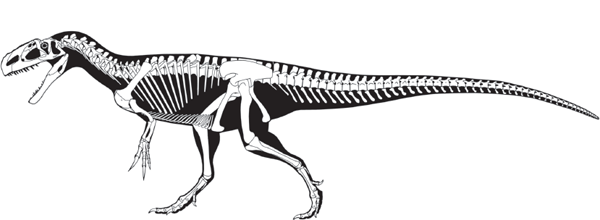
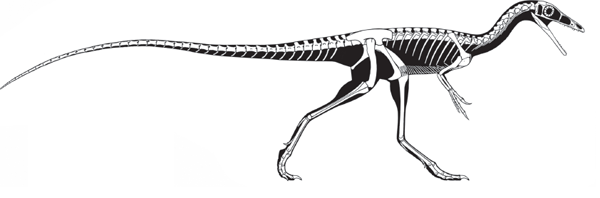
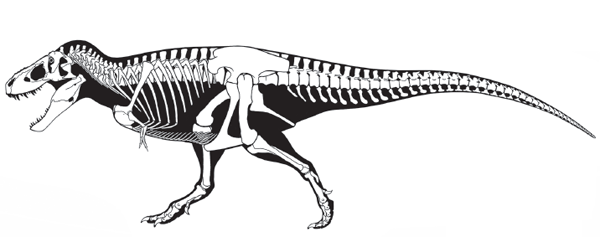
MOST predatory (meat-eating) dinosaurs belong to one group, Theropoda (beast foot) (
box 5.1). The theropod dinosaurs spanned the entire duration of dinosaurs, from the Late Triassic until the end of the Cretaceous. They include animals that ranged in size from the chicken-size
Compsognathus to the most famous land-living predator of all time,
Tyrannosaurus rex. Throughout most of their long history, theropods were a diverse and successful group of dinosaurs.
In this chapter, I examine the main types of theropods and discuss their evolution. But, before doing this, we need to consider some of the complexity associated with the fossil record of theropods and how it affects our view of the evolution of these dinosaurs.
Theropods are the most diverse group of dinosaurs, and recent discoveries have dramatically changed how we view their composition. Clearly, these dinosaurs had a very complex evolution from the Late Triassic through the Late Cretaceous. Indeed, no area of dinosaur phylogeny is more fluid or more debated. At least half a dozen phylogenetic analyses of all the theropods, or of major segments of the Theropoda, can be considered current. The phylogeny used here embodies much of what can be considered consensus (
figure 5.1).
The oldest widely accepted scheme of theropod phylogeny recognized two clades. According to this scheme, the very large theropods with short necks and small forelimbs, the
carnosaurs (meat lizards), diverged early from the smaller, long-necked theropods with large forelimbs, the coelurosaurs (hollow-tailed lizards). This view of theropod phylogeny, however, has been superseded by much more complex phylogenies needed to accommodate the new array of theropods discovered during the past two decades (see
figure 5.1). Thus, primitive theropods, including the ceratosaurs (horned lizards), represent multiple branches in the evolution of theropods. The clade that led to the carnosaurs and coelurosaurs is referred to as the tetanuran theropods. The ancestry of birds lies among the tetanurans and is discussed in
chapter 15.
The Latinized names that paleontologists give to extinct animals usually make some sort of sense. They typically either describe some feature or attribute of the animal, make reference to the place where its fossils were found, or honor another paleontologist (see
box 2.1). Some names are very colorful, like
Tyrannosaurus rex, the “king of the tyrant lizards.” Other names are rather pedestrian, like
Allosaurus, the “different lizard.”
The name applied to all meat-eating dinosaurs is Theropoda, from the Greek therios, meaning “beast,” and podos, meaning “foot.” Although therios literally means “beast,” it usually refers to mammals, so the name Theropoda also means “mammal foot.” Yale paleontologist Othniel Charles Marsh coined the name Theropoda for the meat-eating dinosaurs in 1881. Ten years earlier, Marsh had coined the name Ornithopoda (bird foot), and in 1878 he introduced the name Sauropoda (lizard foot). Indeed, 1882 was a milestone in the history of dinosaur studies, for in that year Marsh published a comprehensive classification of dinosaurs. He divided them into theropods, ornithopods, sauropods, and stegosaurs (plated lizards). Marsh also considered the Hallopoda (leaping foot) to be possible dinosaurs, but they are now thought to be crocodiles.
Clearly, Marsh relied heavily on foot structure as a guide to classifying dinosaurs. In the feet of the dinosaurs he called sauropods, Marsh saw resemblances to the feet of living lizards, hence the name “lizard foot.”
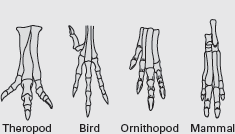
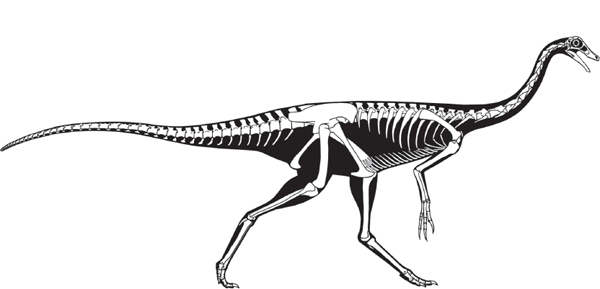
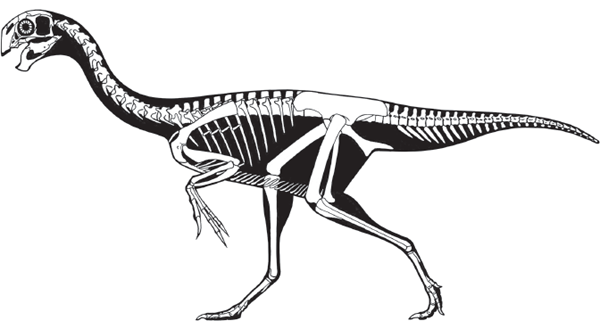
But, what similarities in the feet of meat-eating dinosaurs and mammals did Marsh see? And, given the very bird-like feet of these dinosaurs, why didn’t Marsh name them Ornithopoda? Furthermore, why did Marsh assign the name Ornithopoda to dinosaurs with feet that were more mammal-like than bird-like (
box figure 5.1)?
