PALEONTOLOGISTS long considered dinosaurs to have had the reptilian metabolism popularly referred to as cold-blooded. But, in 1970, paleontologist John Ostrom of Yale University suggested that dinosaur metabolisms may have been more mammal- or bird-like than reptilian. There soon followed an article in Scientific American by paleontologist Robert Bakker titled “Dinosaur Renaissance” in which he presented evidence and analysis to support warm-bloodedness in all dinosaurs. Bakker thereby initiated debate, still ongoing, about the nature of dinosaur metabolism. Much progress has been made in this debate, but some key issues remain unresolved. In this chapter, I review the evidence for dinosaur metabolism and evaluate the debate over this complex and fascinating subject.
Before studying dinosaur metabolism, we need to become conversant with some basic terminology and concepts. Metabolism is best defined as the chemical processes that provide energy to and repair the cells of an organism. In popular terms, vertebrate metabolisms either are cold-blooded (fishes, amphibians, and reptiles) or warm-blooded (mammals and birds). The equivalent technical terms are ectothermic (cold-blooded) and endothermic (warm-blooded or its exaggerated synonym, hot-blooded), based on the Greek roots ecto (outside), endo (inside), and thermos (temperature).
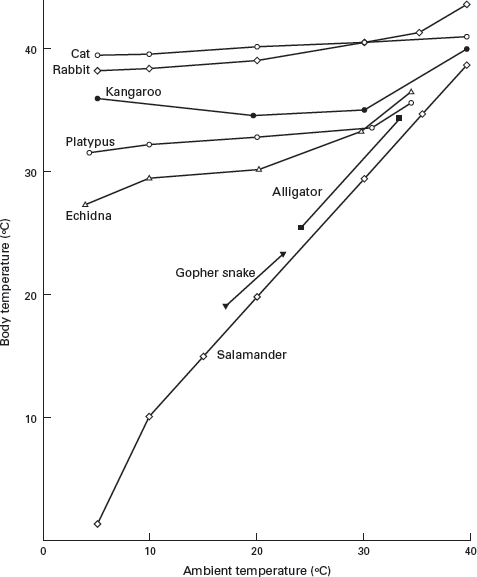
Ectotherms receive most, or all, of their body heat from an external source, usually directly from the sun (
figure 14.1). In contrast, endotherms generate most, or all, of their body heat internally. Endotherms characteristically have high rates of heat production. Ectotherms, however, typically have slower metabolisms. Vertebrates that maintain a nearly constant body temperature (usually + / − 2 degrees Celsius) are called
homeotherms, whereas those in which body temperature varies daily, seasonally, or throughout the life cycle are called
heterotherms (from the Greek
homeos [similar] and
heteros [different]).
FIGURE 14.1
Endotherms maintain a relatively constant body temperature, whereas the body temperature of ectotherms varies with the ambient (outside) temperature. (Drawing by Network Graphics)
Today, different metabolisms are found in different kinds of vertebrates. The very diverse and dominant land vertebrates, mammals and birds, are endotherms, whereas the less diverse reptiles and amphibians are ectotherms. The current success of endotherms relative to ectotherms has incorrectly fueled the notion that ectothermy is an inferior type of metabolism (
box 14.1). Also, the great difference between living endotherms and ectotherms suggests a marked dichotomy in vertebrate metabolism, with endotherms on one side and ectotherms on the other. But, endothermy and ectothermy describe only the extremes of a broad spectrum of vertebrate metabolisms (see
figure 14.1). Some ectotherms, such as tuna, maintain high body temperatures, whereas some endotherms, such as shrew tenrecs, are hard pressed to maintain a high, constant body temperature. There is great complexity and variety in existing vertebrate metabolism, and there is no reason why such variety should not have been present during the past, even in the age of dinosaurs.
The fact that today vertebrate endotherms are more successful than ectotherms has helped create the misconception that ectothermy is an inferior metabolism (
box figure 14.1). Furthermore, because we are mammals, most of us see mammals as superior to cold-blooded amphibians and reptiles. Indeed, many people find reptiles repulsive, for the reasons so well stated by Carolus Linnaeus in 1797: “Reptiles are abhorrent because of their cold body, pale color, cartilaginous skeleton, filthy skin, fierce aspect, calculating eye, offensive smell, harsh voice, squalid habitation, and terrible venom.” Small wonder that most of our pets are mammals and birds, not reptiles.
BOX FIGURE 14.1
Ectothermic reptiles are typically seen as sluggish and inferior to endothermic mammals.
But it is simply a mistake to view ectotherms as inferior to endotherms. Instead, we should realize that both represent equally viable types of vertebrate metabolism, each with advantages and disadvantages. Consider some of the advantages of the relatively slow ectothermic metabolism of an animal receiving most of its body heat from the sun. Such an animal needs to eat much less than a comparably sized endotherm, thus reducing the problems associated with obtaining food. Most ectotherms can endure much greater temperature extremes than endotherms can because their body temperature can vary widely and does not need to be held within a narrow range. These advantages explain, for example, why in some of the hottest deserts on Earth, in western Australia, ectothermic lizards abound, and there are few mammals. The endothermic mammals simply cannot find enough food or endure the extreme temperatures of these deserts.
Although ectothermy may not be inferior to endothermy, the question of superior metabolism has often been behind efforts to demonstrate dinosaurs were endotherms. Some paleontologists simply cannot believe that dinosaurs could have been as successful as they were if they had been ectotherms. No doubt, few living lizards would agree!
