FEW animals are more awe inspiring than the largest land animals of all time, the sauropod dinosaurs. Their anatomy, classification, lifestyle, and evolution are the focus of this chapter, which begins with a consideration of their closest relatives, the prosauropod dinosaurs.
The prosauropods and the sauropods together constitute a group of dinosaurs with the ungainly name Sauropodomorpha. They are distinguished from other dinosaurs by having heads that were very small relative to their bodies, spatulate (spoon-shaped) teeth, at least 10 elongated vertebrae in relatively long necks, short feet, and very large claws on the first digits of their forefeet. Sauropodomorphs spanned the entire age of dinosaurs, from the Late Triassic until the end of the Cretaceous. They were one of the most successful groups of plant-eating dinosaurs as well as the result of nature’s most amazing experiment in animal gigantism.
Prosauropods represent one of the first evolutionary diversifications of plant-eating dinosaurs. Known from fossils found on all continents except Antarctica (
figure 6.1), they range in age from Late Triassic to Early Jurassic and so include some of the oldest dinosaurs. Prosauropods were sauropod-like in general build, though much smaller and more slender.
Prosauropods were once considered ancestral to sauropods (hence their name). Indeed, some paleontologists believed the “missing link” between the two groups was provided by prosauropods such as
Vulcanodon, from the Lower Jurassic of southern Africa, and
Riojasaurus, from the Upper Triassic of Argentina (
figure 6.2). But all prosauropods, including these two types, had some skeletal features, such as the small size of the fifth digit of the hind foot, the existence of which would require a highly unlikely reversal in evolution were prosauropods truly the ancestors of sauropods. Sauropods had large fifth digits on the hind foot, as did, presumably, the ancestors of prosauropods. Therefore, paleontologists now favor the view that prosauropods were the closest relatives, but not the ancestors, of sauropods and that both groups derived from a common ancestor, as yet undiscovered (
figure 6.3).
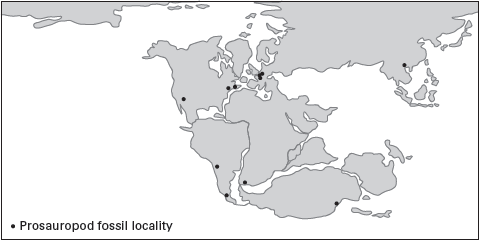
FIGURE 6.1
This map of the occurrences of some well-known prosauropods indicates their worldwide distribution during the Early Jurassic. (Drawing by Network Graphics)
Typical prosauropods include the massive, heavily built 17-meter-long Riojasaurus from Argentina, the lightly built Anchisaurus from the eastern United States, and Yunnanosaurus from southern China, with its chisel-shaped teeth reminiscent of those of some later sauropods. Plateosaurus is a particularly well-known and characteristic prosauropod from the Late Triassic of western Europe.
One of the best-known prosauropods is
Plateosaurus, from the Upper Triassic of western Europe (
figure 6.4). Mass death assemblages of this large (6- to 8 meters in body length) prosauropod are known from Trössingen, Germany, where the animals may have been killed and their skeletons accumulated by mud slides. The complete skeletons of
Plateosaurus from Trössingen give us remarkable knowledge of this early prosauropod.
FIGURE 6.2
Riojasaurus, an 11-meter-long prosauropod from the Upper Triassic of Argentina, may best approximate the ancestry of sauropods. (© Scott Hartman)
FIGURE 6.3
Although some paleontologists see prosauropods as the ancestors of sauropods, the two groups are best regarded as close relatives derived from an as yet undiscovered ancestor. (Drawing by Network Graphics)
Salient features of the skeleton of Plateosaurus include its large size and robustness, the numerous coarsely serrated teeth, and the jaw joint below the level of the teeth. Its small head was attached to a long neck, followed by a long back and tail, in proportions characteristic of prosauropods. The pelvis was typically saurischian and anchored massive hind limbs. The first finger of the hand bore a very large claw, and the fourth and fifth digits of the hind foot were small and must not have borne much weight when the animal walked.
Prosauropods are often restored as bipedal animals (see
figure 6.4), but their footprints indicate that they also could have walked quadrupedally (
figure 6.5). Indeed, the forelimbs of prosauropods were at least two-thirds the length of their hind limbs, as in the later diplodocid sauropods, which are generally considered to have been obligatory quadrupeds. The first digit of the hand was much larger than the others and bore a huge claw, and that digit must have been held clear of the ground while walking. There is little doubt that prosauropods could rear up on their hind limbs to reach vegetation in tall trees, which is consistent with the evidence that prosauropods were herbivores. A large number of features suggest that prosauropods ate plants:
FIGURE 6.4
The skeleton of 6- to 8-meter-long Plateosaurus, from the Upper Triassic of western Europe, shows many characteristic features of prosauropods. (© Scott Hartman)
FIGURE 6.5
Prosauropod footprints like these indicate that prosauropods did sometimes walk on all fours. In the upper-right portion of the drawing, note both hind foot and forefoot impressions. (Drawing by Network Graphics)
1. Their long necks would have extended their vertical feeding range so that they could have cropped vegetation from tall trees, as do living giraffes.
2. The jaw hinge was below the line of the upper tooth row, as in plant-eating ornithischians.
3. The offset of the jaws of prosauropods ranged from small to large. This offset allowed the tooth rows to be almost parallel so that contact would have occurred nearly simultaneously along their entire length when chewing, as in many living plant-eating mammals.
4. The spatulate teeth of prosauropods had about 20 coarse serrations per tooth that were set at an angle of about 45 degrees to the cutting edge. They thus bore an uncanny resemblance to the teeth of the living Iguana, a plant-eating lizard.
5. A mass of small stones (gastric mill or gizzard) found in the stomach region of a skeleton of the African prosauropod Massospondylus may have been used to crush swallowed vegetation.
No review of prosauropods would be complete without mention of
Mussaurus (mouse lizard) of the Upper Triassic, described in the late 1970s from a tiny skeleton and associated eggs from the Upper Triassic of southern Argentina. The total body length of
Mussaurus was about 20 centimeters. Clearly, this tiny baby dinosaur, which could have stood in the palm of your hand, was a hatchling prosauropod. It may have been a juvenile of
Coloradisaurus, a prosauropod dinosaur from the same locality known only from adult bones. The discovery of
Mussaurus not only brought to light one of the world’s smallest dinosaurs, but also demonstrated that prosauropods, like sauropods, were egg layers. Indeed, the eggs of
Mussaurus are the oldest known dinosaur eggs.
Prosauropods were among the first dinosaurs to appear during the Late Triassic. They rapidly achieved a worldwide distribution and were among the largest, and most successful, plant-eating vertebrates of the Late Triassic and Early Jurassic. The great similarity of prosauropods such as Ammosaurus from the United States and Lufengosaurus from southern China suggests that by the Early Jurassic, environments favorable to prosauropods were present across the world’s continents.