BOX FIGURE 5.1
Theropod feet were bird-like, whereas ornithopod feet were mammal-like. (Drawing by Network Graphics)
No clear answer to these questions can be found in Marsh’s writings, which do, however, make it clear that he knew the meanings of the words “theropod” and “ornithopod.” The best guess is that Marsh erred when he introduced the names “ornithopod,” and later, “theropod,” applying them in reverse order from what had been his original intention. The result is that the bird-footed dinosaurs are not called ornithopods, but, rather, nonsensically, theropods. Conversely, the name “ornithopod” is applied to dinosaurs with rather mammal-like feet.
Marsh was a very famous paleontologist whose knowledge of dinosaurs was immense. So, his names for two major groups of dinosaurs, Theropoda and Ornithopoda, were accepted and used by paleontologists worldwide. They continue to be used to this day, despite the fact that neither name quite makes sense.
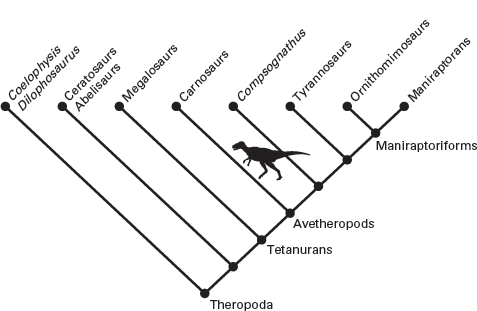
FIGURE 5.1
The phylogeny of theropods currently accepted by most paleontologists shows a complex evolutionary history of this diverse group of dinosaurs. (Drawing by Network Graphics)
Although the picture of theropod phylogeny presented here (see
figure 5.1) seems neat and clear, ongoing debate about the phylogeny of theropods makes it one of the least-agreed-upon phylogenies among the dinosaurs. In part, this is because different paleontologists place greater emphasis on different features as significant evolutionary novelties in theropod evolution. These disagreements cannot be readily resolved at present. So, suffice it to say that several alternative views of theropod phylogeny are plausible, and the one presented here represents a consensus of most paleontologists.
Because theropods comprise most of the meat-eating dinosaurs, it might seem sufficient to identify theropods by the skeletal features, especially the teeth, that were involved in acquiring and processing animal food. It is important to remember, however, that many of the most primitive dinosaurs (such as
Staurikosaurus and
Herrerasaurus) were also meat-eaters (see
chapter 4). Indeed, meat-eating is the primitive condition for all non-dinosaurian archosaurs. Thus, to identify theropods specifically, we must distinguish them from the primitive meat-eating dinosaurs and from other meat-eating archosaurs.
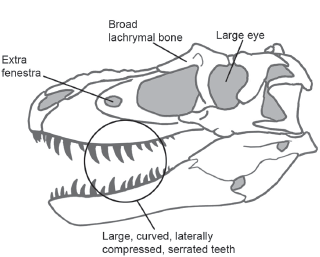
Most of the novelties that distinguish theropods reflect more specialized structures for meat-eating than were present in the primitive, meat-eating dinosaurs. They also reflect the fact that theropods were highly specialized runners and predators. These novelties include broad exposure of the lachrymal bone on the skull roof; a joint (hinge) within the jaw; a sacrum comprising at least five vertebrae; clavicles fused into a wishbone (furcula); four or fewer digits on the hand; and a hind foot that was compact, narrow, and long with three functional digits and a first digit separated from the rest of the foot (
figures 5.2 and
5.3). These novelties and others distinguish theropods from other dinosaurs. Combining these novelties with the primitive features inherited from their nontheropod ancestors gives us an overall picture of theropods as remarkable predators.
FIGURE 5.2
This tyrannosaurid skull shows typical features of theropods. (© Scott Hartman)
The teeth of many theropods were relatively large, curved, compressed, and serrated. The mouth bore numerous such teeth that were held in the jaw by a ligament (see
figure 5.2). The relatively large eyes suggest that theropods located their prey visually. Theropods had the largest and most sophisticated brains of any known dinosaurs (see
chapter 14). Indeed, the brains of the more advanced theropods were very bird-like, suggesting that at least some theropods may have been very sophisticated behaviorally.
The slicing teeth of theropods were set in a lightly built skull in which many of the bones were only loosely joined to each other. Even the bones of the theropod lower jaw were not tightly sutured to each other (see
figure 5.2). These loose joints probably helped the skull compensate for the shocks it encountered when the theropod grabbed and chewed its prey, whereas the rigid lower jaw helped grab and crush the prey.
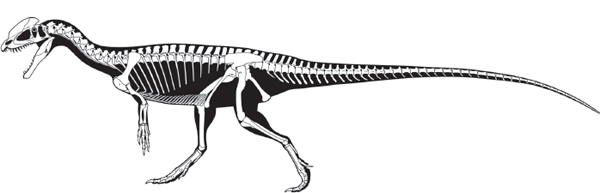
FIGURE 5.3
Dilophosaurus, from the Early Jurassic of Arizona, was a typical primitive theropod. (© Scott Hartman)
The theropod skull was attached to the neck at a highly mobile joint. This joint allowed rapid and precise movement of the head, and its presence further supports the idea that theropods were highly sophisticated visual hunters. Today, birds have the same type of head–neck joint, as did their theropod ancestors.
