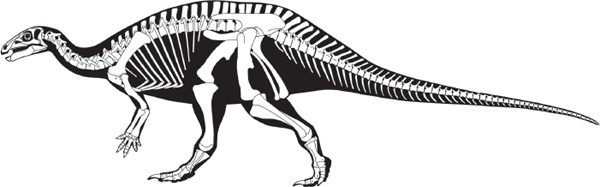
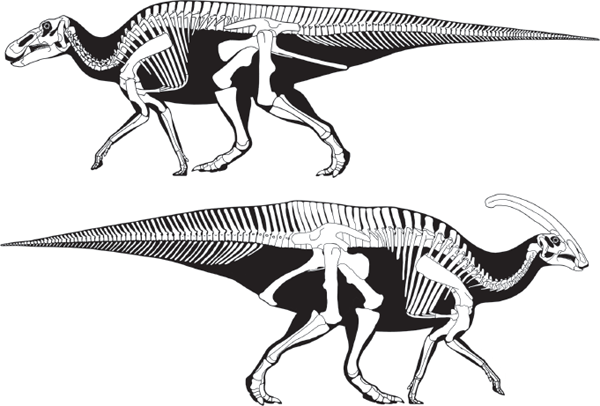
ORNITHOPODS (bird feet) were bipedal ornithischian dinosaurs that lacked body armor (see
box 5.1). They first appeared during the Middle Jurassic and were among the last dinosaurs to become extinct at the end of the Cretaceous. Their fossils are found on all continents. Ornithopods were thus one of the longest-lived dinosaur groups, and they were also one of the most diverse.
Several distinctive clades of ornithopods have been recognized (
figure 7.1), from the primitive ornithopods to the enormous iguanodontians and their close relatives, the hadrosaurids (commonly referred to as the duck-billed dinosaurs). Many features of the skull were evolutionary novelties of the ornithopods, including the cranial kinesis (joints within the skull), the ventrally offset premaxillary tooth row (offset with respect to the maxillary tooth row), the closed or reduced external mandibular fenestra, the lower jaw joint location well below the level of the maxillary and lower tooth rows, and the slender process on the ischium. In this chapter, I review the anatomy and evolution of the ornithopods.
Paleontologists disagree about the phylogenetic position of the heterodontosaurs. They are variously considered very primitive ornithopods, primitive ornithischians, or related to marginocephalians. Although discussed here with ornithopods, we regard them as primitive ornithischians.
Heterodontosaurs are best known from the Lower Jurassic of southern Africa. These small (about 1 to 1.5 meters long) dinosaurs had uniquely shaped cheek teeth that were chisel shaped with tiny cusps (denticles) restricted to the apex of the crown (
figures 7.2 and
7.3). The name “heterodontosaur,” which means “different-toothed lizard,” is based on the canine-like teeth in the front of the mouth (see
figure 7.2), which had a markedly different shape from the chisel-shaped cheek teeth.
FIGURE 7.1
This cladogram of the ornithopods divides them into several clades. (Drawing by Network Graphics)
FIGURE 7.2
This skull of Heterodontosaurus well represents a primitive ornithischian. (© Scott Hartman)
FIGURE 7.3
The chisel-shaped cheek teeth of heterodontosaurs are unique among dinosaurs. (Drawing by Network Graphics)
FIGURE 7.4
Heterodontosaurus, from the Lower Jurassic of southern Africa, was a typical heterodontosaur. (© Scott Hartman)
Heterodontosaurus was a characteristic heterodontosaur (
figure 7.4). This 1.2-meter-long dinosaur had short forelimbs and relatively long hind limbs, indicating it was a biped. The cheek teeth formed a
dental battery set in massive lower jaws hinged to a solidly built skull. The teeth in this dental battery were usually heavily worn. The heavy wear and the chisel shape of the teeth indicate this dinosaur was a plant-eater. Paleontologists disagree as to how
Heterodontosaurus moved its jaws when chewing; either the lower jaw was moved backward and forward, or it was rotated against the upper jaw when the mouth closed.