The extinction of prosauropods at the end of the Early Jurassic marks the first significant disappearance of a major group of dinosaurs. It also coincided with the diversification of sauropod dinosaurs as well as other large plant-eaters, the stegosaurs and ankylosaurs. So it is tempting to believe that the prosauropods were replaced by these larger herbivores.
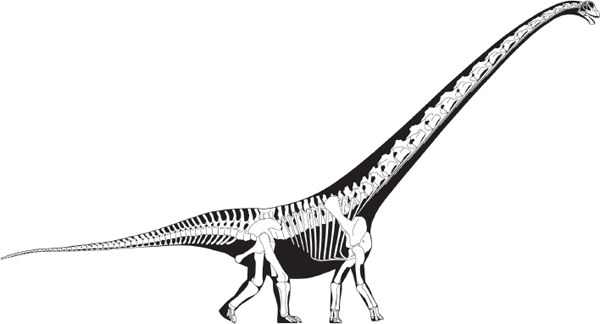
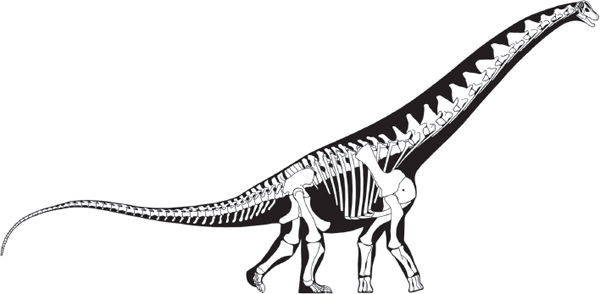
These gigantic quadrupedal herbivores include the largest land animals of all time, behemoths that weighed as much as 59 tons (possibly more) and attained body lengths of at least 35 meters. Brontosaurus, correctly called Apatosaurus, is the sauropod most familiar to the general public, followed by Diplodocus and Brachiosaurus. Paleontologists recognize as many as 75 genera of sauropods, although most of these animals are not known from complete skeletons. Evolutionary novelties of sauropods include their gigantic size, long necks and tails, tiny heads, and dorsally located nostrils, among other features.
The oldest known sauropods are of Late Triassic age, and the group persisted and had a worldwide distribution until the extinction of all the dinosaurs at the end of the Cretaceous (
figure 6.6). In terms of general size and diversity, the zenith of sauropod evolution occurred during the Jurassic–Cretaceous transition. At least three very distinct types of sauropods coexisted: the diplodocoids and primitive macronarians, and the Titanosauria (advanced macronarians), which are distinguished by features of their skulls, vertebrae, and limb skeletons (
box 6.1). Thus, one current and well-accepted phylogeny of sauropods recognizes an array of primitive eusauropods (including the very long-necked
Mamenchisaurus) and two groups of neosauropods: Diplodocoidea and Macronaria. Primitive macronarians include
Brachiosaurus, but most macronarians are titanosaurs, a mostly Cretaceous group known from fossils found on the former Gondwana continents.
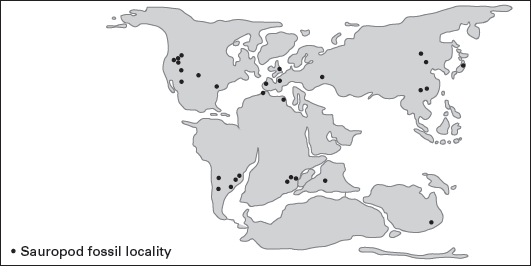
FIGURE 6.6
Sauropod dinosaurs were the largest and among the most successful plant-eating vertebrates of the Jurassic and Cretaceous. Their fossils are known from all continents except Antarctica. (Drawing by Network Graphics)
The skulls of sauropods were small and weakly connected to their vertebral columns. Because of this, they were easily detached and lost after death, so it is the norm to find sauropods without skulls (see
box 6.2). Indeed, the skulls of many sauropod taxa are unknown. This has forced paleontologists to rely on other aspects of the skeletons of sauropods, especially their vertebrae, to classify them. Fortunately, sauropod vertebrae were intricate structures that varied significantly from sauropod to sauropod. So they may offer a useful basis for the classification of these giants.
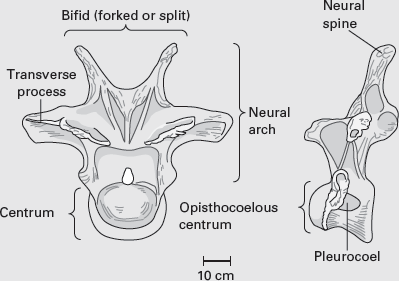
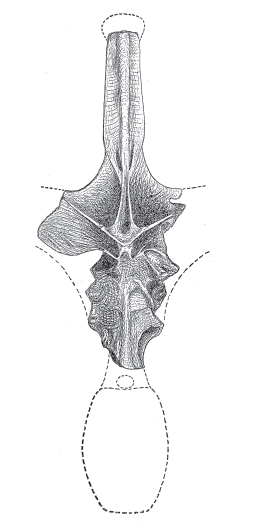
The basic sauropod vertebra, like that of most vertebrates, consists of a
centrum beneath a
neural arch (
box figure 6.1A). The centrum is the body of the vertebra and provides a site of attachment for muscles that support the body. The connected centra together comprised the flexible rod upon which the body of the dinosaur was hung. The centrum also supports the neural arch, a complex structure that encloses the spinal cord and sends out bony struts to which the ribs and other vertebrae are attached. The top of the neural arch is a flange of bone called the neural spine, another site of attachment for tendons, muscles, and ligaments, especially in the shoulder and hip regions.
BOX FIGURE 6.1A
Sauropod vertebrae, such as this dorsal vertebra of Apatosaurus, present a variety of features that can be used in sauropod classification. (Drawing by Network Graphics)
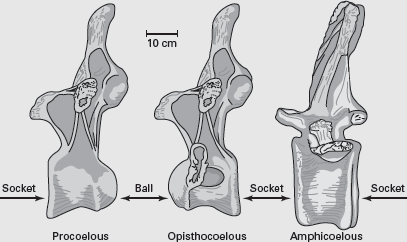
BOX FIGURE 6.1B
Sauropod vertebrae are procoelous, opistocoelous, or amphicoelous, depending on the arrangement of the ball-and-socket joints of the centra. (Drawing by Network Graphics)
As stated in the appendix, “A Primer of Dinosaur Anatomy,” the vertebral column of all dinosaurs, including sauropods (and most other vertebrates), can be divided into four regions: cervical (neck), dorsal (back), sacral (hip), and caudal (tail). In addition, the shapes of the surfaces at which the centra meet each other (articulate) have descriptive names. In sauropods, the centra meet at ball-and-socket joints (
box figure 6.1B). In a single centrum, if the socket is anterior and the ball is posterior, the vertebra is termed
procoelous, from the Greek words
pro, meaning “before,” and
koilus (
coelos), meaning “hollow” or “cavity.” If the situation is reversed—socket posterior and ball anterior—the vertebra is termed
opisthocoelous (
opisthen is Greek for “behind”). Vertebrae bounded by two cavities are called
amphicoelous (
amphi is Greek for “double”). Indeed, one sauropod genus,
Amphicoelias, takes its name from the amphicoelous nature of some of its vertebral centra.