We also see many distinctive features designed for predation in the postcranial skeletons of theropods. Thus, the hands of most theropods had only three digits, one of which (the first digit) was offset from the other two (see
figure 5.3). This digit, like our thumb, may have allowed the theropod hand to grasp. On all three digits of the theropod hand, the next-to-last phalanges were elongated, to extend the reach, and the tips of the digits were long, laterally compressed, and bore curved claws.
Theropod hands clearly were not used for walking. So, theropods were obligate bipeds, and their vertebral columns and hind limbs were modified accordingly. At least five vertebrae made up the theropod sacrum to provide greater rigidity at the attachment point of the powerful hind limbs and backbone. The pelvis was very large and provided considerable space for the attachment of the large hip muscles. The femur was distinctly bowed downward, and the hind foot was elongate and symmetrical around the middle digit. Indeed, the theropod hind foot was the very compact foot of a fast runner and was strikingly bird-like (see
box 5.1).
In many theropods, the tail changed about midway from being flexible proximally to being stiff distally. A similar type of tail is still present in living birds. The rigid distal portion could have been used by theropods as a stabilizer while running and as a counterweight to the body.
A final typical theropod feature was hollow bones. This lightened the theropod skeleton and also increased the strength of the bones to resist bending. Hollow bones evolved in theropods as an aid to rapid, terrestrial locomotion, but later aided their descendants, the birds, in flight.
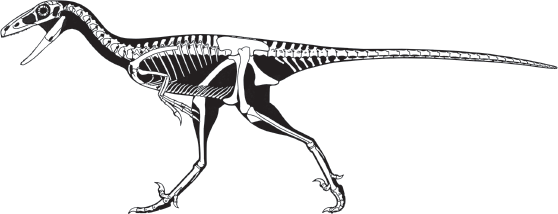
Theropods were already important terrestrial predators in the Late Triassic and became the dominant predators of the land during the Jurassic and Cretaceous. Theropods can be divided into three groups: primitive taxa, such as the coelophysoids and dilophosaurids; the ceratosaurs; and the tetanurans. The primitive theropods were relatively small bipedal hunters, ranging from the 1-meter-long
Procompsognathus to the 3-meter-long
Coelophysis (
box 5.2).
One of the best known primitive theropods is
Dilophosaurus, from the Lower Jurassic of Arizona (see
figure 5.3). This large (6-meter-long) dinosaur displayed many typical theropod features, such as a large head full of long, blade-like teeth; a large antorbital fenestra; and a thick, powerful lower jaw. The neck and tail were long, and the forelimb was much shorter than the hind limb.
Dilophosaurus had many fused bones in the hind limb. It also had a distinct gap between the premaxillary and maxillary bones in the skull, and the hand had four fingers, the first three of which bore claws. The two thin, bony crests on the skull of
Dilophosaurus are the basis of its name, which means “two-crested lizard.”
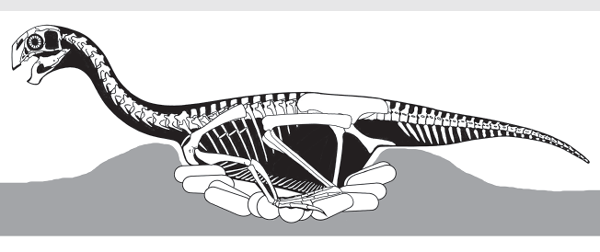
Very few theropod dinosaurs are known from mass death assemblages that represent a sample of a theropod population. Late Triassic Coelophysis is an exception. In 1947, an expedition of the American Museum of Natural History discovered a Coelophysis bonebed in the Upper Triassic rocks of northern New Mexico. At least a few hundred complete skeletons of Coelophysis have been excavated from the bonebed, which is still not completely mined out. Taphonomic analysis indicates that a population of the small theropod was washed into a pond, where the dinosaurs likely drowned, and then were rapidly buried by a second flood event. This population can thus be seen as a large sample of a flock of Coelophysis.
Careful analysis of the Coelophysis skeletons provides rare insight into aspects of the biology of an early theropod dinosaur:
1.
Coelophysis was a visually oriented diurnal hunter that was an obligate biped and a fast, agile runner (
box figure 5.2).
2. The fact that so many dinosaurs were together at death suggests gregarious behavior; they likely aggregated to reproduce.
3. Two kinds of Coelophysis have been found in the bonebed: robust dinosaurs with short skulls, short necks, and large forelimbs, and gracile dinosaurs with long skulls, long necks, and smaller forelimbs. These are likely sexual dimorphs, and the gracile dinosaur is probably the female.
4. Cannibalism of the young is evidenced by bones and coprolites in some of the adult body cavities.
5. The distribution of size classes suggests rapid growth with sexual maturity at about two to three years and that the largest dinosaurs were about seven years old at death.
6. Based on the size classes, there are about 20 yearlings per large adult, so juvenile mortality must have been high.
The Coelophysis bonebed thus gives paleontologists remarkable insight into the lifestyle and population biology of an early theropod dinosaur. Not surprisingly, Coelophysis seems to have been very bird-like in its biology.
BOX FIGURE 5.2
The skull of Coelophysis, with its large eyes, is that of a diurnal, visual hunter.
At the beginning of the Jurassic, Dilophosaurus was the first relatively large dinosaurian predator. Its presence marked the ascent of the theropods to the status of apex predators in Jurassic–Cretaceous terrestrial communities.
Ceratosaurs were a significant group of primitive theropods of the Late Jurassic through to the Late Cretaceous. More than 40 species can be recognized from fossils found in North America, Europe, India, South America, Africa, and Madagascar. Key evolutionary novelties of the Ceratosauria include having six or more sacral vertebrae; very deep coracoids; small forelimbs; and very weak, perhaps even nonfunctional, hands. Ceratosaurs encompass quite a range of theropods, including Limusaurus from the Upper Jurassic of China, with its toothless skull, which was probably the first noncarnivorous theropod.