The long tail and short neck of Heterodontosaurus were features typical of primitive ornithischians. Ossified tendons were present only in the back region. A very bird-like feature of the hind-limb skeleton of Heterodontosaurus was the fusion of the tibia and fibula and their fusion to the tarsals. This fusion stabilized the lower leg and ankle and suggests that Heterodontosaurus was a fast runner. In contrast, the hands of Heterodontosaurus were stout and flexible; they may have been used to dig up or grasp vegetation.
Although
Heterodontosaurus was a plant-eater, it had large tusks near the front of its jaws (see
figure 7.2). These were probably used for defense and display, as are the tusks of some living, large plant-eating mammals, such as pigs. Some heterodontosaur fossils lack these tusks, suggesting that they may have been a sexually dimorphic feature. Males presumably had tusks, and females did not. The tusks might also have been used in feeding, helping the dinosaur to stab and tear vegetation.
Heterodontosaurs are known from the Upper Triassic of Argentina, the Lower Jurassic of southern Africa, the Jurassic of North America, the Cretaceous of Asia, and possibly the Cretaceous of Europe. Their spotty fossil record suggests that many heterodontosaurs remain to be discovered. Most intriguing is Tianyulong, a heterodontosaur recently described from the Lower Cretaceous of northeastern China. The fossil of this dinosaur has filaments preserved in the skin below the neck and above the back and tail. Although these have been called “protofeathers,” their actual relationship to feathers awaits further study.
FIGURE 7.5
Early Cretaceous Hypsilophodon was a typical primitive ornithopod. (© Scott Hartman)
The most primitive ornithopods were long assigned to Hypsilophodontidae, a single family of small to medium-size (2 to 4 meters long) bipedal ornithischians. However, this formerly monophyletic family is now regarded as an array of primitive ornithopods, some more closely related to iguanodontians than others. Late Jurassic to Upper Cretaceous fossils of these primitive ornithopods have been discovered, and the fossils of primitive ornithopods are found on every continent. Best known and typical of the primitive ornithopods is
Hypsilophodon (
figure 7.5).
The limb proportions of Hypsilophodon were those of a bipedal runner. The hind limbs were much longer than the forelimbs, and the distal segments of the hind limb, especially the ankle, were elongate. The long tail, stiffened by calcified tendons, would have counterbalanced the dinosaur while it ran. Although some paleontologists previously believed that Hypsilophodon was a tree-climbing dinosaur, its skeleton is much more that of a fast ground runner.
The skull of
Hypsilophodon resembled that of
Heterodontosaurus but lacked the large tusks. The eyes were large, and a narrow, horny beak was present at the tip of both the upper and lower jaws. The massive jaws of
Hypsilophodon supported a dental battery of interlocking cheek teeth that wore down to produce a continuous, inclined cutting edge. Unlike heterodontosaurs, there was a hinge in the skull of
Hypsilophodon that ran from near the front of the tooth row to near the jaw joint on the posterior corner of the skull. This hinge allowed the jaw to move from side to side as the dinosaur chewed (
box 7.1).
Primitive ornithopods are extremely well known from complete skeletons and eggs. The zenith of their diversity was during the Late Jurassic and Early Cretaceous when these small, bipedal herbivores lived on all continents. Only two Late Cretaceous primitive ornithopods are known, Thescelosaurus and Parksosaurus; their fossils have been found in western North America. The success of the primitive ornithopods was probably due in large part to their ability to grind vegetation.
Many living mammals that eat plants grind their food by moving their lower jaws from side to side while chewing. This transverse movement of the lower jaw is achieved by arranging the jaw muscles and their points of attachment to pull the jaw laterally and medially. In tandem with teeth that provide flat surfaces for grinding and sharp edges for tearing, side-to-side movement of the lower jaw efficiently mills vegetation prior to swallowing.
Ornithopod dinosaurs efficiently milled the vegetation they ate with a mechanism analogous to, but different from, that of living mammals.
Hypsilophodon is a good example. First, instead of moving its lower jaws transversely,
Hypsilophodon had a joint within its skull that allowed the upper jaws to move sideways while chewing (
box figure 7.1). This joint was essentially a diagonal hinge between the premaxillary, the upper jaw bone at the front of the mouth, and the maxillary, the upper jaw bone behind it. When the dinosaur closed its mouth, the teeth sheared past each other and the maxillary rotated slightly outward while the lower jaw rotated inward. Thus, while
Hypsilophodon chomped on vegetation, the outwardly rotating upper jaw produced a grinding movement.