Metabolism is determined by chemical reactions in the enzymes, blood, and internal organs. None of these structures in dinosaurs are known to have fossilized, so it might seem there is very little direct evidence of dinosaur metabolism. Indeed, wouldn’t we need a time machine and a thermometer in order to determine the body temperatures and metabolisms of dinosaurs?
Fortunately, fossils preserve quite a variety of evidence of dinosaur metabolism, although much of it is indirect. This evidence can be organized into several categories, and most of it, as we shall see, has been interpreted in different ways.
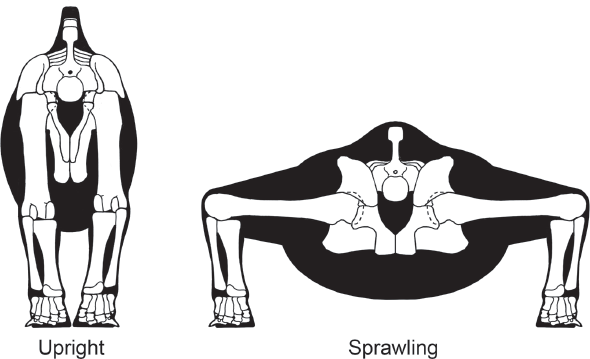
Living ectotherms have sprawling postures and gaits, whereas living endotherms, with a few exceptions, have upright postures and gaits (
figure 14.2). The exceptions are mostly aquatic mammals, such as seals and walruses, which have sprawling limbs admirably adapted to propelling them through water. Living vertebrates show a nearly perfect correlation between posture and metabolism. This correlation may indicate that extinct animals with an upright posture, the dinosaurs, were endotherms.
FIGURE 14.2
Upright posture characterizes living endotherms, whereas living ectotherms have a sprawling posture. (© Scott Hartman)
Two criticisms can be leveled at this conclusion. First, no cause-and-effect relationship has been established between posture and metabolism. Maybe ectotherms can have an upright posture, maybe some (the dinosaurs?) did in the past, and it is only a coincidence that all living ectotherms happen to be sprawlers.
A second criticism is that not all dinosaurs may have had an upright posture. Stegosaurs, ankylosaurs, and ceratopsians have been reconstructed with upright hind limbs and sprawling or semi-sprawling forelimb postures. What does this indicate about their metabolisms? Does it mean their posterior halves were endothermic and anterior halves ectothermic? Of course not. It probably indicates that factors other than metabolism, such as the heavy skulls and armor of stegosaurs, ankylosaurs, and ceratopsians, which forced a slow and powerful sprawling forelimb posture, also determined a dinosaur’s posture.
Despite these criticisms, the correlation between posture and metabolism in living vertebrates is striking. A subset of this correlation, that all living bipeds are endotherms, also suggests that bipedal dinosaurs were endotherms. But, to believe that these correlations indicate that dinosaurs were endothermic, we must agree that posture is controlled by metabolism, a conclusion still open to some debate.
High levels of activity characterize living endotherms, whereas today’s ectotherms generally are slower, more sluggish, and less agile. The qualifier “generally” needs to be used here because some living ectotherms, for example sea turtles, are capable of great speed and/or agility, if only for short periods of time. Nevertheless, evidence for speed, sustained activity, and agility in dinosaurs would be more consistent with their having had an endothermic, rather than ectothermic, metabolism.
Speed, sustained activity, and agility in dinosaurs is evident in their skeletal structures, brain size, brain complexity, and in the speeds estimated from dinosaur trackways. As we saw in
chapters 4 through
9, the skeletons of some dinosaurs, especially small theropods and ornithopods, in many ways resemble those of fast-running birds and mammals. Key features of this resemblance include elongate, slender limbs and limb joints, indicating an ability to flex acutely at the elbow, wrist, knee, and ankle joints. Hollow bones, large claws, limb proportions indicative of habitual bipedality, and long, rigid tails for precision balance while running contribute to the impression of speed and agility in some dinosaurs (
figure 14.3). Some theropod and ornithopod dinosaurs thus seem to have been every bit as quick and agile as living birds and mammals, suggesting that these dinosaurs may have been endotherms.
Some paleontologists assert that the larger dinosaurs, such as ceratopsians, galloped like living endothermic rhinoceroses. But this argument is inconsistent with the limb structures and trackways of the large, quadrupedal plant-eating dinosaurs, which indicate they were slow, powerful walkers. The skeletal evidence for speed, high levels of activity, and agility in dinosaurs, and the inference of endothermy from this evidence, is mostly confined to theropods and ornithopods.
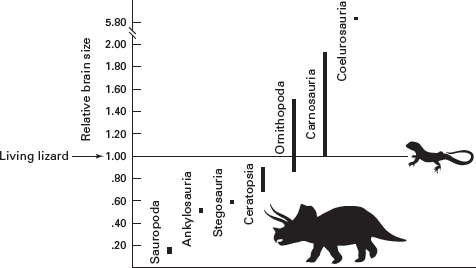
Relative brain size and complexity provide a second line of evidence for speed, high activity levels, and agility in some dinosaurs. These characteristics of living endotherms require great motor and sensory control by larger and more complex brains than those of living ectotherms. If we compare the relative brain sizes of dinosaurs to that of a living ectothermic lizard, theropods and most ornithopods appear to have been relatively “brainy” (
figure 14.4). But sauropod, ankylosaur, stegosaur, and ceratopsian brain sizes fall well below the line set by the lizard.