The 6- to 8-meter-long
Ceratosaurus, from the Upper Jurassic of North America and Europe, is a characteristic, relatively large ceratosaur. However, most of the diversity of ceratosaurs was found in abelisauroids, a Cretaceous group known largely from the Gondwana continents. Abelisauroids are characterized by an enlarged external mandibular fenestra, arched sacral vertebrae, a round head on their humerus, and distinctively doubled grooves on the distal phalanges of their hind feet, among other features.
Aucasaurus, from the Late Cretaceous of Argentina, well represents the abelisaurs (
figure 5.4). Note its extremely small forearms, a characteristic seen in many abelisaurs, thought to have been useless vestigial structures.
FIGURE 5.4
Aucasaurus, from the Upper Cretaceous of Argentina, with its tiny forelimbs, is a characteristic abelisaur. (© Scott Hartman)
Some ceratosaur skeletons show marked sexual dimorphism (differences between presumed males and females). Thus, there seems to have been a “male” form with a longer skull and neck, thicker and more robust limbs, and very large muscle-attachment sites. The “female” form, in contrast, had a juvenile-like skeleton with a shorter skull and neck and slender limbs. Further, there were crests on the skulls of some ceratosaurs. These were made of very thin bone and presumably functioned in display, probably by males.
The strange name
Tetanurae (fused tails) is applied by paleontologists to most of the theropods (see
figure 5.1). These theropods were much more bird-like than the primitive theropods. Indeed, most paleontologists have concluded that the ancestry of birds lies among the tetanuran theropods. Evolutionary novelties of the tetanurans include relatively large heads, a reduction in the dentition so that all teeth were in front of the orbit, and interlocking vertebrae that stiffened the tail.
Three great groups of tetanuran theropods can be recognized: the megalosaurs, the carnosaurs, and the coelurosaurs. The carnosaurs and coelurosaurs are united in a group called the Avetheropoda, because it is from this group that birds originated.
Megalosaurs were large theropods of the Middle Jurassic through to the Late Cretaceous. They were characterized by long skulls with elongated snouts that had small or no crests on the skull roof. This contrasts with the skull of carnosaurs, such as Allosaurus, in which the skull was tall with a short and blunt snout and had horns or crests on the skull roof. The powerful forelimb of megalosaurs also was a distinctive feature.
Megalosaurus, from the Lower and Middle Jurassic of Great Britain and France, is the archetypal megalosaur. It is known from skull fragments, lower jaws, teeth, and various postcranial bones. Described by the British naturalist William Buckland in 1824,
Megalosaurus was the first dinosaur to receive a scientific name. It is estimated to have been about 9 meters long and resembled
Allosaurus in many features. A better known and characteristic Late Jurassic megalosaur is
Monolophosaurus from China (
figure 5.5).
Spinosaurids were megalosaurs of the Cretaceous with very elongate, crocodile-like snouts, conical teeth (lacking serrations,) and tall dorsal neural spines that formed a sailback.
Spinosaurus, from the Lower Cretaceous of Egypt, is characteristic and, at 14 meters long, now holds the record as the largest theropod (
figure 5.6). Known from nearly complete skeletal material,
Spinosaurus had 1.8-meter-long neural spines on its back that formed a “sail” reminiscent of that of
Dimetrodon, a Permian reptile often mistakenly thought to be a dinosaur. Gut contents and the conical teeth indicate it was a fish-eater that swam to hunt. Other spinosaurids include
Baryonyx, from the Lower Cretaceous of Europe;
Suchomimus, from the Lower Cretaceous of North Africa; and
Irritator, from the Lower Cretaceous of Brazil.
FIGURE 5.5
Monolophosaurus, from the Upper Jurassic of China, is a representative megalosaur. (© Scott Hartman)
FIGURE 5.6
Spinosaurus was the largest theropod—a huge, sail-backed fish-eater. (© James McKinnon)
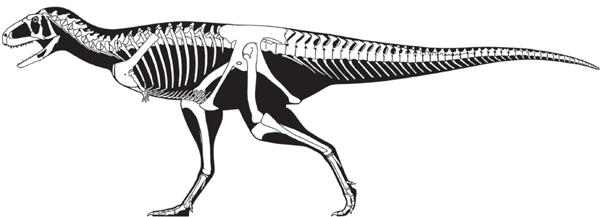
FIGURE 5.7
Late Jurassic Allosaurus (above) and Late Cretaceous Giganotosauros (below) were typical carnosaurs. (© Scott Hartman)
More advanced than megalosaurs, the avetheropods had an additional fenestra in their maxilla, complicated air sac chambers in their vertebrae, and a reduced fourth digit in the manus (“hand”). The dominant large theropods of the Middle Jurassic to Early Cretaceous were the carnosaurs, a group of primitive avetheropods. Carnosaurs are readily distinguished from other avetheropods by having an extra opening in the maxillary and very large nasal bones, among other features.
Allosaurus is the best known carnosaur (
figure 5.7). A unique feature of
Allosaurus was its lightly constructed skull, which had a distinctive, roughened ridge just above and in front of the orbit.
Allosaurus is best known from the remains of at least 44 individuals collected at the Cleveland-Lloyd Dinosaur Quarry in eastern Utah (
figure 5.8).
Cretaceous carnosaurs are carcharodontosaurs. They include
Giganotosaurus (see
figure 5.7), from the Upper Cretaceous of Argentina, a carnosaur almost as large as
Spinosaurus.