BOX FIGURE 7.1
The skull of Hypsilophodon had a joint that allowed the upper jaws to rotate outward when the mouth was closed. The bottom diagram shows the joints between the main elements of the skull of the dinosaur. (© Scott Hartman)
Hypsilophodon had broad, chisel-like teeth that locked together along their crowns to form a continuous cutting edge. A cheek pouch may have been present alongside the teeth to catch and hold food while chewing. This sophisticated mechanism for milling vegetation was very different from the generalized teeth and jaws of primitive ornithischians, such as the heterodontosaurs, dinosaurs that were otherwise rather similar to Hypsilophodon. The mechanism of grinding vegetation must have been one of the keys to ornithopod success.

FIGURE 7.6
Late Jurassic Dryosaurus was a primitive iguanodontian. (© Scott Hartman)
All of the advanced ornithopods are classified in Iguanodontia. They are distinguished by their transversely widened premaxillaries that lacked teeth, deep dentaries, and the loss of a phalanx from the third digit of the hand. Here, we distinguish primitive iguanodontians, well represented by Tenontosaurus, Camptosaurus, and the classic Iguanodon, from the advanced iguanodontians, the hadrosaurids.
Dryosaurus, from the Upper Jurassic of North America and Africa, well represents the primitive iguanodontians (
figure 7.6). It ranged in size from 2 to 3 meters long and had a short skull, long neck, long tail, and three instead of four phalanges in the third finger of the hand, and lacked premaxillary teeth. In many ways,
Dryosaurus represents an ideal transitional form between the small “hypsilophodontids” and the large iguanodontids.
Camptosaurus, from the Upper Jurassic of North America and Europe, is well known from many complete skeletons and is the oldest heavily built ornithopod (
figure 7.7). This relatively small (5 to 7 meters long) iguanodontian did not have a fully developed “thumb” spike and had small hooves on the tips of all its toes.
Camptosaurus had four digits on the hind foot, massive hind limbs, and limb proportions that suggest it was a facultative quadruped. The fused wrist bones provide strong evidence of stable hand walking. The neck of
Camptosaurus was relatively long, and its head was relatively small for an iguanodontian. But, it had the long, horse-like snout and broad, toothless beak characteristic of all iguanodontians.
FIGURE 7.7
Late Jurassic and Early Cretaceous Camptosaurus is the oldest iguanodontian known from a complete skeleton. (© Scott Hartman)
Iguanodontians have a special place in the history of dinosaur studies because
Iguanodon was one of the first dinosaurs to be described scientifically, by Gideon Mantell in 1825 (see
chapter 11).
Iguanodon was as much as 10 meters long and mostly a bipedal herbivore that lived principally during the Late Jurassic and Early Cretaceous in Europe and possibly Asia.
Best known from the Lower Cretaceous of Belgium,
Iguanodon was a much larger and more specialized iguanodontian than
Camptosaurus (
figure 7.8;
box 7.2). Like
Camptosaurus, it had massive hind limbs. But, unlike
Camptosaurus,
Iguanodon had long forelimbs that were 70 to 80 percent as long as its hind limbs. The wrist bones of
Iguanodon were fused, and the central three digits of the hand ended in hooves. The large conical “thumb” spike on the first digit must have been used as a defensive weapon, and the fifth digit was long and slender.
Iguanodon had a large head with a long snout, a long neck and long tail, and an extensive boxwork of ossified tendons that extended along the back from the shoulder region to the middle of the tail (see
figure 7.8). The limb proportions and modifications of the hands of
Iguanodon suggest that it did much more quadrupedal walking than did
Camptosaurus or the smaller ornithopods. It seems likely, however, that
Iguanodon could rear up on its hind limbs and swing its spike-shaped thumb like a knife fighter when attacked.
The skulls of iguanodontians varied greatly, but all had long snouts and toothless beaks. These long snouts included many more teeth (as many as 29 in the maxillary of
Iguanodon) than did the snouts of heterodontosaurs and the primitive ornithopods. The iguanodontian beak was used to crop vegetation. The teeth were leaf shaped with long ridges on the sides and small cusps on the cutting edges and resembled the teeth of living iguanas: hence the name of the dinosaur taxon (see
chapter 11). Paleontologists are certain that iguanodontians were plant eaters.