FIGURE 14.3
The theropod Deinonychus well displays many skeletal features suggestive of speed, sustained high activity, and agility. (© Scott Hartman)
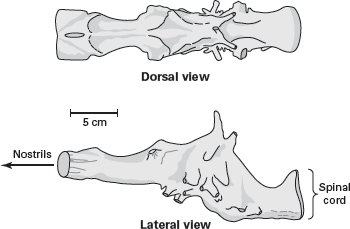
Brain complexity in dinosaurs can be inferred only from endocasts of empty fossil skulls (see
box 8.2). An empty skull acts as a mold that reproduce the brain’s overall configuration and location and the number of associated blood vessels and nerves. Dinosaur brain casts, however, reveal typically reptilian levels of brain complexity (
figure 14.5). These casts do not preserve special structures, such as extremely large cerebral hemispheres, that might be linked to speed, high activity levels, and agility.
FIGURE 14.4
Dinosaurs had a range of relative brain sizes, some larger and some smaller than that of a living lizard. (Drawing by Network Graphics)
FIGURE 14.5
The brain of Tyrannosaurus rex is typically reptilian in its level of complexity. (Drawing by Network Graphics)
A third line of evidence for dinosaur speed, activity, and agility comes from their trackways. In
chapter 12, we saw that dinosaur speeds can be estimated from trackways. Most dinosaur trackways indicate slow walking, which is the normal speed of all living vertebrates, ectotherms and endotherms alike. But, a few trackways document small theropods running as fast as a living antelope.
So, the evidence for speed, high activity levels, and agility among dinosaurs seems to support endothermy only in small theropods and ornithopods. But some paleontologists point out, as I did at the beginning of this discussion, that some living ectotherms can be very fast, active, and agile. It might be that speedy, active, and agile theropods and ornithopods were ectotherms.
Living endotherms maintain a constant, high body temperature and a fast metabolism by consuming and processing large amounts of food (energy). This means they eat more relative to body weight and process that food more quickly than do ectotherms. Key to rapid processing are the teeth, jaws, and skulls of many endotherms, which allow the food to be broken down rapidly into small pieces, thereby increasing the food’s surface area and releasing important nutrients so that the endothermic digestive system can rapidly assimilate the nutrients. If dinosaurs had tooth, jaw, and skull structures that indicated extensive processing of the food in the mouth, this might be evidence that they had an endothermic metabolism.
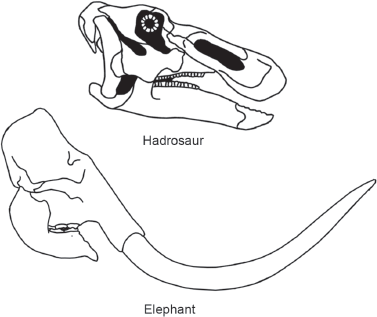
When we look at the teeth, jaws, and skulls of dinosaurs, however, only among ornithopods do we find structures similar to the “
food processors” of living endotherms. The powerful jaws and extensive dental batteries of ornithopods, such as hadrosaurs, are similar to the jaws and teeth of living endothermic horses and elephants that extensively grind the vegetation they consume (
figure 14.6).
FIGURE 14.6
The dental batteries and powerful jaws of hadrosaurs resemble those of some living plant-eating mammals, such as the elephant. (© Scott Hartman)
This might be taken to indicate that only the ornithopods possessed “food processors” suggestive of endothermy. But, the teeth, jaws, and gastric mills of some theropods, prosauropods, and some of the armored ornithischians don’t indicate food processing inferior to that of living plant-eating birds. Predatory theropod food processing doesn’t look much different from the slicing of meat undertaken by living meat-eating mammals, such as wild dogs and cats. We also need to remember that the large size of most dinosaurs would have forced them to process and consume large amounts of food, whether they were ectotherms or endotherms. Furthermore, the ornithopod grinding mechanism may simply reflect their dietary preference for tough, hard-to-process plant foods.
We simply cannot be certain what the “food processors” of dinosaurs indicate about their metabolism. Many needed to consume large amounts of food to support huge body masses. Specialization in tough food items, or the need to feed a rapid, endothermic metabolism, could explain the tooth, jaw, and skull structures of some dinosaurs, especially the ornithopods.
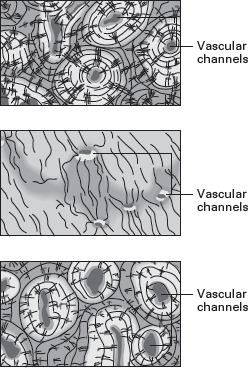
The compact (external) layer of bone of many living ectotherms contains few channels for blood vessels. In contrast, the
compact bone of many living endotherms is full of large numbers of blood vessels (
figure 14.7). This is thought to reflect the quick metabolism of endotherms, which requires the rapid exchange of elements stored in bone, such as calcium and phosphorus. It stands to reason that if dinosaur compact bone had many channels for blood vessels, then this would indicate endothermy.
FIGURE 14.7
Dinosaur compact bone (below) has many channels for blood vessels, like that of living endotherms (above), and unlike that of living ectotherms (center). (Drawing by Network Graphics)
Well-preserved dinosaur bone can be cut into thin wafers, and the microscopic structure of the bone can be determined. This has been done for all major groups of dinosaurs and reveals compact bone with numerous channels similar to that of many living endotherms (see
figure 14.7).
This might seem conclusive evidence for endothermy in all dinosaurs, but the correlation between metabolism and compact-bone microstructure is not a perfect one in living vertebrates. Bone from different parts of the skeleton (for example, limbs versus vertebrae) has different microstructure, in part because of the different stresses placed on different parts of the body. Also, some large ectotherms (turtles and crocodiles) have compact bone with many channels, whereas some small living endotherms (certain mammals and birds) lack numerous blood channels in their compact bone. This suggests that bone microstructure may be more related to size than to metabolism, making the bone microstructure of dinosaurs inconclusive evidence of endothermy.