FIGURE 5.8
This map shows some of the scattered bones at the Cleveland-Lloyd Dinosaur Quarry in Utah, where the remains of at least 44 individuals of Allosaurus were collected. (From J. H. Madsen, Jr. 1976. Allosaurus fragilis: A revised osteology. Utah Geological and Mineralogical Survey Bulletin 109:1–163. Copyright © 1976 James H. Madsen, Jr. Reprinted by permission)
The most bird-like tetanurans were the coelurosaurs, and they include the ancestors of birds. Their key evolutionary novelties include enlarged brains (at least twice the size of other theropods of comparable body size), a tridactyl hand with long second and third digits, long and narrow metatarsals, and boat-shaped chevron bones. Primitive coleurosaurs had what have been called “protofeathers,” so some sort of feathery body covering may also diagnose the group.
Coelurosaurs were mainly a Cretaceous group, best known from Asia and western North America. Several different phylogenies of coelurosaurs have been proposed, so there is a variety of classifications of the many theropods assigned to Coelurosauria. Here, we divide the coelurosaurs into three groups: the primitive coelurosaurs, represented by Compsognathus; the tyrannosauroids; and the Maniraptoriformes.
The Genus Compsognathus
Compsognathus (delicate jaw) is a small dinosaur best known from two virtually complete skeletons from the Upper Jurassic of western Europe (
figure 5.9). The better of the two specimens is exquisitely preserved on lithographic limestone collected near Solnhofen, Bavaria, in southern Germany. In fact, this skeleton of
Compsognathus was one of the first complete dinosaur skeletons ever described.
This skeleton represents a small theropod only 70 centimeters long. Its small size, large eyes, and relatively very large head suggest it belonged to an immature theropod. Indeed, the second skeleton of Compsognathus, from France, is about 50 percent larger than the German specimen.
Compsognathus had the typical proportions of a small tetanuran. It was lightly built and had a long neck, long tail, and short forelimbs that were only 37 percent of the length of the large hind limbs. It also displayed typical tetanuran evolutionary novelties, such as all teeth in the skull and lower jaws being in front of the orbit and a hand lacking a fourth finger.
FIGURE 5.9
The skeleton of Compsognathus, from Germany, well represents a primitive coelurosaur. (© Scott Hartman)
Perhaps one of the most interesting features of
Compsognathus is that the German specimen preserves its last meal. Inside the abdomen of this skeleton is most of the skeleton of an extinct lizard,
Bavarisaurus. This small skeleton was originally identified as an embryo of
Compsognathus and was presented as evidence that the small theropod gave live birth. Closer examination of the skeleton in the
Compsognathus abdomen, however, now convinces paleontologists that just before this dinosaur died, it ran down and ate a small lizard.
Tyrannosauroids
Tyrannosauroids are known primarily from fossils collected in Upper Cretaceous deposits of Asia and western North America, but their fossil record does extend back to the Middle Jurassic. The best known genera are Albertosaurus, Daspletosaurus, and Tyrannosaurus from North America and Tarbosaurus from Asia. Evolutionary novelties of tyrannosauroids include fused nasal bones, incisor-like premaxillary teeth, and a very slender third metacarpal.
The archetypal
tyrannosaurid is
Tyrannosaurus, one of the largest (13 meters long, weighing an estimated 6 tons) land-living meat-eaters of all time (
figure 5.10). This most famous theropod had a 1.3-meter-long head that bore serrated, blade-like teeth, each as much as 20 centimeters long. Its massive head was supported by a short, thick neck, and its short, thick abdomen projected forward from a massive pelvis supported by huge, pillar-like limbs. The long, heavy tail acted to counterbalance the body when running. The tiny forelimbs make a curious impression; their function is not clear (
box 5.3). Other Late Cretaceous tyrannosauroids were in many ways just slightly abbreviated, more primitive versions of
Tyrannosaurus.
During the Late Cretaceous in Asia and western North America, tyrannosauroids were moderately diverse and successful predators. They were the last large-bodied theropods to evolve and include some of the last dinosaurs.
FIGURE 5.10
Late Cretaceous Tyrannosaurus was a typical tyrannosaurid. (© Scott Hartman)
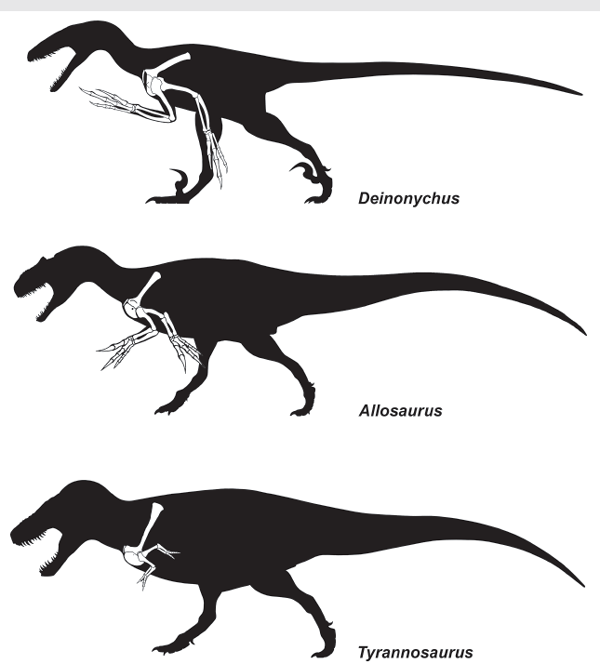
The tiny forelimbs of
Tyrannosaurus rex have long puzzled paleontologists (
box figure 5.3). What did the dinosaur do with those puny arms? Most answers to that question have focused on what the forelimbs could not do—they could not reach the dinosaur’s mouth, and they could not have been used to push the dinosaur up if it laid down. Indeed, this has led many paleontologists to conclude that the forelimbs of
Tyrannosaurus served no useful function. They were simply vestigial structures on their way to being completely lost. Indeed, the forelimbs of some abelisaurs are even proportionately smaller than those of
Tyrannosaurus. They, too, are seen as functionally useless vestiges.