The centra of many sauropod dinosaurs, particularly in the cervical region, were a lightly built framework of delicate struts and buttresses bounding deep cavities, especially along the sides of the centra (see
box figure 6.1A). Such cavities are called
pleurocoels (
pleura is Greek for “side”; thus, “side cavities”). Through evolution, the solid, spool-like vertebrae of sauropod ancestors were greatly modified by removing bone so that the vertebrae became much lighter. The development of pleurocoels may thus have helped lighten an otherwise inordinately heavy structure in a very large animal already fighting a tremendous battle against gravity during its day-to-day activities. Some paleontologists have also suggested that the pleurocoelous vertebrae may have been invested with air sacs that functioned in a fashion similar to the air sacs that modern birds have in their bones, which help to lighten them and increase the efficiency of their respiration. Another suggestion is that the pleurocoels were sites of glycogen (glucose, a natural sugar) storage. These ideas are difficult to evaluate from just the structure of the vertebrae alone, however. What is certain is that the vertebrae were much lightened by the presence of pleurocoels.
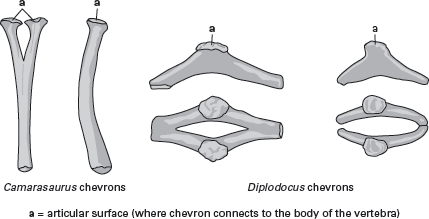
The caudal vertebrae of sauropods had chevron bones on their undersides that surrounded and protected blood vessels for the tail. The shape of these chevrons; the shape and length of the neural spines; and whether various vertebrae are procoelous, opisthocoelous, or amphicoelous are important features used to classify sauropods. Thus, for example, the titanosaurians are identified in part on the basis of their procoelous anterior caudal vertebrae, which differ from the caudals of other sauropods.
The most primitive sauropods are considered primitive
eusauropods. Among the best-known primitive eusauropods are the Chinese genera
Shunosaurus and
Datousaurus, described from the Zigong dinosaur quarry of the Middle Jurassic in Sichuan Province (see
box 10.2). These were relatively small sauropods;
Datousaurus, the longer of the two, was “only” 10 meters long. Their skulls were rather similar to that of
Camarasaurus but had longer muzzles. The numerous teeth were slender but had small, spoon-shaped crowns and were thus intermediate in structure between those of diplodocoids and those of
Camarasaurus. There were 12 cervical vertebrae and 13 in the back. The neck vertebrae were short and lacked much development of the pleurocoels. The neural spines were not divided, and there were three bones in the wrist and two in the ankle. The humerus-to-femur ratio was about 0.66, the same as in diplodocoids. The chevrons were forked, and in
Shunosaurus, there was even a club at the end of the tail.
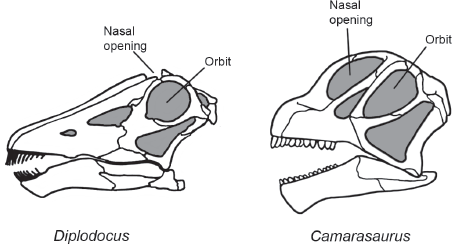
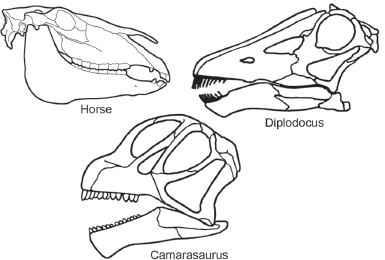
Diplodocoids were Late Jurassic sauropods known mostly from North America, Europe, and Africa. They are most easily distinguished by their skulls, which were long and slender with elongate muzzles (
figure 6.7). The jaws bore slender, peg-like teeth confined to the front of the mouth. The nostrils were on top of the skull, in front of and above the orbits.
Diplodocoid bodies were long and relatively lightly built, and diplodocoids were among the longest, though not necessarily the heaviest, dinosaurs (
figure 6.8). A 26-meter-long
Diplodocus would have weighed about 12 tons, considerably less than the shorter, 40-ton
Brachiosaurus. The necks of diplodocoids were extremely long, with an increased number of vertebrae at the expense of those in the back. This was foreshadowed by the neck of the primitive eusauropod
Mamenchisaurus, from the Cretaceous of China, which had 19 vertebrae but only 12 in the back (
figure 6.9).
FIGURE 6.7
Diplodocoids and primitive macronarians, such as Camarasaurus, can most easily be distinguished by features of their skulls. (© Scott Hartman)
FIGURE 6.8
An adult Diplodocus was a 27-meter-long, lightly built sauropod, characteristic of the diplodocoids. (© Scott Hartman)
The neck vertebrae of diplodocoids had unusually short ribs, and their neural spines were marked by a deep, V-shaped groove, as were those of the anterior dorsal vertebrae. At least one pulley-like muscle or tendon may have occupied this groove as part of the neck- and head-lifting musculature. The spines of the vertebrae in the hip region were high, and the chevrons of the tail were much modified from the type of chevron characteristic of more primitive sauropods. Instead of projecting downward as simple spines, the chevrons of diplodocoids were modified with fore and aft expansions into parallel rods (
figure 6.10). The tails of diplodocoids were very long and made up of at least 80 vertebrae. The last 30 to 40 of these vertebrae were little more than elongate rods that formed what has been called a “whiplash.”
The limbs of diplodocoids were relatively slender, and their rib cages were deep and narrow. The forelimbs were fairly short for sauropods, with a typical humerus-to-femur ratio of 0.66. The number of bones in the wrist and ankle were much reduced: only one or two in the wrist and one (astragalus) in the ankle. Apparently, both the wrist and the ankle were heavily invested with cartilage, which absorbed the stresses produced by so heavy a creature.
FIGURE 6.9
The development of an extremely long neck culminated in Mamenchisaurus, from the Upper Jurassic of China. (© Scott Hartman)
FIGURE 6.10
The normal chevrons of sauropod dinosaurs were shaped like “tuning forks” of bone directed downward. Those of diplodocids were modified to have fore-and-aft expansions that, in some posterior caudal vertebrae, became rod-like extensions. (Drawing by Network Graphics)
Diplodocus, of course, is the typical and best-known diplodocoid (see
figure 6.8). Its fossils are known from Upper Jurassic deposits in the western United States, as are those of the closely related diplodocoid,
Barosaurus.
Barosaurus has also been reported from eastern Africa. It had much longer neck vertebrae and a shorter tail than did
Diplodocus.
Perhaps the most famous diplodocoid is
Apatosaurus, also known as
Brontosaurus (
box 6.2).
Apatosaurus was one of the bulkiest diplodocoids, with robust limb bones, heavy cervical ribs, and a single bone in its wrist.