FIGURE 7.8
Early Cretaceous Iguanodon was a powerful quadrupedal walker. (© Scott Hartman)
Bernissart is a small mining town in southwestern Belgium. In 1878, miners there discovered a crevice full of clay that cut across a bed of coal and was full of dinosaur bones. This crevice, at a depth of 322 meters, was excavated from 1878 until 1881. The fossils of 39 individuals of
Iguanodon, many of them complete skeletons (
box figure 7.2), were recovered, as were the fossils of many plants, fishes, and other reptiles. Bernissart thus stands as one of the greatest dinosaur localities on Earth.
BOX FIGURE 7.2
Sketch of one of the many complete skeletons of Iguanodon from Bernissart as it was found in the rock. (From L. Dollo. 1882. Première note sur les dinosauriens de Bernissart. Bulletin du Musée royal d’histoire naturelle de Belgique 1:55–74)
For many years, paleontologists thought a single catastrophe led to the accumulated Iguanodon skeletons at Bernissart. One popular image shows the dinosaurs falling off a cliff into the crevice, which later filled with clay. Indeed, the “evidence” of a single catastrophe at Bernissart suggested to some paleontologists that Iguanodon lived in groups (“herds”).
Careful restudy of the Bernissart Iguanodon, however, indicated that the Iguanodon skeletons accumulated in groups on at least three different occasions. The skeletons were buried and fossilized in a marshy environment, but the precise cause of death of the dinosaurs remains uncertain. What is certain, though, is that a single catastrophe did not kill the Bernissart Iguanodon, so the accumulation of skeletons here does not necessarily indicate that Iguanodon lived in herds.
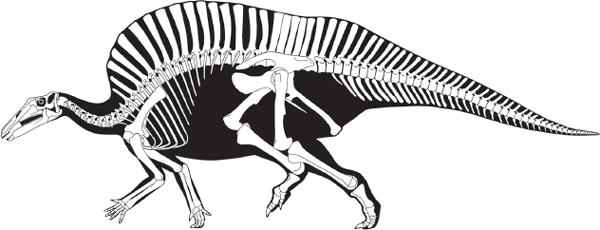
FIGURE 7.9
Sail-backed Ouranosaurus, from the Middle Cretaceous of Niger, was one of the most distinctive iguanodontians. (© Scott Hartman)
The zenith of primitive iguanodontian diversity was during the Early to Middle Cretaceous, and iguanodontian fossils are known from Europe, North America, Africa, Asia, and Australia. A particularly unusual iguanodontian from this time period is
Ouranosaurus from Niger with its very distinctive head and long neural spines that formed a sail along its back (
figure 7.9). Clearly, primitive iguanodontians were a diverse, widespread, and significant group of medium to large herbivorous dinosaurs of the Early Cretaceous world. There was a marked decline of primitive iguanodontians during the Late Cretaceous, probably because of the appearance of the hadrosaurids.
Advanced iguanodontians belong to a single family: Hadrosauridae. Hadrosaurids, popularly known as the duck-billed dinosaurs, first appeared during the Late Cretaceous and thus were the last major group of ornithopods to evolve. Much more is probably known about hadrosaurids than about any other single group of dinosaurs. Their outstanding fossil record includes many complete skeletons, eggs, footprints, and coprolites and several mummified individuals with skin intact.
The hadrosaurids were large ornithopods, 7 to 12 meters long (
figure 7.10), characterized by their broad beaks lacking teeth, intricate dental batteries in which three or more replacement teeth existed for each tooth position (
figure 7.11), and the loss of the first digit (thumb) on the hands, among other features. The jaws were deep and long, and the jaw muscles were evidently powerful. Hadrosaurids chomped their food and achieved a side-to-side motion of the jaws by slightly rotating the upper tooth rows outward when the mouth was closed.
Hadrosaurid teeth were arranged in dental batteries consisting of literally hundreds of teeth cemented together in each jaw (see
figure 7.11). The teeth thus formed washboard-like grinding surfaces that milled the vegetation cropped by the horny beak at the front of the mouth. The hadrosaurid jaw mechanism could process resistant, fibrous vegetation and even twigs. Indeed, the two mummified hadrosaurids discovered in Alberta, Canada, have stomachs full of conifer needles and twigs, seeds, and other tough plant debris, and hadrosaur coprolites are packed with conifer debris.