BOX FIGURE 5.3
Tiny Tyrannosaurus forelimbs are typically considered useless by paleontologists. (© Scott Hartman)
There have been, however, a couple of positive suggestions about possible forelimb function by
Tyrannosaurus rex. One is that the tiny arms helped to balance the extremely large head and prevent the dinosaur from tipping over. And, a more compelling analysis demonstrates that the tiny arms of
Tyrannosaurus were actually strong and powerful—certainly more so than human arms. Thus, reconstructions of the forelimb musculature and strength calculations indicate that they could be moved quickly and resist strong forces. This suggests that they may have been used to clutch prey. Perhaps these forelimbs were vestigial but still were useful to what was one of the great dinosaurian predators.
Maniraptoriforms
The derived coleurosaurs are a mainly Cretaceous group formally called the Maniraptoriformes. They were distinguished from other coelurosaurs by many features, including even larger brains, small skulls relative to body size, and an increased number of relatively smaller teeth. Many of the maniraptoriforms were not carnivorous and instead were either omnivores or herbivores. Here, we divide the maniraptoriforms into the ornithomimosaurs and the maniraptorans.
Ornithomimosaurs Well-known and truly abundant primitive maniraptoriforms are the “bird-mimic lizards,” the ornithomimosaurids. Best known from the Upper Cretaceous of Asia and western North America, the
ornithomimosaurs strongly resemble modern flightless birds, such as emus and ostriches, in having small heads, long necks, and long legs (
figure 5.11). But their long arms with clawed hands and their long, bony tails are the two most obvious features that distinguish them from birds.
FIGURE 5.11
Ornithomimosaurids were very similar in overall body shape and proportions to living ostriches. (Drawing by Network Graphics)
Evolutionary novelties that distinguish ornithomimosaurs from other maniraptoriforms include the lightly built skull with very large orbits and shallow snout, the lack of (or near lack of) teeth (presumably a horny bill covered much of the jaws), and the long neck, which accounts for about 40 percent of the length of the vertebral column in front of the sacrum.
Struthiomimus, from the Upper Cretaceous of western Canada, was a typical ornithomimosaur and possessed these evolutionary novelties (
figure 5.12). Other features of
Struthiomimus include the very flexible joints in this dinosaur’s long neck, which conferred great mobility. The ventral part of the abdomen was protected by belly ribs (gastralia), and deep scars on the spines of the dorsal vertebrae suggest the presence of powerful ligaments along the back. Such ligaments and the gastralia would have produced a very rigid abdomen that was held forward and nearly horizontal as the dinosaur ran. Large attachment sites for muscles on the distal caudal vertebrae would have helped to hold the tail rigid as a counterbalance.
The forelimbs of Struthiomimus were extremely long and slender, about 50 percent the length of the hind limbs. The long, slender hands had three functional fingers that bore long, pointed, but only slightly curved claws. The long, slender hind limbs had three functional toes as well, but they bore flattened claws.
The anatomy of Struthiomimus and some other ornithomimosaurs strongly suggests that they were fast ground runners, perhaps as swift as living, similarly proportioned ostriches, which achieve top speeds of 50 kilometers per hour. Their diet, however, is less clearly interpreted from their toothless, beak-like mouths. Most paleontologists view ornithomimosaurs as omnivores that had a mixed diet of plant and animal food.
FIGURE 5.12
Late Cretaceous Struthiomimus was a typical ornithomimosaur. (© Scott Hartman)
Maniraptorans The remaining coelurosaurs are placed in the Maniraptora (hand grabbers). They range in age from Middle Jurassic to Late Cretaceous and are the single most diverse group of dinosaurs. Their even larger brains, large bony sternum, and elongate forelimbs are evolutionary novelties that distinguish them from ornithomimosaurs.
Among the best known maniraptorans are the oviraptorosaurs, from the Cretaceous of Asia and western North America. Unique features of most oviraptorosaurs include their toothless jaws (Early Cretaceous forms still had teeth); short snouts in short, boxy skulls; very large fenestrae in the mandible; very deep and strong lower jaws; and a crest of sponge-like bone on the tip of the snout (possibly for display).
Khaan, from the Upper Cretaceous of Mongolia, was a characteristic oviraptorosaur (
figure 5.13). This 2-meter-long coelurosaur had large claws on the hands and slender hind limbs.
Khaan had a skull and jaws designed to crush tough food items. At first, paleontologists thought that a close relative of
Khaan,
Oviraptor, ate the eggs of other dinosaurs; hence its name, which means “egg stealer” (
box 5.4). But, now it is believed that oviraptorosaurs primarily ate the freshwater clams that are common fossils in the lake-margin deposits where most oviraptorosaur fossils are found.
A second diverse group of maniraptorans is the deinonychosaurs, popularly known as the “raptors.” The sickle-shaped retractable claw on pedal digit II is their most distinctive evolutionary novelty. They comprise two families: the
Dromaeosauridae and the
Troodontidae. Dromaeosaurids are known from the Cretaceous of Asia, North America, Europe, Africa, and Madagascar.
Deinonychus (terrible claw), from the Lower Cretaceous of Wyoming and Montana, is one of the best known and characteristic dromaeosaurids (
figure 5.14). This rather large (3 to 3.3 meters long) dromaeosaurid displays characteristic features of this group of dinosaurs. Note especially the large skull full of serrated teeth, the backward-directed pubis, the stiffened tail, and the huge claws on the second toes of the hind feet. Aside from these specializations, much of the anatomy of
Deinonychus was typically theropod.
FIGURE 5.13
Late Cretaceous Khaan was a typical oviraptorosaur. (© Scott Hartman)
Discovered in Mongolia in 1923 by the Central Asiatic Expeditions of the American Museum of Natural History (AMNH), Oviraptor is a small maniraptoran theropod. Found near several nests of dinosaur eggs that were thought to have been laid by the ceratopsian Protoceratops, Oviraptor was perceived to be an egg-eater. Hence, its name, conferred by AMNH Chief Paleontologist Henry Fairfield Osborn, which in Latin means “egg thief” or “egg taker.”