Another aspect of dinosaur bone microstructure is the presence of growth rings in the bone. These growth rings have been found in the bone of many kinds of dinosaurs, including ornithopods, theropods, and ceratopsians. The rings are characteristic of the bone of living ectothermic reptiles. Periodic pauses in bone growth due to seasonal (or annual) temperature fluctuation cause these rings to form. However, the bone of some dinosaurs, such as the ornithopod Dryosaurus, lacks growth rings and more resembles endothermic bone. Furthermore, the microstructure of some dinosaur bone closely resembles the microstructure of bird bone, which indicates rapid bone growth in dinosaurs, as in endothermic birds.
An endothermic metabolism requires high
blood pressure and rapid blood circulation to move energy quickly through the body. Thus, living endotherms have consistently higher blood pressures than do ectotherms (
figure 14.8). If we could estimate dinosaur blood pressures, this might indicate whether they were endotherms.
But, how can we do this? It turns out quite easily if we recognize that a primary function of the vertebrate heart is to pump blood to the brain, which is usually elevated above the level of the heart. In other words, the vertical distance between the heart and the brain should be related, in some way, to blood pressure, because sufficient pressure must be maintained to move blood to the brain, or the animal will die.
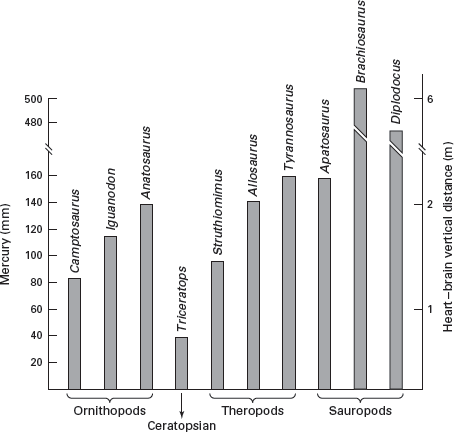
It is possible to estimate blood pressures based on the vertical distance between the dinosaur’s heart and its brain (
figure 14.9). Such estimates are based on the relationship between blood pressure and the distance between the heart and the brain of living vertebrates and also require certainty of the posture of the dinosaur. These estimates suggest high endothermic blood pressures for most dinosaurs (
Triceratops is a notable exception) and incredibly high blood pressures (more than 400 millimeters of mercury!) for sauropods. It is questionable whether blood pressures as high as those estimated for sauropods could be maintained by any vertebrate circulatory system without an explosion. Thus, it seems likely that sauropods may have used arterial valves or muscular contractions in the neck, as do living giraffes, to help bring blood to the brain and thus would have had much lower blood pressures than estimated. Estimates of blood pressures, however, are consistent with endothermy in most dinosaurs, although critics point out that large vertical heart–brain distances in dinosaurs could be the determining factor here, not metabolism.
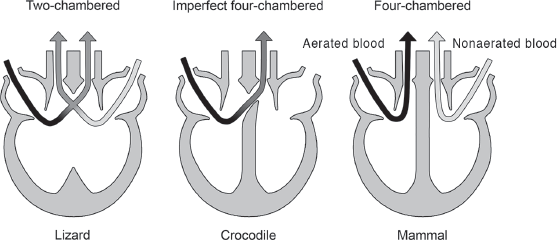
A second, and speculative, aspect of dinosaur blood pressure concerns the structure of the dinosaurian heart. Living endotherms have a fully divided
four-chambered heart that separates aerated from nonaerated blood and thus acts as a double pump, producing high pressure for the aerated blood and lower pressure for the nonaerated blood. This mechanism is especially significant because lowering the pressure of the nonaerated blood prevents the rupture of tiny blood vessels characteristic of the lungs of endotherms. In contrast, ectothermic hearts have only two chambers that do not efficiently separate the aerated from the nonaerated blood. An exception is living crocodiles, which have an imperfectly divided four-chambered heart (
figure 14.10).
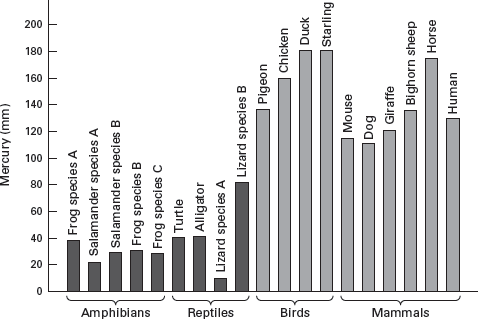
FIGURE 14.8
Living endotherms have higher blood pressures than do ectotherms. (Drawing by Network Graphics; modified from J. H. Ostrom. 1980. The evidence for endothermy in dinosaurs, pp. 82–105, in R. D. K. Thomas and E. C. Olson, eds., A Cold Look at the Warm-Blooded Dinosaurs. Copyright © 1980 American Association for the Advancement of Science)
FIGURE 14.9
Dinosaur blood pressures can be estimated from their heart–brain vertical distances. (Drawing by Network Graphics; modified from J. H. Ostrom. 1980. The evidence for endothermy in dinosaurs, pp. 82–105, in R. D. K. Thomas and E. C. Olson, eds., A Cold Look at the Warm-Blooded Dinosaurs. Copyright © 1980 American Association for the Advancement of Science)
FIGURE 14.10
Living endotherms have a four-chambered heart, crocodiles have an imperfectly divided four-chambered heart, and other ectotherms have a two-chambered heart. (© Scott Hartman)
The closest living relatives of dinosaurs—birds and crocodiles—have four-chambered hearts. So, many paleontologists believe that dinosaurs had four-chambered hearts as well. Dinosaur hearts have not fossilized, so this remains speculation. Indeed, a supposed “fossilized heart” of a dinosaur, with four chambers, has been demonstrated to be nothing more than a piece of ironstone. But, if dinosaurs did have four-chambered hearts, and if their high estimated blood pressures don’t simply reflect large vertical heart–brain distances, then these lines of evidence suggest endothermy among most dinosaurs.