Brontosaurus stands out as one of the most popular names for a dinosaur. Yet few grade-school children shrink from pointing out that
Brontosaurus is not technically the correct name for this sauropod;
Apatosaurus is. This is because scientists have, since 1903, agreed that
Apatosaurus, named by Yale paleontologist Othniel Charles Marsh in 1877, and
Brontosaurus, also named by Marsh, but in 1879, represent the same kind of sauropod dinosaur. When more than one name exists for the same genus, the internationally accepted rules for naming animals, living and extinct, force paleontologists to use the earliest proposed name (see
box 2.1). The later name, in this case
Brontosaurus, is then branded a synonym and abandoned.
Marsh’s research also lies at the crux of another aspect of the
Brontosaurus business, the question of what type of head this sauropod had. In 1883, Marsh produced the first reconstruction of a sauropod dinosaur, that of
Brontosaurus excelsus (
box figure 6.2). In so doing, he relied primarily on a fairly complete skeleton from a dinosaur quarry at Como Bluff, Wyoming. But, this skeleton lacked a head. To complete his reconstruction Marsh had to guess which type of head, from among the sauropod heads known to him, belonged to
Brontosaurus. Unfortunately, he guessed incorrectly when he based the head in the reconstruction on two incomplete detached skulls, one from a dinosaur quarry east of Como Bluff and another from Garden Park, Colorado. So,
Brontosaurus was presented to the public with a
Camarasaurus-like skull, one with large, spatulate teeth.
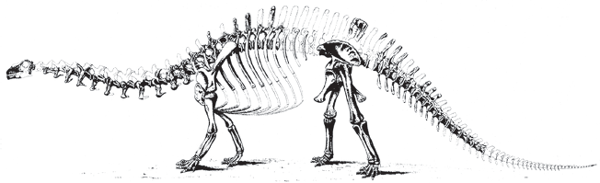
BOX FIGURE 6.2
Othniel Charles Marsh’s classic reconstruction of the skeleton of Apatosaurus included a Camarasaurus-like head. Only 75 years later was the correct, Diplodocus-like skull placed on the body of Apatosaurus. (Courtesy of Yale Peabody Museum of Natural History, New Haven, Connecticut)
More than a decade after Marsh’s death, between the years 1909 and 1915, the Carnegie Museum began to excavate the famous dinosaur quarry near Jensen, Utah, now known as Dinosaur National Monument. The most complete Apatosaurus known was one of the fruits of their labors, but it was also missing a head connected to the vertebral column. In 1916, this skeleton was named Apatosaurus louisae by W. J. Holland, the director of the Carnegie Museum, after Louise Carnegie, wife of the museum’s patron, Andrew Carnegie. The Apatosaurus louisae skeleton was found lying on top of another smaller, headless Apatosaurus skeleton. Nearby lay part of the skeleton of a third sauropod thought to be a Diplodocus. Also among those bodies was a Diplodocus-like skull. Holland thought this skull was too large to belong to the supposed Diplodocus skeleton or to the smaller Apatosaurus. In his mind, it made sense to attach it to the skeleton of Apatosaurus louisae. Thus, Holland reasoned, Marsh had guessed wrong.
The skeleton of Apatosaurus louisae was mounted at the Carnegie Museum, and Holland planned to place the Diplodocus-like skull on it. But, Henry Fairfield Osborn, one of the most eminent vertebrate paleontologists of the time, believed that Marsh was right and “convinced” Holland not to do so. Or did he? Holland left the Apatosaurus louisae skeleton on display without a head for more than 20 years. Only after Holland’s death in 1932 was a head placed on the skeleton—a copy of the Camarasaurus-like skull originally chosen by Marsh!
Holland, as the saying goes, must have rolled over in his grave. But, in the late 1970s he was vindicated. Further preparation of the skeletons from the Utah quarry showed that the skeleton Holland thought was a Diplodocus was actually another Apatosaurus. Three Apatosaurus skeletons in the same quarry as a Diplodocus-like skull provides very strong evidence that the skull belonged to Apatosaurus, too. Indeed, modern studies of sauropods have listed numerous similarities shared by the skeletons of Diplodocus and Apatosaurus, making it no surprise that their skulls would also be similar. So, the Camarasaurus-like skulls that rode the necks of Apatosaurus skeletons for nearly 100 years came off and were replaced by Diplodocus-like skulls with rod-shaped teeth.

FIGURE 6.11
Camarasaurus, one of the best known primitive macronarians, would have reached a length of as much as 18 meters. (© Scott Hartman)
Camarasaurus is the best known of all sauropods, known from dozens of skulls and many skeletons. It is a characteristic primitive macronarian and thus provides a remarkable contrast to the diplodocoids (
figure 6.11). This is most obvious from its skull (see
figure 6.7). That of
Camarasaurus was short and heavy with a blunt snout. The jaws bore relatively large, spatulate teeth along their entire length. The nostrils were large (hence the name
Macronaria, which means “large nose”) and located on the sides of the skull, just in front of the eyes.
The body of Camarasaurus was solidly built, but neither exceptionally long nor overly heavy. There were only 12 neck vertebrae (the number found in primitive sauropods), and a U-shaped trough in the neural spines extended along the first four dorsal vertebrae. The spines of the sacral vertebrae were lower and thicker than in diplodocoids. In the neck and back vertebrae, the pleurocoels were very extensive. The tail chevrons were of the standard sauropod type.
Camarasaurus had limbs that were stout, and the typical humerus-to-femur ratio was 0.70 or more. So, the forelimbs were relatively longer than in diplodocoids. The wrist and ankle each had two bones, and the forefoot was elongated. Besides
Camarasaurus, the best-known primitive macronarians are its compatriot
Haplocanthosaurus, from the Upper Jurassic of the western United States, and
Brachiosaurus, a behemoth best known from the Upper Jurassic of Tanzania (
figure 6.12).
In features of the skull and teeth,
Brachiosaurus closely resembled
Camarasaurus. But, in
Brachiosaurus, the forelimb was relatively long—the humerus-to-femur ratio was greater than 1.0. Thus, the shoulders of
Brachiosaurus were positioned higher than its hips. Although there were only 13 neck vertebrae and 11 or 12 vertebrae in the back, those of the neck were quite elongate, resulting in a very long neck. The neural spines were all undivided, and those in the sacral region were low. Although there were about 50 vertebrae in the tail, each vertebra was short, which made for a relatively short tail. The chevrons were simple, and the hand bore a very short claw on its first digit.
Brachiosaurus, from the Upper Jurassic of the western United States, Tanzania, and Portugal, was not even as long as
Apatosaurus, but it was at least twice as heavy and could raise its head 12 meters off the ground.