Indeed, the original skull of Oviraptor was found just inches away from what was thought to be a nest of Protoceratops eggs. Further, Osborn saw the toothless beak of the small theropod as a crushing mechanism to break eggs. This induced Osborn to imagine the dinosaur as a likely egg-eater that plundered Protoceratops nests. He thus coined the colorful name Oviraptor but cautioned that this “may entirely mislead us as to the feeding habits and belie its character.” Indeed, Osborn went on to conclude that the dinosaur was “herbivorous or omnivorous.” One has to wonder why, then, he named it Oviraptor!
Decades later, in the 1990s, a new AMNH expedition to the Gobi Desert found more of the same kinds of dinosaur egg, but with
Oviraptor embryos inside them. And, they found an
Oviraptor skeleton sitting on top of a nest of eggs (
box figure 5.4). This dinosaur had evidently been buried in a sandstorm as it brooded more than 15 eggs in its nest. Clearly,
Oviraptor was not likely to have been eating its own eggs, so the name of the dinosaur mistakes its diet, at the very least.
BOX FIGURE 5.4
The skeleton of the theropod Khaan brooding its nest of eggs. (© Scott Hartman)
So, what did Oviraptor eat? This has been discussed at length, because the peculiar toothless beak of the dinosaur suggests it may not have been just a meat-eater. Still, a lizard skeleton in the stomach region of a close relative of Oviraptor indicates some predation. Thus, an omnivorous diet seems most likely for Oviraptor.
Perhaps most significant is the bird-like behavior inferred for Oviraptor from the discovery of the skeleton on the nest. The eggs in the nest were arranged, which suggests the dinosaur manipulated them into a pattern, as do many modern birds. And, the dinosaur sat on the nest to incubate the eggs, also a very avian behavior.
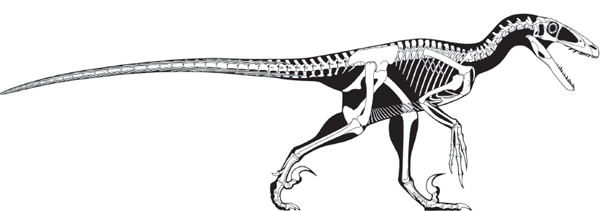
FIGURE 5.14
Early Cretaceous Deinonychus was a typical dromaeosaurid. (© Scott Hartman)
A quick examination of the skeleton of Deinonychus convincingly demonstrates it was a habitual biped. How the dinosaur might have used the huge, curved claws on its hind feet, however, is puzzling. The only way to use these claws would have been to have one or both hind feet off the ground, stabbing the prey. To do so would have required great agility and balance, aided in part by the stiff counterbalancing tail. The skeleton of Deinonychus was clearly that of a very active bipedal predator.
The second group of deinonychosaurs is the troodontids. These coelurosaurs, best known from
Saurornithoides (includes
Troodon of some paleontologists) from the Upper Cretaceous of North America and Asia, are closely related to dromaeosaurids (
figure 5.15). Characteristic features of the troodontids include a long skull with a narrow snout, a large braincase (troodontids have the largest brains relative to body size of any dinosaur), and an inflated bony casing for the middle ear. The teeth of troodontids were small and numerous (as many as 35 in the lower jaw), and many were serrated along their posterior edges. Troodontids had ankle bones fused to each other and to the tibia and small claws on the second digits of their hind feet.
Other features that distinguish Saurornithoides and other troodontids include their extremely large eyes and possible stereoscopic vision. Like dromaeosaurids, troodontids were agile bipedal predators. The earliest known troodontids are from the Lower Cretaceous of Asia, but they are best (though still poorly) known from the Upper Cretaceous of Asia and western North America.
A very unusual group of maniraptorans is the Therizinosauria (scythe reptiles), from the Cretaceous of Asia and North America. They had small skulls, long necks, huge claws, and short legs. Nearly the size of
Tyrannosaurus,
Nothronychus had 1-meter-long claws on its hands (
figure 5.16).
FIGURE 5.15
Late Cretaceous Troodon was a typical troodontid. (© Scott Hartman)
FIGURE 5.16
Nothronychus, with huge claws, was a characteristic therizinosaur. (© Scott Hartman)
Theropod dinosaurs appeared as highly specialized, running predators almost at the outset of dinosaur evolution. Early theropods of the Late Triassic were important terrestrial predators up to 3 meters long. By the Early Jurassic, 6-meter-long theropods were the apex predators of terrestrial communities. For the remainder of the Jurassic, and through the Cretaceous, theropods remained the dominant terrestrial carnivores.
The evolutionary split between ceratosaurs and tentanurans took place during the Early Jurassic. Although ceratosaurs were relatively rare during the Jurassic, they include the abelisauroids, which were the dominant predatory dinosaurs of the southern continents and Europe during the Cretaceous.
Tetanurans comprise three major groups: the megalosauroids, the carnosaurs, and the maniraptoriforms. Jurassic megalosaurs were large predators, but the Cretaceous megalosaurs, notably the spinosauroids, were a group of fish-eaters that include the largest known theropods. Carnosaurs were large theropods of the Middle Jurassic and Cretaceous and include the gigantic carcharodontosaurs of the Cretaceous of South America and Africa.
Coelurosaurs first appeared as small carnivorous forms during the Middle Jurassic. However, by the Cretaceous, there were herbivorous and omnivorous coelurosaurs as well. Small, Compsognathus-like theropods were primitive coelurosaurs. Tyranosauroids and maniraptoriforms were more advanced. The maniraptoriforms were the most diverse dinosaurs and are best known from Cretaceous records in Asia and North America. They include the oviraptorosaurs, ornithomimosaurs, therizinosaurs, and deinonychosaurs. It is among the deinonychosaurs, more specifically the dromaeosaurids, that most paleontologists locate the ancestry of birds.