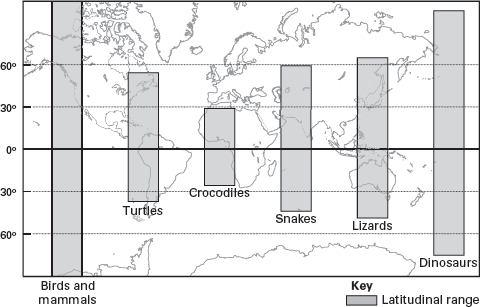
Today, ectotherms cannot live in the extremely cold climates near the poles simply because there is not enough solar energy there with which to warm their bodies (
figure 14.11). However, endothermic mammals and birds, such as polar bears and penguins, are capable of living in those colder regions of the globe. So, if the geographic distribution of dinosaur fossils indicates that they lived in cold climates, this also might suggest endothermy in dinosaurs.
The current geographic distribution of dinosaur fossils encompasses Cretaceous localities as far north as Alaska, the Northwest Territories of Canada, and Svalbard (Spitzbergen) and as far south as Antarctica. These are places where ectotherms do not live today. But, the Cretaceous world was not as cold toward the poles as is today’s world, and because of continental drift, these Cretaceous dinosaur localities were also not as poleward as they are today. Despite this, these locations would still have been far enough poleward during the Cretaceous to have experienced the “polar night”—winter months of virtually continuous darkness. This darkness would have excluded ectotherms from living in poleward regions, even if there were no polar ice caps and temperatures were warmer during the Cretaceous.
FIGURE 14.11
Today, ectotherms do not live in cold, poleward regions. (Drawing by Network Graphics)
It also has been suggested that the poleward Cretaceous dinosaur fossils do not represent dinosaurs that lived in these regions year round. Instead, they may represent dinosaurs that migrated over large areas and were only living poleward during the warmer, brighter portion of the year. So, a hadrosaurid might have migrated the 3,000 kilometers from Alaska to Alberta in a given year, a trek of 60 days at a speed of 50 kilometers per day. To have migrated so far, a dinosaur would have to be endothermic—or would it? Given the possibility of migration, it is difficult to argue that the geographical distribution of dinosaurs tells us something conclusive about their metabolism.
Dinosaurs are the ancestors of birds (see
chapter 15). Living birds are endotherms, and it may be that all extinct birds, including the Late Jurassic
Archaeopteryx, the first bird, were endotherms. Indeed, some have argued that the small size, skeletal structure, insulating feathers and powered flight of
Archaeopteryx strongly suggest an endothermic metabolism (
figure 14.12). The question thus arises, did endothermy in birds first evolve in
Archaeopteryx, or did the theropod ancestors and close relatives of birds have an endothermic metabolism? Or, how many times did endothermy arise in the origin and evolution of birds?
FIGURE 14.12
Living birds are endotherms.
This question can’t be answered definitively. As already discussed, many features of the small theropods are consistent with endothermy. Indeed, some small theropods from the Lower Cretaceous of China have feather-like body coverings. These and many other theropods are strikingly bird-like. That the theropods gave rise to the birds and are very bird-like in many ways is also consistent with endothermy. Although the endothermy of birds may suggest endothermy in at least some theropods, it provides no clues to the metabolism of the other dinosaurs.
Today, complex social behaviors are characteristic of many endotherms and uncommon among ectotherms. The evidence reviewed in
chapter 13 suggests that some dinosaurs may have lived in groups and had some form of sociality based on their trackways, visual display, and inferred parental care of young dinosaurs. On face value, this is consistent with endothermic dinosaurs.
But, some living reptiles form social groups to hunt and administer minimal care to their young, and not all living endotherms form social groups; some live solitary lives. Indeed, social behavior is not normally viewed as being caused by metabolism, but instead is related to other factors, such as the distribution of food resources in an animal’s habitat. So, the inferred social behavior of some dinosaurs at best is consistent with, but not strong evidence for, their endothermy.
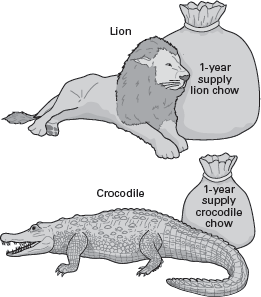
Living endotherms need to consume more energy than do comparably sized ectotherms. This means that a 150-kilogram lion eats more food, and more frequently, than a 150-kilogram crocodile. Therefore, in the wild, a lion must have access to more food items (prey) than a crocodile (
figure 14.13). The
predator–prey ratio, the ratio of the body mass of predators to that of their potential prey, thus should differ for endothermic and ectothermic predators. Thus, endothermic predator–prey ratios should be lower (less predator mass per prey) than the ratio for ectotherms. Extrapolating this to dinosaurs, one can predict that predatory dinosaurs should be rare relative to their potential prey dinosaurs if the predatory dinosaurs were endotherms and more common if they were ectotherms.