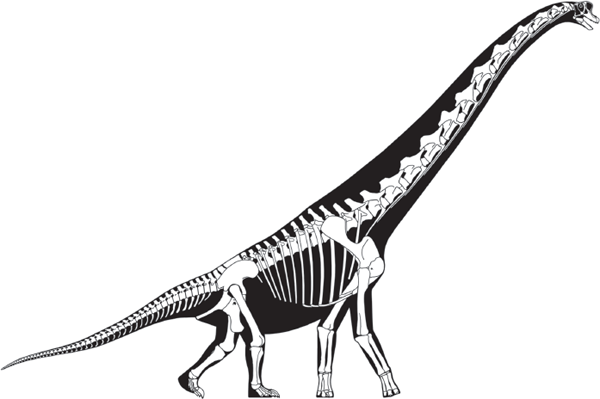
FIGURE 6.12
Brachiosaurus (called Giraffatitan by some paleontologists) was one of the heaviest land animals of all time, weighing as much as 50 tons and measuring 22 meters from snout to tail tip. (© Scott Hartman)
Titanosauria encompasses about 50 species of derived macronarians. These sauropods are identified by their procoelous anterior tail vertebrae, widened sacrum, short and robust limbs, and short forefeet, among other features (
figure 6.13). Furthermore, the shape of a titanosaur’s humerus and femur indicate that these bones were bowed out (not held strictly vertical) while walking.
FIGURE 6.13
This composite skeleton of Alamosaurus, from the Upper Cretaceous of North America, is of a relatively well-known titanosaur. (© Scott Hartman)
One titanosaur,
Saltasaurus, from the Cretaceous of Argentina, had body armor (
figure 6.14). So it is reasonable to assume that at least some other titanosaurids were armored. Most titanosaurs were of Cretaceous age and inhabited Argentina (
Antarctosaurus and
Saltasaurus), India (
Titanosaurus), and other parts of the Mesozoic southern supercontinent called Gondwana. A rare exception is
Opisthocoelicaudia, a titanosaur from the Upper Cretaceous of Mongolia.
In North America, the last known sauropod of the Late Cretaceous is
Alamosaurus. This fairly large titanosaur is known from just a few teeth (of the diplodocoid type), a shoulder blade and forelimb, some pelvic bones, a femur, a foot, some cervical and dorsal vertebrae, and a tail, yet it is one of the better known titanosaurs. It almost certainly emigrated from South America into North America at about the end of the Cretaceous (
box 6.3).
FIGURE 6.14
Armor plates like this one adorned the back of the titanosaurian Saltasaurus and may have characterized all titanosaurians. (Drawing by Network Graphics)
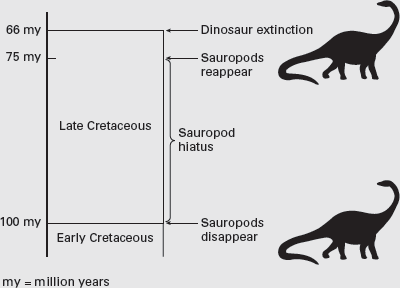
It is a curious fact that sauropod dinosaurs have a temporally disjunct distribution in the western United States during the Cretaceous. In Lower Cretaceous strata, sauropod bones and footprints are found in Wyoming, Texas, Oklahoma, and Arkansas. Thereafter, sauropod fossils are absent for about 30 million years, reappearing solely in the form of the titanosaurid Alamosaurus, in the uppermost Cretaceous deposits of Texas, New Mexico, and Utah. Two explanations have been offered to explain this 30-million-year absence, the sauropod hiatus.
The first explanation is that sauropods were not absent in western North America during the hiatus but were actually there all along. The environments the sauropods lived in during the hiatus have either not been preserved in the rock record or have not yet been sampled by paleontologists. So, sauropods may have lived in the western United States during the hiatus, but their remains either were not fossilized or are still to be discovered.
The second explanation posits the extinction of sauropods in western North America at about the end of the Early Cretaceous, about 100 million years ago. Thirty million years later, Alamosaurus arrived from elsewhere, probably from South America, where sauropods prospered throughout the Cretaceous.
The balance of evidence favors the second explanation. Particularly important is the fact that there are some very well-known dinosaur faunas in the western United States during the 30-million-year-long hiatus, representing a variety of environments from which no trace of a sauropod fossil is known. Also significant is the fact that the last North American sauropod,
Alamosaurus, is a titanosaurian closely related to South American Cretaceous sauropods. Although it does seem reasonable to believe that in the western United States, sauropods became extinct at the end of the Early Cretaceous and invaded 30 million years later (
box figure 6.3), the reasons for their extinction and subsequent invasion remain unknown.
BOX FIGURE 6.3
The sauropod hiatus lasted for 30 million years. (Drawing by Network Graphics)
Recent discoveries of huge sauropods have increased the known limits of body size in terrestrial vertebrates and raised the question, which was the largest sauropod? (
box 6.4). When using the word “large,” however, it is important to distinguish between weight (body mass) and length. As the previous discussion indicates, some of the longest sauropods were relatively lightly built and thus generally lighter than the shorter, more robust sauropods such as
Brachiosaurus.
Some recent candidates for the award for largest, in terms of weight, are Antarctosaurus, a titanosaur from the Cretaceous of Argentina estimated to have weighed up to 69 tons; Argentinosaurus, also from the Argentine Cretaceous, estimated at up to 73 tons; Brachiosaurus, weighing in at up to 78 tons; Paralititan from the Upper Cretaceous of Egypt and Dreadnoughtus from the Upper Cretaceous of Argentina, both weighing up to 59 tons; and Sauroposeidon, from the Lower Cretaceous of Oklahoma, weighing as much as 60 tons. For perspective, a Boeing 737 airplane weighs about 50 tons, and an African elephant weighs about 5 tons.
Other than
Brachiosaurus, none of these huge sauropods are known from a complete skeleton, so their weight estimates are based on various extrapolations. Yet, even based on complete skeletal material, published weight estimates of
Brachiosaurus range from 16 to more than 78 tons, a nearly fivefold difference. This shows how problematic dinosaur weight estimates can be. Different methods can yield very different estimates. About 50 tons seems to be the most reliable upper limit for known humongous sauropods (see
box 6.4).
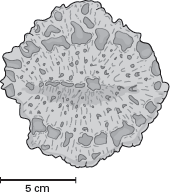
In 1878, Edward Drinker Cope described
Amphicoelias fragillimus, long considered to be the largest sauropod ever. He based the new dinosaur species on parts of a huge vertebra and femur from the Upper Jurassic Morrison Formation in Colorado. The vertebra was only part of the neural arch (
box figure 6.4), but Cope reported it to be a whopping 1.5 meters tall, leading him to estimate a total height of more than 1.8 meters for the dorsal vertebra that it represented.
In the 1890s, Cope sold his fossil collection to the American Museum of Natural History, including the bones of Amphicoelias fragillimus. However, these bones disappeared by 1900, perhaps discarded because they were very fragile and broken. Fortunately, the lost dinosaur has remained part of the scientific literature. Estimates of its size based on the data and illustration in Cope’s original article are of a 58- to 60-meter-long monster weighing as much as 150 tons. For comparison, most Late Jurassic sauropods weighed an estimated 7 to 26 tons. And, a typical living blue whale, generally considered the largest of all animals, is about 24 meters long and weighs in at about 190 tons.