By the Late Cretaceous, abelisaurids were the top theropod predators everywhere but Asia and North America. In these areas, tyrannosaurids dominated. They were among the last of the dinosaurs.
1. Most predatory dinosaurs were theropods.
2. Theropod phylogeny is complex and not fully agreed upon.
3. Nevertheless, theropods can be distinguished from other dinosaurs by many skeletal features, most of which identify them as bird-like, bipedal cursors. Indeed, theropods include the ancestors of birds.
4. The most primitive theropods were the Late Triassic–Late Jurassic coelophysoids and the Jurassic–Cretaceous ceratosaurs.
5. The very bird-like tetanurans were advanced theropods.
6. The megalosaurs of the Jurassic–Cretaceous were large tetanurans and include the largest of all theropods, Spinosaurus.
7. Carnosaurs were large theropods of the Jurassic–Cretaceous.
8. The tyrannosaurids, some of the largest meat-eating land animals of all time, were advanced, extremely large carnosaurs of the Late Cretaceous.
9. The coelurosaurs were advanced tetanurans with small skulls and long forelimbs, among other features. Late Jurassic
Compsognathus is a characteristic primitive coelurosaur.
10. Maniraptoriforms were advanced coelurosaurs that include the ornithomimosaurs and manirpatorans.
11. The maniraptorans encompass many taxa, including the bizarre therizinosaurs, the oviraptorosaurs, and the deinonychosaurs (“raptors”).
12. Bird ancestry lies among the deinonychosaurs.
Albertosaurus
Allosaurus
carnosaur
Ceratosaurus
Coelophysis
coelurosaur
Daspletosaurus
Deinonychus
Dilophosaurus
Dromaeosauridae
Khaan
Ornithomimosaur
Oviraptor
oviraptorosaur
Saurornithoides
Struthiomimus
Tarbosaurus
Tetanurae
Theropoda
Troodontidae
Tyrannosaurus
1. What features are diagnostic of theropods, and what types of behavior can we infer from these features?
2. What relationship exists between the theropod fossil record and our understanding of theropod phylogeny?
3. What are the bird-like features of theropods?
4. How are avetheropods distinguished from other theropods, and what is the significance of their distinctive features?
5. Draw an evolutionary tree of the coelurosaurs. Where would you place the origin of birds on this tree?
6. Compare and contrast the anatomy and behavior of Struthiomimus and Deinonychus.
Carrano, M. T., R. B. J. Benson, and S. D. Sampson. 2012. The phylogeny of Tetanurae (Dinosauria: Theropoda). Journal of Systematic Palaeontology 10:211–300. (Detailed cladistic analysis of tetanuran relationships)
Holtz, T. R., Jr. 2012. Theropods, pp. 347–378, in M. K. Brett-Surman, T. R. Holtz, Jr., and J. O. Farlow, eds., The Complete Dinosaur. 2nd ed. Bloomington: Indiana University Press. (Concise review of the theropods)
Larson, P., and K. Carpenter, eds. 2008. Tyrannosaurus rex, the Tyrant King. Bloomington: Indiana University Press. (Collection of 21 technical articles on Tyrannosaurus rex)
Lee, Y., et al. 2014. Resolving the long-standing enigmas of a giant ornithomimosaur
Deinocheirus mirificus.
Nature 515:257–260. (Describes a nearly complete skeleton of this therizinosaur)
Madsen, J. H., Jr. 1976. Allosaurus fragilis: A revised osteology. Utah Geological and Mineralogical Survey Bulletin 109:1–163. (A detailed, technical, and extensive bone-by-bone description of Allosaurus)
Ostrom, J. H. 1969. Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana. Peabody Museum of Natural History, Yale University Bulletin 30:1–165. (A comprehensive, technical description of the anatomy, behavior, and relationships of Deinonychus)
Ostrom, J. H. 1978. The osteology of Compsognathus longipes Wagner. Zitteliana [Abhandlungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie] 4:73–118. (A comprehensive, technical description of the anatomy, behavior, and relationships of Compsognathus)
Parrish, J. M., et al., eds. 2013. Tyrannosaurid Paleobiology. Bloomington: Indiana University Press. (Collection of 15 technical articles on all aspects of the biology of tyrannosaurs)
Rinehart, L. F., et al. 2009. The paleobiology of Coelophysis bauri (Cope) from the Upper Triassic (Apachean) Whitaker quarry, New Mexico, with an analysis of a single quarry block. Bulletin of the New Mexico Museum of Natural History and Science 45:1–260. (Comprehensive study of the biology of Coelophysis)
You can find a real fossil of Tyrannosaurus rex on display at several natural history museums in the United States and Canada. The type specimen of T. rex, for which the name was coined, is exhibited at the Carnegie Museum of Natural History (Pittsburgh, Pennsylvania). The American Museum of Natural History (New York) also has a skeleton on display. Legendary fossil collector Barnum Brown (1873–1963) collected these T. rex fossils in the early twentieth century, and they were the first to be described. In Chicago, the Field Museum of Natural History displays the T. rex skeleton nicknamed “Sue,” after its discoverer, Sue Hendrickson. A similarly nicknamed skeleton, “Stan,” can be seen at the museum of the Black Hills Institute of Geological Research (Hill City, South Dakota). Sue and Stan are remarkably complete skeletons of T. rex, more complete than the skeletons displayed in Pittsburgh and New York. Other skeletons and skulls of T. rex can be seen in the exhibit halls of the Smithsonian’s National Museum of Natural History (Washington, D.C.), at the Royal Tyrrell Museum of Palaeontology (Drumheller, Alberta), and at the Natural History Museum of Los Angeles County (Los Angeles, California). Most of the T. rex on display in other natural history museums are replicas (casts) made from original fossils.