FIGURE 14.13
A 150-kilogram lion (endotherm) eats more food than a 150-kilogram crocodile (ectotherm) in a given period of time. (Drawing by Network Graphics)
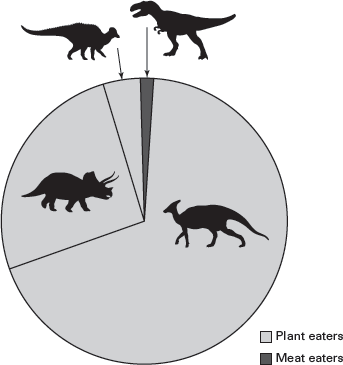
FIGURE 14.14
Predatory dinosaurs are extremely rare in the dinosaur collections from the Upper Cretaceous of Dinosaur Provincial Park in Alberta, Canada. (Drawing by Network Graphics)
A survey of dinosaur fossil collections reveals that predatory dinosaurs are relatively rare. For example, the most famous dinosaurian predator of all time,
Tyrannosaurus rex, is known from a few skeletons. One of the most extensive dinosaur collections ever made, from the Upper Cretaceous badlands in Dinosaur Provincial Park, in Alberta, Canada, contains only 3 to 5 percent predatory dinosaurs (
figure 14.14). On face value, the scarcity of predatory dinosaurs suggests that they (and only they, because this fact tells us nothing about the metabolism of their prey) were endotherms.
There are, however, two insurmountable problems with using predator–prey ratios to infer dinosaur metabolism. First, it is not clear that the ratio of predators to prey today is determined simply by the food requirements of the predators. Other factors, such as food availability for the prey, are important as well. The second problem, and a very large one, is that we cannot be certain that collections of dinosaur fossils reflect the actual predator–prey ratios of dinosaurs. Taphonomic processes destroy many potential fossils and may have biased the dinosaur fossil record against predators or toward prey (see
box 3.1). If dinosaur predators and prey did not always inhabit the same environments, then fossil collections would not necessarily reflect their actual abundances. In light of these problems, the predator–prey ratios of dinosaurs seem an unsatisfactory way to infer the metabolism of predatory dinosaurs.
Body size has already been mentioned as a factor in the evaluation of several of the lines of evidence for dinosaur metabolism. It is a very important factor in any consideration of vertebrate metabolism because body size influences metabolism, and the metabolism of a vertebrate must be consistent with its body size.
To understand why this is so, we need to understand the relationship between the body size of an animal and its surface area. This relationship is best portrayed by two spheres, one large and one small (
figure 14.15). The volume of a sphere is 4/3π
r3, where
r is the radius of the sphere. The surface area of a sphere is 2/3π
r2, where, again,
r is the radius. As a sphere becomes larger, its volume increases as the cube of its radius, whereas its surface area increases only as a square of its radius. The guaranteed result is that a larger sphere has a smaller surface area relative to its volume than does a smaller sphere, which has much more surface area relative to volume.
Although this may not strike you as intuitively correct, a quick calculation should convince you. Consider two spheres, one with a radius of 5 centimeters and the other with a radius of 10 centimeters. Calculate the surface areas and volumes of both spheres, and then divide the surface area of each sphere by its volume. The result is that the surface area of the smaller sphere is 59 percent of its volume, whereas the surface area of the larger sphere is only 30 percent of its volume. Smaller spheres have larger surface areas relative to larger spheres.
If we transfer this basic geometry to animals, we realize that small animals have much more surface area relative to their volume than do large animals. Much of metabolism is generating body heat, and the surface area of an animal determines how readily it can acquire heat from an external source, or how rapidly it loses its body heat. Small animals, such as mice and hamsters, have such large surface areas relative to their volume (or mass) that they lose heat very fast. As endotherms, they have an insulating coat of fur to help retard heat loss and use their very rapid metabolism to generate more heat to replace that being lost rapidly. They also shiver frequently to generate more body heat and hide in burrows or under vegetation to slow heat loss.
FIGURE 14.15
Because of the surface-area-to-volume relationship, smaller spheres (and animals) have more surface area relative to volume than larger spheres (and animals). (Drawing by Network Graphics)
In contrast, an elephant has a much lower surface-area-to-volume ratio than a mouse. Its problem, as an endotherm, lies in overheating because an elephant loses heat very slowly. For this reason, elephants have little in the way of insulating body hair and use their large ears as heat radiators by pumping blood into large vessels in the ears, thereby cooling it by increasing the blood’s surface area. Elephants also employ behavioral mechanisms to avoid overheating, such as bathing in rivers several times a day.
Ectotherms that receive most of their body heat from the sun also are affected by the surface-area-to-volume relationship. Because of their relatively large surface areas, small lizards can heat and cool themselves rapidly. But, because of their large size, crocodiles are slower to warm up and cool down.
Dinosaurs also must have been affected by the
surface-area-to-volume relationship, and this must constrain our interpretation of dinosaur metabolism. Large dinosaurs, those that weighed about 1,000 kilograms or more, had very low surface areas relative to their volumes. This has inspired calculations that suggest that very large dinosaurs, especially full-grown sauropods, would have overheated if they had had an endothermic metabolism. Indeed, these dinosaurs would have been inertial homeotherms (“
gigantotherms”), huge animals with a nearly constant body temperature (homeotherms) due to their large size (which, via the surface-area-to-volume relationship, produces thermal inertia) even though they might have had relatively slow, ectothermic metabolisms (
box 14.2). Small dinosaurs, including baby and juvenile sauropods, based on their surface-area-to-volume relationship, could have been either ectotherms or endotherms.