FIGURE 7.10
Edmontosaurus, a hadrosaurine (above), and Parasaurolophus, a lambeosaurine (below), are representative hadrosaurids. (© Scott Hartman)
FIGURE 7.11
The complex dental batteries of hadrosaurids included three or more replacement teeth at each tooth position. (Drawing by Network Graphics)
The skeletons of hadrosaurids were very similar to those of primitive iguanodontians but lacked thumbs, differed in the shape of the pelvis, and had as many as 8 to 10 sacral vertebrae. Few differences are evident among the skeletons of different types of hadrosaurids, so identifying them is done almost exclusively by comparing their skulls.
FIGURE 7.12
The flat skulls of hadrosaurines (far left) contrast with the crested skulls of lambeosaurines (center and right), among which there was great variety. (© Mark Hallett. Reproduced with permission of Mark Hallett Paleoart)
Although the variation in skull type among hadrosaurids is astounding, just two hadrosaurid subfamilies are generally recognized:
Hadrosaurinae and
Lambeosaurinae (
figure 7.12). Hadrosaurines are the more primitive subfamily and had rather flat skull roofs. They are sometimes called the “Roman-nosed” hadrosaurs after their large and long nasals that often peaked near the posterior end of the nostrils.
Edmontosaurus, from the Upper Cretaceous of North America, is a characteristic hadrosaurine (see
figure 7.10).
The lambeosaurines are the advanced subfamily, often referred to as the “crested” hadrosaurs. They are distinguished by the convoluted
tubes and crests on the tops of their skulls, which contained the modified nasal passages and a nasal cavity relocated above the orbits. Other distinctive lambeosaurine features include elongated neural spines on the vertebrae and relatively robust limbs. The varied crests and tubes on lambeosaurine heads likely served as visual display and probably also acted as resonators to produce distinctive calls (
box 7.3). Many of the differences in crest and tube size and shape reflect growth and sexual dimorphism (
figure 7.13).
Parasaurolophus, from the Upper Cretaceous of North America, is a representative lambeosaurine (see
figure 7.10).
Most of the features of the hadrosaurid skeleton suggest that these dinosaurs were powerful quadrupedal walkers also well suited to bipedal locomotion. The hadrosaur mummies from Alberta, however, have flaps of skin between their fingers that suggest a webbed hand. And, the tall, paddle-like tail of hadrosaurids looks suited (though its flexibility was limited) to propelling these dinosaurs in water. These features, and the fact that many hadrosaurid fossils, including complete, articulated skeletons, are found in rocks deposited in rivers, lakes, swamps, and even the sea, have led some paleontologists to view hadrosaurids as aquatic. It seems most certain, however, that hadrosaurids were very well suited to living on dry land and may have entered water only for defense or feeding. Much more has been suggested about hadrosaurid behavior, especially based on their nests of eggs, and is discussed in
chapter 13.
The crests and tubes on the heads of lambeosaurines have elicited speculation about their function for decades (
box figure 7.3). An early idea was that the crests and tubes allowed for feeding underwater either by acting as snorkels (but no upward openings were present in the crests and tubes), as air-storage tanks (but the volume of air that could be stored in the crests was very small relative to lung volume), or as “air locks” to prevent water from entering the lungs (although how this “air lock” would have worked is unclear). Other, more plausible suggestions are that the hollow spaces inside the tubes and crests provided space for glands or, by increasing the surface area for tissue inside the nasal cavity, warmed and moistened inhaled air and improved the sense of smell. Another suggestion is that the tubes and crests were used to deflect foliage when lambeosaurines crashed through dense forests.