Most paleontologists have accepted the colossal size of
Amphicoelias fragillimus, even though this makes it a sauropod three to four times larger than any contemporary Late Jurassic sauropod. Furthermore, the size estimates of
Amphicoelias fragillimus are of a sauropod much heavier than the 75 tons considered to be the maximum weight limit of any terrestrial organism, based on calculations using biomechanical and gravitational constraints.
These considerations encouraged a recent re-evaluation of Amphicoelias fragillimus by paleontologists Cary Woodruff and John Foster. They suggest that Cope made a typographical error, reporting the preserved portion of the vertebra as 1.5 meters tall, when it was more likely 1.05 meters tall. That still makes it the largest Late Jurassic sauropod, but one at least one-third smaller than had been estimated.
Of course, without the fossil and without Edward Drinker Cope (who died in 1897), we will never know whether the measurement Cope published was correct or a typo. What we do know is that no sauropod that approaches the size of Amphicoelias fragillimus has been discovered since 1878 and that the size estimates based on Cope’s publication seem implausible.
BOX FIGURE 6.4
Edward Drinker Cope’s original drawing of part of a vertebra of Amphicoelias fragillimus. (From E. D. Cope. 1878. A new species of Amphiocoelias. American Naturalist 12:563–564)

FIGURE 6.15
At 50 or more tons, Brachiosaurus (or Giraffatitan, left), Puertasaurus (center), and Argentinosaurus (right) were the heaviest dinosaurs and would have dwarfed a human and a living elephant but still were significantly lighter than a blue whale. (© Scott Hartman)
Length estimates based on incomplete skeletons are also tricky. The longest estimates that may be reliable are 35 meters for Mamenchisaurus, 36 meters for Argentinosaurus, and 39 meters for Turiasaurus, from the Upper Jurassic of Spain. Length estimates as long as 58 meters were made for “Seismosaurus,” a synonym of Diplodocus, but did not stand up to critical scrutiny.
At present, conservative estimates of the upper limits of known sauropod size are about 50 tons for an extremely heavy
Brachiosaurus and about 40 meters long for a very elongate
Turiasaurus (
figure 6.15).
The enormous size and the unusual body shape (especially the long neck and minuscule head) of sauropod dinosaurs have led to much speculation about their way of life. Particularly significant topics for discussion are sauropod diet, metabolism, locomotion, habitat preferences, and reproduction, as well as evidence of social behavior among sauropods.
Diet
The two primary types of sauropod teeth, well represented by the peg-like teeth of diplodocoids and the spatulate teeth of
Camarasaurus, appear to have been well suited to cropping vegetation in much the same way that a living horse uses its incisors (
figure 6.16). Some sauropod teeth are found worn, but sauropods clearly had no grinding or shearing teeth for smashing and pulping vegetation as did the duck-billed dinosaurs, with their dental batteries, or living horses, with their premolars and molars. Sauropods must have swallowed vegetation with little chewing and relied on chemical or bacterial mechanisms in the gut to break it down.
FIGURE 6.16
The teeth of sauropods probably functioned like the incisors of living horses and were used to crop vegetation. (© Scott Hartman)
Some paleontologists have argued that the rod-like teeth of the diplodocids were used as a sieve to collect algae and invertebrates. This is plausible, though it would necessitate an incredible supply of algae and invertebrates for the diplodocids to feed on. Sauropod cropping of different kinds of plants, quickly swallowed and broken down in the gut chemically or bacterially, seems most plausible. Assuming modern nutritional values for the vegetation of the Mesozoic, it has been estimated that a 29-ton sauropod would have had to eat 50 kilograms of vegetation each day!
Metabolism
The 50-kilogram-per-day calculation is based on the notion that sauropods had metabolic rates comparable to those of living reptiles. Of course, if they had higher metabolic rates, comparable to those of living mammals, they would have had to eat more. There is no conclusive evidence, however, that this was so (see
chapter 14), so it is reasonable to conclude that they had an ectothermic (“cold-blooded”) reptilian metabolism, with one important difference.
This difference is evident considering the great bulk of sauropods; their massiveness would have made them what physiologists term inertial homeotherms (also called gigantotherms). In other words, sauropods would have maintained an essentially constant body temperature (homeothermy) because of the resistance to heat transference inherent in their large mass (thermal inertia). Thus, by virtue of its size alone, once it reached its ideal body temperature, a sauropod would have retained that temperature much longer than a smaller animal. Of course it would also have taken much longer to reach that temperature in the first place!
FIGURE 6.17
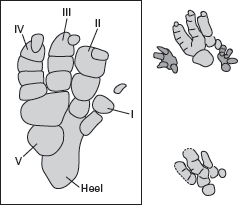
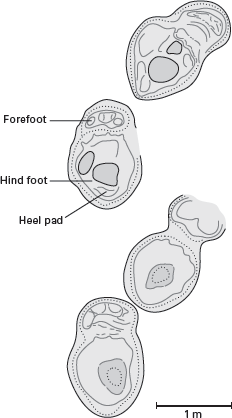
Sauropod footprints like these from Morocco indicate that sauropods walked on all fours and that their heels were supported by thick, fleshy pads, as are the heels of living elephants. (Drawing by Network Graphics)
Locomotion and Habitat Preferences
The nearly equal length of the fore and hind limbs; the thick, solid limb bones; the massive pelvic and shoulder girdles; and the short, broad feet of sauropods indicate that they were obligatory quadrupeds. Sauropod foot structure indicates that they walked on the tips of their front toes with their hind-foot heels resting on large pads, similar to living elephants. Sauropod footprints confirm these conclusions (
figure 6.17). The limb structure of sauropods is what anatomists term
graviportal, meaning designed to bear great weight on land.
The oldest idea about sauropod lifestyle is that they were aquatic. For nearly a century, paleontologists believed that sauropod limbs were not strong enough to support their bulky bodies, that the “weak” teeth of sauropods could be used only to eat soft, aquatic vegetation, and that the nostrils on the top of some sauropod skulls functioned as snorkels, allowing the animal to breathe while most of the head was submerged. The image of a sauropod dinosaur standing in deep water, its head emergent like the periscope of a submarine, is classic (
figure 6.18). Indeed, one paleontologist even went so far as to conclude that the differing neck lengths and limb proportions indicated habitation of different depths of water. According to this scheme,
Brachiosaurus lived in water depths of about 8 meters,
Diplodocus in depths of 4 to 5 meters, and
Apatosaurus in shallow depths of 2 to 3 meters.
A modern view of sauropod habitat preferences has resulted from a reappraisal of sauropod anatomy. This reappraisal demonstrates that there is little, if any, evidence to support the idea of aquatic sauropods. Indeed, it has been estimated that water pressure would have prevented expansion and contraction of a sauropod’s lungs if the dinosaur were submerged. The massive limbs of sauropods could certainly have supported their huge bodies; the graviportal limb skeletons of sauropods seem to have been designed to do just that. The teeth of sauropods were as robust as a horse’s incisors and could easily have cropped tough vegetation. And, the nasal openings on top of some sauropod heads were remarkably like those of living tapirs, land-living mammals that have a short trunk (proboscis) like a miniature version of an elephant’s trunk. The nostrils far back on a tapir’s skull are an outgrowth of the presence of a proboscis. Perhaps some sauropods, most likely the diplodocoids, had a tapir-like proboscis. The evidence thus seems to support a view of sauropods as land-living treetop browsers analogous to living giraffes and elephants.