Dinosaur body size considered in light of the surface-area-to-volume relationship makes endothermy in adult sauropods and some of the other larger dinosaurs (those with a body weight well above 1,000 kilograms) seem improbable. It suggests that the largest dinosaurs were gigantotherms, but leaves open the question of the metabolism of small dinosaurs. Indeed, small juvenile dinosaurs may have had a metabolism different from their metabolism later in life at large adult sizes, so many dinosaurs could have been heterotherms.
The most common form (what chemists call an isotope) of the oxygen atom in nature has an atomic weight of 16 (abbreviated 16O), whereas the next most common form has an atomic weight of 18 (18O). The bone of any vertebrate incorporates both forms of oxygen into its mineral matrix. The relative amount (ratio) of 18O to 16O in the bone depends on temperature. Some scientists have argued that in an endotherm, the ratio of 18O and 16O should vary little between the limb bones and the bones in the core of the body (such as vertebrae) because the body temperature is nearly the same at both locations. Conversely, they argue, the temperatures of the extremities and body core are very different in an ectotherm, so the oxygen ratios should be very different at each location.
A consistency of oxygen ratios has been demonstrated in the extremities and core of the skeleton of
Tyrannosaurus rex, suggestive of endothermy. However, there may be a problem with the basic argument underlying this conclusion. Various studies of living mammals and birds show that temperature varies considerably between their cores and extremities, at least as much as in living alligators. Also, factors other than body temperature, including diet and water consumption, can affect the oxygen isotope ratios in bone. The reasoning behind using oxygen ratios to determine metabolism thus appears to be flawed.
Chapter 6 discussed the metabolism of sauropod dinosaurs, identifying them as animals that maintained a nearly constant body temperature by virtue of their great bulk; such creatures are referred to as inertial homeotherms. The term “inertial homeotherm” can be replaced by the more colorful word
gigantotherm, referring to an animal that maintains a constant, high body temperature by virtue of its large size.
How large does an animal have to be to be a gigantotherm? Recent studies suggest that ectotherms at least as large as 1,000 kilograms are gigantotherms. The most studied example of a living ectothermic gigantotherm is the leatherback turtle. This large reptile lives in the sea, migrating from the tropics to the Arctic oceans on a regular basis. The 1,000-kilogram adult leatherbacks have metabolic rates well below those of comparably sized mammals. Yet gigantothermy enables leatherbacks to stay warm in frigid waters. If anything, the leatherbacks have trouble dumping heat and staying cool in tropical waters and on the warm beaches where they lay their eggs.
BOX FIGURE 14.2
Very large dinosaurs, such as sauropods, were gigantotherms. (© Mark Hallett. Reproduced with permission of Mark Hallett Paleoart)
The example of the leatherback shows that large size can be conducive to maintaining a constant body temperature. All large vertebrates, especially the large dinosaurs (
box figure 14.2), must experience some degree of gigantothermy.
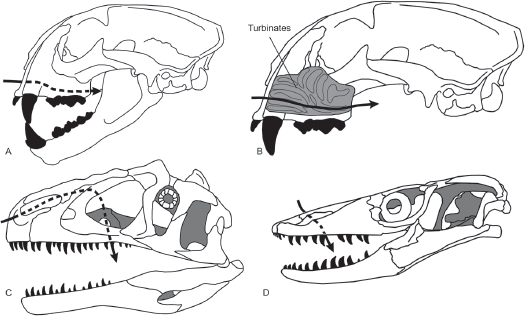
FIGURE 14.16
Living endotherms (A) have respiratory turbinates (B) that warm and moisten the air that is inhaled. Theropod dinosaurs (C) lacked respiratory turbinates, so air would have flown directly into the mouth, as in a living ectothermic lizard (D). (© Scott Hartman)
Living mammals and birds have small bones in their nasal passages called
respiratory turbinates (
figure 14.16). These bones increase the surface area over which blood and moist tissues are exposed to air. By conserving water, respiratory turbinates can thus play a role in the rapid breathing and high rate of oxygen consumption characteristic of the endotherms.
Most dinosaur skulls appear to lack respiratory turbinates, so air would have flowed directly from their nostrils into their mouths without first being warmed or moistened. Some argue that this provides strong evidence that dinosaurs were not endotherms like living birds and mammals. Nevertheless, recent studies show that in living birds, respiratory turbinates are not important for water conservation. These endotherms conserve water in other ways, and endothermic dinosaurs may also have conserved water without using respiratory turbinates. Thus, the case for dinosaur ectothermy based on their lack of respiratory turbinates is not as strong as has been argued by some paleontologists.
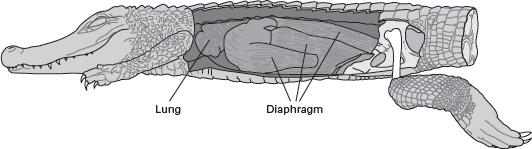
Living tetrapods have two kinds of lungs. Mammals have
alveolar lungs made of millions of tiny, very vascularized, spherical alveoli. Other tetrapods, including birds, have
septate lungs in which the entire lung is like one huge alveolus and has vascularized septa that penetrate it from its perimeter. Airflow (ventilation) is bidirectional in both kinds of lung, but there are two very different ways the lungs are ventilated. Mammals and crocodiles use a diaphragm muscle between the lung and the pelvis to act like a piston to change lung volume (
figure 14.17). In contrast, birds expand and contract their rib cage, thus using their jointed ribs and sternum and their rib (costal) muscles to change the volume of their lungs.