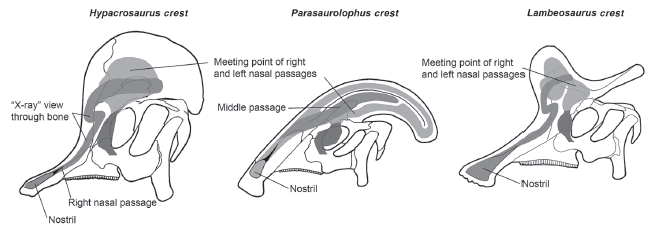
BOX FIGURE 7.3
The tubes and crests on lambeosaurine skulls were hollow and connected to the nasal passages. (© Scott Hartman)
These plausible functions of the crests and tubes do not exclude what may have been their most important function—as signaling devices. This signaling would have been both visual and auditory. The fact that particular shapes of crests and tubes were specific to particular types of lambeosaurines, and that the crests of males and females of a specific type of lambeosaurine were of different sizes and shapes, strongly supports their identification as visual signaling devices. Distinctive-looking tubes and crests thus allowed lambeosaurines to recognize members of their own species and to distinguish males from females.
Another kind of display was also possible with the lambeosaurine crests and tubes because they were hollow and connected to the air passages. Such hollow structures could have acted as resonating chambers, with differently shaped tubes and crests producing distinctive sounds like the differently shaped wind instruments of a symphony orchestra. So, not only could lambeosaurines identify each other visually by their crests and tubes, they could signal to others with distinctive sounds. The Late Cretaceous landscapes they inhabited must have been noisy places!

FIGURE 7.13
The tubes and crests on the skulls of lambeosaurines changed during growth and were sexually dimorphic. (© Scott Hartman)
Hadrosaurids first appeared in Asia during the Late Cretaceous. Later in the Late Cretaceous, they had spread to North America, Asia, Europe, South America, and Antarctica, though they were most diverse and abundant in Asia and North America. Hadrosaurids lived in a variety of habitats, from inland floodplains to coastal swamps and jungles. In the New World, their distribution extended from Alaska to Argentina. Hadrosaurids were among the dominant plant-eating dinosaurs of the Late Cretaceous and were among the last dinosaurs to become extinct.
As mentioned at the beginning of this chapter, the ornithopods were one of the most long-lived and diverse groups of dinosaurs. The dental battery was a hallmark of the ornithopods and one of the keys to their great success. They first appeared during the Middle Jurassic as small bipeds and were widespread by the Late Jurassic.
Indeed, during the Late Jurassic, it would have been virtually impossible to visit any place on Earth without encountering an ornithopod dinosaur. Small to medium-size primitive ornithopods and large iguanodontians were very diverse and abundant plant-eating dinosaurs of the Late Jurassic through to the Early Cretaceous and had an essentially worldwide distribution.
The hadrosaurids arose by Late Cretaceous time. Their first appearance coincided with a marked decline in the types and numbers of other ornithopods. It is thus tempting to believe that hadrosaurids contributed to the demise of their more primitive relatives. Hadrosaurids were extremely diverse and abundant during the Late Cretaceous and inhabited a variety of environments until their own demise 66 million years ago.
1. Ornithopods were bipedal ornithischian dinosaurs that lived from the Middle Jurassic to the Late Cretaceous. Their fossils are found on all continents.
2. Heterodontosaurs were primitive ornithischians considered by some paleontologists to be closely related to ornithopods. They were small bipedal dinosaurs with unique, chisel-like teeth best known from the Lower Jurassic of southern Africa.
3. Hypsilophodon and its relatives were small bipedal ornithopods that were especially diverse and widespread during the Late Jurassic and Early Cretaceous.
4. The success of Hypsilophodon and its relatives may have been based in part on their ability, via hinges in the skull, to move their dental batteries sideways in order to grind vegetation.
5. Primitive iguanodontians are well represented by Tenontosaurus, Camptosaurus, and the famous Iguanodon.
6. The primitive iguanodontians were large, facultatively bipedal ornithopods that were particularly successful during the Late Jurassic and Early Cretaceous.
7. Hadrosaurids, the duck-billed dinosaurs, first appeared during the Late Cretaceous and were particularly diverse and abundant in Asia and North America.
8. The appearance of hadrosaurids nearly coincides with the decline of the other ornithopods.
9. Hadrosaurids encompass two subfamilies: the hadrosaurines with flat skulls and the lambeosaurines with skulls bearing dorsal tubes or crests.
10. The hollow tubes and crests on lambeosaurine skulls were most likely signaling devices used for both visual display and as resonating chambers to produce distinctive sounds.