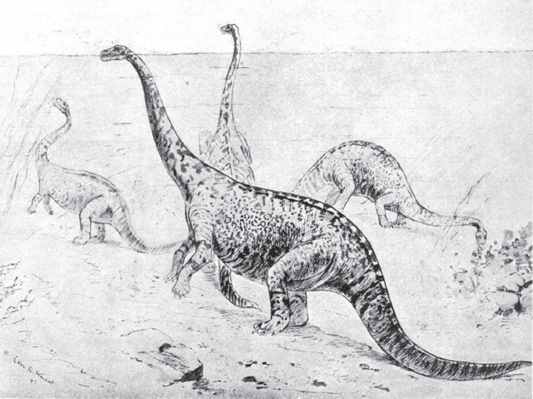
FIGURE 6.18
Classic reconstructions of sauropods (drawn by paleontologist Edward Drinker Cope and published in 1897) show them submerged in water. Paleontologists now envision sauropods as tree-top browsers that spent most of their lives on dry land.
Reproduction
Like prosauropods, sauropod dinosaurs were egg layers. Sauropod eggs are well known from the Jurassic of the United States and Portugal and from the Cretaceous of the United States, Argentina, France, Spain, Romania, and India. Titanosaur embryos in eggs are known from the Upper Cretaceous of Argentina. At Aix-en-Provence in France, numerous eggs as much as 25 centimeters long, with bumpy shell textures, have been attributed to the titanosaur Hypselosaurus.
Although discussed at greater length in
chapter 13, it is appropriate to mention the circumstantial evidence for social behavior in sauropods. Many of the great dinosaur quarries of the Late Jurassic, such as those at Como Bluff, Wyoming, are populated mostly by sauropod skeletons. These mass death assemblages of sauropods may indicate group (gregarious) behavior among these giant reptiles, but some of the dinosaur quarries, such as the one at Dinosaur National Monument, Utah, are populated mostly by river-transported carcasses that accumulated in sand banks. The evidence of group behavior among sauropods provided by the mass death assemblages is thus not totally convincing.
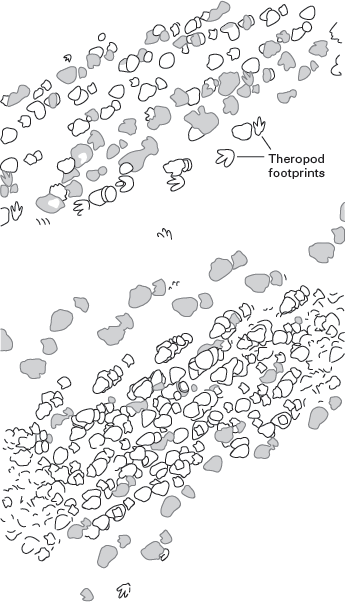
Sauropod trackways also support, to some extent, the notion of gregarious behavior among sauropods. For example, Early Cretaceous footprints from near Glen Rose, Texas (
figure 6.19), seem to document a group of sauropods walking together in the same direction. But, to call this group a herd, which implies a complex social structure, may be overly generous. Not only is there no unequivocal evidence that all these sauropods walked together at one time, but the distribution of track sizes, which would indicate the presence and number of sauropods of different ages, is scattered, which suggests that no orderly movement of young and old was involved.
FIGURE 6.19
These sauropod trackways, from the Lower Cretaceous of Texas, are taken by some paleontologists as evidence of group behavior among sauropods. (Drawing by Network Graphics)
The evidence for sauropod group behavior is at best circumstantial. Mass death assemblages and multiple trackways suggest some sort of group behavior among sauropods, but little else can be concluded at present.
Sauropod dinosaurs evolved from a prosauropod-like ancestry late in the Triassic. Although no known prosauropod is a strong candidate for sauropod ancestry (unless evolutionary reversals be accepted), Riojasaurus, from the Late Triassic of Argentina, or Vulcanodon, from the Early Jurassic of southern Africa, may best approximate that ancestor.
The oldest sauropods are of Late Triassic and Early Jurassic age and include Barapasaurus from India, Isanosaurus from Thailand, and Vulcanodon from southern Africa. They and Middle Jurassic sauropods, such as Datousaurus and Shunosaurus (China), Cetiosaurus (England), and Patagosaurus (Argentina), were long included in a group of primitive sauropods nominally termed cetiosaurids. This “family” is clearly a grade level (not a clade) in the evolution of sauropods characterized by relatively small (up to 12 meters long), short-necked forms with Camarasaurus-like heads and teeth. It is interesting that these large animals appear suddenly in the fossil record with most of the graviportal limb specializations we associate with sauropods. Their geographic distribution suggests a likely origin of sauropods in the southern supercontinent of Gondwana.
The zenith in size and diversity of sauropods was achieved during the Jurassic–Cretaceous transition. The terrestrial giants had a worldwide distribution by this time, and Diplodocus, Brachiosaurus, and Camarasaurus well represent their structural diversity. In terms of size, diversity, and sheer abundance, sauropod dinosaurs were the dominant herbivores on land during the Jurassic–Cretaceous transition.
Sauropod fortunes seem to have waned during the Cretaceous, when there were fewer and mostly smaller animals. It is tempting to suggest that the rise of ornithopod dinosaurs, potential herbivorous competitors of sauropods, and the appearance and evolution of flowering plants during the Cretaceous, may have contributed to or resulted from the decline in the sauropods. Sauropods were plant eaters, but they may not have been well adapted to eating flowering plants. Particularly significant is that most sauropod diversity during the Cretaceous was limited to the southern continents where ornithopods were not very successful. The extinction of sauropods coincided with the extinction of the dinosaurs at the end of the Cretaceous. Though reduced in diversity and numbers, sauropods can truly be said to have met the end with “their heads held high.”
1. Sauropodomorph dinosaurs comprise two closely related groups, prosauropods and sauropods, distinguished from other dinosaurs by having had small heads, spatulate teeth, long necks, short feet, and large claws on the first digits of their forefeet.
2. No known prosauropod is a suitable ancestor of sauropods, although Riojasaurus, from the Upper Triassic of Argentina, may best approximate that ancestry.
3. Prosauropods lived during the Late Triassic and Early Jurassic and so are some of the oldest dinosaurs, representing the first evolutionary diversification of plant-eating dinosaurs.
4. Sauropod dinosaurs first appeared in the Late Triassic, and by the Late Jurassic they were the dominant very large plant-eating dinosaurs.
5. At least four types of sauropods can be recognized in the Jurassic–Cretaceous: primitive eusauropods, diplodocoids, primitive macronarians, and titanosaurs.