FIGURE 14.17
Dinosaurs likely had septate lungs ventilated by a diaphragm muscle, like living crocodiles. (Drawing by Network Graphics)
Given their relationship to birds and crocodiles, dinosaurs almost certainly had septate lungs. But, they do not have jointed ribs or sternal bones. Indeed, their hip structure suggests dinosaurs used a diaphragm-like piston to ventilate their lungs. Archaeopteryx is similar to the dinosaurs in this regard. Thus, the question is, did dinosaurs ventilate their lungs rapidly enough to maintain an endothermic metabolism? In other words, would a diaphragm muscle ventilate a septate lung rapidly enough to provide enough oxygen to an endotherm? This is a tough question to answer, given that no living endotherm ventilates a septate lung with a diaphragm. Lung structure thus seems to tell us little about dinosaur metabolism.
Having reviewed many lines of information bearing on hypotheses of dinosaur metabolism, you should see how difficult it is to present a simple answer to the question about what type(s) of metabolism dinosaurs had. The following answers, however, are offered by different paleontologists:
1. All dinosaurs were ectotherms.
2. All dinosaurs were endotherms.
3. Dinosaurs were a diverse group of animals metabolically; some (at least some theropods and ornithopods) were endotherms, and others were ectotherms.
4. Large dinosaurs, the sauropods and big ornithischians, were gigantotherms as adults. Juveniles of these large dinosaurs, and the smaller dinosaurs, may have been either endotherms or ectotherms.
The information presented in this chapter suggests that neither of the first two extreme views of dinosaur metabolism is correct. The third and fourth views, some combination of them or some variant, appear to best explain the evidence. There is, indeed, no simple answer to the question of what type of metabolism the dinosaurs had.
1. Warm-blooded vertebrate metabolisms are endothermic and fast.
2. Cold-blooded vertebrate metabolisms are ectothermic and slow.
3. Numerous lines of evidence have been brought to bear on the question of dinosaur metabolism, including posture and gait; speed, activity level, and agility; feeding adaptations; bone microstructure; blood pressure; geographic distribution; bird ancestry; social behavior; predator–prey ratios; body size; the ratio of oxygen isotopes in bone; the presence versus absence of respiratory turbinates; and lung function.
4. Many of these lines of evidence are consistent with endothermy in at least some theropods and ornithopods, but most evidence does not support endothermy in the other dinosaurs.
5. Very large dinosaurs, such as the sauropods, were gigantotherms.
6. The evidence does not support extreme views of dinosaur metabolism; that is, that all dinosaurs were ectothermic or all were endothermic.
7. The evidence suggests a probable variety of metabolisms in dinosaurs, including endotherms and ectotherms, some of which may also have been heterotherms or gigantotherms.
alveolar lung
blood pressure
bone microstructure
brain complexity
cold-blooded
compact bone
ectothermic
endothermic
“food processor”
four-chambered heart
gigantotherm
heterotherm
homeotherm
hot-blooded
metabolism
“polar night”
predator–prey ratio
relative brain size
respiratory turbinate
septate lung
surface-area-to-volume relationship
warm-blooded
1. Define the following terms: endotherm, ectotherm, homeotherm, heterotherm, gigantotherm.
2. What are some common misconceptions about ectotherms, and why are they wrong?
3. Which of the 13 lines of evidence presents the most convincing evidence for endothermic dinosaurs? Which presents the weakest?
4. Explain the surface-area-to-volume relationship and its bearing on dinosaur metabolism.
5. What type(s) of metabolism did dinosaurs have? Defend your answer.
Bakker, R. T. 1975. Dinosaur renaissance. Scientific American 232:48–78. (The now classic, original article that argues all dinosaurs were endotherms)
Bakker, R. T. 1986. The Dinosaur Heresies. New York: Morrow. (Much of this book argues the case for endothermic dinosaurs)
Cleland, T. P., M. K. Stoskopf, and M. H. Schweitzer. 2011. Histological, chemical, and morphological reexamination of the “heart” of a small Late Cretaceous Thescelosaurus. Naturwissenschaften 98:203–211. (Re–evaluates the supposed fossil dinosaur heart and concludes that it is inorganic)
Paul, G. 2012. Evidence for avian-mammalian aerobic capacity and thermoregulation in Mesozoic dinosaurs, pp. 819–872, in M. K. Brett-Surman, T. R. Holtz, Jr., and J. O. Farlow, eds., The Complete Dinosaur. 2nd ed. Bloomington: Indiana University Press. (Argues the case for dinosaur endothermy)
Reid, R. E. H. 2012. “Intermediate” dinosaurs: The case updated, pp. 873–921, in M. K. Brett-Surman, T. R. Holtz, Jr., and J. O. Farlow, eds., The Complete Dinosaur. 2nd ed. Bloomington: Indiana University Press. (Argues the case for unique dinosaur metabolism, somewhere between ectothermy and endothermy)
Ruben, J. A., et al. 2012. Metabolic physiology of dinosaurs and early birds, pp. 785–817, in M. K. Brett-Surman, T. R. Holtz, Jr., and J. O. Farlow, eds., The Complete Dinosaur. 2nd ed. Bloomington: Indiana University Press. (Argues the case for dinosaur ectothermy)
Spotila, J. R., M. P. O’Connor, P. Dodson, and F. V. Paladino. 1991. Hot and cold running dinosaurs: Body size, metabolism and migration. Modern Geology 16:203–227. (Presents the evidence for dinosaur gigantothermy)