11. Hadrosaurids were powerful quadrupedal walkers that may also have been amphibious.
Camptosaurus
dental battery
Dryosaurus
Edmontosaurus
Hadrosauridae
Hadrosaurinae
Heterodontosaurus
Hypsilophodon
Iguanodon
Iguanodontia
Lambeosaurinae
ornithopod
Ouranosaurus
Parasaurolophus
resonating chamber
Tianyulong
tubes and crests
Uteodon
1. What are the distinctive features of ornithopods?
2. To what feature(s) might you attribute the success of ornithopods?
3. How did primitive ornithopods differ from heterodontosaurs, and how might these differences explain the greater success of the primitive ornithopods?
4. How did hadrosaurids resemble and differ from primitive ornithopods?
5. What features of the skeletons of different groups of ornithopods identify them as either bipeds or facultative bipeds?
6. What defensive strategies did ornithopods employ?
7. How do the jaw, tooth, and skull structures of the various ornithopods differ from each other? Is a progression in the evolution of chewing mechanisms evident among the ornithopods?
8. Why do paleontologists think the crests and tubes of lambeosaurines functioned as signaling devices?
Butler, R. J., and P. M. Barrett. 2012. Ornithopods, pp. 551–566, in M. K. Brett-Surman, T. R. Holtz, Jr., and J. O. Farlow, eds., The Complete Dinosaur. 2nd ed. Bloomington: Indiana University Press. (Concise review of the ornithopods)
Case, J. A., et al. 2000. The first duck-billed dinosaur (family Hadrosauridae) from Antarctica. Journal of Vertebrate Paleontology 20:612–614. (Describes an Upper Cretaceous hadrosaur tooth found in Antarctica)
Eberth, D. A., and D. C. Evans, eds. 2014. Hadrosaurs. Bloomington: Indiana University Press. (A collection of 36 technical articles presenting recent research on hadrosaurs)
Godefroit, P., ed. 2012. Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems. Bloomington: Indiana University Press. (A collection of 33 technical articles on Bernissart and much more about Early Cretaceous dinosaurs and ecosystems)
Prieto-Márquez, A. 2010. Global phylogeny of Hadrosauridae (Dinosauria; Ornithopoda). Zoological Journal of the Linnean Society of London 159:435–502. (Recent cladistic analysis of hadrosaur phylogeny)
Weishampel, D. B. 1981. Acoustical analysis of potential vocalization in lambeosaurine dinosaurs (Reptilia: Ornithischia). Paleobiology 7:252–261. (A technical analysis of the acoustical properties of lambeosaur cranial crests)
Weishampel, D. B. 1984. Evolution of jaw mechanisms in ornithopod dinosaurs. Advances in Anatomy, Embryology and Cell Biology 87:1–109. (A detailed examination of the skulls and teeth of ornithopods)
Zheng, X., H. You, X. Xu, and Z. Dong. 2009. An Early Cretaceous heterodontosaurid dinosaur with filamentous integumentary structures. Nature 458:333–336. (Describes Tianyulong)
Ornithopod dinosaurs can be found at many of the world’s great natural history museums. In Brussels, Belgium, the Royal Belgian Institute of Natural Science displays 30 of the Iguanodon skeletons found during the nineteenth century at Bernissart, Belgium, by miners. Do take note of the large thumb spikes on those Iguanodon skeletons, which were originally thought to be nose horns. In North America, hadrosaurs (duck-billed dinosaurs) are mainstays of the dinosaur exhibits at the American Museum of Natural History (New York) and the Museum of the Rockies (Bozeman, Montana). The Bozeman display features the well-known Upper Cretaceous hadrosaur Maiasaura and its nests of eggs and baby hadrosaurs found in Montana. In Albuquerque, New Mexico, the bizarre tube-headed hadrosaur Parasaurolophus is featured as both skull and sculpture at the New Mexico Museum of Natural History and Science. Most notable, however, are the many superb hadrosaur skeletons and skulls on display in Canada at the Royal Ontario Museum (Toronto, Ontario) and the Royal Tyrrell Museum of Palaeontology (Drumheller, Alberta). And, if you are in Thailand, the Sirindhorn Museum and Phu Kum Khao Dinosaur Excavation Site near Sahatsakhan displays replicas of ornithopod skeletons and many other dinosaurs.