6. The heaviest known sauropod was Brachiosaurus, weighing up to 50 tons, and the longest sauropod was Turiasaurus, with a length of 39 meters.
7. Sauropods were plant eaters, egg layers, and inertial homeotherms. There is no evidence to support the long popular notion of aquatic sauropods. Sauropods had elephant-like limb structures and probably spent most of their time on dry land.
8. By the Jurassic–Cretaceous transition, sauropods had reached their evolutionary zenith in terms of size and diversity.
9. Sauropods were much less successful after the Jurassic but did survive until the end of the Cretaceous.
Alamosaurus
Amphicoelias
amphicoelous
Apatosaurus
Brachiosaurus
Brontosaurus
Camarasaurus
centrum
chevron
diplodocoid
Diplodocus
eusauropod
graviportal
Hypselosaurus
Macronaria
Mamenchisaurus
Mussaurus
neural arch
opisthocoelous
Plateosaurus
pleurocoel
procoelous
prosauropod
Riojasaurus
sauropod
sauropod hiatus
Sauropodomorpha
Seismosaurus
Titanosauria
Vulcanodon
1. Why do most paleontologists exclude any known prosauropod from the ancestry of sauropods?
2. What characteristic features of prosauropods are exemplified by Plateosaurus?
3. What anatomical evidence supports the notion that prosauropods and sauropods were herbivores?
4. Distinguish diplodocoid from macronarian sauropods.
5. What other groups of sauropods are recognized by paleontologists, and what are their salient characteristics?
6. Describe the vertebrae of sauropods, and explain how they are relevant to sauropod classification.
7. How was the confusion about the name and the skull of Brontosaurus resolved?
8. Why did paleontologists believe sauropods were aquatic, and why do they no longer hold this view?
9. Review the evolutionary history of the sauropodomorph dinosaurs.
Berman, D. S., and J. S. McIntosh. 1978. Skull and relationships of the Upper Jurassic sauropod Apatosaurus (Reptilia, Saurischia). Bulletin of the Carnegie Museum of Natural History 8:1–35. (An in-depth technical description of the skull of Apatosaurus and related sauropods as well as an authoritative review of the “Brontosaurus business”)
Coombs, W. P., Jr. 1975. Sauropod habits and habitats. Palaeogeography, Palaeoclimatology, Paleoecology 17:1–33. (A thorough review of the evidence for aquatic habits among sauropods and the key article debunking this notion)
Curry Rogers, K. A., and J. A. Wilson. 2005. The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press. (A collection of 11 articles on diverse sauropod research)
Desmond, A. J. 1976. The Hot-Blooded Dinosaurs. New York: Dial. (Chapter 5, pp. 100–133, recounts the history of ideas on sauropod behavior, habitat preferences, and the rise and demise of the idea of aquatic sauropod)
Fowler, D. W., and R. M. Sullivan. 2011. The first giant titanosaurian sauropod from the Upper Cretaceous of North America. Acta Palaeontologica Polonica 56:685–690. (Giant bones of Alamosaurus suggest a sauropod as large as Argentinosaurus)
Klein, N., K. Remes, C. T. Gee, and P. M. Sander, eds. 2011. Biology of the Sauropod Dinosaurs: Understanding the Life of Giants. Bloomington: Indiana University Press. (A collection of 18 articles on diverse aspects of sauropod biology)
Lavocara, K. J., et al. 2014. A gigantic, exceptionally complete titanosaurian sauropod dinosaur from southern Patagonia, Argentina. Nature Scientific Reports 4:6196. (Description of a new behemoth, Dreadnoughtus)
Lucas, S. G., and A. P. Hunt. 1989. Alamosaurus and the sauropod hiatus in the Cretaceous of the North American Western Interior. Geological Society of America Special Papers 238:75–85. (Develops and evaluates explanations for the sauropod hiatus)
Ostrom, J. H., and J. S. McIntosh. 1966. Marsh’s Dinosaurs: The Collections from Como Bluff. New Haven, Conn.: Yale University Press. (A thorough review of the collections from one of the world’s great Jurassic–Cretaceous dinosaur quarries, including publication of 65 lithographs prepared for Othniel Charles Marsh of Diplodocus, Apatosaurus, and Camarasaurus, among others)
Rainforth, E. C. 2003. Revision and re-evaluation of the Early Jurassic dinosaurian ichnogenus Otozoum. Palaeontology 46:803–808. (Study of early prosauropod footprints)
Sander, P. M., et al. 2011. Biology of the sauropod dinosaurs: The evolution of gigantism. Biological Reviews 86:117–155. (Detailed review of the biology of sauropod gigantism)
Wilson, J. A., and K. A. Curry Rogers. 2012. Sauropoda, pp. 445–481, in M. K. Brett-Surman, T. R. Holtz, Jr., and J. O. Farlow, eds.,
The Complete Dinosaur. 2nd ed. Bloomington: Indiana University Press. (Concise review of the sauropods)
Woodruff, D. C., and J. R. Foster. 2014. The fragile legacy of Amphicoelias fragillimus (Dinosauria: Sauropoda; Morrison Formation, latest Jurassic). Volumina Jurassica 12:211–220. (Argues that the huge size estimates for Amphicoelias fragillimus may have been based on a typographical error)
Yates, A. M. 2012. Basal sauropodomorpha, pp. 425–433, in M. K. Brett-Surman, T. R. Holtz, Jr., and J. O. Farlow, eds., The Complete Dinosaur. 2nd ed. Bloomington: Indiana University Press. (Concise review of the prosauropods)
Where can you find a giant sauropod dinosaur? Perhaps the most spectacular sauropod fossil in public view is the skeleton of Brachiosaurus (some call it Giraffatitan) on display in the Museum für Naturkunde (Berlin, Germany). Collected in the early twentieth century from Upper Jurassic rocks in eastern Africa, this monster towers almost 13 meters above the museum floor and is advertised as the tallest dinosaur skeleton on display anywhere. Closer to home, the American Museum of Natural History (New York) exhibits skeletons of Apatosaurus and Barosaurus. In Washington, D.C., the Smithsonian’s National Museum of Natural History has Camarasaurus and Diplodocus skeletons on display. The Carnegie Museum of Natural History (Pittsburgh, Pennsylvania) has the renowned original skeleton of Diplodocus carnegii, named after the museum’s benefactor. And, don’t forget the infamous skeleton of “Brontosaurus” (with the correct head) that is the centerpiece of the Great Hall of Dinosaurs at the Yale Peabody Museum of Natural History (New Haven, Connecticut). In California, the Natural History Museum of Los Angeles County has the long-necked Chinese sauropod Mamenchisaurus in its exhibit hall. And, way down south, in Buenos Aires, Argentina, you can see a mounted skeleton of Argentinosaurus (not all of it real), touted by some as the largest of all sauropods, at the Museo Argentino de Ciencias Naturales Bernadino Rivadavia. Closer to home, you can see a replica of the Argentinosaurus skeleton on display at the Fernbank Museum of Natural History (Atlanta, Georgia).